Molecular Properties and New Potentials of Plant Nepenthesins


Abstract
1. Introduction
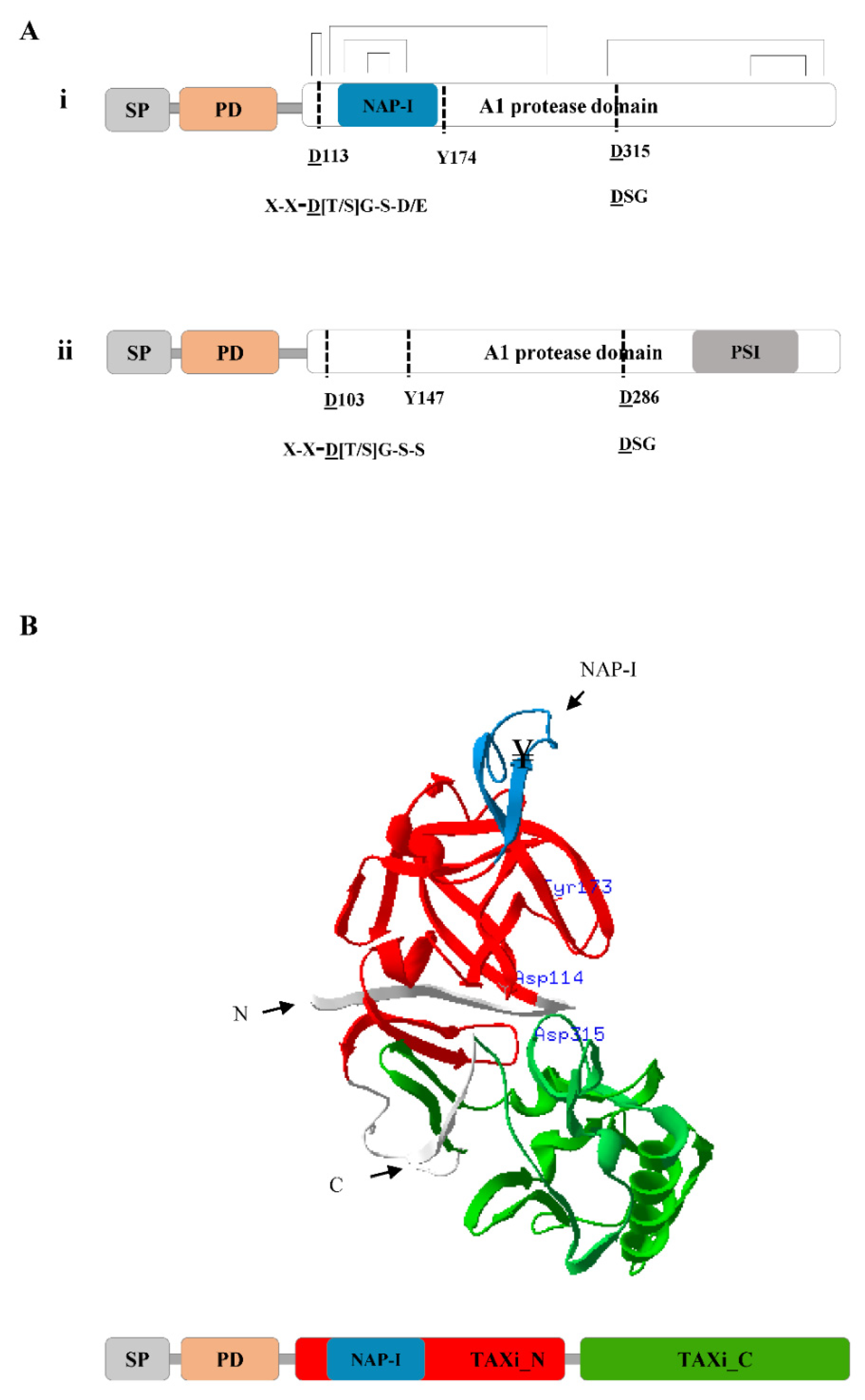
2. Molecular Structure of Plant Nepenthesins
3. Regulation of Plant Nepenthesins
4. Biochemistry of Plant Nepenthesins
5. Biological Roles of Nepenthesins in Carnivorous Plants
5.1. Nepenthesins Contributing to Plant Nutrition
5.2. Nepenthesins Play a Role in Plant Defense
6. Potentials of Plant Nepenthesins
6.1. Industrial Applications: Tool for Digestion in Hydrogen/Deuterium Exchange Mass Spectrometry
6.2. Treatment of Celiac Disease
6.3. Trait Improvement of Crop Plants
7. Conclusions and Future Perspectives
Author Contributions
Funding
Conflicts of Interest
References
- Simões, I.; Faro, C. Structure and function of plant aspartic proteinases. Eur. J. Biochem. 2004, 271, 2067–2075. [Google Scholar] [CrossRef]
- Hiraiwa, N.; Kondo, M.; Nishimura, M.; HaraNishimura, I. An aspartic endopeptidase is involved in the breakdown of propeptides of storage proteins in protein-storage vacuoles of plants. Eur. J. Biochem. 1997, 246, 133–141. [Google Scholar] [CrossRef]
- Mutlu, A.; Gal, S. Plant aspartic proteinases: Enzymes on the way to a function. Physiol. Plant. 1999, 105, 569–576. [Google Scholar] [CrossRef]
- Garciamartinez, J.L.; Moreno, J. Proteolysis of ribulose-1,5-bisphosphate carboxylase oxygenase in citrus leaf extracts. Physiol. Plant. 1986, 66, 377–383. [Google Scholar] [CrossRef]
- An, C.I.; Fukusaki, E.; Kobayashi, A. Aspartic proteinases are expressed in pitchers of the carnivorous plant Nepenthes alata Blanco. Planta 2002, 214, 661–667. [Google Scholar] [CrossRef] [PubMed]
- Tokes, Z.A.; Woon, W.C.; Chambers, S.M. Digestive enzymes secreted by carnivorous plant Nepenthes macferlanei L. Planta 1974, 119, 39–46. [Google Scholar] [CrossRef] [PubMed]
- Ge, X.C.; Dietrich, C.; Matsuno, M.; Li, G.J.; Berg, H.; Xia, Y.J. An Arabidopsis aspartic protease functions as an anti-cell-death component in reproduction and embryogenesis. EMBO Rep. 2005, 6, 282–288. [Google Scholar] [CrossRef] [PubMed]
- Vieira, M.; Pissarra, J.; Verissimo, P.; Castanheira, P.; Costa, Y.; Pires, E.; Faro, C. Molecular cloning and characterization of cDNA encoding cardosin B, an aspartic proteinase accumulating extracellularly in the transmitting tissue of Cynara cardunculus L. Plant Mol. Biol. 2001, 45, 529–539. [Google Scholar] [CrossRef]
- Ramalho-Santos, M.; Pissarra, J.; Veríssimo, P.; Pereira, S.; Salema, R.; Pires, E.; Faro, C.J. Cardosin A, an abundant aspartic proteinase, accumulates in protein storage vacuoles in the stigmatic papillae of Cynara cardunculus L. Planta 1997, 203, 204–212. [Google Scholar] [CrossRef]
