Silver Can Induce Oxidative Stress in Parallel to Other Chemical Elicitors to Modulate the Ripening of Chili Cultivars



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
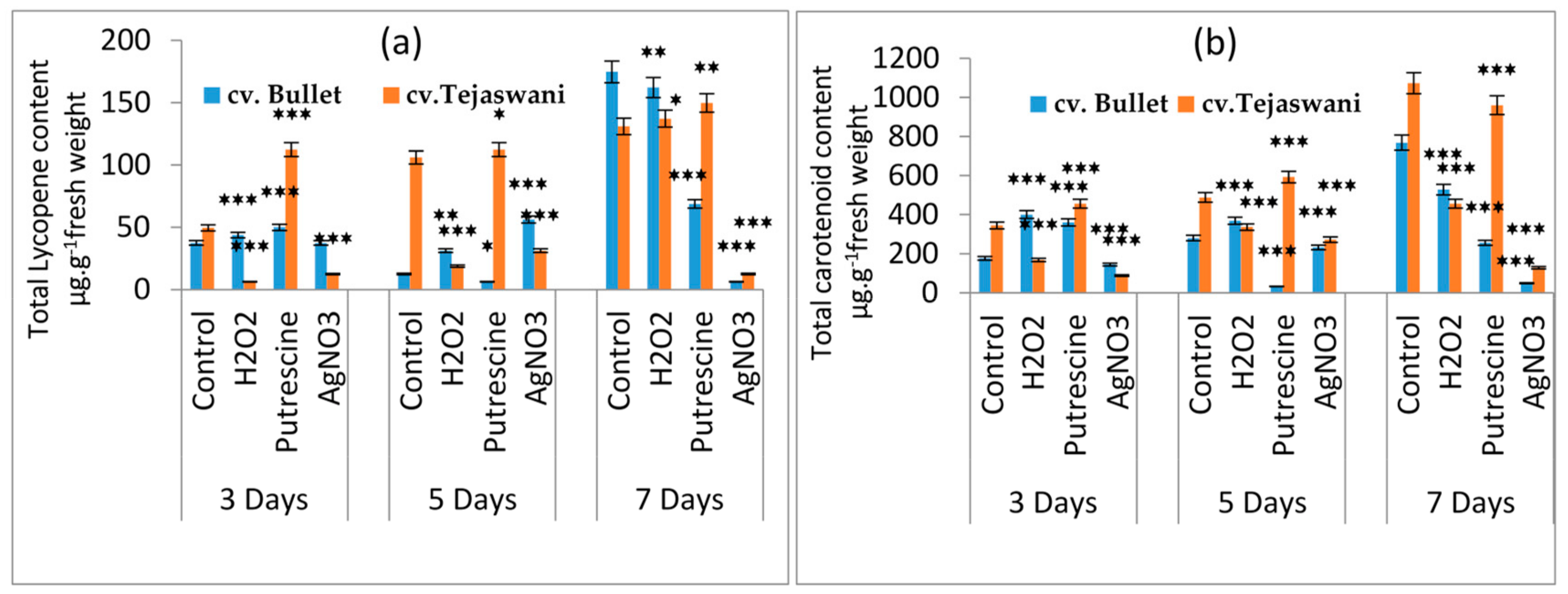
2.1. Effect of Chemical Treatments on Lycopene and Carotenoid Content
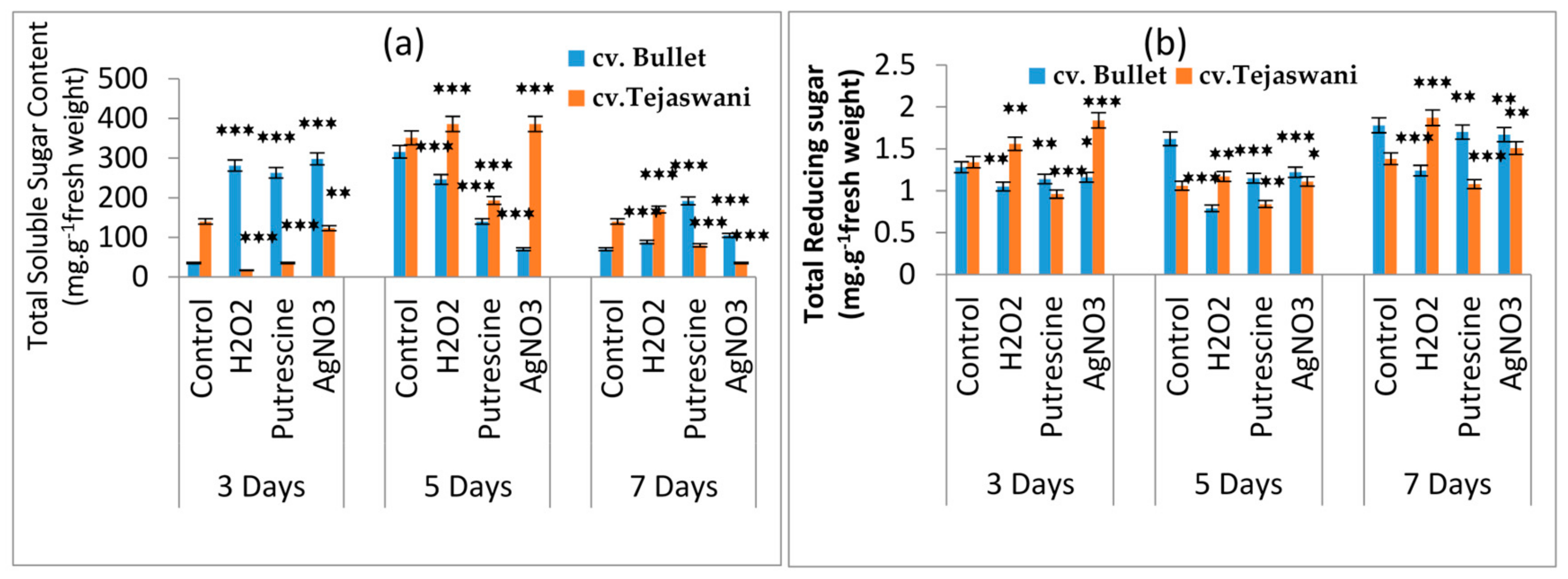
2.2. Changes of Reducing Sugar and Total Sugar Content
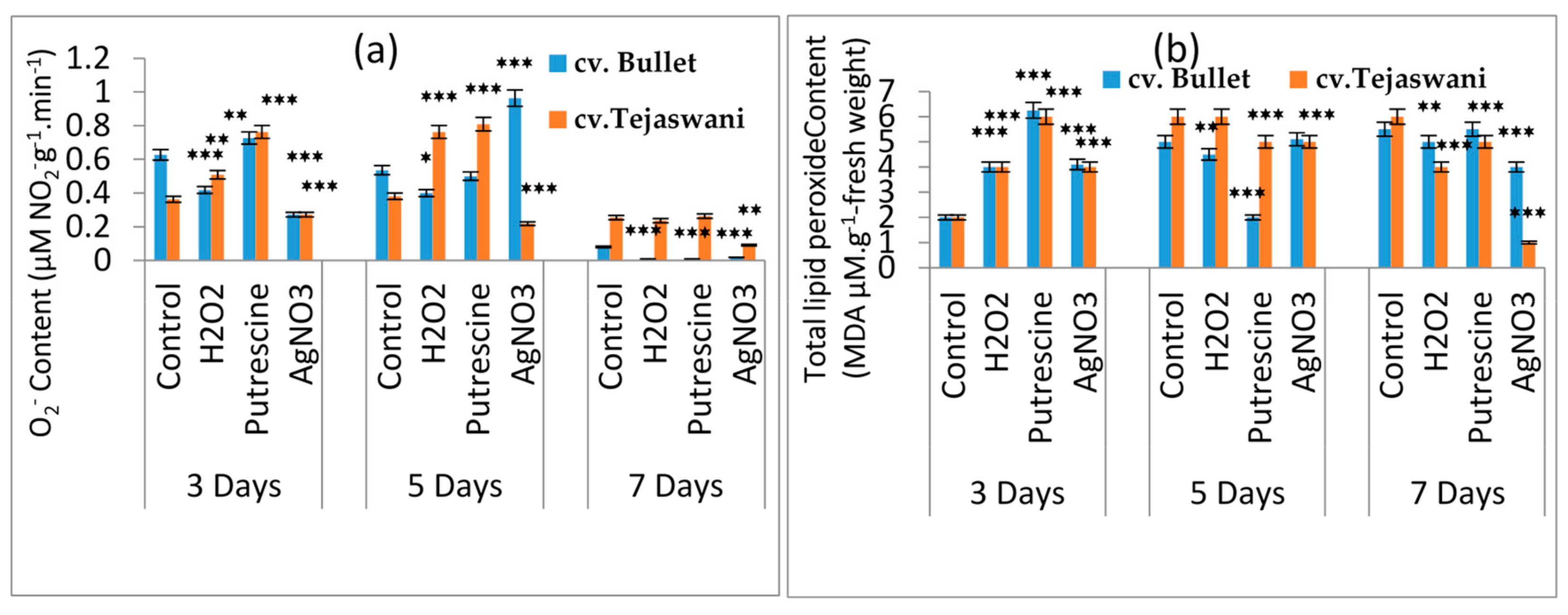
2.3. Influence of Treatments on H2O2 and Lipid Peroxide Production
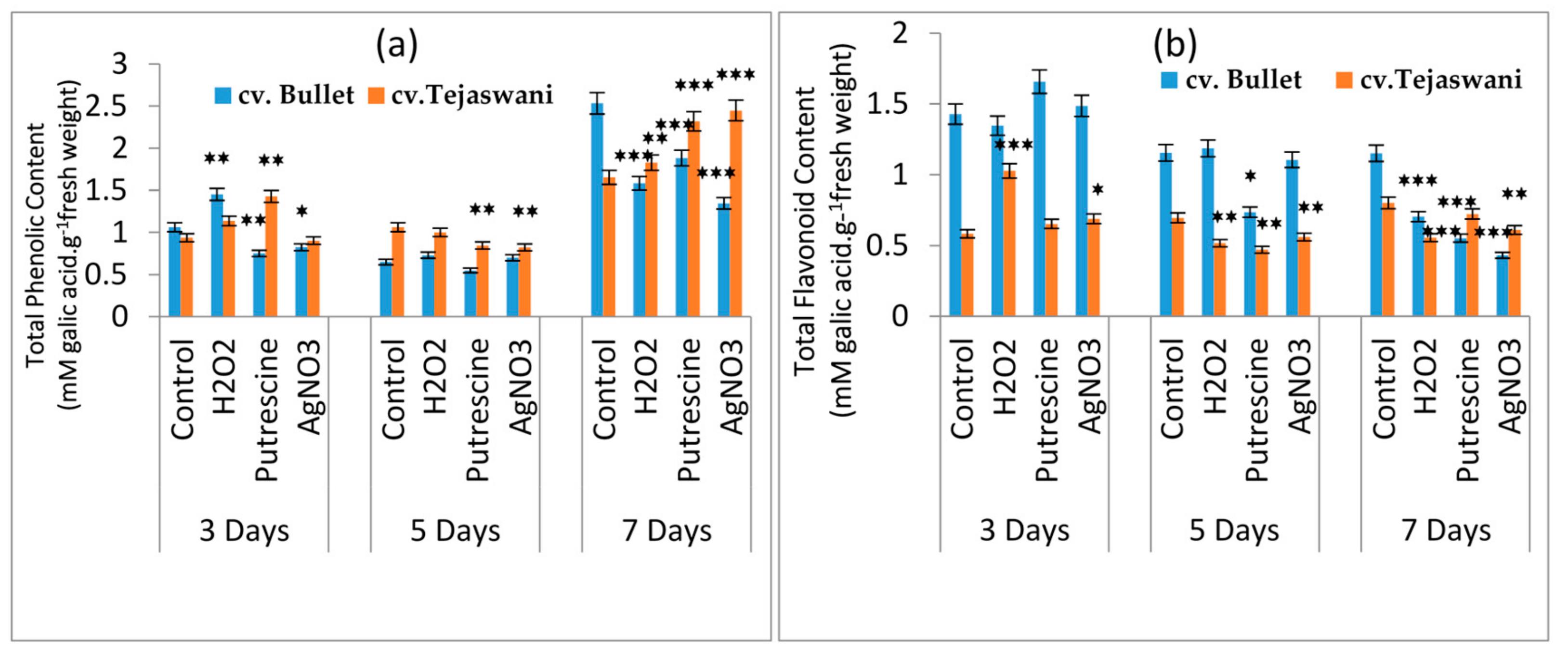
2.4. Changes of Phenolic Content and total Flavonoid Content
2.5. Activity of Pectin Methylesterase (PME) under Different Treatments
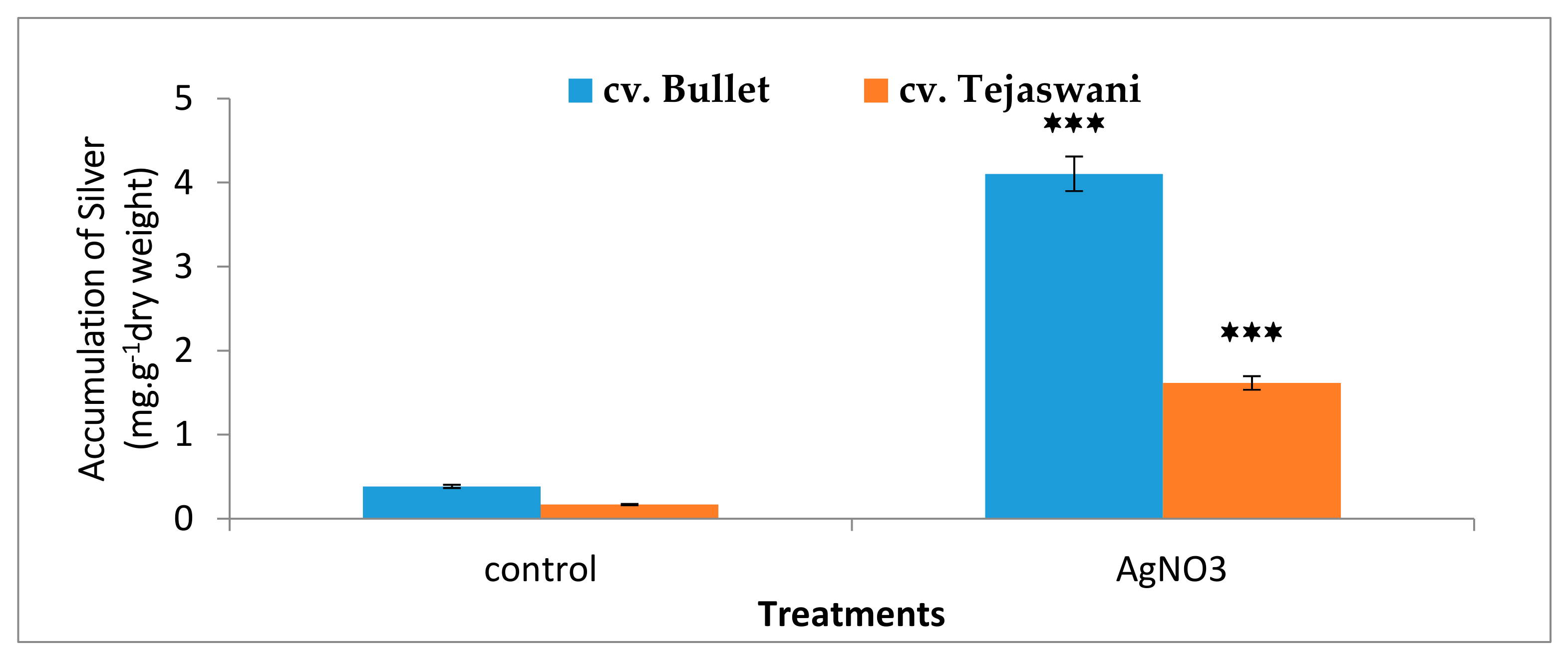
2.6. Bioaccumulation of Silver in the Tissue
3. Discussion
4. Materials and Methods
4.1. Plant Materials and Treatments
4.2. Estimation of Carbohydrate Content
4.3. Estimation of Total Lycopene and Carotenoid Content
4.4. Estimation of Total Phenolic and Flavonoid Content
4.5. Detection of H2O2, O2-, and Lipid Peroxide
4.6. Assay of Pectin Methylesterase (PME)
4.7. Quantification of Silver Content
4.8. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Martínez-Lüscher, J.; Morales, F.; Sánchez-Díaz, M.; Delrot, S.; Aguirreolea, J.; Gomes, E.; Pascual, I. Climate change conditions (elevated CO2 and temperature) and UV-B radiation affect grapevine (Vitisvinifera cv. Tempranillo) leaf carbon assimilation, altering fruit ripening rates. Plant Sci. 2015, 236, 168–176. [Google Scholar] [CrossRef]
