Comparative Transcriptome Analysis Revealing the Different Germination Process in Aryloxyphenoxypropionate-Resistant and APP-Susceptible Asia Minor Bluegrass (Polypogon fugax)


Abstract
1. Introduction
2. Results
2.1. Characterization of Seed Germination in LS and LR
2.2. De Novo Assembly of P. fugax Reference Transcriptome
2.3. Identification of Differentially Expressed Genes (DEGs)
2.4. GO (Gene Ontology) and KEGG (Kyoto Encyclopedia of Genes and Genomes) Enrichment of DEGs
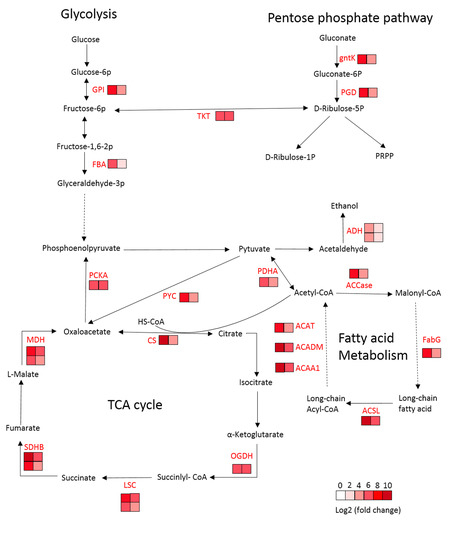
2.5. DEGs Related to Fatty Acid Metabolism
2.6. DEGs Related to Carbohydrate Metabolism
2.7. DEGs Related to Hormones Biosynthesis and Signal Transduction
2.8. Hormones Regulate the Seed Germination of LS and LR
2.9. Validation of DEGs by Quantitative Real-Time PCR (qRT-PCR)
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Growth Condition
4.2. Dynamic Changes in Germination Percentage, Soluble Sugar and Protein Content
4.3. Effect of Exogenous Hormones on Seed Germination of LS and LR
4.4. RNA Isolation and Transcriptome Sequencing
4.5. Transcriptome Assembly and Functional Classification
4.6. Differential Gene Expression and Enrichment Analysis
4.7. qRT-PCR Validation
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Casale, F.A.; Giacomini, D.A.; Tranel, P.J. Empirical investigation of mutation rate for herbicide resistance. Weed Sci. 2019, 67, 361–368. [Google Scholar] [CrossRef]
- Busi, R.; Gaines, T.A.; Walsh, M.J.; Powles, S.B. Understanding the potential for resistance evolution to the new herbicide pyroxasulfone: Field selection at high doses versus recurrent selection at low doses. Weed Res. 2012, 52, 489–499. [Google Scholar] [CrossRef]
- Neve, P.; Powles, S. High survival frequencies at low herbicide use rates in populations of Lolium rigidum result in rapid evolution of herbicide resistance. Heredity 2005, 95, 485–492. [Google Scholar] [CrossRef]
- Maxwell, B.D.; Mortimer, A.M. Selection for Herbicide Resistance. In Herbicide Resistance in Plants: Biology and Biochemistry; Powles, S.B., Holtum, J.A.M., Eds.; Lewis: Boca Raton, FL, USA, 1994; pp. 1–25. [Google Scholar]
- Délye, C.; Michel, S. Universal primers for PCR-sequencing of grass chloroplastic acetyl-CoA carboxylase domains involved in resistance to herbicides. Weed Res. 2005, 45, 323–330. [Google Scholar] [CrossRef]
- Yu, Q.; Cairns, A.; Powles, S. Glyphosate, paraquat and ACCase multiple herbicide resistance evolved in a Lolium rigidum biotype. Planta 2007, 225, 499–513. [Google Scholar] [CrossRef]
- Powles, S.B.; Yu, Q. Evolution in action: Plants resistant to herbicides. Annu. Rev. Plant Biol. 2010, 61, 317–347. [Google Scholar] [CrossRef] [PubMed]
- Cousens, R.D.; Fournier-Level, A. Herbicide resistance costs: What are we actually measuring and why? Pest Manag. Sci. 2018, 741, 539–1546. [Google Scholar] [CrossRef]