- de Carvalho, M.H.C.; d’Arcy-Lameta, A.; Roy-Macauley, H.; Gareil, M.; El Maarouf, H.; Pham-Thi, A.T.; Zuily-Fodil, Y. Aspartic protease in leaves of common bean (Phaseolus vulgaris L.) and cowpea (Vigna unguiculata L. Walp): Enzymatic activity, gene expression and relation to drought susceptibility. Febs Lett. 2001, 492, 242–246. [Google Scholar] [CrossRef]
- Simoes, I.; Faro, R.; Bur, D.; Faro, C. Characterization of recombinant CDR1, an Arabidopsis aspartic proteinase involved in disease resistance. J. Biol. Chem. 2007, 282, 31358–31365. [Google Scholar] [CrossRef] [PubMed]
- Schaller, A.; Ryan, C.A. Molecular cloning of a tomato leaf cDNA encoding an aspartic protease, a systemic wound response protein. Plant Mol. Biol. 1996, 31, 1073–1077. [Google Scholar] [CrossRef] [PubMed]
- Guevara, M.G.; Daleo, G.R.; Oliva, C.R. Purification and characterization of an aspartic protease from potato leaves. Physiol. Plant. 2001, 112, 321–326. [Google Scholar] [CrossRef] [PubMed]
- Cordeiro, M.C.; Pais, M.S.; Brodelius, P.E. Tissue-specific expression of multiple forms of cyprosin (aspartic proteinase) in flowers of Cynara cardunculus. Physiol. Plant. 1994, 92, 645–653. [Google Scholar] [CrossRef]
- Chen, F.Q.; Foolad, M.R. Molecular organization of a gene in barley which encodes a protein similar to aspartic protease and its specific expression in nucellar cells during degeneration. Plant Mol. Biol. 1997, 35, 821–831. [Google Scholar] [CrossRef]
- Rawlings, N.D.; Alan, J.; Thomas, P.D.; Huang, X.D.; Bateman, A.; Finn, R.D. The MEROPS database of proteolytic enzymes, their substrates and inhibitors in 2017 and a comparison with peptidases in the PANTHER database. Nucleic Acids Res. 2018, 46, D624–D632. [Google Scholar] [CrossRef]
- Vines, S.H. The Proteolytic Enzyme of Nepenthes. Ann. Bot. 1897, 11, 563–584. [Google Scholar] [CrossRef]
- Vines, S.H. The Proteolytic Enzyme of Nepenthes. Ann. Bot. 1901, 15, 563–573. [Google Scholar] [CrossRef][Green Version]
- Owen, T.P.; Lennon, K.A. Structure and development of the pitchers from the carnivorous plant Nepenthes alata (Nepenthaceae). Am. J. Bot. 1999, 86, 1382–1390. [Google Scholar] [CrossRef]
- Nakayama, S.; Amagase, S. Acid protease in nepenthes—Partial purification and properties of enzyme. Proc. Jpn. Acad. 1968, 44, 358. [Google Scholar] [CrossRef][Green Version]
- Amagase, S.; Nakayama, S.; Tsugita, A. Acid protease in Nepenthes. 2. Study on specificity of nepenthesin. J. Biochem. 1969, 66, 431. [Google Scholar] [CrossRef] [PubMed]
- Amagase, S. Digestive enzymes in insectivorous plants part 3 acid proteases in the genus Nepenthes and Drosera peltata. J. Biochem. 1972, 72, 73–81. [Google Scholar] [CrossRef] [PubMed]
- Athauda, S.B.P.; Matsumoto, K.; Rajapakshe, S.; Kuribayashi, M.; Kojima, M.; Kubomura-Yoshida, N.; Iwamatsu, A.; Shibata, C.; Inoue, H.; Takahashi, K. Enzymic and structural characterization of nepenthesin, a unique member of a novel subfamily of aspartic proteinases. Biochem. J. 2004, 381, 295–306. [Google Scholar] [CrossRef] [PubMed]
