Adventitious Shoot Regeneration from In Vitro Leaf Explants of the Peach Rootstock Hansen 536



Abstract
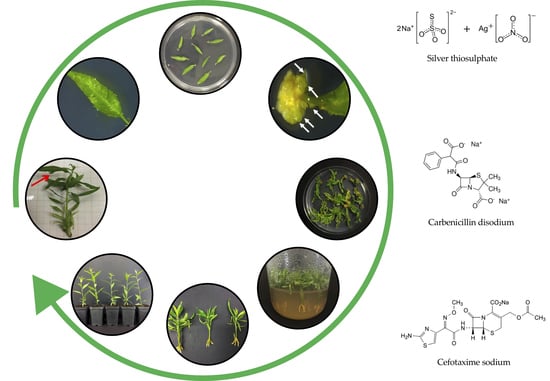
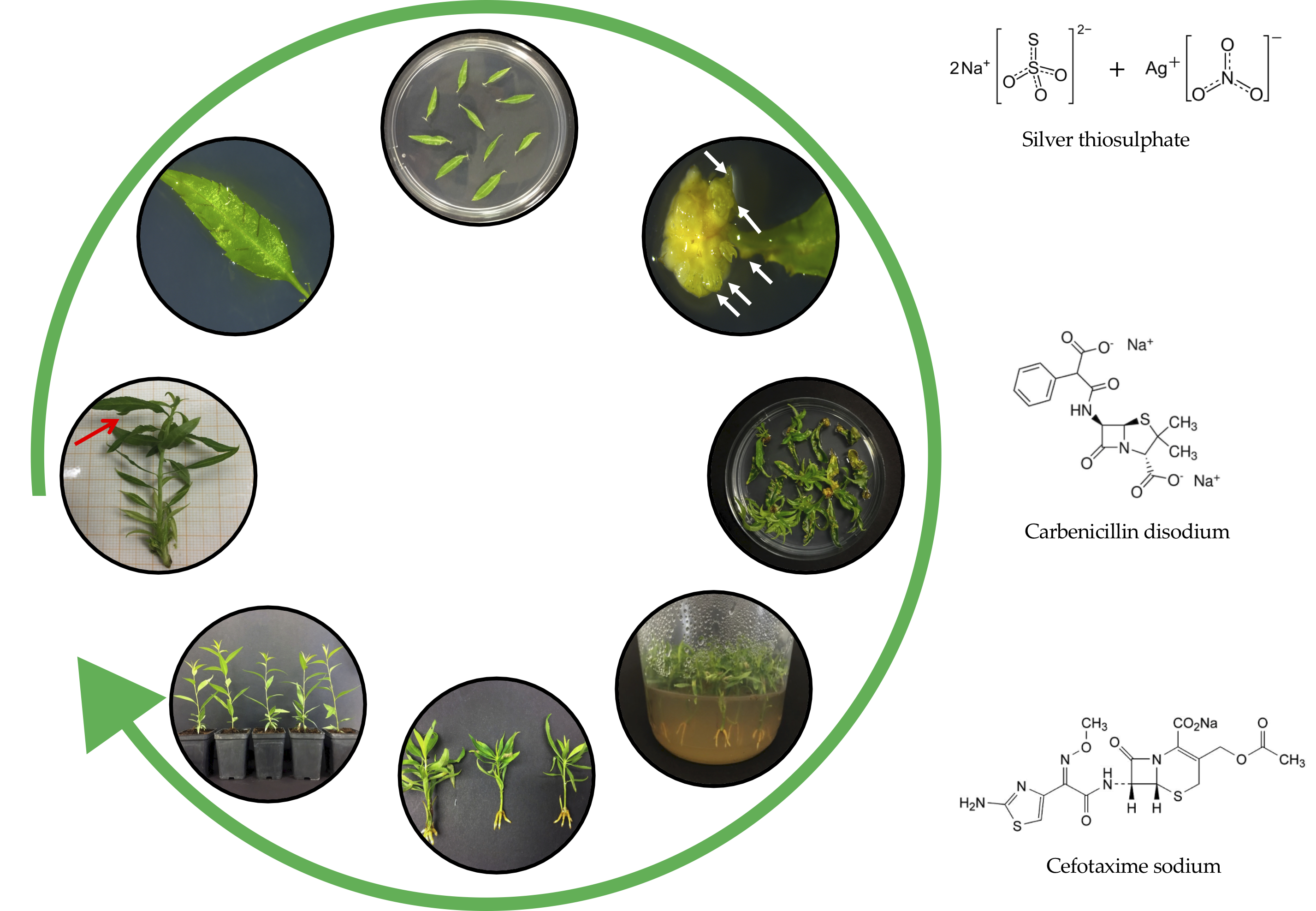
1. Introduction
2. Results
2.1. Influence of Different Combinations and Concentrations of PGRs on Regeneration Efficiency from Leaf Explants
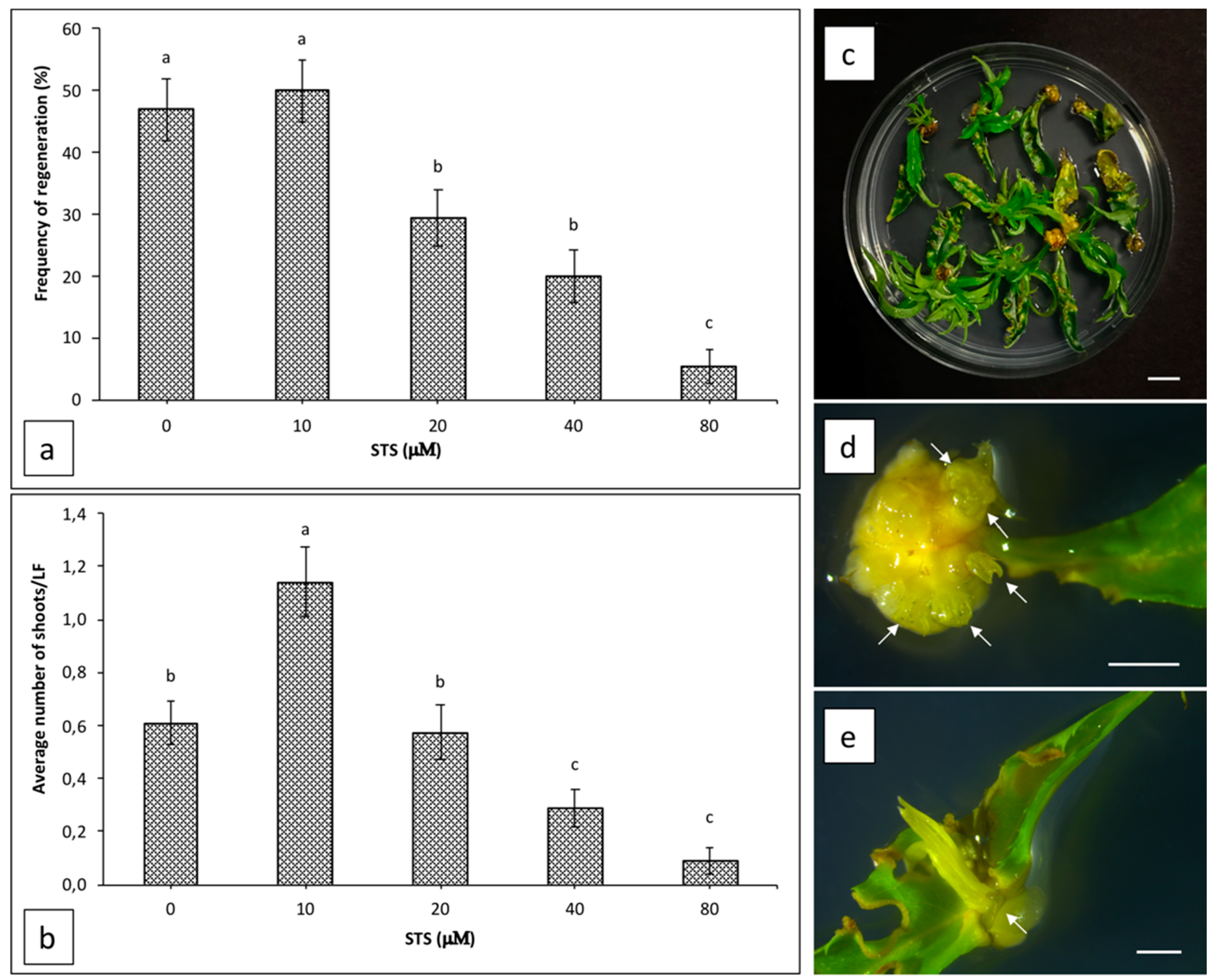
2.2. Influence of Silver Thiosulphate on Regeneration Efficiency from Leaf Explants
2.3. Influence of Antibiotics on Regeneration Efficiency from Leaf Explants
2.4. In Vitro Rooting and Acclimatization of Adventitious Shoots
3. Discussion
4. Materials and Methods
4.1. Establishment of In Vitro Shoots
4.2. General Approach for Adventitious Shoot Regeneration
4.3. Effect of Different Combinations and Concentrations of PGRs
4.4. Effect of Silver Thiosulphate
4.5. Effect of Antibiotics
4.6. Shoots Elongation, Rooting and Acclimatization
4.7. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Xu, S.; Lai, E.; Zhao, L.; Cai, Y.; Ogutu, C.; Cherono, S.; Han, Y.; Zheng, B. Development of a fast and efficient root transgenic system for functional genomics and genetic engineering in peach. Sci. Rep. 2020, 10, 2836. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Jiménez, M.; Carrillo-Navarro, A.; Cos-Terrer, J. Regeneration of peach (Prunus persica L. Batsch) cultivars and Prunus persica × Prunus dulcis rootstocks via organogenesis. Plant Cell Tissue Organ Cult. 2012, 108, 55–62. [Google Scholar] [CrossRef]
- Sabbadini, S.; Ricci, A.; Limera, C.; Baldoni, D.; Capriotti, L.; Mezzetti, B. Factors Affecting the Regeneration, via Organogenesis, and the Selection of Transgenic Calli in the Peach Rootstock Hansen 536 (Prunus persica × Prunus amygdalus) to Express an RNAi Construct against PPV Virus. Plants 2019, 8, 178. [Google Scholar] [CrossRef] [PubMed]
- Hammerschlag, F.A.; Bauchan, G.; Scorza, R. Regeneration of peach plants from callus derived from immature embryos. Theor. Appl. Genet. 1985, 70, 248–251. [Google Scholar] [CrossRef] [PubMed]
