Investigating Effects of Bordered Pit Membrane Morphology and Properties on Plant Xylem Hydraulic Functions—A Case Study from 3D Reconstruction and Microflow Modelling of Pit Membranes in Angiosperm Xylem

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Role of the Pit Membrane Morphology in Plant Hydraulics
2.1. Pit Membrane Thickness Variation and Its Hydraulic Functional Role
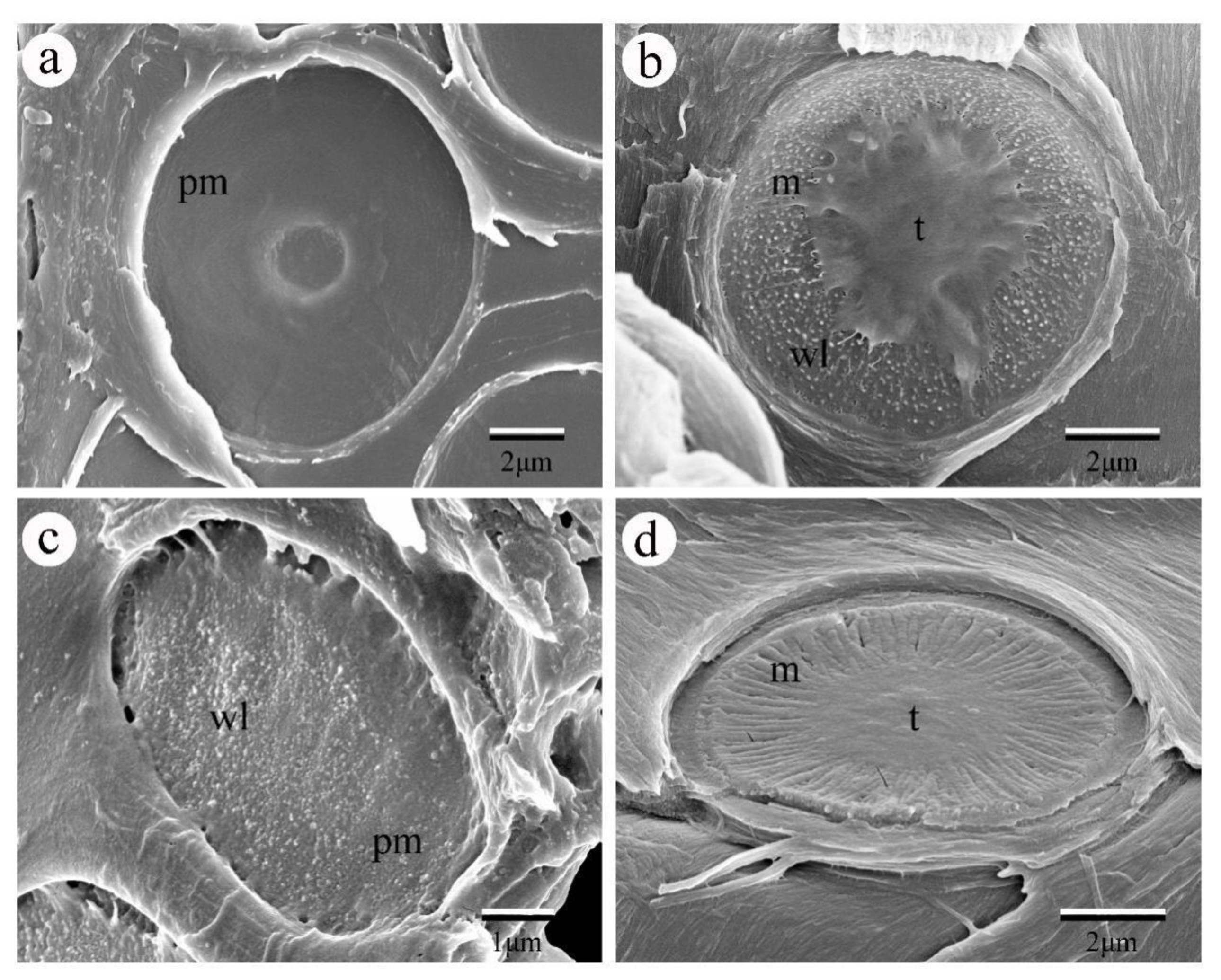
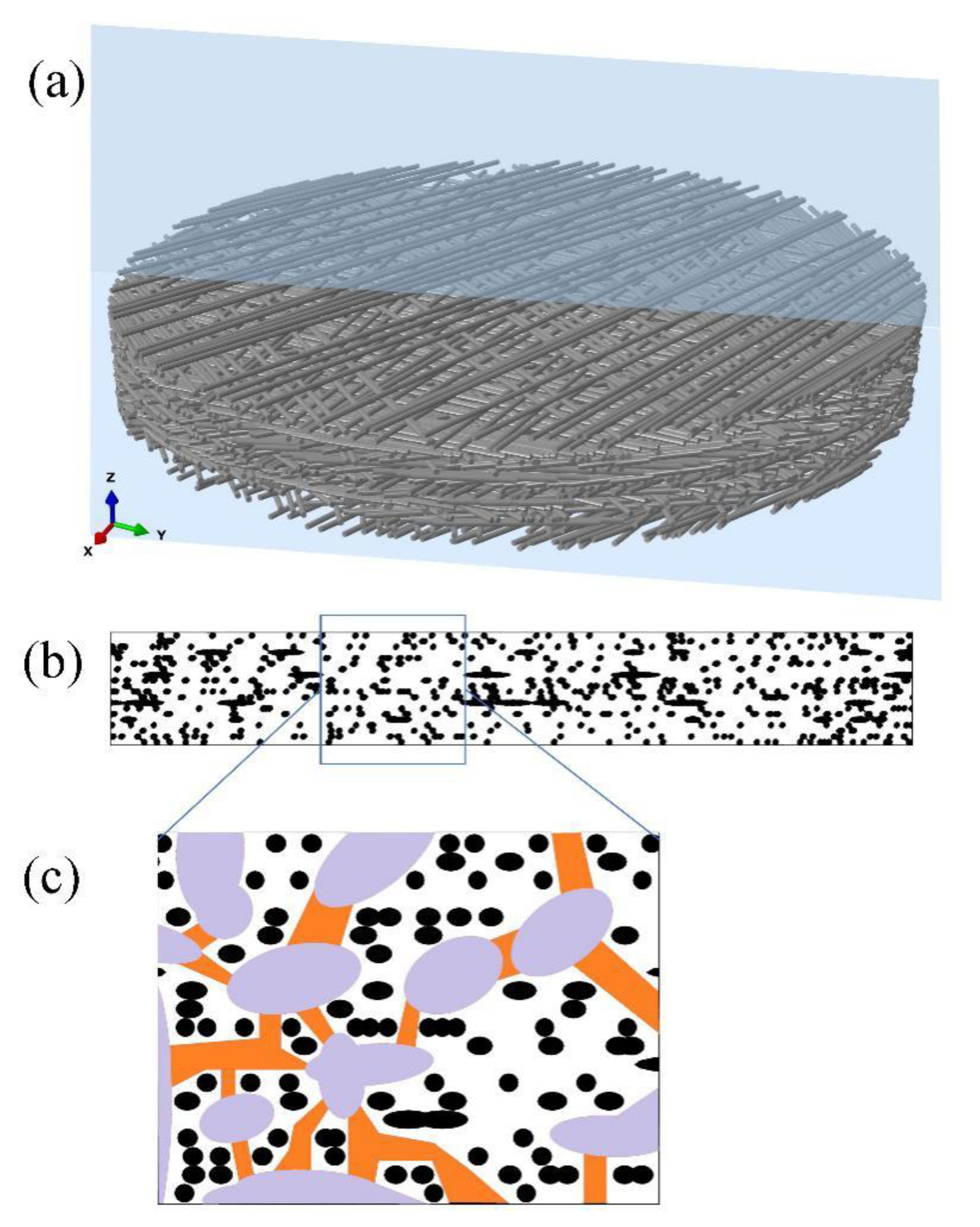
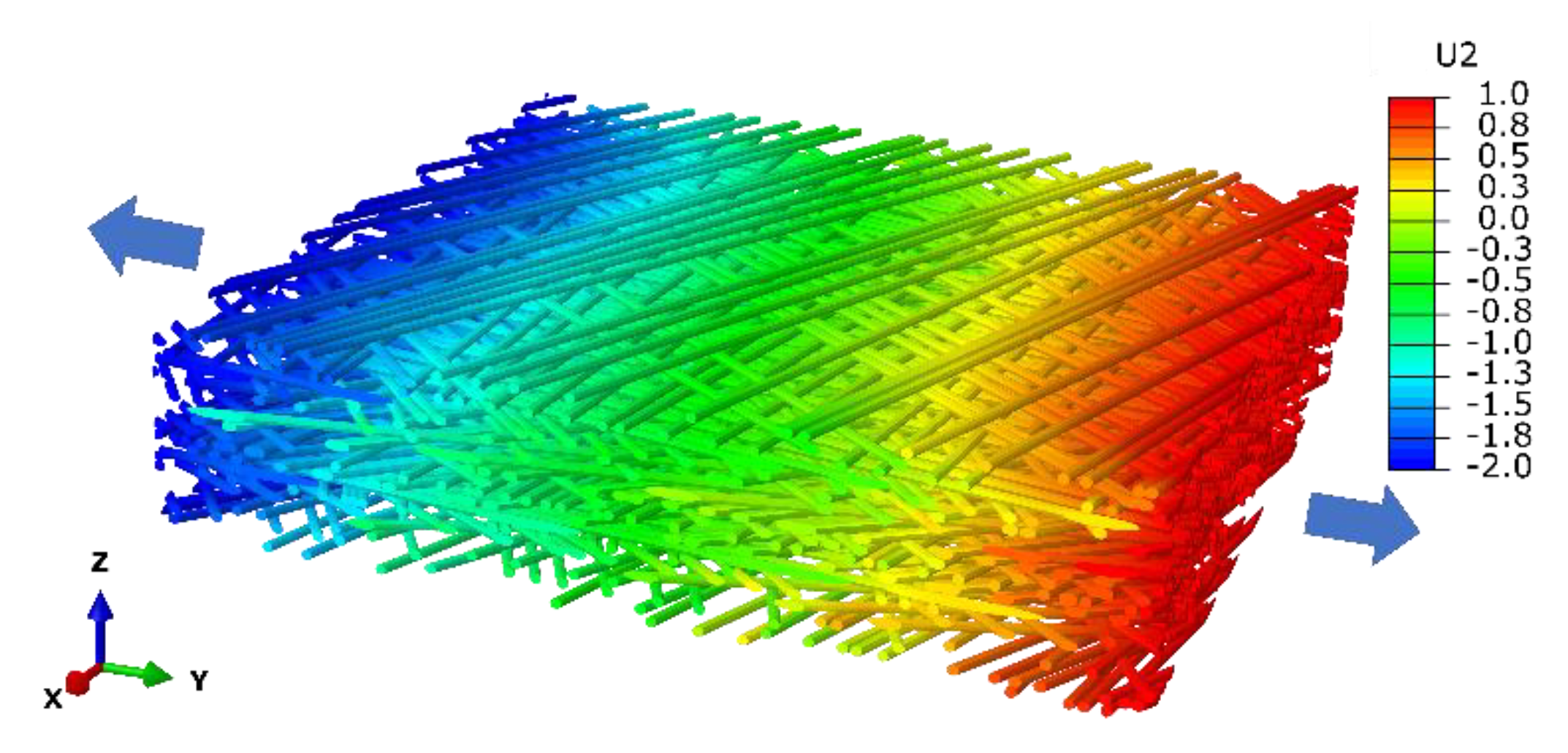
2.2. Three Dimension of Intervessel Pit Membrane and Its Function
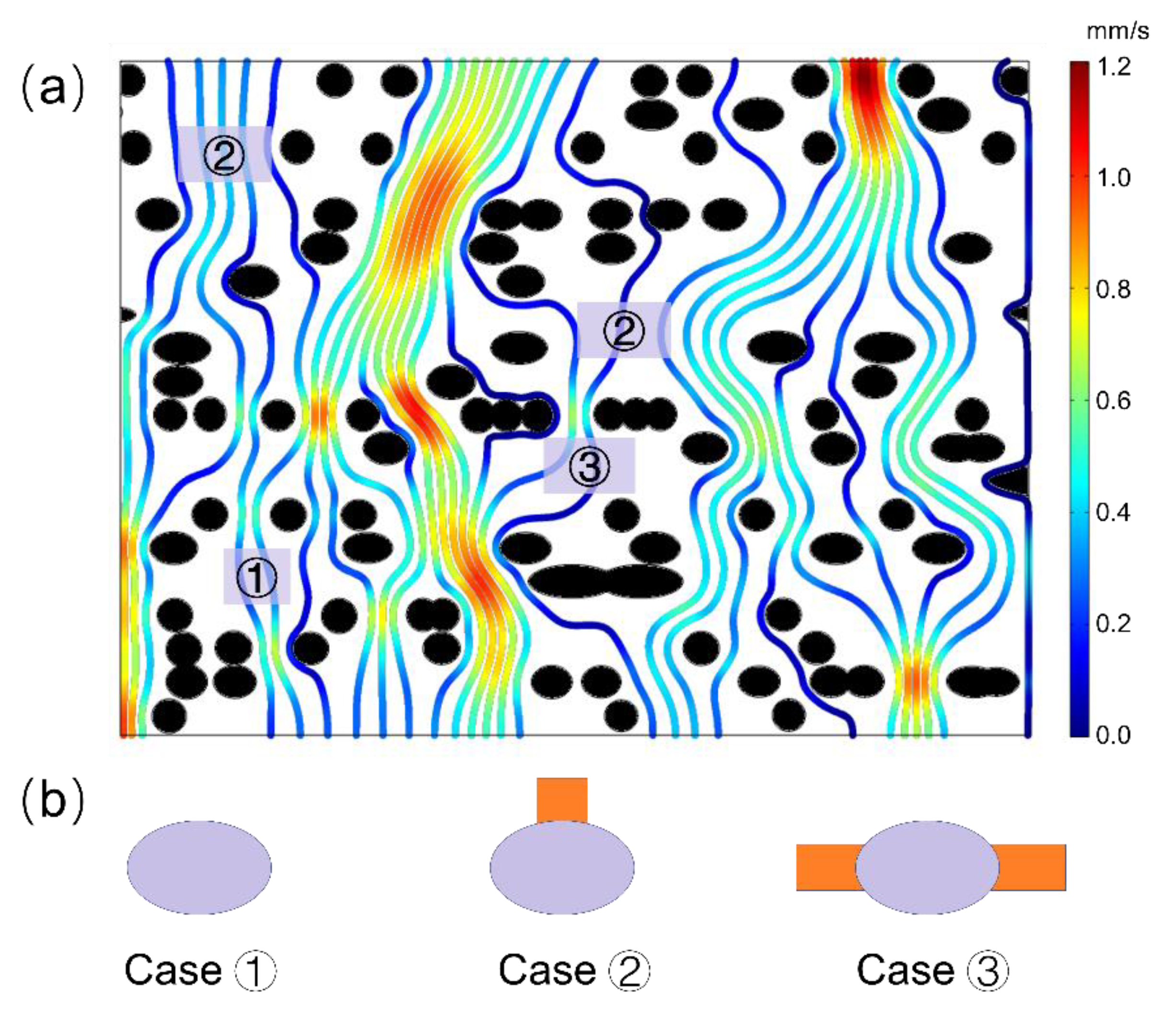
2.3. Microflow Simulation across Intervessel Pit Membrane and Its Implication
2.4. Other Pit Membrane Related Traits and Functional Roles
3. Role of Pit Membrane Chemical Composition in Plant Hydraulics
3.1. Chemical Components of Pit Membranes
3.2. The Ionic Effect and Controversy of Pectin Presence in Pit Membrane
4. Role of Pit Membrane Mechanical Properties in Plant Hydraulics
4.1. Techniques in Characterizing Mechanical Properties of Pit Membrane
4.2. Modelling Mechanical Properties of Pit Membrane
4.3. The Reversibility of Pit Membranes under Environmental Changes
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Dixon, H.H.; Joly, J., II. On the ascent of sap. Proc. R. Soc. Lond. 1895, 57, 3–5. [Google Scholar]
- Tyree, M.T.; Zimmermann, M.H.; Tyree, P.M.T. Xylem structure and the ascent of sap; Springer Science and Business Media: Berlin/Heidelberg, Germany, 2002. [Google Scholar]