- Ravee, R.; Mohd Salleh, F. ‘Imadi; Goh, H.-H. Discovery of digestive enzymes in carnivorous plants with focus on proteases. PeerJ 2018, 6, e4914. [Google Scholar] [CrossRef]
- Takahashi, K.; Matsumoto, K.; Nishii, W.; Muramatsu, M.; Kubota, K.; Shibata, C.; Athauda, S.B.P.; Peradeniya; Lanka, S. Comparative Studies on the Acid Proteinase Activities in the Digestive Fluids of Nepenthes, Cephalotus, Dionaea and Drosera; Carnivorous Plant Newsletter: Pinole, CA, USA, 2009; Volume 38, pp. 75–82. [Google Scholar]
- Takahashi, K.; Nishii, W.; Shibata, C. The digestive fluid of Drosera indica contains a cysteine endopeptidase similar to dionain from Dionaea muscipula. Carniv. Plant Newsl. 2012, 41, 132–134. [Google Scholar]
- Fukushima, K.; Fang, X.; Alvarez-Ponce, D.; Cai, H.; Carretero-Paulet, L.; Chen, C.; Chang, T.-H.; Farr, K.M.; Fujita, T.; Hiwatashi, Y.; et al. Genome of the pitcher plant Cephalotus reveals genetic changes associated with carnivory. Nat. Ecol. Evol. 2017, 1, 0059. [Google Scholar] [CrossRef]
- Schulze, W.X.; Sanggaard, K.W.; Kreuzer, I.; Knudsen, A.D.; Bemm, F.; Thøgersen, I.B.; Bräutigam, A.; Thomsen, L.R.; Schliesky, S.; Dyrlund, T.F.; et al. The protein composition of the digestive fluid from the venus flytrap sheds light on prey digestion mechanisms. Mol. Cell. Proteomics 2012, 11, 1306–1319. [Google Scholar] [CrossRef]
- Takahashi, K.; Niwa, H.; Yokota, N.; Kubota, K.; Inoue, H. Widespread tissue expression of nepenthesin-like aspartic protease genes in Arabidopsis thaliana. Plant Physiol. Biochem. 2008, 46, 724–729. [Google Scholar] [CrossRef]
- Chen, J.; Ouyang, Y.; Wang, L.; Xie, W.; Zhang, Q. Aspartic proteases gene family in rice: Gene structure and expression, predicted protein features and phylogenetic relation. Gene 2009, 442, 108–118. [Google Scholar] [CrossRef]
- Soares, A.; Ribeiro Carlton, S.M.; Simões, I. Atypical and nucellin-like aspartic proteases: Emerging players in plant developmental processes and stress responses. J. Exp. Bot. 2019. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, K.; Athauda, S.B.P.; Matsumoto, K.; Rajapakshe, S.; Kuribayashi, M.; Kojima, M.; Kubomura-Yoshida, N.; Iwamatsu, A.; Shibata, C.; Inoue, H. Nepenthesin, a unique member of a novel subfamily of aspartic proteinases: Enzymatic and structural characteristics. Curr. Protein Pept. Sci. 2005, 6, 513–525. [Google Scholar] [CrossRef] [PubMed]
- Potter, S.C.; Luciani, A.; Eddy, S.R.; Park, Y.; Lopez, R.; Finn, R.D. HMMER web server: 2018 update. Nucleic Acids Res. 2018, 46, W200–W204. [Google Scholar] [CrossRef] [PubMed]
- Guex, N.; Peitsch, M.C. SWISS-MODEL and the Swiss-Pdb Viewer: An environment for comparative protein modeling. Electrophoresis 1997, 18, 2714–2723. [Google Scholar] [CrossRef] [PubMed]