- Basu, S.; Roychoudhury, A.; Saha, P.P.; Sengupta, D.N. Differential antioxidative responses of indica rice cultivars to drought stress. Plant Growth Regul. 2009, 60, 51–59. [Google Scholar] [CrossRef]
- Wang, T.-D.; Zhang, H.-F.; Wu, Z.-C.; Li, J.-G.; Huang, X.-M. Sugar Uptake in the Aril of Litchi Fruit Depends on the Apoplasmic Post-Phloem Transport and the Activity of Proton Pumps and the Putative Transporter LcSUT4. Plant Cell Physiol. 2015, 56, 377–387. [Google Scholar] [CrossRef] [PubMed]
- Decros, G.; Baldet, P.; Beauvoit, B.; Stevens, R.; Flandin, A.; Colombié, S.; Gibon, Y.; Pétriacq, P. Get the Balance Right: ROS Homeostasis and Redox Signalling in Fruit. Front. Plant Sci. 2019, 10, 1091. [Google Scholar] [CrossRef] [PubMed]
- Del Río, L.A.; López-Huertas, E. ROS generation in peroxisomes and its role in cell signaling. Plant Cell Physiol. 2016, 57, 1364–1376. [Google Scholar] [CrossRef] [PubMed]
- Ramos, P.; Parra-Palma, C.; Figueroa, C.R.; Zuñiga, P.E.; Valenzuela-Riffo, F.; Gonzàlez, J.; Gaete-Eastman, C.; Morales-Quintana, L. Cell wall-related enzymatic activities and transcriptional profiles in four strawberry (Fragaria x ananassa) cultivars during fruit development and ripening. Sci. Hortic. 2018, 238, 325–332. [Google Scholar] [CrossRef]