- Bhansali, R.R.; Driver, J.A.; Durzan, D.J. Rapid multiplication of adventitious somatic embryos in peach and nectarine by secondary embryogenesis. Plant Cell Rep. 1990, 9, 280–284. [Google Scholar] [CrossRef] [PubMed]
- Scorza, R.; Morgens, P.H.; Cordts, J.M.; Mante, S.; Callahan, A.M. Agrobacterium-mediated transformation of peach (Prunus persica L. batsch) leaf segments, immature embryos, and long-term embryogenic callus. Vitr. Cell. Dev. Biol. 1990, 26, 829–834. [Google Scholar]
- Smigocki, A.C.; Hammerschlag, F.A. Regeneration of Plants from Peach Embryo Cells Infected with a Shooty Mutant Strain of Agrobacterium. J. Am. Soc. Hortic. Sci. 1991, 116, 1092–1097. [Google Scholar] [CrossRef]
- Pérez-Clemente, R.M.; Pérez-Sanjuán, A.; García-Férriz, L.; Beltrán, J.-P.; Cañas, L.A. Transgenic peach plants (Prunus persica L.) produced by genetic transformation of embryo sections using the green fluorescent protein (GFP) as an in vivo marker. Mol. Breed. 2005, 14, 419–427. [Google Scholar] [CrossRef]
- Sabbadini, S.; Pandolfini, T.; Girolomini, L.; Molesini, B.; Navacchi, O. Peach (Prunus persica L.) BT—Agrobacterium Protocols: Volume 2; Wang, K., Ed.; Springer: New York, NY, USA, 2015; pp. 205–215. ISBN 978-1-4939-1658-0. [Google Scholar]
- Gentile, A.; Monticelli, S.; Damiano, C. Adventitious shoot regeneration in peach [Prunus persica (L.) Batsch]. Plant Cell Rep. 2002, 20, 1011–1016. [Google Scholar] [CrossRef]
- Zhou, H.; Li, M.; Zhao, X.; Fan, X.; Guo, A. Plant regeneration from in vitro leaves of the peach rootstock ‘Nemaguard’ (Prunus persica × P. davidiana). Plant Cell Tissue Organ Cult. 2010, 101, 79–87. [Google Scholar] [CrossRef]
- San, B.; Li, Z.; Hu, Q.; Reighard, G.L.; Luo, H. Adventitious shoot regeneration from in vitro cultured leaf explants of peach rootstock Guardian® is significantly enhanced by silver thiosulfate. Plant Cell Tissue Organ Cult. 2015, 120, 757–765. [Google Scholar] [CrossRef]
- Zong, X.; Denler, B.J.; Danial, G.H.; Chang, Y.; Song, G. Adventitious Shoot Regeneration and Agrobacterium tumefaciens-mediated Transient Transformation of Almond × Peach Hybrid Rootstock ‘Hansen 536’. Hortscience 2019, 54, 936–940. [Google Scholar] [CrossRef]