- Fierens, K.; Brijs, K.; Courtin, C.M.; Gebruers, K.; Goesaert, H.; Raedschelders, G.; Robben, J.; Van Campenhout, S.; Volckaert, G.; Delcour, J.A. Molecular identification of wheat endoxylanase inhibitor TAXI-I1, member of a new class of plant proteins. FEBS Lett. 2003, 540, 259–263. [Google Scholar] [CrossRef]
- Sansen, S.; De Ranter, C.J.; Gebruers, K.; Brijs, K.; Courtin, C.M.; Delcour, J.A.; Rabijns, A. Structural basis for inhibition of Aspergillus niger xylanase by Triticum aestivum xylanase inhibitor-I. J. Biol. Chem. 2004, 279, 36022–36028. [Google Scholar] [CrossRef]
- Pollet, A.; Sansen, S.; Raedschelders, G.; Gebruers, K.; Rabijns, A.; Delcour, J.A.; Courtin, C.M. Identification of structural determinants for inhibition strength and specificity of wheat xylanase inhibitors TAXI-IA and TAXI-IIA. FEBS J. 2009, 276, 3916–3927. [Google Scholar] [CrossRef]
- Prasad, B.D.; Creissen, G.; Lamb, C.; Chattoo, B.B. Heterologous expression and characterization of recombinant OsCDR1, a rice aspartic proteinase involved in disease resistance. Protein Expr. Purif. 2010, 72, 169–174. [Google Scholar] [CrossRef]
- Kato, Y.; Murakami, S.; Yamamoto, Y.; Chatani, H.; Kondo, Y.; Nakano, T.; Yokota, A.; Sato, F. The DNA-binding protease, CND41, and the degradation of ribulose-1,5-bisphosphate carboxylase/oxygenase in senescent leaves of tobacco. Planta 2004, 220, 97–104. [Google Scholar] [CrossRef]
- Kato, Y.; Yamamoto, Y.; Murakami, S.; Sato, F. Post-translational regulation of CND41 protease activity in senescent tobacco leaves. Planta 2005, 222, 643–651. [Google Scholar] [CrossRef]
- Faro, C.; Gal, S. Aspartic Proteinase Content of the Arabidopsis Genome. Curr. Protein Pept. Sci. 2005, 6, 493–500. [Google Scholar] [CrossRef]
- Kadek, A.; Tretyachenko, V.; Mrazek, H.; Ivanova, L.; Halada, P.; Rey, M.; Schriemer, D.C.; Man, P. Expression and characterization of plant aspartic protease nepenthesin-1 from Nepenthes gracilis. Protein Expr. Purif. 2014, 95, 121–128. [Google Scholar] [CrossRef] [PubMed]
- Kubota, K.; Metoki, Y.; P Athauda, S.B.; Shibata, C.; Takahashi, K. Stability Profiles of Nepenthesin in Urea and Guanidine Hydrochloride: Comparison with Porcine Pepsin A. Biotechnol. Biochem. 2010, 74, 2323–2326. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Hoeppner, M.; Rey, M.; Kadek, A.; Man, P.; Schriemer, D.C. Recombinant Nepenthesin II for Hydrogen/Deuterium Exchange Mass Spectrometry. Anal. Chem. 2015, 87, 6681–6687. [Google Scholar] [CrossRef] [PubMed]
- van der Hoorn, R.A.L. Plant proteases: From phenotypes to molecular mechanisms. Annu. Rev. Plant Biol. 2008, 59, 191–223. [Google Scholar] [CrossRef]
- Athauda, S.B.P.; Inoue, H.; Iwamatsu, A.; Takahashi, K. Acid proteinase from Nepenthes distillatoria (Bandura). In Aspartic Proteinases: Retroviral and Cellular Enzymes; James, M.N.G., Ed.; Advances in Experimental Medicine and Biology; Plenum Press: New York, NY, USA, 1998; Volume 59, pp. 453–458. ISBN 0-306-45809-8. [Google Scholar]