- Bortolin, R.C.; Caregnato, F.F.; Junior, A.M.D.; Zanotto-Filho, A.; Moresco, K.S.; Rios, A.D.O.; Salvi, A.D.O.; Ortmann, C.F.; De Carvalho, P.; Reginatto, F.H.; et al. Chronic ozone exposure alters the secondary metabolite profile, antioxidant potential, anti-inflammatory property, and quality of red pepper fruit from Capsicum baccatum. Ecotoxicol. Environ. Saf. 2016, 129, 16–24. [Google Scholar] [CrossRef]
- Brandes, N.; Zude-Sasse, M. Respiratory patterns of European pear (Pyruscommunis L. ‘Conference’) throughout pre- and post-harvest fruit development. Heliyon 2019, 5, e01160. [Google Scholar] [CrossRef]
- Kumar, V.; Parvatam, G.; Ravishankar, G.A. AgNO3—A potential regulator of ethylene activity and plant growth modulator. Electron. J. Biotechnol. 2009, 12, 8–9. [Google Scholar] [CrossRef]
- Hussain, M.; Raja, N.I.; Iqbal, M.; Aslam, S. Applications of plant flavonoids in the green synthesis of colloidal silver nanoparticles and impacts on human health. Iran. J. Sci. Technol. Trans. A Sci. 2019, 43, 1381–1392. [Google Scholar] [CrossRef]
- Tavakoli, S.A.; Mirzaei, S.; Rahimi, M.; Tavassolian, I. Assessment of peppermint, clove, cumin essential oils and silver nano particles on biochemical and shelf life of Citrus limon(L.). Indian J. Biochem. Biophys. IJBB. 2019, 56, 269–275. [Google Scholar]
- Al-Saikhan, M.S.; Shalaby, T.A. Effect of hydrogen peroxide (H2O2) treatment on physicochemical characteristics of tomato fruits during post-harvest storage. Aust. J. Crop Sci. 2019, 13, 798. [Google Scholar] [CrossRef]
