Exploiting Biological Nitrogen Fixation: A Route Towards a Sustainable Agriculture

 , ,
, ,  and
and
Abstract
1. Introduction
2. Plant Available Nitrogen
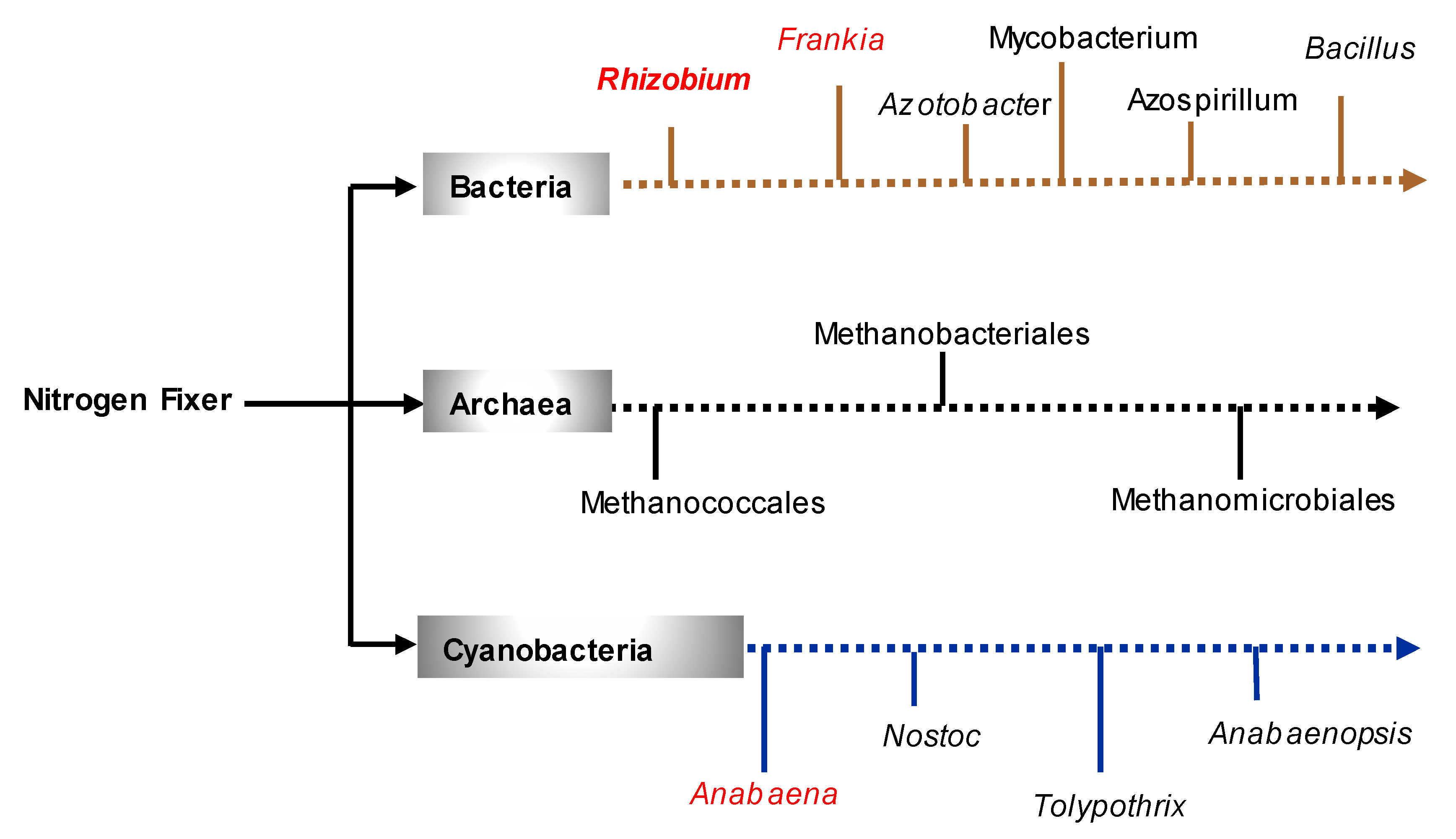
3. Nitrogen Fixation
4. Retrospect on the Isolation of Nitrogen-Fixing Bacteria and Launch of N-Fixing Biofertilizers
4.1. Discovery
4.2. Commercialization
5. BNF and their Contribution to Agriculture
5.1. Inoculants for Legume Crops
5.2. Inoculants for Non-Legume Crops
5.3. Success-Limiting Factors of BNF Application in Agriculture
5.4. Beneficial Mechanisms Other Than N-Fixation Provides by Diazotrophs Bacteria
5.5. Synergistic Benefits
6. Challenges of Extending the BNF Ability to Non-Legumes
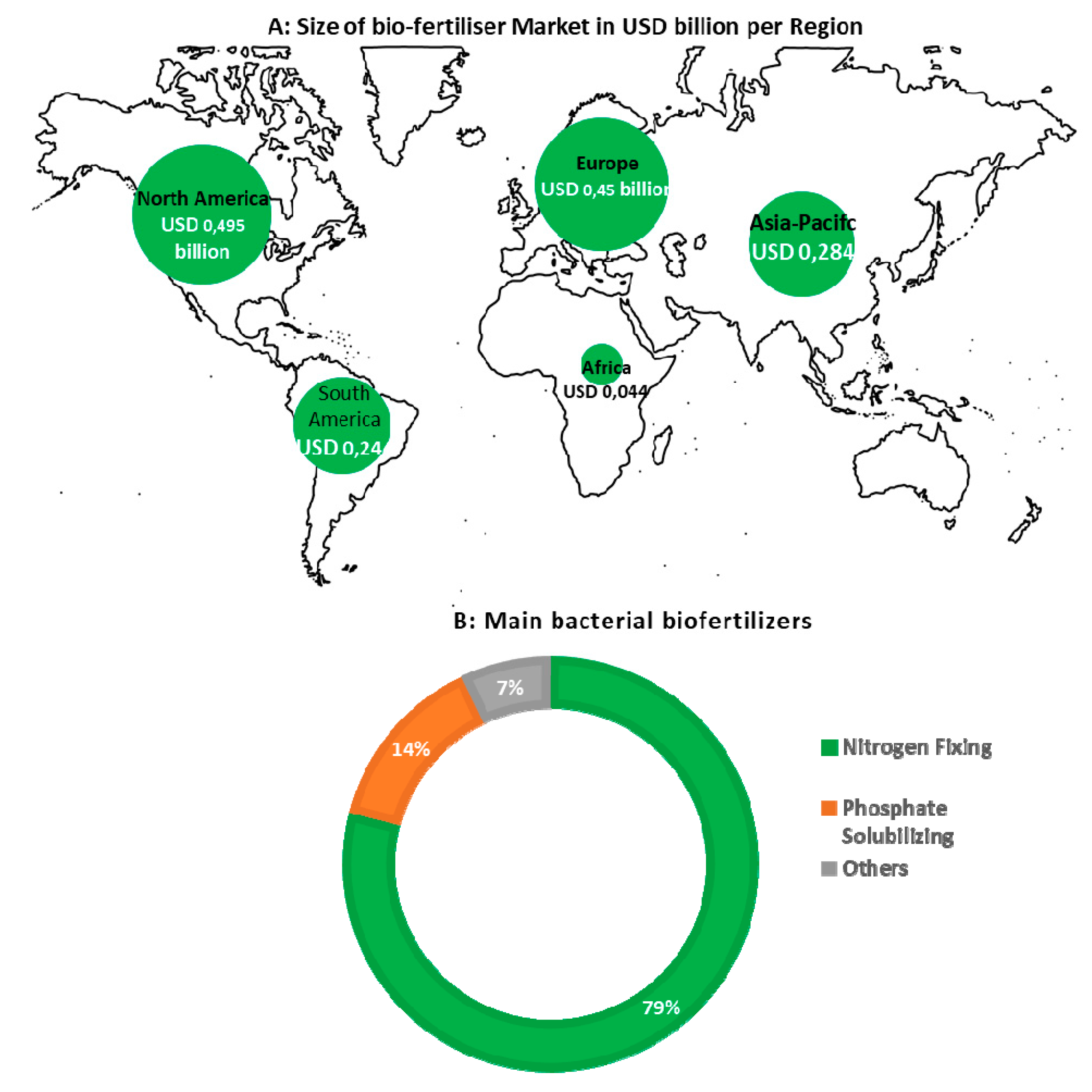
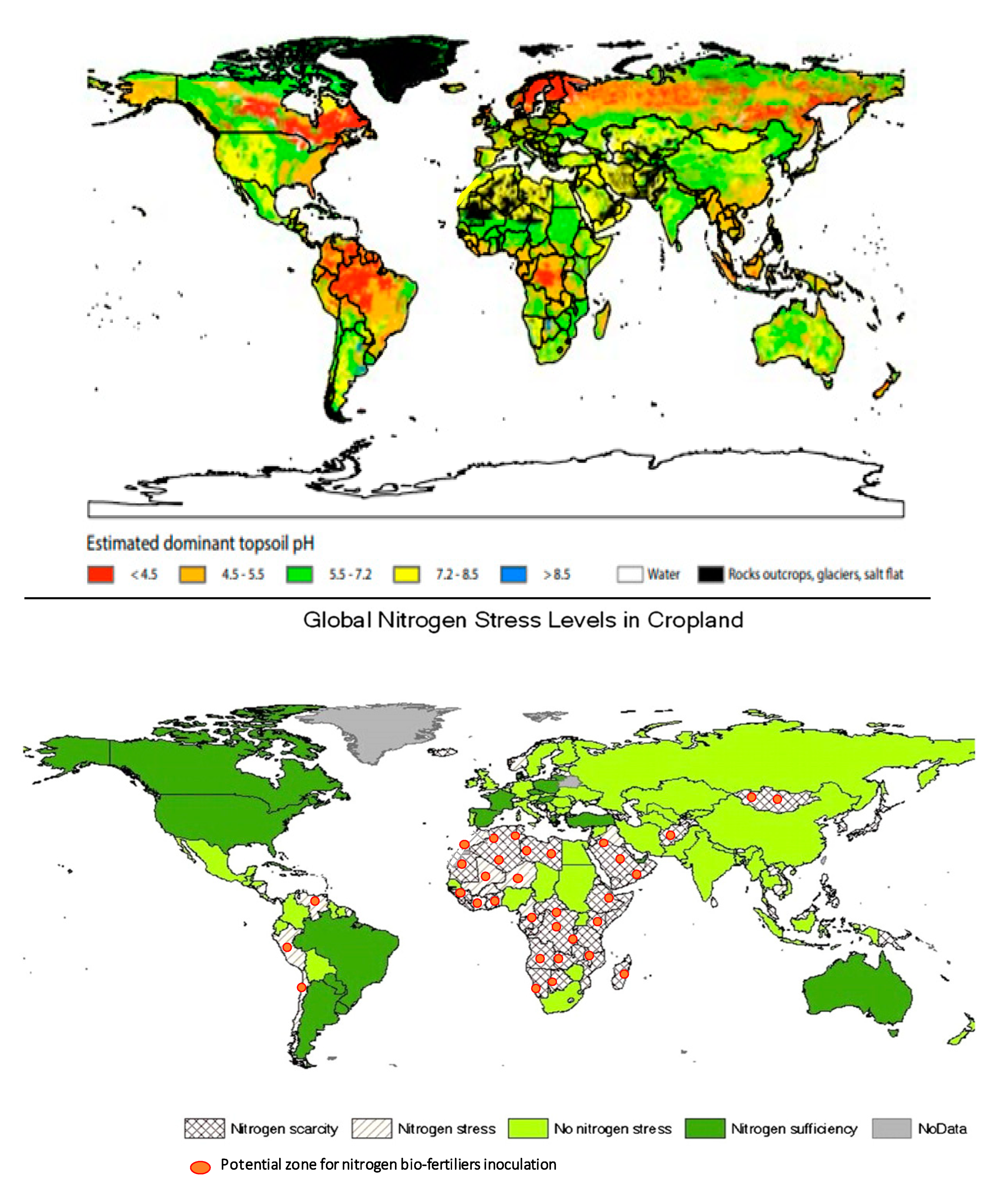
7. Road Map for Successful and Large-Scale Adoption of N-Fixing Biofertilizers
8. Conclusion and Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- FAO, World fertilizer trends and outlook to 2018. Food and Agriculture Organization of the United Nations—Rome 2015. Available online: www.fao.org/publication (accessed on 12 May 2020).
- World Religion Database: International Religious Demographic Statistics and Sources; Johnson, T.M., Grim, B.J., Eds.; Brill: Leiden, The Netherlands; Boston, MA, USA, 2010; Available online: http://www.worldreligiondatabase.org (accessed on 10 May 2020).
- Hirel, B.; Tétu, T.; Lea, P.J.; Dubois, F. Improving Nitrogen Use Efficiency in Crops for Sustainable Agriculture. Sustainabilty 2011, 3, 1452–1485. [Google Scholar] [CrossRef]
- Mueller, N.D.; Gerber, J.S.; Johnston, M.; Ray, D.K.; Ramankutty, N.; Foley, J.A. Closing yield gaps through nutrient and water management. Nature 2012, 490, 254–257. [Google Scholar] [CrossRef]
- Fertilizer Consumption. The World Bank Data Bank. 2013. Available online: http://data.worldbank.org/indicator/AG.CON.FERT.ZS?view=chart (accessed on 5 May 2020).
- Liu, J.; Ma, K.; Ciais, P.; Polasky, S. Reducing human nitrogen use for food production. Sci. Rep. 2016, 6, 30104. [Google Scholar] [CrossRef] [PubMed]
- IEA/IRENA, Perspectives for the Energy Transition (IEA/IRENA, 2017). Available online: go.nature.com/2pgkfwd (accessed on 5 May 2020).
- FAO. Plant Nutrition for Food Security; Fertilizer and Plant Nutrition Bulletin No. 16; Publishing Management Service Information Division FAO: Rome, Italy, 2006; p. 366. [Google Scholar]
- Maheswari, M.; Murthy, A.N.G.; Shanker, A.K. Nitrogen Nutrition in Crops and Its Importance in Crop Quality—In: The Indian nitrogen assessment: Sources of reactive nitrogen, environmental and climate effects, management options, and policies. Agron. Hortic. 2017, 175–186. [Google Scholar]
- Yang, X.; Fang, S. Practices, perceptions, and implications of fertilizer use in East-Central China. Ambio 2015, 44, 647–652. [Google Scholar] [CrossRef] [PubMed]
- Usha, B. Agriculture and the Dark Side of Chemical Fertilizers. Environ. Anal. Ecol. Stud. 2018, 1, 3. [Google Scholar]
- Zheng, M.; Zhou, Z.; Luo, Y.; Zhao, P.; Mo, J. Global pattern and controls of biological nitrogen fixation under nutrient enrichment: A meta-analysis. Glob. Chang. Biol. 2019, 25, 3018–3030. [Google Scholar] [CrossRef] [PubMed]
- Araujo, A.; Leite, L.; De Iwata, B.; De Lira, M.; Xavier, G.; Figueiredo, M.V.-B. Microbiological process in agroforestry systems. A review. Agronomy for Sustainable Development. Agron. Sustain. Dev. 2012, 32, 215–226. [Google Scholar] [CrossRef]
- Shah, F.; Wu, W. Soil and Crop Management Strategies to Ensure Higher Crop Productivity within Sustainable Environments. Sustainability 2019, 11, 1485. [Google Scholar] [CrossRef]
- Saikia, S.P.; Jain, V. Biological nitrogen fixation with non-legumes: An achievable target or a dogma. Curr. Sci. 2007, 92, 317–322. [Google Scholar]
- Sur, S.; Bothra, A.K.; Sen, A. Symbiotic Nitrogen Fixation-A Bioinformatics Perspective. Biotechnology 2010, 9, 257–273. [Google Scholar] [CrossRef]
- Ravikumar, S.; Kathiresan, K.; Alikhan, S.L.; Williams, G.P.; Anitha, N.; Gracelin, A. Growth of Avicennia marina and Ceriops decandra seedlings inoculated with Halophilic Azotobacters. J. Environ. Biol. 2007, 28, 601–603. [Google Scholar] [PubMed]
- Ininbergs, K.; Bay, G.; Rasmussen, U.; Wardle, D.A.; Nilsson, M.C. Composition and diversity of nifH genes of nitrogen-fixing cyanobacteria associated with boreal forest feather mosses. N. Phytol. 2011, 192, 507–517. [Google Scholar] [CrossRef] [PubMed]
- Welte, C.U. Revival of Archaeal Methane Microbiology. mSystems 2018, 3, e00181-17. [Google Scholar] [CrossRef]