- Venturas, M.D.; Sperry, J.S.; Hacke, U.G. Plant xylem hydraulics: What we understand, current research, and future challenges. J. Integr. Plant Boil. 2017, 59, 356–389. [Google Scholar] [CrossRef] [PubMed]
- Li, S.; Lens, F.; Espino, S.; Karimi, Z.; Klepsch, M.; Schenk, H.J.; Schmitt, M.; Schuldt, B.; Jansen, S. Intervessel pit membrane thickness as a key determinant of embolism resistance in angiosperm xylem. IAWA J. 2016, 37, 152–171. [Google Scholar] [CrossRef]
- Sperry, J.S.; Hacke, U.G.; Wheeler, J.K. Comparative analysis of end wall resistivity in xylem conduits. Plant, Cell Environ. 2005, 28, 456–465. [Google Scholar] [CrossRef]
- Tyree, M.T. Plant hydraulics: The ascent of water. Nature 2003, 423, 923. [Google Scholar] [CrossRef] [PubMed]
- Hacke, U.; Sauter, J.J. Xylem dysfunction during winter and recovery of hydraulic conductivity in diffuse-porous and ring-porous trees. Oecologia 1996, 105, 435–439. [Google Scholar] [CrossRef] [PubMed]
- Urli, M.; Porté, A.J.; Cochard, H.; Guengant, Y.; Burlett, R.; Delzon, S. Xylem embolism threshold for catastrophic hydraulic failure in angiosperm trees. Tree Physiol. 2013, 33, 672–683. [Google Scholar] [CrossRef]
- Hammond, W.M.; Yu, K.; Wilson, L.A.; Will, R.E.; Anderegg, W.R.L.; Adams, H.D. Dead or dying? Quantifying the point of no return from hydraulic failure in drought-induced tree mortality. New Phytol. 2019, 223, 1834–1843. [Google Scholar] [CrossRef]