- Hosseini, M.S.; Babalar, M.; Askari, M.A.; Zahedi, S.M. Comparison the effect of putrescine application on postharvest quality of Pyruscommunis cv. “Shah-Miveh” and “Spadona.”. Food Sci. Nutr. 2018, 7, 14–21. [Google Scholar] [CrossRef] [PubMed]
- Müller, L.; Caris-Veyrat, C.; Lowe, G.; Böhm, V. Lycopene and Its Antioxidant Role in the Prevention of Cardiovascular Diseases—A Critical Review. Crit. Rev. Food Sci. Nutr. 2015, 56, 1868–1879. [Google Scholar] [CrossRef]
- Ahmed, B.; Hashmi, A.; Khan, M.S.; Musarrat, J. ROS mediated destruction of cell membrane, growth and biofilms of human bacterial pathogens by stable metallic AgNPs functionalized from bell pepper extract and quercetin. Adv. Powder Technol. 2018, 29, 1601–1616. [Google Scholar] [CrossRef]
- An, J.; Almasaud, R.A.; Bouzayen, M.; Zouine, M.; Chervin, C. Auxin and ethylene regulation of fruit set. Plant Sci. 2020, 292, 110381. [Google Scholar] [CrossRef]
- Seifi, H.S.; Shelp, B.J. Spermine Differentially Refines Plant Defense Responses against Biotic and Abiotic Stresses. Front. Plant Sci. 2019, 10, 117. [Google Scholar] [CrossRef]
- Zhang, Y.; Gong, Y.; Chen, L.; Peng, Y.; Wang, Q.; Shi, J. Hypotaurine delays senescence of peach fruit by regulating reactive oxygen species metabolism. Sci. Hortic. 2019, 253, 295–302. [Google Scholar] [CrossRef]