- Rey, M.; Yang, M.; Burns, K.M.; Yu, Y.; Lees-Miller, S.P.; Schriemer, D.C. Nepenthesin from Monkey Cups for Hydrogen/Deuterium Exchange Mass Spectrometry. Mol. Cell. Proteomics 2013, 12, 464–472. [Google Scholar] [CrossRef]
- Jentsch, J. Enzymes from carnivorous plants (Nepenthes). Isolation of the protease nepenthacin. FEBS Lett. 1972, 21, 273–276. [Google Scholar] [CrossRef]
- Kadek, A.; Mrazek, H.; Halada, P.; Rey, M.; Schriemer, D.C.; Man, P. Aspartic Protease Nepenthesin-1 as a Tool for Digestion in Hydrogen/Deuterium Exchange Mass Spectrometry. Anal. Chem. 2014, 86, 4287–4294. [Google Scholar] [CrossRef]
- Buch, F.; Kaman, W.E.; Bikker, F.J.; Yilamujiang, A.; Mithöfer, A.; Mithoefer, A. Nepenthesin Protease Activity Indicates Digestive Fluid Dynamics in Carnivorous Nepenthes Plants. PLoS ONE 2015, 10, e118853. [Google Scholar] [CrossRef]
- Lee, L.; Zhang, Y.; Ozar, B.; Sensen, C.W.; Schriemer, D.C. Carnivorous Nutrition in Pitcher Plants (Nepenthes spp.) via an Unusual Complement of Endogenous Enzymes. J. Proteome Res. 2016, 15, 3108–3117. [Google Scholar] [CrossRef]
- Bekalu, Z.E.; Dionisio, G.; Madsen, C.K.; Holme, I.B.; Etzerodt, T.P.; Fomsgaard, I.; Jørgensen, L.N.; Brinch-Pedersen, H. Nepenthesin-1 Derived Resistance to Fungal Pathogens in Major Crop Plants; WO 2019/057845 A1, filed 20 September 2018, and issued 28 March 2019; World Intellectual Property Organization: Geneva, Switzerland, 2019. [Google Scholar]
- Ellison, A.M.; Gotelli, N.J. Evolutionary ecology of carnivorous plants. Trends Ecol. Evol. 2001, 16, 623–629. [Google Scholar] [CrossRef]
- Saganová, M.; Bokor, B.; Stolárik, T.; Pavlovič, A. Regulation of enzyme activities in carnivorous pitcher plants of the genus Nepenthes. Planta 2018, 248, 451–464. [Google Scholar] [CrossRef] [PubMed]
- Lei, X.; Porres, J.; Mullaney; Edward; Brinch-Pedersen, H. Phytase: Source, structure and application. In Industrial Enzymes: Structure, Function and Applications; Polina, J., Mac Cabe, A.P., Eds.; Springer: Dordrecht, The Netherlands, 2007; pp. 505–529. [Google Scholar]
- Nasmith, C.G.; Walkowiak, S.; Wang, L.; Leung, W.W.Y.; Gong, Y.; Johnston, A.; Harris, L.J.; Guttman, D.S.; Subramaniam, R. Tri6 Is a Global Transcription Regulator in the Phytopathogen Fusarium graminearum. PLoS Pathog. 2011, 7, e1002266. [Google Scholar] [CrossRef] [PubMed]
- Hohn, T.M.; Vanmiddlesworth, F. Purification and characterization of the sesquiterpene cyclase trichodiene synthetase from Fusarium sporotrichioides. Arch. Biochem. Biophys. 1986, 251, 756–761. [Google Scholar] [CrossRef]