- Sperry, J.S.; Tyree, M.T. Mechanism of water stress-induced xylem embolism. Plant Physiol. 1988, 88, 581–587. [Google Scholar] [CrossRef]
- Brodersen, C.R.; McElrone, A.J.; Choat, B.; Lee, E.F.; Shackel, K.A.; Matthews, M.A. In vivo visualizations of drought-induced embolism spread in Vitis vinifera. Plant Physiol. 2013, 161, 1820–1829. [Google Scholar] [CrossRef]
- Zimmermann, M.H. Hydraulic architecture of some diffuse-porous trees. Can. J. Bot. 1978, 56, 2286–2295. [Google Scholar] [CrossRef]
- Pittermann, J.; Sperry, J.S.; Hacke, U.G.; Wheeler, J.K.; Sikkema, E.H. Inter-tracheid pitting and the hydraulic efficiency of conifer wood: The role of tracheid allometry and cavitation protection. Am. J. Bot. 2006, 93, 1265–1273. [Google Scholar] [CrossRef] [PubMed]
- Brodersen, C.; Jansen, S.; Choat, B.; Rico, C.; Pittermann, J. Cavitation resistance in seedless vascular plants: The structure and function of interconduit pit membranes. Plant Physiol. 2014, 165, 895–904. [Google Scholar] [CrossRef] [PubMed]
- Jansen, S.; Choat, B.; Pletsers, A. Morphological variation of intervessel pit membranes and implications to xylem function in angiosperms. Am. J. Bot. 2009, 96, 409–419. [Google Scholar] [CrossRef]
- Scholz, A.; Rabaey, D.; Stein, A.; Cochard, H.; Smets, E.; Jansen, S. The evolution and function of vessel and pit characters with respect to cavitation resistance across 10 Prunus species. Tree Physiol. 2013, 33, 684–694. [Google Scholar] [CrossRef]