- Podlešáková, K.; Ugena, L.; Spíchal, L.; Doležal, K.; De Diego, N. Phytohormones and polyamines regulate plant stress responses by altering GABA pathway. New Biotechnol. 2019, 48, 53–65. [Google Scholar] [CrossRef]
- Kärkönen, A.; Kuchitsu, K. Reactive oxygen species in cell wall metabolism and development in plants. Phytochemistry 2015, 112, 22–32. [Google Scholar] [CrossRef]
- De Oliveira, A.B.; de Almeida Lopes, M.M.; Moura, C.F.; de Siqueira Oliveira, L.; de Souza, K.O.; Gomes Filho, E.; Urban, L.; de Miranda, M.R. Effects of organic vs. conventional farming systems on quality and antioxidant metabolism of passion fruit during maturation. Sci. Hortic. 2017, 222, 84–89. [Google Scholar] [CrossRef]
- Berni, R.; Luyckx, M.; Xu, X.; Legay, S.; Sergeant, K.; Hausman, J.-F.; Lutts, S.; Cai, G.; Guerriero, G. Reactive oxygen species and heavy metal stress in plants: Impact on the cell wall and secondary metabolism. Environ. Exp. Bot. 2019, 161, 98–106. [Google Scholar] [CrossRef]
- Zhang, L.; Liu, P.; Li, L.; Huang, Y.; Pu, Y.; Hou, X.; Song, L. Identification and Antioxidant Activity of Flavonoids Extracted from Xinjiang Jujube (Ziziphus jujube Mill.) Leaves with Ultra-High Pressure Extraction Technology. Molecules 2018, 24, 122. [Google Scholar] [CrossRef] [PubMed]
- Fortes, A.M.; Agudelo-Romero, P.; Pimentel, D.; Alkan, N. Transcriptional Modulation of Polyamine Metabolism in Fruit Species under Abiotic and Biotic Stress. Front. Plant Sci. 2019, 10, 816. [Google Scholar] [CrossRef]