- Vila-Aiub, M.M.; Yu, Q.; Powles, S.B. Do plants pay a fitness cost to be resistant to glyphosate? New Phytol. 2019, 223, 532–547. [Google Scholar] [CrossRef]
- Anthimidou, E.; Ntoanidou, S.; Madesis, P.; Eleftherohorinos, I. Mechanisms of Lolium rigidum multiple resistance to ALS- and ACCase-inhibiting herbicides and their impact on plant fitness. Pestic. Biochem. Physiol. 2019, 164, 65–72. [Google Scholar] [CrossRef]
- Menchari, Y.; Chauvel, B.; Darmency, H.; Délye, C. Fitness Costs Associated with Three Mutant Acetylco enzyme a Carboxylase Alleles Endowing Herbicide Resistance in Black-Grass Alopecurus myosuroides. J. Appl. Ecol. 2008, 45, 939–947. [Google Scholar] [CrossRef]
- Shergill, L.S.; Boutsalis, P.; Preston, C.; Gill, G.S. Fitness costs associated with 1781 and 2041 ACCase-mutant alleles conferring resistance to herbicides in Hordeum glaucum Steud. Crop Prot. 2016, 87, 60–67. [Google Scholar] [CrossRef]
- Li, Y.H. Weed Flora of China, 1st ed.; China Agriculture Press: Beijing, China, 1998; pp. 1313–1315. (In Chinese) [Google Scholar]
- Xu, Z.H.; Qi, H.Y.; Lu, Y.L.; Yang, W.D.; Xie, G.X. Weed Identification and Management, 1st ed.; Zhejiang University: Hangzhou, China, 2014; p. 372. (In Chinese) [Google Scholar]
- Wang, K.; Qiang, Q. Quantitative analysis of distribution of weed communities in wheat fields in the south of Jiangsu province. J. Biomath. 2005, 20, 107–114. (In Chinese) [Google Scholar]
- Tang, W.; Zhou, F.; Chen, J.; Zhou, X. Resistance to ACCase-inhibiting Herbicides in an Asia Minor Bluegrass (Polypogon fugax) Population in China. Pestic. Biochem. Physiol. 2014, 108, 16–20. [Google Scholar] [CrossRef] [PubMed]
- Tang, W.; Xu, X.; Shen, G.; Chen, J. Effect of Environmental Factors on Germination and Emergence of Aryloxyphenoxy Propanoate Herbicide-Resistant and -Susceptible Asia Minor Bluegrass (Polypogon fugax). Weed Sci. 2015, 63, 669–675. [Google Scholar] [CrossRef]
- Tang, W.; Chen, J.; Yu, X.; Zhang, J.; Lu, Y. Growth, fecundity, and competition between aryloxyphenoxypropionate-resistant and -susceptible Asia Minor bluegrass (Polypogon fugax). Weed Sci. 2019, 67, 546–551. [Google Scholar] [CrossRef]
- Zhou, F.Y.; Zhang, Y.; Tang, W.; Gao, T. Transcriptomics analysis of the flowering regulatory genes involved in the herbicide resistance of Asia minor bluegrass (Polypogon fugax). BMC Genom. 2017, 18, 953. [Google Scholar] [CrossRef]
- Yu, Q.; Han, H.; Vila-Aiub, M.M.; Powles, S.B. AHAS herbicide resistance endowing mutations: Effect on AHAS functionality and plant growth. J. Exp. Bot. 2010, 61, 3925–3934. [Google Scholar] [CrossRef]
- Ashigh, J.; Tardif, F. An Ala205Val substitution in acetohydroxyacid synthase of Eastern black nightshade (Solanum ptychanthum) reduces sensitivity to herbicides and feedback inhibition. Weed Sci. 2007, 55, 558–565. [Google Scholar] [CrossRef]
- Purrington, C.B.; Bergelson, J. Exploring the physiological basis of costs of herbicide resistance in Arabidopsis thaliana. Am. Nat. 1999, 154, S82–S91. [Google Scholar] [CrossRef]