- Biddington, N.L. The influence of ethylene in plant tissue culture. Plant Growth Regul. 1992, 11, 173–187. [Google Scholar] [CrossRef]
- Veen, H.; van de Geijn, S.C. Mobility and ionic form of silver as related to longevity of cut carnations. Planta 1978, 140, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Burgos, L.; Alburquerque, N. Ethylene inhibitors and low kanamycin concentrations improve adventitious regeneration from apricot leaves. Plant Cell Rep. 2003, 21, 1167–1174. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.; Pijut, P.M. Plant regeneration from in vitro leaves of mature black cherry (Prunus serotina). Plant Cell Tissue Organ Cult. 2008, 94, 113–123. [Google Scholar] [CrossRef]
- Petri, C.; Scorza, R. Factors affecting adventitious regeneration from in vitro leaf explants of ‘Improved French’ plum, the most important dried plum cultivar in the USA. Ann. Appl. Biol. 2010, 156, 79–89. [Google Scholar] [CrossRef]
- Byrne, D.H.; Raseira, M.B.; Bassi, D.; Piagnani, M.C.; Gasic, K.; Reighard, G.L.; Moreno, M.A.; Pérez, S. Peach BT—Fruit Breeding; Badenes, M.L., Byrne, D.H., Eds.; Springer: Boston, MA, USA, 2012; pp. 505–569. ISBN 978-1-4419-0763-9. [Google Scholar]
- Connell, J.H.; Buchner, R.; Duncan, R.; Edstrom, J.; Micke, W.; Verdegaal, P.; Viveros, M. Evaluation of Potential Rootstocks for California Almonds. In Proceedings of the Acta Horticulturae; International Society for Horticultural Science (ISHS): Leuven, Belgium, 2002; pp. 45–52. [Google Scholar]
- Pinochet, J. ‘Greenpac’, a New Peach Hybrid Rootstock Adapted to Mediterranean Conditions. HortScience 2009, 44, 1456–1457. [Google Scholar] [CrossRef]
- Kester, D.E.; Asay, R.N. “Hansen 2168” and “Hansen 536”: Two new Prunus rootstock clones. HortScience 1986, 21, 331–332. [Google Scholar]
- Felipe, A.J.; Gómez-Aparisi, J.; Socías, R.; Carrera, M. The Almond X Peach Hybrid Rootstocks Breeding Program At Zaragoza (Spain). In Proceedings of the Acta Horticulturae; International Society for Horticultural Science (ISHS): Leuven, Belgium, 1997; pp. 259–262. [Google Scholar]