- Jain, V.; Chawla, S.; Choudhary, P.; Jain, S. Post-harvest calcium chloride treatments influence fruit firmness, cell wall components and cell wall hydrolyzing enzymes of Ber(ZiziphusmauritianaLamk.) fruits during storage. J. Food Sci. Technol. 2019, 56, 4535–4542. [Google Scholar] [CrossRef]
- Zhang, L.; Wang, P.; Chen, F.; Lai, S.; Yu, H.; Yang, H. Effects of calcium and pectin methylesterase on quality attributes and pectin morphology of jujube fruit under vacuum impregnation during storage. Food Chem. 2019, 289, 40–48. [Google Scholar] [CrossRef]
- Hodge, J.E. Determination of reducing sugars and carbohydrates. Methods Carbohydr. Chem. 1962, 1, 380–394. [Google Scholar]
- Miller, G.L. Estimation of reducing sugar by dinitrosalicylic acid method. Anal. Chem. 1972, 31, 426–428. [Google Scholar] [CrossRef]
- Zakaria, M.; Simpson, K.; Brown, P.R.; Krstulović, A. Use of reversed-phase high-performance liquid chromatographic analysis for the determination of provitaminA carotenes in tomatoes. J. Chromatogr. A 1979, 176, 109–117. [Google Scholar] [CrossRef]
- Heath, R.L.; Packer, L. Photoperoxidation in isolated chloroplasts: I. Kinetics and stoichiometry of fatty acid peroxidation. Arch. Biochem. Biophys. 1968, 125, 189–198. [Google Scholar] [CrossRef]
- Hobson, G.; Norris, F.W.; Schryver, S.B.; Resch, C.E.; Buston, H.W.; Nanji, H.R. Polygalacturonase in normal and abnormal tomato fruit. Biochem. J. 1964, 92, 324–332. [Google Scholar] [CrossRef] [PubMed]