- Paris, M.; Roux, F.; Berard, A.; Reboud, X. The effects of the genetic background on herbicide resistance fitness cost and its associated dominance in Arabidopsis thaliana. Heredity 2008, 101, 499–506. [Google Scholar] [CrossRef]
- Frenkel, E.; Matzrafi, M.; Rubin, B.; Peleg, Z. Effects of environmental conditions on the fitness penalty in herbicide resistant Brachypodium hybridum. Front. Plant Sci. 2017, 8, 94. [Google Scholar] [CrossRef] [PubMed]
- Williams, M.M.I.; Jordan, N.; Yerkes, C. The fitness cost of triazine resistance in jimsonweed (Datura stramonium L.). Am. Midl. Nat. 1995, 133, 131–137. [Google Scholar]
- Han, C.; Yang, P. Studies on the molecular mechanisms of seed germination. Proteomics 2015, 15, 1671–1679. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Liu, S.; Yuan, G.; Zhao, P.; Yang, W.; Jia, J.; Cheng, L.; Qi, D.; Chen, S.; Liu, G. Comparative transcriptome analysis provides insights into the distinct germination in sheepgrass (Leymus chinensis) during seed development. Plant Physiol. Biochem. 2019, 139, 446–458. [Google Scholar] [CrossRef]
- Dal Degan, F.; Rocher, A.; Cameron-Mills, V.; von Wettstein, D. The expression of serine carboxypeptidases during maturation and germination of the barley grain. Proc. Natl. Acad. Sci. USA 1994, 91, 8209–8213. [Google Scholar] [CrossRef]
- Brown, A.P.; Slabas, A.R.; Rafferty, J.B. Fatty Acid Biosynthesis in Plants Metabolic Pathways, Structure and Organization. In Lipids in Photosynthesis. Advances in Photosynthesis and Respiration; Wada, H., Murata, N., Eds.; Springer: Dordrecht, The Netherlands, 2009; Volume 30. [Google Scholar]
- Ohlrogge, J.; Browse, J. Lipid Biosynthesis. Plant Cell 1995, 7, 957–970. [Google Scholar]
- Vila-Aiub, M.M.; Neve, P.; Steadman, K.J.; Powles, S.B. Ecological fitness of a multiple herbicide-resistant Lolium rigidum population: Dynamics of seed germination and seedling emergence of resistant and susceptible phenotypes. J. Appl. Ecol. 2005, 42, 288–298. [Google Scholar] [CrossRef]
- Délye, C.; Menchari, Y.; Michel, S.; Cadet, É.; Le Corre, V. A new insight into arable weed adaptive evolution: Mutations endowing herbicide resistance also affect germination dynamics and seedling emergence. Ann. Bot. 2013, 111, 681–691. [Google Scholar] [CrossRef]
- Vila-Aiub, M.M. Fitness of Herbicide-Resistant Weeds: Current Knowledge and Implications for Management. Plants 2019, 8, 469. [Google Scholar] [CrossRef]
- Nikolau, B.J.; Ohlrogge, J.B.; Wurtele, E.S. Plant biotin-containing carboxylases. Arch. Biochem. Biophys. 2003, 414, 211–222. [Google Scholar] [CrossRef]
- Zhang, H.; Tweel, B.; Tong, L. Molecular basis for the inhibition of the carboxytransferase domain of acetyl-coenzyme-A carboxylase by haloxyfop and diclofop. Proc. Natl. Acad. Sci. USA 2004, 101, 5910–5915. [Google Scholar] [CrossRef]