- Declerck, V.; Korban, S.S. Influence of growth regulators and carbon sources on callus induction, growth and morphogenesis from leaf tissues of peach (Prunus persica L. Batsch). J. Hortic. Sci. 1996, 71, 49–55. [Google Scholar] [CrossRef]
- Escalettes, V.; Dosba, F. In vitro adventitious shoot regeneration from leaves of Prunus spp. Plant Sci. 1993, 90, 201–209. [Google Scholar] [CrossRef]
- Miguel, C.M.; Druart, P.; Oliveira, M.M. Shoot regeneration from adventitious buds induced on juvenile and adult almond (Prunus dulcis mill.) explants. Vitr. Cell. Dev. Biol. Plant 1996, 32, 148–153. [Google Scholar] [CrossRef]
- Pérez-Tornero, O.; Egea, J.; Vanoostende, A.; Burgos, L. Assessment of factors affecting adventitious shoot regeneration from in vitro cultured leaves of apricot. Plant Sci. 2000, 158, 61–70. [Google Scholar] [CrossRef]
- Mazumdar, P.; Basu, A.; Paul, A.; Mahanta, C.; Sahoo, L. Age and orientation of the cotyledonary leaf explants determine the efficiency of de novo plant regeneration and Agrobacterium tumefaciens-mediated transformation in Jatropha curcas L. S. Afr. J. Bot. 2010, 76, 337–344. [Google Scholar] [CrossRef]
- Cardoso, J.C.; Habermann, G. Adventitious shoot induction from leaf segments in Anthurium andreanum is affected by age of explant, leaf orientation and plant growth regulator. Hortic. Environ. Biotechnol. 2014, 55, 56–62. [Google Scholar] [CrossRef]
- James, D.J.; Passey, A.J.; Rugini, E. Factors Affecting High Frequency Plant Regeneration from Apple Leaf Tissues Cultured in vitro. J. Plant Physiol. 1988, 132, 148–154. [Google Scholar] [CrossRef]
- Bhatia, P.; Ashwath, N.; Midmore, D.J. Effects of genotype, explant orientation, and wounding on shoot regeneration in tomato. Vitr. Cell. Dev. Biol. Plant 2005, 41, 457–464. [Google Scholar] [CrossRef]
- Meng, R.; Chen, T.H.H.; Finn, C.E.; Li, Y. Improving in vitro plant regeneration from leaf and petiole explants of “Marion” blackberry. HortScience 2004, 39, 316–320. [Google Scholar] [CrossRef]
- Sarwar, M.; Skirvin, R.M. Effect of thidiazuron and 6-benzylaminopurine on adventitious shoot regeneration from leaves of three strains of “McIntosh” apple (Malus X domestica Borkh.) in vitro. Sci. Hortic. (Amsterdam) 1997, 68, 95–100. [Google Scholar] [CrossRef]