- Cochard, H.; Barigah, S.; Kleinhentz, M.; Eshel, A. Is xylem cavitation resistance a relevant criterion for screening drought resistance among Prunus species? J. Plant Physiol. 2008, 165, 976–982. [Google Scholar] [CrossRef]
- Larter, M.; Pfautsch, S.; Domec, J.; Trueba, S.; Nagalingum, N.; Delzon, S. Aridity drove the evolution of extreme embolism resistance and the radiation of conifer genus Callitris. New Phytol. 2017, 215, 97–112. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.X.; Yu, Z.; Shui, Y.; Yan, Y. Determining the curing parameters of furfuryl alcohol for wood modification by nanoindentation. Eur. J. Wood Wood Prod. 2017, 75, 81–87. [Google Scholar] [CrossRef]
- Schmid, R.; Machado, P. Pit membranes in hard wood-fine structure and development. Protoplasma 1968, 66, 185–204. [Google Scholar] [CrossRef]
- Roth-Nebelsick, A.; Voigt, D.; Gorb, S. Cryo-scanning electron microscopy studies of pits in Pinus Wallichiana and Mallotus Japonicus. IAWA J. 2010, 31, 257–267. [Google Scholar] [CrossRef]
- Plavcová, L.; Jansen, S.; Klepsch, M.; Hacke, U.G. Nobody’s perfect: Can irregularities in pit structure influence vulnerability to cavitation? Front. Plant Sci. 2013, 4, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Liese, W.; Ingeborg, J. Experimenfelle untersuchungen fiber die feinsfruktur der hoftfupfel bei den koniferen. Naturwissenschaften 1954, 41, 579. [Google Scholar] [CrossRef]