- Santi, C.; Bogusz, D.; Franche, C. Biological nitrogen fixation in non-legume plants. Ann. Bot. 2013, 111, 743–767. [Google Scholar] [CrossRef]
- Mus, F.; Crook, M.B.; Garcia, K.; Garcia Costas, A.; Geddes, B.A.; Kouri, E.D.; Paramasivan, P.; Ryu, M.-H.; Oldroyd, G.E.; Poole, P.S.; et al. Symbiotic nitrogen fixation and the challenges to its extension to nonlegumes. Appl. Environ. Microbiol. 2016, 82, 3698–3710. [Google Scholar] [CrossRef]
- Liu, C.W.; Murray, J.D. The Role of Flavonoids in Nodulation Host-Range Specificity: An Update. Plants 2016, 5, 33. [Google Scholar] [CrossRef]
- Jimenez-Jimenez, S.; Santana, O.; LaraRojas, F.; Arthikala, M.K.; Armada, E.; Hashimoto, K.; Kuchitsu, K.; Salgado, S.; Aguirre, J.; Quinto, C.; et al. Differential tetraspanin genes expression and subcellular localization during mutualistic interactions in Phaseolus vulgaris. PLoS ONE 2019, 14, e0219765. [Google Scholar] [CrossRef]
- Cissoko, M.; Hocher, V.; Gherbi, H.; Gully, D.; Carré-Mlouka, A.; Sane, S.; Pignoly, S.; Champion, A.; Ngom, M.; Pujic, P.; et al. Actinorhizal Signaling Molecules: Frankia Root Hair Deforming Factor Shares Properties With NIN Inducing Factor. Front. Plant. Sci. 2018, 9, 1494. [Google Scholar] [CrossRef]
- Suzaki, T.; Takeda, N.; Nishida, H.; Hoshino, M.; Ito, M.; Misawa, F.; Handa, Y.; Miura, K. Masayoshi KawaguchiCorrection: Lack of symbiont accommodation controls intracellular symbiont accommodation in root nodule and arbuscular mycorrhizal symbiosis in Lotus japonicus. PLoS Genet. 2019, 15, e1007966. [Google Scholar]
- Mugabe, J. Research on biofertilizers: Kenya, Zimbabwe and Tanzania, Biotechnol. Dev. Monitor. 1994, 18, 9–10. [Google Scholar]
- Chianu, J.N.; Nkonya, E.M.; Mairura, F.S.; Chianu, J.N.; Akinnifesi, F.K. Biological nitrogen fixation and socioeconomic factors for legumeproduction in sub-Saharan Africa: A review. Agron. Sustain. Dev. 2011, 31, 139–154. [Google Scholar] [CrossRef]
- Beyan, S.M.; Wolde-meskel, E.; Dakora, F.D. An assessment of plant growth and N2 fixation in soybean genotypes grown in uninoculated soils collected from different locations in Ethiopia. Symbiosis 2018, 75, 189–203. [Google Scholar] [CrossRef] [PubMed]
- Van Heerwaarden, J.; Baijukya, F.; Kyei-Boahen, S.; Adjei-Nsiah, S.; Ebanyat, P.; Kamai, N.; Wolde-meskel, E.; Kanampiu, F.; Vanlauwe, B.; Giller, K. Soyabean response to rhizobium inoculation across sub-Saharan Africa: Patterns of variation and the role of promiscuity. Agric. Ecosyst. Environ. 2018, 261, 211–218. [Google Scholar] [CrossRef] [PubMed]
- Dent, D.; Cocking, E.C. Establishing symbiotic nitrogen fixation in cereals and other non-legume crops: The Greener Nitrogen Revolution. Agric. Food Secur. 2017, 6, 7. [Google Scholar] [CrossRef]
- Smercina, D.N.; Evans, S.E.; Friesen, M.L.; Tiemann, L.K. To Fix or Not to Fix: Controls on Free-Living Nitrogen-Fixation in the rhizosphere. Appl. Environ. Microbiol. 2019, 85, 6–22. [Google Scholar] [CrossRef] [PubMed]
- Peoples, M.B.; Craswell, E.T. Biological nitrogen-fixation—Investments, expectations and actual contributions to agriculture. Plant Soil 1992, 141, 13–39. [Google Scholar] [CrossRef]
- Kennedy, I.R.; Islam, N. The current and potential contribution of asymbiotic nitrogen fixation to nitrogen requirements onfarms: A review. Aust. J. Exp. Agric. 2001, 41, 447–457. [Google Scholar] [CrossRef]
- Oliveira, A.L.M.; Canuto, E.L.; Reis, V.M.; Baldani, J.I. Response of micropropagated sugarcane varieties to inoculation with endophytic diazotrophic bacteria. Braz. J. Microbiol. 2003, 34, 59–61. [Google Scholar] [CrossRef]
- Pankievicz, V.C.S.; do Amaral, F.P.; Santos, K.F.D.N.; Agtuca, B.; Xu, Y.; Schueller, M.J.; Arisi, A.C.M.; Steffens, M.B.R.; de Souza, E.M.; Pedrosa, F.O.; et al. Robust biological nitrogen fixation in a model grass-bacterial association. Plant J. 2015, 81, 907–919. [Google Scholar] [CrossRef] [PubMed]
- Van Deynze, A.; Zamora, P.; Delaux, P.-M.; Heitmann, C.; Jayaraman, D.; Rajasekar, S.; Graham, D.; Maeda, J.; Gibson, D.; Schwartz, K.D.; et al. Nitrogen fixation in a landrace of maize is supported by a mucilage-associated diazotrophic microbiota. PLoS Biol. 2018, 16, e2006352. [Google Scholar] [CrossRef]
- Liu, C.W.; Sung, Y.; Chen, B.C. Effects of Nitrogen Fertilizers on the Growth and Nitrate Content of Lettuce (Lactuca sativa L.). Int. J. Environ. Res. Public Health 2014, 11, 4427–4440. [Google Scholar] [CrossRef] [PubMed]
- Tamme, T.; Reinik, M.; Roasto, M. Nitrates and Nitrites in Vegetables: Occurrence and Health Risks. In Bioactive Foods Promoting Health: Fruits and Vegetables; Watson, R.R., Preedy, V.R., Eds.; Academic Press: Salt Lake City, UT, USA, 2009; pp. 307–321. [Google Scholar]