- Chi, G.-L.; Barfield, D.G.; Sim, G.-E.; Pua, E.-C. Effect of AgNO3 and aminoethoxyvinylglycine on in vitro shoot and root organogenesis from seedling explants of recalcitrant Brassica genotypes. Plant Cell Rep. 1990, 9, 195–198. [Google Scholar] [CrossRef]
- Kumar, V.; Parvatam, G.; Ravishankar, G.A. AgNO3: A potential regulator of ethylene activity and plant growth modulator. Electron. J. Biotechnol. 2009, 12, 8–9. [Google Scholar] [CrossRef]
- Möllers, C.; Zhangm, S.; Wenzel, G. The Influence of Silver Thiosulfate on Potato Protoplast Cultures. Plant Breed. 1992, 108, 12–18. [Google Scholar] [CrossRef]
- Trujillo-Moya, C.; Gisbert, C. The influence of ethylene and ethylene modulators on shoot organogenesis in tomato. Plant Cell Tissue Organ Cult. 2012, 111, 41–48. [Google Scholar] [CrossRef]
- Dimasi-Theriou, K.; Economou, A.S. Ethylene enhances shoot formation in cultures of the peach rootstock GF-677 (Prunus persica × P. amygdalus). Plant Cell Rep. 1995, 15, 87–90. [Google Scholar] [CrossRef]
- Nakano, M.; Mii, M. Antibiotics Stimulate Somatic Embryogenesis without Plant Growth in Several Dianthus Cultivars. J. Plant Physiol. 1993, 141, 721–725. [Google Scholar] [CrossRef]
- Dai, W.; Castillo, C. Factors Affecting Plant Regeneration from Leaf Tissues of Buddleia Species. HortScience 2007, 42, 1670–1673. [Google Scholar] [CrossRef]
- Yu, Y.; Wei, Z.-M. Influences of cefotaxime and carbenicillin on plant regeneration from wheat mature embryos. Biol. Plant 2008, 52, 553–556. [Google Scholar] [CrossRef]
- Bosela, M.J. Effects of β-lactam antibiotics, auxins, and cytokinins on shoot regeneration from callus cultures of two hybrid aspens, Populustremuloides × P. tremula and P. xcanescens × P. gradidentata. Plant Cell Tissue Organ Cult. 2009, 98, 249–261. [Google Scholar] [CrossRef]
- Silvestri, C.; Cristofori, V.; Ceccarelli, M.; Caceres, M.E.; Escribà-Lacuesta, J.; Rugini, E. Adventitious shoot organogenesis from leaf and petiole explants of European hazelnut. Plant Cell Tissue Organ Cult. 2016, 126, 59–65. [Google Scholar] [CrossRef]