- Jansen, S.; Lamy, J.-B.; Burlett, R.; Cochard, H.; Gasson, P.; Delzon, S. Plasmodesmatal pores in the torus of bordered pit membranes affect cavitation resistance of conifer xylem. Plant Cell Environ. 2012, 35, 1109–1120. [Google Scholar] [CrossRef] [PubMed]
- Oda, Y.; Fukuda, H. Initiation of cell wall pattern by a Rho- and Microtubule-driven symmetry breaking. Science 2012, 337, 1333–1336. [Google Scholar] [CrossRef] [PubMed]
- Sasaki, T.; Fukuda, H.; Oda, Y. Cortical microtubule disordering1 is required for secondary cell wall patterning in xylem vessels. Plant Cell 2017, 29, 3123–3139. [Google Scholar] [CrossRef]
- Dória, L.C.; Podadera, D.S.; Lima, R.S.; Lens, F.; Marcati, C.R. Axial sampling height outperforms site as predictor of wood trait variation. IAWA J. 2019, 40, 1–S3. [Google Scholar] [CrossRef]
- Pfautsch, S.; Aspinwall, M.J.; E Drake, J.; Chacon-Doria, L.; A Langelaan, R.J.; Tissue, D.T.; Tjoelker, M.G.; Lens, F. Traits and trade-offs in whole-tree hydraulic architecture along the vertical axis of Eucalyptus grandis. Ann. Bot. 2018, 121, 129–141. [Google Scholar] [CrossRef]
- Schmitz, N.; Koch, G.; Beeckman, H.; Koedam, N.; Robert, E.M.; Schmitt, U. A structural and compositional analysis of intervessel pit membranes in the sapwood of some mangrove woods. IAWA J. 2012, 33, 243–256. [Google Scholar] [CrossRef]
- Li, S.; Li, X.; Link, R.; Li, R.; Deng, L.; Schuldt, B.; Jiang, X.; Zhao, R.; Zheng, J.; Li, S.; et al. Influence of cambial age and axial height on the spatial patterns of xylem traits in Catalpa bungei, a ring-porous tree species native to China. Forests 2019, 10, 662. [Google Scholar] [CrossRef]