- Rosswall, T. The internal nitrogen cycle between microorganisms, vegetation and soil. Ecol. Bull. 1976, 22, 157–167. [Google Scholar]
- Stein, L.Y.; Klotz, M.G. The nitrogen cycles. Curr. Biol. 2016, 26, 83–101. [Google Scholar] [CrossRef] [PubMed]
- Keramidas, K.; Tchung-Ming, S.; Diaz-Vazquez, A.; Weitzel, M.; Vandyck, T.; Després, J.; Schmitz, A.; Rey, L.; Los Santos, K.; Wojtowicz, B.; et al. Soria-Ramirez, Global Energy and Climate Outlook 2018: Sectoral Mitigation Options Towards a Low-Emissions Economy—Global Context to the EU Strategy for long-Term Greenhouse Gas Emissions Reduction; EUR 29462 EN; Publications Office of the European Union: Luxembourg, 2018; ISBN 978-92-79-97462-5. [Google Scholar]
- Sammauria, R.; Kumawat, S.; Kumawat, P.; Singh, J.K.; Jatwa, T. Microbial inoculants: Potential tool for sustainability of agricultural production systems. Arch. Microbiol. 2020, 202, 677–693. [Google Scholar] [CrossRef] [PubMed]
- Braakhekke, M.C.; Rebel, K.T.; Dekker, S.C.; Smith, B.; Beusen, A.H.W.; Wassen, M.J. Nitrogen leaching from natural ecosystems under global change: A modelling study. Earth Syst. Dynam. 2017, 8, 1121–1139. [Google Scholar] [CrossRef]
- Mabrouk, Y.; Hemissi, I.; Salem, I.B.; Mejri, S.; Saidi, M.; Belhadj, O. Potential of Rhizobia in Improving Nitrogen Fixation and Yields of Legumes. Symbiosis 2018, 107. [Google Scholar] [CrossRef]
- Gueye, M.; Bordeleau, L.M. Nitrogen fixation in Bambara groundnut (Vigna subterranea (L.) Thouars. Mircen. J. 1988, 4, 365–375. [Google Scholar] [CrossRef]
- Igarashi, R.Y.; Seefeldt, L.C. Nitrogen fixation: The mechanism of the Modependent nitrogenase. Crit. Rev. BMC Mol. Biol. 2003, 38, 351–384. [Google Scholar] [CrossRef]
- Buscot, F.; Varma, A. Microorganisms in Soils: Roles in Genesis and Functions; Springer Science Business Media: Berlin/Heidelberg, Germany, 2005; Volume 3, p. 422. [Google Scholar]
- Gilchrist, M.; Benjamin, N. From Atmospheric Nitrogen to Bioactive Nitrogen Oxides. In Nitrite and Nitrate in Human Health and Disease, 2nd ed.; Bryan, N.S., Loscalzo, J., Eds.; Humana Press: New York, NY, USA, 2017; pp. 11–20. [Google Scholar] [CrossRef]
- Burén, S.; Rubio, L.M. State of the art in eukaryotic nitrogenase engineering. FEMS Microbiol. Lett. 2018, 365, fnx274. [Google Scholar] [CrossRef]
- Dai, Z.; Guo, X.; Yin, H.; Liang, Y.; Cong, J.; Liu, X. Identification of Nitrogen-Fixing Genes and Gene Clusters from Metagenomic Library of Acid Mine Drainage. PLoS ONE 2014, 9, e87976. [Google Scholar] [CrossRef] [PubMed]
- Seefeldt, L.C.; Hoffman, B.M.; Peters, J.W.; Raugei, S.; Beratan, D.N.; Antony, E.; Dean, D.R. Energy Transduction in Nitrogenase. Acc. Chem. Res. 2018, 51, 2179–2186. [Google Scholar] [CrossRef] [PubMed]
- Nonaka, A.H.; Yamamoto, N.; Kamiya, H.; Kotani, H.; Yamakawa, R.; Tsujimoto, Y. Fujita, Accessory Proteins of the Nitrogenase Assembly, NifW, NifX/NafY, and NifZ, Are Essential for Diazotrophic Growth in the Non heterocystous Cyanobacterium Leptolyngbya boryana. Front. Microbiol. 2019, 10, 495. [Google Scholar] [CrossRef]
- Harris, D.F.; Lukoyanov, D.A.; Shaw, S.; Compton, P.; Tokmina-Lukaszewska, M.; Bothner, B.; Kelleher, N.; Dean, D.R.; Hoffman, B.M.; Seefeldt, L.C. Mechanism of N2 Reduction Catalyzed by Fe-Nitrogenase Involves Reductive Elimination of H2. Biochemistry 2018, 57, 701–710. [Google Scholar] [CrossRef]
- Berman-Frank, I.; Chen, Y.B.; Gerchman, Y.; Dismukes, G.C.; Falkowski, P.G. Inhibition of nitrogenase by oxygen in marine cyanobacteria controls the global nitrogen and oxygen cycles. Biogeosci. Discuss. 2005, 2, 261–273. [Google Scholar] [CrossRef]
- Fay, P. Oxygen Relations of Nitrogen Fixation in Cyanobacteria. Microbiol. Rev. 1992, 56, 340–373. [Google Scholar] [CrossRef] [PubMed]
- Rice, W.A.; Paul, E.A. The organisms and biological processes involved in a symbiotic nitrogen fixation in waterlogged soil amended with straw. Can. J. Microbiol. 1972, 18, 715–723. [Google Scholar] [CrossRef]
- Ropera, M.M.; Gupta, V.V.S.R. Enhancing Non-symbiotic N2 Fixation in Agriculture. Open Agric. 2016, 10, 7–27. [Google Scholar] [CrossRef]
- Berman-Frank, I.P.; Lundgren, Y.B.; Chen, H.; Kupper, Z.; Kolber, B.; Bergman Falkowski, P. Segregation of nitrogen fixation and oxygenic photosynthesis in the marine Cyanobacterium trichodesmium. Science 2001, 294, 1534–1547. [Google Scholar] [CrossRef]