- Tokai, T.; Koshino, H.; Takahashi-Ando, N.; Sato, M.; Fujimura, M.; Kimura, M. Fusarium Tri4 encodes a key multifunctional cytochrome P450 monooxygenase for four consecutive oxygenation steps in trichothecene biosynthesis. Biochem. Biophys. Res. Commun. 2007, 353, 412–417. [Google Scholar] [CrossRef]
- Bekalu, Z.E.; Krogh Madsen, C.; Dionisio, G.; Bæksted Holme, I.; Jørgensen, L.N.; Fomsgaard, I.S.; Brinch-Pedersen, H. Overexpression of Nepenthesin HvNEP-1 in Barley Endosperm Reduces Fusarium Head Blight and Mycotoxin Accumulation. Agronomy 2020, 10, 203. [Google Scholar] [CrossRef]
- Marcsisin, S.R.; Engen, J.R. Hydrogen exchange mass spectrometry: What is it and what can it tell us? Anal. Bioanal. Chem. 2010, 397, 967–972. [Google Scholar] [CrossRef]
- Konermann, L.; Pan, J.; Liu, Y.H. Hydrogen exchange mass spectrometry for studying protein structure and dynamics. Chem. Soc. Rev. 2011, 40, 1224–1234. [Google Scholar] [CrossRef]
- Rey, M.; Yang, M.; Lee, L.; Zhang, Y.; Sheff, J.G.; Sensen, C.W.; Mrazek, H.; Halada, P.; Man, P.; McCarville, J.L.; et al. Addressing proteolytic efficiency in enzymatic degradation therapy for celiac disease. Sci. Rep. 2016, 6, 30980. [Google Scholar] [CrossRef]
- Schriemer, D.; Man, P.; Mrazek, H.; Rey, M. Treatment of Gluten Intolerance and Related Conditions. US 2015/0265686 A1, filed 3 October 2014, and issued 24 September 2015. U.S. Patent 9005610, 15 March 2013. [Google Scholar]
- Abadie, V.; Sollid, L.M.; Barreiro, L.B.; Jabri, B. Integration of Genetic and Immunological Insights into a Model of Celiac Disease Pathogenesis. Annu. Rev. Immunol. 2011, 29, 493–525. [Google Scholar] [CrossRef]
- Bethune, M.T.; Khosla, C. Oral enzyme therapy for celiac sprue. Methods Enzymol. 2012, 502, 241–271. [Google Scholar]
- Shan, L.; Molberg, Ø.; Parrot, I.; Hausch, F.; Filiz, F.; Gray, G.M.; Sollid, L.M.; Khosla, C. Structural basis for gluten intolerance in Celiac Sprue. Science (80-.) 2002, 297, 2275–2279. [Google Scholar] [CrossRef] [PubMed]
- Shan, L.; Qiao, S.-W.; Arentz-Hansen, H.; Molberg, Ø.; Gray, G.M.; Sollid, L.M.; Khosla, C. Identification and Analysis of Multivalent Proteolytically Resistant Peptides from Gluten: Implications for Celiac Sprue. J. Proteome Res. 2005, 4, 1732–1741. [Google Scholar] [CrossRef] [PubMed]
- Bethune, M.T.; Strop, P.; Tang, Y.; Sollid, L.M.; Khosla, C. Heterologous Expression, Purification, Refolding, and Structural-Functional Characterization of EP-B2, a Self-Activating Barley Cysteine Endoprotease. Chem. Biol. 2006, 13, 637–647. [Google Scholar] [CrossRef] [PubMed]
- Gil-Humanes, J.; Pistón, F.; Altamirano-Fortoul, R.; Real, A.; Comino, I.; Sousa, C.; Rosell, C.M.; Barro, F. Reduced-Gliadin Wheat Bread: An Alternative to the Gluten-Free Diet for Consumers Suffering Gluten-Related Pathologies. PLoS ONE 2014, 9, e90898. [Google Scholar] [CrossRef] [PubMed]