- Kotowska, M.M.; Thom, R.; Zhang, Y.; Schenk, H.J.; Jansen, S. Within-tree variability and sample storage effects of bordered pit membranes in xylem of Acer pseudoplatanus. Trees 2019, 34, 61–71. [Google Scholar] [CrossRef]
- Anfodillo, T.; Petit, G.; Crivellaro, A. Axial conduit widening in woody species: A still neglected anatomical pattern. IAWA J. 2013, 34, 352–364. [Google Scholar] [CrossRef]
- Zhong, M.; Castro-Díez, P.; Puyravaud, J.-P.; Sterck, F.J.; Cornelissen, J.H.C. Convergent xylem widening among organs across diverse woody seedlings. New Phytol. 2019, 222, 1873–1882. [Google Scholar] [CrossRef]
- Cai, J.; Tyree, M.T. The impact of vessel size on vulnerability curves: Data and models for within-species variability in saplings of aspen, Populus tremuloides Michx. Plant Cell Environ. 2010, 33, 1059–1069. [Google Scholar] [CrossRef] [PubMed]
- Tyree, M.T.; Cochard, H.; Cruiziat, P.; Sinclair, B.; Ameglio, T. Drought-induced leaf shedding in walnut: Evidence for vulnerability segmentation. Plant Cell Environ. 1993, 16, 879–882. [Google Scholar] [CrossRef]
- Charrier, G.; Torres-Ruiz, J.M.; Badel, E.; Burlett, R.; Choat, B.; Cochard, H.; Delmas, C.E.L.; Domec, J.-C.; Jansen, S.; King, A.; et al. Evidence for hydraulic vulnerability segmentation and lack of xylem refilling under tension. Plant Physiol. 2016, 172, 1657–1668. [Google Scholar] [CrossRef] [PubMed]