- Moshiri, F.; Crouse, B.R.; Johnson, M.K.; Maier, R.J. The “nitrogenase-protective” FeSII protein of Azotobacter vinelandii: Overexpression, characterization, and crystallization. Biochemistry 1995, 34, 12973–12982. [Google Scholar] [CrossRef]
- Kuhla, J.; Oelze, J. Dependence of nitrogenase switch-off upon oxygen stress on the nitrogenase activity in Azotobacter vinelandii. J. Bacteriol. 1988, 170, 5325–5329. [Google Scholar] [CrossRef] [PubMed]
- Sabra, W.; Zeng, A.-P.; Lünsdorf, H.; Deckwer, W.D. Effect of Oxygen on Formation and Structure of Azotobacter vinelandii Alginate and Its Role in Protecting Nitrogenase. Appl. Environ. Microbial. 2000, 66, 4037–4044. [Google Scholar] [CrossRef] [PubMed]
- Franche, C.; Lindström, K.; Elmerich, C. Nitrogen-fixing bacteria associated with leguminous and non-leguminous plants. Plant Soil 2009, 321, 35–59. [Google Scholar] [CrossRef]
- Beijerinck, M.W. Über ologonitrophile mikroben. Zentralbl. Bakteriol. Parasitenkd. Infektionskr. II Abt. Technol. Eng. 1901, 561–582. [Google Scholar]
- Winogradsky, S. Recherche sur assimilation de l’azote libre de l’atmosphere par ls microbes. Arch. Sci. Biol. 1895, 3, 297–352. [Google Scholar]
- Gurthie, F.B. inoculation of the soil for leguminous crops. Agric. Gaz. NSW 1896, 7, 690–694. [Google Scholar]
- Bullard, G.K.; Roughley, R.J.; Pulsford, D.J. The legume inoculant industry and inoculant quality control in Australia: 1953–2003. Aust. J. Exp. Agric. 2005, 45, 127. [Google Scholar] [CrossRef]
- Döbereiner, J. Azotobacter paspali sp. Nov. uma bactéria fixadora de nitrogênio na rizosfera de Paspalum. Pesq. Agropec. Bras. 1966, 1, 357–365. [Google Scholar]
- Baldani, J.I.; Baldani, V.L.D. History on the biological nitrogen fixation research in graminaceous plants: Special emphasis on the Brazilian experience. Anais da Academia Brasileira de Ciências 2005, 77, 549–579. Available online: www.scielo.br/aabc (accessed on 4 May 2020). [CrossRef]
- Döbereiner, J. Fixação de nitrogênio atmosférico em gramíneas tropicais. In Proceedings of the Congresso Brasileiro de Ciência do Solo, Colombia, Cali, 7 July 1976; Volume 15, pp. 593–602. [Google Scholar]
- Kizilkaya, R. Nitrogen fixation capacity of Azotobacter spp. strains isolated from soils in different ecosystems and relationship between them and the microbiological properties of soils. J. Environ. Biol. 2009, 30, 73–82. [Google Scholar]
- Stewart, W.D.P. Biological and ecological aspects of nitrogen fixation by free-living microorganisms. Proc. R. Soc. B 1969, 172, 367–388. [Google Scholar]
- Murray, P.A.; Zinder, S.H. Nitrogen fixation by a methanogenic archaebacterium. Nature 1984, 312, 284–286. [Google Scholar] [CrossRef]
- Leigh Nitrogen, J.A. Fixation in Methanogens: The Archaeal Perspective. Curr. Issues Mol. Biol. 2000, 2, 125–131. [Google Scholar]
- Belay, N.; Sparling, R.; Daniels, L. Dinitrogen fixation by a thermophilic methanogenic bacterium. Nature 1984, 312, 286–288. [Google Scholar] [CrossRef] [PubMed]
- Smil, V. Enriching the Earth; Massachusetts Institute of Technology: Cambridge, MA, USA, 2001. [Google Scholar]
- Timonin, M.I. Azotobacter Preparation (Azotogen) as a Fertilizer for Cultivated Plants1. Soil Sci. Soc. Am. J. 1949, 13, 246. [Google Scholar] [CrossRef]
- O’Hara, G.W. The role of nitrogen fixation in crop production. J. Crop. Prod. 1998, 1, 115–138. [Google Scholar] [CrossRef]
- Von Bulow, J.F.W.; Dobereiner, J. Potential for nitrogen fixation in maize genotypes in Brazil. Proc. Natl. Acad. Sci. USA 1975, 72, 2389–2393. [Google Scholar] [CrossRef]
- Parnell, J.J.; Berka, R.; Young, H.A.; Sturino, J.M.; Kang, Y.; Barnhart, D.M.; Di Leo, M.V. From the lab to the farm: An industrial perspective of plant beneficial microorganisms. Front Plant Sci. 2016, 7, 1110. [Google Scholar] [CrossRef]
- Gaby, J.B.; Buckley, D.H. A global census of nitrogenase diversity. Environ. Microbiol. 2011, 13, 1790–1799. [Google Scholar] [CrossRef]
- Khosro, M.; Yousef, S. Bacterial bio-fertilizers for sustainable crop production: A review. ARPN J. Agric. Biol. Sci. 2012, 7, 307–316. [Google Scholar]
- Swarnalakshmi, K.; Vandana, Y.; Senthilkumar, M.; Dolly, W.D. Bio fertilizers for higher pulse production in India: Scope, accessibility and challenges. Indian J. Agron. 2016, 61, 173–181. [Google Scholar]
- Karanja, N.; Freire, J.; Gueye, M.; DaSilva, E. MIRCEN Networking: Capacity-Building and BNF Technology Transfer in Africa and Latin America. AgBiotechNet 2000, 2, 1–5. [Google Scholar]
- Global Biofertilizers Market-Growth, Trends and Forecast. (2019–2024). Available online: https://www.mordorintelligence.com/industry-reports/global-biofertilizers-market-industry (accessed on 16 June 2020).