- Jouanin, A.; Boyd, L.; Visser, R.G.F.; Smulders, M.J.M. Development of Wheat With Hypoimmunogenic Gluten Obstructed by the Gene Editing Policy in Europe. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Sánchez-León, S.; Gil-Humanes, J.; Ozuna, C.V.; Giménez, M.J.; Sousa, C.; Voytas, D.F.; Barro, F. Low-gluten, nontransgenic wheat engineered with CRISPR/Cas9. Plant Biotechnol. J. 2018, 16, 902–910. [Google Scholar] [CrossRef]
- Martinez, M.; Gómez-Cabellos, S.; Giménez, M.J.; Barro, F.; Diaz, I.; Diaz-Mendoza, M. Plant Proteases: From Key Enzymes in Germination to Allies for Fighting Human Gluten-Related Disorders. Front. Plant Sci. 2019, 10. [Google Scholar] [CrossRef]
- Gass, J.; Bethune, M.T.; Siegel, M.; Spencer, A.; Khosla, C. Combination Enzyme Therapy for Gastric Digestion of Dietary Gluten in Patients with Celiac Sprue. Gastroenterology 2007, 133, 472–480. [Google Scholar] [CrossRef]
- Desjardins, A.E. Fusarium Mycotoxins: Chemistry, Genetics and Biology. Plant Pathol. 2007, 56, 337. [Google Scholar]
- European Commission. Commission regulation (EU) 1881/2006 of 19 December 2006 setting maximum levels for certain contaminants in foodstuffs. Off. J. Eur. Union L 2006, 364, 5–24. [Google Scholar]
- Muñoz, F.F.; Mendieta, J.R.; Pagano, M.R.; Paggi, R.A.; Daleo, G.R.; Guevara, M.G. The swaposin-like domain of potato aspartic protease (StAsp-PSI) exerts antimicrobial activity on plant and human pathogens. Peptides 2010, 31, 777–785. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Source (Species) | Proteases | Molecular Weight (kDa) | Specific Activity | Substrates | Optimum | Inhibitors | Substrate Specificity | References | ||
|---|---|---|---|---|---|---|---|---|---|---|
| pH | T (°C) | Substrates | Cleavage Specificity | |||||||
| Nepenthes sp. | Nepenthesin | n.d. | 0.55 U mL−1 | Casein | 2.8 | 40 | n.d. | Peptides (8) | Leu-Asp, Ser-Asp, Thr-Asp, Ala-Ala, Tyr-Asp | [21] |
| N. maxima N. rafflesiana N. ampullaria N. x dyeriana N. x mixta Drosera peltata | Nepenthesin | n.d. | n.d. | Casein | 3.0 | 40 | n.d. | Peptides (6) | Asp-Glu, Asp-Ala, Ala-Ala, Lys-Arg | [22] |
| Nepenthes sp. | Nepenthesin | n.d. | n.d. | Casein | 2.9 | 40 | n.d. | n.d. | n.d. | [48] |
| N. macfarlanei | Nepenthesin I Nepenthesin II | 59.00 21.00 | n.d. n.d. | Bovine fibrin Bovine serum albumin Horse-heart cytochrome c | n.d. n.d. 2.2 | 37 37 37 | Pepstatin | Horse-heart cytochrome c | Lys-Ala, Glu-Asp, Glu-Thr, Lys-Thr, Gly-Gly, Leu-Phe | [6] |
| N. distillatoria | Nepenthesin I Nepenthesin II | 51.00 45.00 | 874 U mg−1 809 U mg−1 | Acid-denatured haemoglobin | 2.6 2.6 | 55 45 | Pepstatin, DAN | Oxidized insulin B chain | Phe-Phe, Glu-Ala, Leu-Cya, Leu-Tyr, Tyr-Leu | [23] |
| N. alata,
Cephalotus follicularis D. muscipula D. capensis | Nepenthesin I Nepenthesin II | n.d. n.d. | n.d. n.d. | Haemoglobin Haemoglobin Oxidized insulin B chain | 2.5 3.0 3.5 | 47–57 60 47 | Pepstatin | Oxidized insulin B chain | Leu-Tyr, Phe-Phe, Glu-Ala, Ala-Leu, Tyr-Leu, Tyr-Thr, Lys-Ala, Gly-Phe, | [25] |