- Orlikowska, T.K.; Cranston, H.J.; Dyer, W.E. Factors influencing Agrobacterium tumefaciens-mediated transformation and regeneration of the safflower cultivar ‘centennial’. Plant Cell Tissue Organ Cult. 1995, 40, 85–91. [Google Scholar] [CrossRef]
- Hammerschlag, F.A.; Zimmerman, R.H.; Yadava, U.L.; Hunsucker, S.; Gercheva, P. Effect of Antibiotics and Exposure to an Acidified Medium on the Elimination of Agrobacterium tumefaciens from Apple Leaf Explants and on Shoot Regeneration. J. Am. Soc. Hortic. Sci. JASHS 1997, 122, 758–763. [Google Scholar] [CrossRef]
- Yu, T.A.; Yeh, S.D.; Yang, J.S. Effects of carbenicillin and cefotaxime on callus growth and somatic embryogenesis from adventitious roots of papaya. Bot. Bull. Acad. Sin. 2001, 42, 281–286. [Google Scholar]
- Shehata, A.M.; Wannarat, W.; Skirvin, R.M.; Norton, M.A. The dual role of carbenicillin in shoot regeneration and somatic embryogenesis of horseradish (Armoracia rusticana) in vitro. Plant Cell Tissue Organ Cult. 2010, 102, 397–402. [Google Scholar] [CrossRef]
- Holford, P.; Newbury, H.J. The effects of antibiotics and their breakdown products on the in vitro growth of Antirrhinum majus. Plant Cell Rep. 1992, 11, 93–96. [Google Scholar] [CrossRef] [PubMed]
- Nauerby, B.; Billing, K.; Wyndaele, R. Influence of the antibiotic timentin on plant regeneration compared to carbenicillin and cefotaxime in concentrations suitable for elimination of Agrobacterium tumefaciens. Plant Sci. 1997, 123, 169–177. [Google Scholar] [CrossRef]
- Borrelli, G.M.; Di Fonzo, N.; Lupotto, E. Effect of Cefotaxime on Callus Culture and Plant Regeneration in Durum Wheat. J. Plant Physiol. 1992, 140, 372–374. [Google Scholar] [CrossRef]
- Yepes, L.M.; Aldwinekle, H.S. Factors that effect leaf regeneration efficiency in apple, and effect of antibiotics in morphogenesis. Plant Cell Tissue Organ Cult. 1994, 37, 257–269. [Google Scholar]
- Sarma, K.S.; Evans, N.E.; Selby, C. Effect of carbenicillin and cefotaxime on somatic embryogenesis of Sitka spruce (Picea sitchensis (Bong.) Carr.). J. Exp. Bot. 1995, 46, 1779–1781. [Google Scholar] [CrossRef]