- Market Data Forecast 2018. Available online: https://www.marketdataforecast.com/market-reports/asia-pacific-biofertilizers-market (accessed on 15 June 2020).
- Vitousek, P.M.; Menge, D.N.L.; Reed, S.C.; Cleveland, C.C. Biological nitrogen fixation: Rates, patterns and ecological controls in terrestrial ecosystems. Philos. Trans. R. Soc. Lond. B. Biol. Sci. 2013, 368, 1621. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, M.V.B.; Mergulhão, A.E.S.; Sobral, J.K.; Junio, M.A.L.; Araújo, A.S.F. Biological Nitrogen Fixation: Importance, Associated Diversity, and Estimates. In Plant Microbe Symbiosis: Fundamentals and Advances; Arora, N.K., Ed.; Springer India: Berlin/Heidelberg, Germany, 2013; p. 267. [Google Scholar] [CrossRef]
- Rao, D.L.N.; Balachandar, D. Nitrogen inputs from Biological Nitrogen Fixation in Indian Agriculture. In The Indian Nitrogen Assessment. Sources of Reactive Nitrogen, Environmental and Climate Effects, Management Options, and Policies; Abrol, Y.P., Ed.; Elsevier: Amsterdam, The Netherlands, 2017; pp. 117–132. [Google Scholar]
- Hungria, M.; Mendes, I.C. Nitrogen Fixation with Soybean: The Perfect Symbiosis? Biological Nitrogen Fixation; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2015; Volume 2, pp. 1009–1024. [Google Scholar] [CrossRef]
- Gyogluu, C.; Jaiswal, S.K.; Kyei-Boahen, S.; Dakora, F.D. Identification and distribution of microsymbionts associated with soybean nodulation in Mozambican soils. Syst. Appl. Microbiol. 2018, 41, 506–515. [Google Scholar] [CrossRef] [PubMed]
- Aloril, E.T.; Babalola, O.O. Microbial Inoculants for Improving Crop Quality and Human Health in Africa. Front. Microbial. 2018, 9, 2213. [Google Scholar] [CrossRef] [PubMed]
- Thilakarathna, M.S.; Chapagain, T.; Ghimire, B.; Pudasaini, R.; Tamang, B.B.; Gurung, K.; Choi, K.; Rai, L.; Magar, S.; Bishnu, B.K.; et al. Evaluating the Effectiveness of Rhizobium Inoculants and Micronutrients as Technologies for Nepalese Common Bean Smallholder Farmers in the Real-World Context of Highly Variable Hillside Environments and Indigenous Farming Practices. Agriculture 2019, 9, 20. [Google Scholar] [CrossRef]
- Bruno, J.R.A.; Boddey, R.M.; Urquiaga, S. The success of BNF in soybean in Brazil. In Plant and Soil 660 FAO; Kluwer Academic Publishers: Dordrecht, The Netherlands, 2003; Volume 252, pp. 1–9. [Google Scholar]
- Córdova, S.C.; Castellano, M.J.; Dietzel, R.; Licht, M.A.; Togliatti, K.; Martínez-Feria, R.; Archontoulis, S.V. Soybean nitrogen fixation dynamics in Iowa, USA. Field Crops Res. 2019, 236, 165–176. [Google Scholar] [CrossRef]
- Pastor-Bueis, R.; Sánchez-Cañizares, C.; James, E.K.; González-Andrés, F. Formulation of a Highly Effective Inoculant for Common Bean Based on an Autochthonous Elite Strain of Rhizobium leguminosarum bv. phaseoli, and Genomic-Based Insights Into Its Agronomic Performance. Front. Microbial. 2019, 10, 2724. [Google Scholar] [CrossRef] [PubMed]
- Ulzen, J.; Abaidoo, R.C.; Mensah, N.E.; Masso, C.; AbdelGadir, A.H. Bradyrhizobium Inoculants Enhance Grain Yields of Soybean and Cowpea in Northern Ghana. Front. Plant Sci. 2016, 7, 1770. [Google Scholar]
- Ronner, E.; Franke, A.; Vanlauwe, B.; Dianda, M.; Edeh, E.; Ukem, B.; Giller, K. Understanding variability in soybean yield and response to P-fertilizer and rhizobium inoculants on farmers’ fields in northern Nigeria. Field Crops Res. 2016, 186, 133–145. [Google Scholar] [CrossRef]
- Thuita, M.; Vanlauwe, B.; Mutegi, E.; Masso, C. Reducing spatial variability of soybean response to rhizobia inoculants in farms of variable soil fertility in Siaya Country of west Kenya. Biol. Fert. Soils 2018, 261, 153–160. [Google Scholar]
- Mathenge, C.; Thuita, M.; Masso, C.; Gweyi-Onyango, J.; Vanlauwe, B. Variability of soybean response to rhizobia inoculant, vermicompost, and a legume-specific fertilizer blend in Siaya County of Kenya. Soil Till. Res. 2019, 194, 104–290. [Google Scholar] [CrossRef]
- Thuita, M.; Pypers, P.; Herrmann, L.; Okalebo, R.J.; Othieno, C.; Muema, E.; Lesueur, D. Commercial rhizobial inoculants significantly enhance growth and nitrogen fixation of a promiscuous soybean variety in Kenyan soils. Biol. Fertil. Soils 2012, 48, 87–96. [Google Scholar] [CrossRef]
- Balla, A.; Karanja, N.; Murwira, M.; Lwimbi, L.; Abaidoo, R.; Giller, K. Production and Use of Rhizobial Inoculants in Africa, 2011, 21. Available online: www.N2Africa.org (accessed on 6 July 2020).