- Rylott, E.L. The gluconeogenic enzyme phosphoenolpyruvate carboxykinase in Arabidopsis is essential for seedling establishment. Plant Physiol. 2009, 131, 1834–1842. [Google Scholar] [CrossRef] [PubMed]
- Sew, Y.S.; Stroher, E.; Fenske, R.; Millar, A.H. Loss of mitochondrial malate dehydrogenase activity alters seed metabolism impairing seed maturation and post-germination growth in Arabidopsis. Plant Physiol. 2016, 171, 849–863. [Google Scholar] [CrossRef] [PubMed]
- Pracharoenwattana, I.; Smith, C.S.M. Arabidopsis peroxisomal citrate synthase is required for fatty acid respiration and seed germination. Plant Cell 2005, 17, 2037–2048. [Google Scholar] [CrossRef] [PubMed]
- Shu, K.; Liu, X.; Xie, Q.; He, Z. Two Faces of One Seed: Hormonal Regulation of Dormancy and Germination. Mol. Plant 2016, 9, 34–45. [Google Scholar] [CrossRef]
- Sun, T.P.; Kamiya, Y. The Arabidopsis GA1 locus encodes the cyclase ent-kaurene synthetase A of gibberellin biosynthesis. Plant Cell 1994, 6, 1509–1518. [Google Scholar]
- Lee, S.; Cheng, H.; King, K.E.; Wang, W.; Peng, J. Gibberellin regulates Arabidopsis seed germination via rgl2, a GAI/RGA-like gene whose expression is up-regulated following imbibition. Gene Dev. 2002, 16, 646–658. [Google Scholar] [CrossRef]
- Yamaguchi, S.; Smith, M.W.; Brown Robert, G.S.; Kamiya, Y.; Sun, T. Phytochrome regulation and differential expression of gibberellin 3β-hydroxylase genes in germinating Arabidopsis seeds. Plant Cell 1998, 10, 2115–2126. [Google Scholar] [CrossRef]
- De Lucas, M.; Davière, J.M.; Rodríguez-Falcón, M.; Pontin, M.; Iglesias-Pedraz, J.M.; Lorrain, S.; Fankhauser, C.; Blázquez, M.A.; Titarenko, E.; Prat, S. A molecular framework for light and gibberellin control of cell elongation. Nature 2008, 451, 480–484. [Google Scholar] [CrossRef]
- Lefebvre, V.; North, H.; Frey, A.; Sotta, B.; Seo, M.; Okamoto, M.; Nambara, E.; Marion-Poll, A. Functional analysis of Arabidopsis NCED6 and NCED9 genes indicates that ABA synthesized in the endosperm is involved in the induction of seed dormancy. Plant J. 2006, 45, 309–319. [Google Scholar] [CrossRef]
- Lindgren, L.O.; Stalberg, K.G.; Hoglund, A.S. Seed-specific overexpression of an endogenous Arabidopsis phytoene synthase gene results in delayed germination and increased levels of carotenoids, chlorophyll, and abscisic acid. Plant Physiol. 2003, 132, 779–785. [Google Scholar] [CrossRef]
- Chae, M.J.; Lee, J.S.; Nam, M.H.; Cho, K.; Hong, J.Y.; Yi, S.A.; Suh, S.C.; Yoon, I.S. A rice dehydration-inducible SNF1-related protein kinase 2 phosphorylates an abscisic acid responsive element-binding factor and associates with ABA signaling. Plant Mol. Biol. 2007, 63, 151–169. [Google Scholar] [CrossRef] [PubMed]
- Fujii, H.; Zhu, J.K. Arabidopsis mutant deficient in 3 abscisic acid-activated protein kinases reveals critical roles in growth, reproduction, and stress. Proc. Natl. Acad. Sci. USA 2009, 106, 8380–8385. [Google Scholar] [CrossRef] [PubMed]