- Grzebelus, E.; Skop, L. Effect of β-lactam antibiotics on plant regeneration in carrot protoplast cultures. Vitr. Cell. Dev. Biol. Plant 2014, 50, 568–575. [Google Scholar] [CrossRef]
- Sujana, P.; Naidu, C.V. Influence of Bavistin, Cefotaxime, Kanamycin and Silver Thiosulphate on Plant Regeneration of Mentha piperita (L.)—An Important Multipurpose Medicinal Plant. J. Phytol. 2011, 3, 36–40. [Google Scholar]
- Tambarussi, E.V.; Rogalski, M.; Nogueira, F.T.S.; Brondani, G.E.; De Martin, V.F.; Carrer, H. Influence of antibiotics on indirect organogenesis of Teak. Ann. For. Res. 2015, 58, 177–183. [Google Scholar] [CrossRef]
- Rugini, E.; Caricato, G. Somatic embryogenesis and plant recovery from mature tissues of olive cultivars (Olea europaea L.) “canino” and “moraiolo”. Plant Cell Rep. 1995, 14, 257–260. [Google Scholar] [CrossRef] [PubMed]
- Lloyd, G.; McCown, B. Commercially-feasible micropropagation of mountain laurel, Kalmia latifolia, by use of shoot-tip culture. Commer. Feasible Micropropag. Mt. Laurel Kalmia Latifolia Use Shoot Tip Cult. 1980, 30, 421–427. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Regeneration Media | Frequency of Regeneration (%) ± SE * | Average Number of Regenerating Shoots/LF ± SE |
|---|---|---|
| WPM 1 | 0 (g) | 0 (e) |
| WPM 2 | 0 (g) | 0 (e) |
| WPM 3 | 8.7 ± 2.3 (fg) | 0.12 ± 0.03 (de) |
| WPM 4 | 9.3 ± 2.38 (fg) | 0.11 ± 0.04 (de) |
| WPM 5 | 6.7 ± 2.04 (fg) | 0.07 ± 0.02 (de) |
| WPM 6 | 13.3 ± 2.78 (defg) | 0.15 ± 0.03 (cde) |
| WPM 7 | 3.3 ± 1.47 (g) | 0.03 ± 0.0.1 (e) |
| WPM 8 | 2 ± 1.15 (g) | 0.02 ± 0.01 (e) |
| WPM 9 | 26.7 ± 3.62 (bcd) | 0.41 ± 0.06 (b) |
| WPM 10 | 24.7 ± 3.53 (bcde) | 0.39 ± 0.06 (b) |
| WPM 11 | 53 ± 4.02 (a) | 0.77 ± 0.08 (a) |
| WPM 12 | 29.3 ± 3.73 (bc) | 0.47 ± 0.07 (b) |
| WPM 13 | 32 ± 3.82 (bc) | 0.39 ± 0.05 (b) |
| WPM 14 | 37 ± 3.84 (b) | 0.48 ± 0.06 (b) |
| WPM 15 | 28 ± 3.88 (bc) | 0.48 ± 0.06 (b) |
| WPM 16 | 20 ± 3.59 (cdef) | 0.3 ± 0.06 (bcd) |
| WPM 17 | 24 ± 3.49 (bcde) | 0.36 ± 0.07 (bc) |
| WPM 18 | 12 ± 3.19 (efg) | 0.16 ± 0.05 (de) |
| WPM 19 | 2 ± 2.72 (g) | 0.02 ± 0.04 (e) |