- William, S.; Akiko, A. History of Soybeans and Soyfoods in Eastern Europe (Including All of Russia) (1783–2020): Extensively Annotated Bibliography and Sourcebook; Soyinfo Center: Lafayette, CA, USA, 2020; ISBN1 1948436175. ISBN2 9781948436175. [Google Scholar]
- Reis, V.M. Uso de Bactérias Fixadores de Nitrogênio omo Inoculante para Aplicação em Gramíneas. In Seropédica: Embrapa Agrobiologia; Embrapa Agrobiologia: Rodovia, Brasilia, 2007; Volume 232, p. 22. ISSN 1517-8498. [Google Scholar]
- El-Sayed, A.A.; Elenein, R.A.; Shalaby, E.E.; Shalan, M.A.; Said, M.A. Response of barley to biofertilizer with N and P application under newly reclaimed areas in Egypt. In Proceedings of the 3rd International Crop Science Congress (ICSC), Hamburg, Germany, 17–22 August 2000; pp. 17–22. [Google Scholar]
- Rose, M.T.; Phuong, T.L.; Nhan, D.K.; Cong, P.T.; Hien, N.T.; Kennedy, I.R. Up to 52% N fertilizer replaced by biofertilizer in lowland rice via farmer participatory research. Agron. Sustain. Dev. 2014, 34, 857–868. [Google Scholar] [CrossRef]
- Herridge, D.F.; Peoples, M.B.; Boddey, R.M. Global inputs of biological nitrogen fixation in agricultural systems. Plant Soil 2008, 311, 1–18. [Google Scholar] [CrossRef]
- Serna-Cock, L.; Arias-García, C.; Valencia Hernandez, L.J. Effect of biofertilization on the growth of potted sugarcane plants (Saccharum officinarum). Rev. Biol. Agroind. 2011, 9, 85–95. [Google Scholar]
- Antunes, J.E.L.; De Freitas, A.D.S.; Oliveira, L.M.S.; De Lyra, M.D.C.C.P.; Fonseca, M.A.C.; Santos, C.E.R.S.; Oliveira, J.P.; De Araújo, A.S.F.; Figueiredo, M.V.B. Sugarcane inoculated with endophytic diazotrophic bacteria: Effects on yield, biological nitrogen fixation and industrial characteristics. Anais da Academia Brasileira de Ciências 2019, 91, e20180990. [Google Scholar] [CrossRef]
- Lesueur, D.; Deaker, R.; Herrmann, L.; Bräu, L.; Jansa, J. Bioformulations: For Sustainable Agriculture; Kumar, A.N., Samina, M., Raffaella, B., Eds.; Springer India: New Delhi, India, 2016; pp. 71–92. [Google Scholar]
- Ouma, E.W.; Asango, A.M.; Maingi, J.; Njeru, E.M. Elucidating the potential of native rhizobial isolates to improve biological nitrogen fixation and growth of common bean and soybean in smallholder farming systems of Kenya. Int. J. Agron. 2016, 1–7. [Google Scholar] [CrossRef]
- Koskey, G.; Mburu, S.W.; Njeru, E.M.; Kimiti, J.M.; Omwoyo, O.; Maingi, J.M. Potential of Native Rhizobia in Enhancing Nitrogen Fixation and Yields of Climbing Beans (Phaseolus vulgaris L.) in Contrasting Environments of Eastern Kenya. Front. Plant Sci. 2017, 8, 443. [Google Scholar] [CrossRef] [PubMed]
- Douglas, A.E.; Werren, J.H. Holes in the hologenome: Why host–microbe symbioses are not holobionts. MBio 2016, 7, e02099-15. [Google Scholar] [CrossRef]
- Wang, Q.; Liu, J.; Zhu, H. Genetic and Molecular Mechanisms Underlying Symbiotic Specificity in Legume-Rhizobium Interactions. Front. Plant Sci. 2018, 9, 313. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, L.; Atieno, M.; Brau, L.; Lesueur, D. Microbial Quality of Commercial Inoculants to Increase BNF and Nutrient Use Efficiency. In Molecular Microbial Ecology of the Rhizosphere; De Bruijn, F.J., Ed.; Wiley: Blackwell, UK; Hoboken, NJ, USA, 2015. [Google Scholar]
- Singleton, P.W.; Boonkerd, N.; Carr, T.J.; Thompson, J.A. Technical and market constraints limiting legume inoculant use in Asia. In Extending Nitrogen Fixation Research to Farmers’ Fields; Rupela, O.P., Johansen, C., Herridge, D.F., Eds.; ICRISAT: Pantacheru, India, 1997; pp. 17–38. [Google Scholar]
- Callaghan, M.O. Microbial inoculation of seed for improved crop performance: Issues and opportunities. Appl. Microbiol. Biotecnol. 2016, 100, 5729–5746. [Google Scholar] [CrossRef] [PubMed]
- Herrmann, L.; Lesueur, D. Challenges of formulation and quality of biofertilizers for successful inoculation. Appl. Microbiol. Biotechnol. 2013, 97, 8859–8873. [Google Scholar] [CrossRef]
- Nguyen, T.H.; Phan, T.C.; Choudhury, A.T.M.A.; Rose, M.T.; Deaker, R.J.; Kennedy, I.R. BioGro: A Plant Growth-Promoting Biofertilizer Validated by 15 Years’ Research from Laboratory Selection to Rice Farmer’s Fields of the Mekong Delta. Agro-Environ. Sustain. 2017, 237–254. [Google Scholar] [CrossRef]
- Chien, Y.T.; Zinder, S.H. Cloning, functional organization, transcript studies, and phylogenetic analysis of the complete nitrogenase structural genes (nifHDK2) and associated genes in the archaeon Methanosarcina barkeri 227. J. Bacteriol. 1996, 178, 143–148. [Google Scholar] [CrossRef] [PubMed]
- Owen, D.; Williams, A.P.; Griffith, G.W.; Withers, P.J.A. Use of commercial bio-inoculants to increase agricultural production through improved phosphrous acquisition. Appl. Soil Ecol. 2015, 86, 41–54. [Google Scholar] [CrossRef]
- Trabelsi, D.; Mhamdi, R. Microbial Inoculants and Their Impact on Soil Microbial Communities: A Review. BioMed Res. Int. 2013, 1–11. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Srinivas, V.; Prakash, B.; Sathya, A.; Vijayabharathi, R. Plant growth-promoting traits of Pseudomonas geniculata isolated from chickpea nodules. 3 Biotech 2015, 5, 653–661. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Srinivas, V.; Samineni, S. Nitrogen fixation, plant growth and yield enhancements by diazotrophic growth-promoting bacteria in two cultivars of chickpea (Cicero arietinum L.). Biocatal. Agric. Biotechnol. 2017, 11, 116–123. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.V.; Srinivas, A.; Vemula, S.; Samineni, A. Rathore, Influence of diazotrophic bacteria on nodulation, nitrogen fixation, growth promotion and yield traits in five cultivars of chickpea. Biocatal. Agric. Biotechnol. 2018, 15, 35–42. [Google Scholar] [CrossRef]
- Yanni, Y.G.; Rizk, R.Y.; Corich, V.; Squartini, A.; Ninke, K.; Philip-Hollingsworth, S.; Orgambide, G.; de Bruijn, F.; Stoltzfus, J.; Buckley, D.; et al. Natural endophytic association between Rhizobium leguminosarum bv. trifolii and rice roots and assessment of its potential to promote rice growth. Plant Soil 1997, 194, 99–114. [Google Scholar] [CrossRef]
- Gebhardt, C.; Turner, G.L.; Gibson, A.H.; Dreyfus, B.L.; Bergersen, F.J. Nitrogen-fixing Growth in Continuous Culture of a Strain of Rhizobium sp. Isolated from Stem Nodules on Sesbania rostrata. Microbiology 1984, 130, 843–848. [Google Scholar] [CrossRef]
- Mia, M.A.B.; Shamsuddin, Z.H. Rhizobium as a crop enhancer and biofertilizer for increased cereal production. Afric. J. Biotechnol. 2010, 9, 6001–6009. [Google Scholar]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Varshney, R.K.; Gowda, C.L.L.; Krishnamurthy, L. Plant growth promoting rhizobia: Challenges and opportunities. 3 Biotech 2015, 5, 355. [Google Scholar] [CrossRef]
- Das, K.; Prasanna, R.; Saxena, A.K. Rhizobia: A potential biocontrol agent for soilborne fungal pathogens. Folia Microbiologica 2017, 62, 425–435. [Google Scholar] [CrossRef] [PubMed]
- Smith, S.E.; Read, D.J. Mycorrhizal Symbiosis; Academic Press: San Diego, CA, USA, 2008; p. 800. ISBN 9780123705266. [Google Scholar]
- Daniel, M.M.; Ezekiel, M.N.; Methuselah, M.N.; John, M.M. Arbuscular mycorrhizal fungi and Bradyrhizobium coinoculation enhances nitrogen fixation and growth of green grams (Vigna radiata L.) under water stress. J. Plant Nutr. 2020, 43, 1036–1047. [Google Scholar] [CrossRef]
- De Novais, C.B.; Sbrana, C.; da Conceição Jesus, E.; Rouws, L.F.M.; Giovannetti, M.; Avio, L.; Siqueira, J.O.; Saggin Júnior, O.J.; da Silva, E.M.R.; de Faria, S.M. Mycorrhizal networks facilitate the colonization of legume roots by a symbiotic nitrogen-fixing bacterium. Mycorrhiza 2020, 30, 389–396. [Google Scholar] [CrossRef]
- Parniske, M. Arbuscular mycorrhiza: The mother of plant root endosymbioses. Nat. Rev. Microbiol. 2008, 6, 763–775. [Google Scholar] [CrossRef]
- Dellagi, A.; Quillere, I.; Hirel, B. Beneficial soil-borne bacteria and fungi: A promising way to improve plant nitrogen acquisition. J. Exp. Bot. 2020, eraa112. [Google Scholar] [CrossRef]