- Bradford, M.M. A rapid method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Hansen, J.; Møller, I. Percolation of starch and soluble carbohydrates from plant tissue for quantitative determination with anthrone. Anal. Biochem. 1975, 68, 87–94. [Google Scholar] [CrossRef]
- Grabherr, M.G.; Haas, B.J.; Yassour, M.; Levin, J.Z.; Thompson, D.A.; Amit, I.; Adiconis, X.; Fan, L.; Raychowdhury, R.; Zeng, Q.; et al. Full-length transcriptome assembly from RNA-Seq data without a reference genome. Nat. Biotechnol. 2011, 29, 644–652. [Google Scholar] [CrossRef]
- Zheng, Y.; Jiao, C.; Sun, H.; Rosli, H.G.; Pombo, M.A.; Zhang, P.; Banf, M.; Dai, X.; Martin, G.B.; Giovannoni, J.J.; et al. iTAK: A program for genome-wide prediction and classification of plant transcription factors, transcriptional regulators, and protein kinases. Mol. Plant 2016, 9, 1667–1670. [Google Scholar] [CrossRef]
- Trapnell, C.; Williams, B.A.; Pertea, G.; Mortazavi, A.; Kwan, G.; van Baren, M.J.; Salzberg, S.L.; Wold, B.J.; Pachter, L. Transcript assembly and quantification by RNA-Seq reveals unannotated transcripts and isoform switching during cell differentiation. Nat. Biotechnol. 2010, 28, 511–515. [Google Scholar] [CrossRef]
- Love, M.I.; Huber, W.; Anders, S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014, 15, 550. [Google Scholar] [CrossRef]
- Young, M.D.; Wakefield, M.J.; Smyth, G.K. Gene ontology analysis for RNA-seq: Accounting for selection bias. Genome Biol. 2010, 11, R14. [Google Scholar] [CrossRef]
- Mao, X.; Cai, T.; Olyarchuk, J.G.; Wei, L. Automated genome annotation and pathway identification using the KEGG Orthology (KO) as a controlled vocabulary. Bioinformatics 2005, 21, 3787–3793. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene ID | log2FC (LS_6d vs. LS_3d) | log2FC (LR_6d vs. LR_3d) | Encoded Protein | Description |
|---|---|---|---|---|
| Carbohydrate metabolism | ||||
| Cluster-37472.6545 | 7.36 | 3.84 | GPI | glucose-6-phosphate isomerase |
| Cluster-37472.71503 | 5.65 | 2.84 | FBA | fructose-bisphosphate aldolase |
| Cluster-40718.0 | 5.96 | 4.88 | PDHA | pyruvate dehydrogenase E1 component alpha subunit |
| Cluster-35171.2 | 6.98 | 5.61 | PCKA | phosphoenolpyruvate carboxykinase (ATP) |
| Cluster-37472.39187 | 3.80 | 1.85 | ADH | alcohol dehydrogenase class-P |
| Cluster-37472.47005 | 4.19 | 2.04 | ADH | alcohol dehydrogenase class-P |
| Cluster-37472.85108 | 8.43 | 6.07 | MDH | malate dehydrogenase |
| Cluster-15133.3 | 6.95 | 3.19 | MDH | malate dehydrogenase |
| Cluster-37472.21027 | 9.41 | 4.35 | CS | citrate synthase |
| Cluster-33485.0 | 6.88 | 5.55 | OGDH | 2-oxoglutarate dehydrogenase E1 component |
| Cluster-38121.0 | 7.15 | 5.66 | LSC1 | succinyl-CoA synthetase alpha subunit |
| Cluster-34010.0 | 5.12 | 3.97 | LSC2 | succinyl-CoA synthetase beta subunit |