| WPM 20 | 6 ± 2.04 (fg) | 0.06 ± 0.03 (de) |
| Antibiotic(s) w | μM y | Frequency of Regeneration (%) ± SE z | Average Number of Regenerating Shoots/LF ± SE |
|---|---|---|---|
| None | 0 | 49 ± 5.02 (a) | 0.61 ± 0.08 (b) |
| Carbenicillin | 475 | 19.3 ± 3.68 (b) | 0.4 ± 0.07 (bc) |
| Cefotaxime | 420 | 21.4 ± 3.86 (b) | 0.33 ± 0.06 (bc) |
| Timentin x | 514 | 9.3 ± 1.41 (b) | 0.17 ± 0.01 (c) |
| Carbenicillin/Cefotaxime | 238/210 | 43 ± 4.94 (a) | 0.91 ± 0.13 (a) |
| Timentin/Cefotaxime | 171/210 | 21.3 ± 3.94 (b) | 0.39 ± 0.07 (bc) |
| Plant Growth Regulators (μM) | ||||
|---|---|---|---|---|
| KIN | BAP | TDZ | NAA | |
| WPM 1 | - | - | - | - |
| WPM 2 | - | - | - | 0.25 |
| WPM 3 | 14 | - | - | - |
| WPM 4 | 16.3 | - | - | - |
| WPM 5 | 19 | - | - | - |
| WPM 6 | 14 | - | - | 0.25 |
| WPM 7 | 16.3 | - | - | 0.25 |
| WPM 8 | 19 | - | - | 0.25 |
| WPM 9 | - | 11 | - | - |
| WPM 10 | - | 13.3 | - | - |
| WPM 11 | - | 15.5 | - | - |
| WPM 12 WPM 13 WPM 14 WPM 15 WPM 16 WPM 17 WPM 18 | - - - - - - - | 11 13.3 15.5 - - - - | - - - 2.25 4.5 9 2.25 | 0.25 0.25 0.25 - - - 0.25 |
| WPM 19 | - | - | 4.5 | 0.25 |
| WPM 20 | - | - | 9 | 0.25 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ricci, A.; Capriotti, L.; Mezzetti, B.; Navacchi, O.; Sabbadini, S. Adventitious Shoot Regeneration from In Vitro Leaf Explants of the Peach Rootstock Hansen 536. Plants 2020, 9, 755. https://doi.org/10.3390/plants9060755
Ricci A, Capriotti L, Mezzetti B, Navacchi O, Sabbadini S. Adventitious Shoot Regeneration from In Vitro Leaf Explants of the Peach Rootstock Hansen 536. Plants. 2020; 9(6):755. https://doi.org/10.3390/plants9060755
Chicago/Turabian StyleRicci, Angela, Luca Capriotti, Bruno Mezzetti, Oriano Navacchi, and Silvia Sabbadini. 2020. "Adventitious Shoot Regeneration from In Vitro Leaf Explants of the Peach Rootstock Hansen 536" Plants 9, no. 6: 755. https://doi.org/10.3390/plants9060755
APA StyleRicci, A., Capriotti, L., Mezzetti, B., Navacchi, O., & Sabbadini, S. (2020). Adventitious Shoot Regeneration from In Vitro Leaf Explants of the Peach Rootstock Hansen 536. Plants, 9(6), 755. https://doi.org/10.3390/plants9060755