- Agnolucci, M.; Avio, L.; Pepe, A.; Turrini, A.; Cristani, C.; Bonini, P.; Cirino, V.; Colosimo, F.; Ruzzi, M.; Giovannetti, M. Bacteria Associated With a Commercial Mycorrhizal Inoculum: Community Composition and Multifunctional Activity as Assessed by Illumina Sequencing and Culture-Dependent Tools. Front. Plant Sci. 2019, 9, 1956. [Google Scholar] [CrossRef]
- Goulding, K.W.T. Soil acidification and the importance of liming agricultural soils with particular reference to the United Kingdom. Soil Use Manag. 2016, 32, 390–399. [Google Scholar] [CrossRef]
- Geddes, B.A.; Paramasivan, P.; Joffrin, A.; Thompson, A.L.; Christensen, K.; Jorrin, B.; Brett, P.; Conway, S.J.; Oldroyd, G.E.D.; Poole, P.S. Engineering transkingdom signalling in plants to control gene expression in rhizosphere bacteria. Nat. Commn. 2019, 10, 3430. [Google Scholar] [CrossRef]
- Batista, M.B.; Dixon, R. Manipulating nitrogen regulation in diazotrophic bacteria for agronomic benefit. Biochem. Soc. Trans. 2019, 47, 603–614. [Google Scholar] [CrossRef] [PubMed]
- Beatty, P.H.; Good, A.G. Future prospects for cereals that fix nitrogen. Science 2011, 333, 416–417. [Google Scholar] [CrossRef]
- Griesmann, M.; Chang, Y.; Liu, X.; Song, Y.; Haberer, G.; Matthew, B.C.; Billault-Penneteau, B.; Lauressergues, D.; Keller, J.; Imanishi, L.; et al. Phylogenomic reveals multiple losses of nitrogen-fixing root nodule symbiosis. Science 2018, 361, eaat1743. [Google Scholar] [CrossRef] [PubMed]
- Rogers, C.; Oldroyd, G.E. Synthetic biology approaches to engineering the nitrogen symbiosis in cereals. J. Exp. Bot. 2014, 65, 1939–1946. [Google Scholar] [CrossRef]
- Burén, S.; López-Torrejón, G.; Rubio, L.M. Extreme bioengineering to meet the nitrogen challenge. Proc. Natl. Acad. Sci. USA 2018, 115, 12247. [Google Scholar] [CrossRef]
- Vicente, E.J.; Dean, D.R. Keeping the nitrogen-fixation dream alive. Proc. Natl. Acad. Sci. USA 2017, 114, 3009–3011. [Google Scholar] [CrossRef]
- Van Velzen, R.; Holmer, R.; Bu, F.; Rutten, L.; van Zeijl, A.; Liu, W.; Santuari, L.; Cao, Q.; Sharma, T.; Shen, D.; et al. Comparative genomics of the nonlegumeParasponiareveals insights into evolution of nitrogen-fixing rhizobium symbioses. Proc. Natl. Acad. Sci. USA 2018, 115, E4700–E4709. [Google Scholar] [CrossRef]
- López-Torrejón, G.; Jiménez-Vicente, E.; Buesa, J.M.; Hernandez, J.A.; Verma, H.K.; Rubio, L.M. Expression of a functional oxygen-labile nitrogenase component in the mitochondrial matrix of aerobically grown yeast. Nat. Commun. 2016, 7, 11426. [Google Scholar] [CrossRef]
- Wardhani, T.A.; Roswanjaya, Y.P.; Dupin, S.; Li, H.; Linders, S.; Hartog, M.; Geurts, R.; van Zeijl, A. Transforming, Genome Editing and Phenotyping the Nitrogen-fixing Tropical Cannabaceae Tree Parasponia andersonii. J. Vis. Exp. 2019, 150, e59971. [Google Scholar] [CrossRef]
- Mahmud, K.; Makaju, S.; Ibrahim, R.; Missaoui, A. Current Progress in Nitrogen Fixing Plants and Microbiome Research. Plants 2020, 9, 97. [Google Scholar] [CrossRef]
- Ivleva, N.B.; Groat, J.; Staub, J.M.; Stephens, M. Expression of Active Subunit of Nitrogenase via Integration into Plant Organelle Genome. PLoS ONE 2016, 11, e0160951. [Google Scholar] [CrossRef] [PubMed]
- Dixon, R.A.; Postgate, J.R. genetic transfer of nitrogen fixation from Klebsiella pneumoniae to Escherichia-coli. Nature 1972, 237, 102–103. [Google Scholar] [CrossRef]
- Temme, K.; Zhao, D.; Voigt, C.A. Refactoring the nitrogen fixation gene cluster from Klebsiella oxytoca. Proc. Natl Acad. Sci. USA 2012, 109, 7085–7090. [Google Scholar] [CrossRef]
- Wang, X.; Yang, J.-G.; Chen, L.; Wang, J.-L.; Cheng, Q.; Dixon, R.; Wang, Y.P. Using Synthetic Biology to Distinguish and Overcome Regulatory and Functional Barriers Related to Nitrogen Fixation. PLoS ONE 2013, 8, e68677. [Google Scholar] [CrossRef] [PubMed]
- Yang, J.; Xie, X.; Wang, X.; Dixon, R.; Wang, Y.P. Reconstruction and minimal gene requirements for the alternative iron-only nitrogenase in Escherichia coli. Proc. Natl. Acad. Sci. USA 2014, 111, 35. [Google Scholar] [CrossRef] [PubMed]
- Han, Y.; Lu, N.; Chen, Q.; Zhan, Y.; Liu, W.; Wei, L.; Zhu, B.; Lin, M.; Yang, Z.; Yan, Y. Interspecies Transfer and Regulation of Pseudomonas stutzeri A1501 Nitrogen Fixation Island in Escherichia coli. J. Microbiol. Biotechnol. 2015, 25, 1339–1348. [Google Scholar] [CrossRef]
- Burén, S.; Young, E.M.; Sweeny, E.A.; Lopez-Torrejón, G.; Veldhuizen, M.; Voigt, C.A.; Rubio, L.M. Formation of Nitrogenase NifDK Tetramers in the Mitochondria of Saccharomyces cerevisiae. ACS Synth. Biol. 2017, 6, 1043–1055. [Google Scholar] [CrossRef]
- Oldroyd, G.E.; Dixon, R. Biotechnological solutions to the nitrogen problem. Curr. Opin. Biotechnol. 2014, 26, 19–24. [Google Scholar] [CrossRef]
- Dwivedi, S.L.; Sahrawat, K.L.; Upadhyaya, H.D.; Mengoni, A.; Galardini, M.; Bazzicalupo, M.; Biondi, E.G.; Hungria, M.; Kaschuk, G.; Blair, M.W.; et al. Advances in Host Plant and Rhizobium Genomics to Enhance Symbiotic Nitrogen Fixation in Grain Legumes. Adv. Agron. 2015, 129, 1–116. [Google Scholar]
- Checcucci, A.; Azzarello, E.; Bazzicalupo, M.; Galardini, M.; Lagomarsino, A.; Mancuso, S.; Squartini, A.; Zanardo, M.; Mengoni, A. Mixed nodule infection in Sinorhizobium meliloti-Medicago sativa symbiosis suggest the presence of cheating behavior. Front. Plant Sci. 2016, 7, 835. [Google Scholar] [CrossRef]
- diCenzo, G.C.; Zamani, M.; Checcucci, A.; Fondi, M.; Griffitts, J.; Finan, T.; Mengoni, A. Multi-disciplinary approaches for studying rhizobium-legume symbioses. Can. J. Microbiol. 2019, 65, 1–33. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Mens, C.; Hastwell, A.H.; Zhang, M.; Su, H.; Jones, C.H.; Chu, X.; Gresshoff, P.M. Legume nodulation: The host controls the party. Plant Cell Environ. 2018, 42, 41–51. [Google Scholar] [CrossRef] [PubMed]
- Miransari, M.; Smith, D. Overcoming the stressful effects of salinity and acidity on soybean nodulation and yields using signal molecule genistein under field conditions. J. Plant Nutr. 2007, 30, 1967–1992. [Google Scholar] [CrossRef]
- Miransari, M.; Balakrishnan, P.; Smith, D.; Mackenzie, A.; Bahrami, H.; Malakouti, M.; Rejali, F. Overcoming the stressful effect of low pH on soybean root hair curling using lipochitooligosacharides. Commun. Soil Sci. Plant Anal. 2006, 37, 1103–1110. [Google Scholar] [CrossRef]
- Suliema, S.; Tran, L.S.P. Legume Nitrogen Fixation in a Changing Environment—Achievements and Challenges; Sulieman, S., Phan Tran, L.-S., Eds.; Springer: Cham, Switzerland, 2015; p. 133. [Google Scholar] [CrossRef]
- Pedrosa, F.O. Genome of Herbaspirillum seropedicae strain SmR1, a specialized diazotrophic endophyte of tropical grasses. PLoS Genet. 2011, 7, e1002064. [Google Scholar] [CrossRef]
- Yonebayashi, K.; Katsumi, N.; Nishi, T.; Okazaki, M. Activation of Nitrogen-Fixing Endophytes Is Associated with the Tuber Growth of Sweet Potato. Mass Spectrum. 2014, 3, A0032. [Google Scholar] [CrossRef]
- Farrar, K.; Bryant, D.; Cope-delby, N. Understanding and engineering beneficial plant-microbe interactions: Plant growth promotion in energy crops. Plant Biotechnol. J. 2014, 12, 1193–1206. [Google Scholar] [CrossRef]
- Straub, D.; Rothballer, M.; Hartmann, A.; Ludewig, U. The genome of the endophytic bacterium H. frisingense GSF30(T) identifies diverse strategies in the Herbaspirillum genus to interact with plants. Front. Microbiol. 2013, 4, 168. [Google Scholar] [CrossRef]
- Adesemoye, A.O.; Torbert, H.A.; Kloepper, J.W. Plant growth-promoting rhizobacteria allow reduced application rates of chemical fertilizers. Microb. Ecol. 2009, 5, 921–929. [Google Scholar] [CrossRef]
- Burghardt, L.T. Evolving together, evolving apart: Measuring the fitness of rhizobial bacteria in and out of symbiosis with leguminous plants. New Phytol. 2019. [Google Scholar] [CrossRef]
- Soumare, A.; Diop, T.; Manga, A.; Ndoye, I. Role of arbuscular mycorrhizal fungi and nitrogen fixing bacteria on legume growth under various environmental stresses. Int. J. Biosci. 2015, 7, 31–46. [Google Scholar] [CrossRef]
- FAO/IIASA/ISRIC/ISS-CAS/JRC, Harmonized World Soil Database (version 1.1). FAO. 2009, Rome, Italy and IIASA, Laxenburg, Austria. Available online: http://www.fao.org/3/a-aq361e.pdf (accessed on 13 July 2020).