| Cluster-37472.82495 | 9.25 | 6.45 | SDHB | succinate dehydrogenase (ubiquinone) iron-sulfur subunit |
| Cluster-39676.0 | 7.51 | 4.56 | SDHB | succinate dehydrogenase (ubiquinone) iron-sulfur subunit |
| Cluster-41938.0 | 7.25 | 4.02 | PYC | pyruvate carboxylase |
| Cluster-37472.72264 | 8.37 | 3.93 | PGD | 6-phosphogluconate dehydrogenase |
| Cluster-37472.83492 | 6.66 | 5.71 | TKT | transketolase |
| Cluster-37472.237 | 7.72 | 4.88 | gntK | gluconokinase |
| Fatty acid metabolism | ||||
| Cluster-39490.0 | 7.28 | 4.20 | ACACA | acetyl-CoA carboxylase/biotin carboxylase 1 |
| Cluster-31226.0 | 7.10 | 4.82 | FabG | 3-oxoacyl-[acyl-carrier protein] reductase |
| Cluster-37472.4287 | 9.16 | 5.52 | ACSL | long-chain acyl-CoA synthetase |
| Cluster-33548.0 | 9.20 | 5.34 | ACAA1 | acetyl-CoA acyltransferase 1 |
| Cluster-37472.85335 | 9.90 | 5.52 | ACADM | acyl-CoA dehydrogenase |
| Cluster-30044.0 | 7.77 | 4.75 | ACAT | acetyl-CoA C-acetyltransferase |
| Hormones biosynthesis and signal transduction | ||||
| Cluster-37472.24032 | 9.69 | 3.87 | KS | ent-copalyl diphosphate synthase |
| Cluster-37472.23672 | 6.42 | 4.78 | KS | ent-kaurene synthase |
| Cluster-37472.70387 | 3.31 | 2.02 | GA3ox | gibberellin 3-beta-dioxygenase |
| Cluster-51396.0 | −1.69 | 3.01 | PIF4 | phytochrome-interacting factor 4 |
| Cluster-37472.23110 | −1.97 | 1.05 | NCED | 9-cis-epoxycarotenoid dioxygenase |
| Cluster-37472.23992 | 2.73 | 3.36 | PSY | phytoene synthase |
| Cluster-37472.76697 | 1.64 | 2.53 | LcyE | lycopene epsilon-cyclase |
| Cluster-37472.6593 | 1.32 | 2.78 | SNRK2 | serine/threonine-protein kinase SRK2 |
| Cluster-48355.0 | 1.63 | 2.58 | ABF | ABA responsive element binding factor |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, X.; Tang, W.; Yang, Y.; Zhang, J.; Lu, Y. Comparative Transcriptome Analysis Revealing the Different Germination Process in Aryloxyphenoxypropionate-Resistant and APP-Susceptible Asia Minor Bluegrass (Polypogon fugax). Plants 2020, 9, 1191. https://doi.org/10.3390/plants9091191
Yu X, Tang W, Yang Y, Zhang J, Lu Y. Comparative Transcriptome Analysis Revealing the Different Germination Process in Aryloxyphenoxypropionate-Resistant and APP-Susceptible Asia Minor Bluegrass (Polypogon fugax). Plants. 2020; 9(9):1191. https://doi.org/10.3390/plants9091191
Chicago/Turabian StyleYu, Xiaoyue, Wei Tang, Yongjie Yang, Jianping Zhang, and Yongliang Lu. 2020. "Comparative Transcriptome Analysis Revealing the Different Germination Process in Aryloxyphenoxypropionate-Resistant and APP-Susceptible Asia Minor Bluegrass (Polypogon fugax)" Plants 9, no. 9: 1191. https://doi.org/10.3390/plants9091191
APA StyleYu, X., Tang, W., Yang, Y., Zhang, J., & Lu, Y. (2020). Comparative Transcriptome Analysis Revealing the Different Germination Process in Aryloxyphenoxypropionate-Resistant and APP-Susceptible Asia Minor Bluegrass (Polypogon fugax). Plants, 9(9), 1191. https://doi.org/10.3390/plants9091191