- Choleva, T.G.; Tsogas, G.Z.; Giokas, D.L. Determination of silver nanoparticles by atomic absorption spectrometry after dispersive suspended microextraction followed by oxidative dissolution back-extraction. Talanta 2019, 196, 255–261. [Google Scholar] [CrossRef] [PubMed]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ghosh, A.; Saha, I.; Dolui, D.; De, A.K.; Sarkar, B.; Adak, M.K. Silver Can Induce Oxidative Stress in Parallel to Other Chemical Elicitors to Modulate the Ripening of Chili Cultivars. Plants 2020, 9, 238. https://doi.org/10.3390/plants9020238
Ghosh A, Saha I, Dolui D, De AK, Sarkar B, Adak MK. Silver Can Induce Oxidative Stress in Parallel to Other Chemical Elicitors to Modulate the Ripening of Chili Cultivars. Plants. 2020; 9(2):238. https://doi.org/10.3390/plants9020238
Chicago/Turabian StyleGhosh, Arijit, Indraneel Saha, Debabrata Dolui, Arnab Kumar De, Bipul Sarkar, and Malay Kumar Adak. 2020. "Silver Can Induce Oxidative Stress in Parallel to Other Chemical Elicitors to Modulate the Ripening of Chili Cultivars" Plants 9, no. 2: 238. https://doi.org/10.3390/plants9020238
APA StyleGhosh, A., Saha, I., Dolui, D., De, A. K., Sarkar, B., & Adak, M. K. (2020). Silver Can Induce Oxidative Stress in Parallel to Other Chemical Elicitors to Modulate the Ripening of Chili Cultivars. Plants, 9(2), 238. https://doi.org/10.3390/plants9020238