- Liu, J.; You, L.; Amini, M.; Obersteiner, M.; Herrero, M.; Zehnder, A.J.B.; Yang, H. A high-resolution assessment on global nitrogen flows in cropland PNAS. Proc. Natl. Acad. Sci. USA 2010, 107, 8035–8040. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Date | Events | References |
|---|---|---|
| 1836 | Identification of nitrogen as a nutrient for plants | [75] |
| 1886 | Hellriegel and Wilfarth demonstrated the ability of legumes to convert N2 | [20] |
| 1888 | First rhizobia were isolated from nodules | [64] |
| 1893 | Isolation of Clostridium pasteurianum (Free-living N fixers) | |
| 1895 | First commercial inoculant (Nitragen) | |
| 1901 | Isolation Azotobacter | [75] |
| 1913 | Carl Bosch performed Haber’s ammonia synthesis on an industrial scale | |
| 1946 | Second commercial inoculant (Azotogen) | [76] |
| 1953 | Identification of two nitrogen fixing bacteria: Beijerinckia fluminensis and Azotobacter paspali | [67,68] |
| 1969 | Positive results for 15N2 uptake by cyanobacteria | [71] |
| 1972 | Isolation of Enterobacter cloacae from corn roots | [77] |
| 1975 | Isolation of Spirillum sp. and demonstration of their nitrogenase activity | [78] |
| 1984 | Nitrogen fixation in Methanogens (archaea) | [74] |
| 2011 | The European Nitrogen Assessment provides the first integrated and comprehensive look at N use in Europe | [79] |
| 2012 | Database of all nifH sequences available in the Genbank nucleotide database | [80] |
| Name of Manufactured Products and Producer (in Italic) | Strain | Formulation | Crops Suited | Benefits According to the Authors | References |
|---|---|---|---|---|---|
| BioGro Nguyen Thanh Hien in Hanoi University (Vietnam) | Pseudomonas fluorescens Bacillus subtilis Bacillus amyloliquefaciens Candida tropicalis | Solid in peat | Rice (Oryza sativa) | Improve rice yield | [118] |
| Biofix MEA company limited (Kenya) | Rhizobium | solid | -Soya bean (Glycine max) -Common bean (Phaseolus vulgaris L) -Alfalfa (Medicago sativa) | Cheaper than chemical nitrogen Lighter to transport, requires less labor effective for many crops | [109] |
| Bio N Nutri-Tech Solutions (Australia) | Azotobacter spp. | liquid | Horticulture | Access free atmospheric nitrogen. Increase yield and quality. Reduce soil erosion. Improve water retention. Enhance germination. Promote root growth. Phosphate release | [114] |
| Microbin and Azottein Egyptian Ministry of Agriculture | Klebsiella, Bacillus, Azotobacter Azospirillum | Carrier material | Barley cultivar Giza | Increased the different plant characteristic increases in grain yield reached approximately 24.8 and 27.2% | [92,104] |
| Legumefix Legume Technology (UK) | Bradyrhizobium japonicum | Sterile peat inoculant | Soybean and cowpea | grain yield (12–19%) relative to the control | [96] |
| Leguspirflo SoyGro (South Africa) | Azospirillum brasilense | Liquid | soybean | Inefficient | [97] |
| TerraMax’s Micro AZ product TerraMax (Minnesota, USA) | Azospirillum brasilense A. lipoferum. | Liquid | Wheat, Corn and Grain Sorghum | Improve root structure and stimulate root growth Provide biological nitrogen fixation Increases yields Stimulates rooting Increases yields | [119,120] |
| Nitrofix P Agro-Input Suppliers Limited (AISL) (Malawi) | Bradyrhizobium japonicum and Bradyrhizobium elkanii | Dry- inoculum based on gamma-sterilized peat | Soybeans | Promotes an increase in the yield by an average of 14.3–20.3% Reduced the nitrogen requirement | [79] |
| Vault LVL BASF (Badische Anilin- & Soda-Fabrik) Germany | B. japonicum + Bacillus subtilis | Liquid | Soybeans | Biomass yield improved | [98,100] |
| Current Progress in Nitrogenase Transfer | References |
|---|---|
| Nif gene cluster transfer from nitrogen-fixing bacteria Klebsiella pneumoniae into Escherichia coli | [150] |
| Refactoring the nitrogen fixation gene to a simple cluster easy to access, engineering, and transferability. | [151] |
| Nif cluster gene transfer from nitrogen-fixing bacteria Paenibacillus sp. into Escherichia coli | [152] |
| Conception of an artificial FeFe nitrogenase system in Escherichia coli | [153] |
| Tranfer and expression of Pseudomonas stutzeri A1501 Nitrogen Fixation Island in Escherichia coli | [154] |
| Tranfer nitrogenase components (nifH) in Saccharomyces cerevisiae mitochondria (as model of eukaryotic cell) | [155] |
| Transgenic production of nitrogenase and expressing nitrogenase genes in plant plastids (tobacco as model) | [149] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soumare, A.; Diedhiou, A.G.; Thuita, M.; Hafidi, M.; Ouhdouch, Y.; Gopalakrishnan, S.; Kouisni, L. Exploiting Biological Nitrogen Fixation: A Route Towards a Sustainable Agriculture. Plants 2020, 9, 1011. https://doi.org/10.3390/plants9081011
Soumare A, Diedhiou AG, Thuita M, Hafidi M, Ouhdouch Y, Gopalakrishnan S, Kouisni L. Exploiting Biological Nitrogen Fixation: A Route Towards a Sustainable Agriculture. Plants. 2020; 9(8):1011. https://doi.org/10.3390/plants9081011
Chicago/Turabian StyleSoumare, Abdoulaye, Abdala G. Diedhiou, Moses Thuita, Mohamed Hafidi, Yedir Ouhdouch, Subramaniam Gopalakrishnan, and Lamfeddal Kouisni. 2020. "Exploiting Biological Nitrogen Fixation: A Route Towards a Sustainable Agriculture" Plants 9, no. 8: 1011. https://doi.org/10.3390/plants9081011
APA StyleSoumare, A., Diedhiou, A. G., Thuita, M., Hafidi, M., Ouhdouch, Y., Gopalakrishnan, S., & Kouisni, L. (2020). Exploiting Biological Nitrogen Fixation: A Route Towards a Sustainable Agriculture. Plants, 9(8), 1011. https://doi.org/10.3390/plants9081011