| N. alata | Nepenthesin I Nepenthesin IIa Nepenthesin IIb | n.d. n.d. n.d. | n.d. n.d. n.d. | Haemoglobin | n.d. n.d. n.d. | n.d. n.d. n.d. | n.d. n.d. n.d. | n.d. n.d. n.d. | n.d. n.d. n.d. | [43] |
| N. gracilis | Nepenthesin 1 | 43.73 | n.d. | Haemoglobin Myoglobin | 2.5 n.d. | 50–60 n.d. | n.d. | n.d. | n.d. | [42,49] |
| N. mirabilis | Nepenthesin I Nepenthesin II | n.d. 45.00 | n.d. n.d. | PFU-093 (FRET peptide substrate) | 8.0 8.0 | 42 42 | Pepstatin A | n.d. | n.d. | [50] |
| N. alata | Nepenthesin I Nepenthesin II | n.d. n.d. | n.d. n.d. | PFU-093 (FRET peptide substrate) | 8.0 8.0 | 42 42 | Pepstatin A | n.d. | n.d. | |
| N. reinwardtiana N. distillatoria N. eymae N. wittei N. hookeriana N. boschiana N. maxima | Nepenthesin I Nepenthesin II | n.d. n.d. | n.d. n.d. | PFU-093 (FRET peptide substrate) | 8.0 8.0 | 42 42 | Pepstatin A | n.d. | n.d. | |
| N. gracilis * | Nepenthesin I Nepenthesin II | n.d. 37.50 | n.d. n.d. | Haemoglobin | 2.5 2.5 | 50 55 | n.d. | XRCC4, XLF, PNK, BRCT, myoglobin | Ser-Ilu/Pro/Thr//Phe, Glu-Glu, Leu-Tyr, Phe-Phe, Glu-Ala, Ala-Leu | [44] |
| N. rafflesiana | Nepenthesin-1 Nepenthesin-2 Nepenthesin-3 Nepenthesin-4 Nepenthesin-5 | 47.11 46.63 49.09 48.99 48.81 | n.d. n.d. n.d. n.d. n.d. | None | n.d. n.d. n.d. n.d. n.d. | n.d. n.d. n.d. n.d. n.d. | n.d. | n.d. | n.d. | [51] |
| H. vulgare | Nepenthesin-1 (HvNEP-1) | 48.90 | n.d. | Aspergillus ficuum phytase | 5.0 | 40 | Pepstatin A | n.d. | n.d. | [52] |
| Applications | Proteases | Purification | Substrates | Molar Ratio (Protease: Substrate) | References |
|---|---|---|---|---|---|
| HDX-MS | Porcine pepsin | Reagent grade (Sigma) | XRCC4, XLF, BRCT, PNK, myoglobin, cytochrome C | 1:0.00192 | [47] |
| Nepenthesin II | Crude | XRCC4, XLF, BRCT, PNK, myoglobin, cytochrome C | 1:2.63 | ||
| Celiac treatment | CysProt EP-B2 | Recombinant (E. coli) | α2-gliadin | 1:10 | [68] |
| Nepenthesin II | Crude | α2-gliadin | 1:12000 | [62] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bekalu, Z.E.; Dionisio, G.; Brinch-Pedersen, H. Molecular Properties and New Potentials of Plant Nepenthesins. Plants 2020, 9, 570. https://doi.org/10.3390/plants9050570
Bekalu ZE, Dionisio G, Brinch-Pedersen H. Molecular Properties and New Potentials of Plant Nepenthesins. Plants. 2020; 9(5):570. https://doi.org/10.3390/plants9050570
Chicago/Turabian StyleBekalu, Zelalem Eshetu, Giuseppe Dionisio, and Henrik Brinch-Pedersen. 2020. "Molecular Properties and New Potentials of Plant Nepenthesins" Plants 9, no. 5: 570. https://doi.org/10.3390/plants9050570
APA StyleBekalu, Z. E., Dionisio, G., & Brinch-Pedersen, H. (2020). Molecular Properties and New Potentials of Plant Nepenthesins. Plants, 9(5), 570. https://doi.org/10.3390/plants9050570