- Klepsch, M.; Zhang, Y.; Kotowska, M.M.; Lamarque, L.J.; Nolf, M.; Schuldt, B.; Torres-Ruiz, J.M.; Qin, D.-W.; Choat, B.; Delzon, S.; et al. Is xylem of angiosperm leaves less resistant to embolism than branches? Insights from micro CT, hydraulics, and anatomy. J. Exp. Bot. 2018, 69, 5611–5623. [Google Scholar] [PubMed]
- Pivovaroff, A.L.; Sack, L.; Santiago, L.S. Coordination of stem and leaf hydraulic conductance in southern California shrubs: A test of the hydraulic segmentation hypothesis. New Phytol. 2014, 203, 842–850. [Google Scholar] [CrossRef]
- Scholz, F.G.; Bucci, S.J.; Goldstein, G. Strong hydraulic segmentation and leaf senescence due to dehydration may trigger die-back in Nothofagus dombeyi under severe droughts: A comparison with the co-occurring Austrocedrus chilensis. Trees 2014, 28, 1475–1487. [Google Scholar] [CrossRef]
- Torres-Ruiz, J.M.; Diaz-Espejo, A.; Perez-Martin, A.; Hernandez-Santana, V. Role of hydraulic and chemical signals in leaves, stems and roots in the stomatal behaviour of olive trees under water stress and recovery conditions. Tree Physiol. 2014, 35, 415–424. [Google Scholar] [CrossRef]
- Skelton, R.P.; Anderegg, L.D.L.; Papper, P.; Reich, E.; Dawson, T.E.; Kling, M.; Thompson, S.E.; Diaz, J.; Ackerly, D.D. No local adaptation in leaf or stem xylem vulnerability to embolism, but consistent vulnerability segmentation in a North American oak. New Phytol. 2019, 223, 1296–1306. [Google Scholar] [CrossRef]
- Bouche, P.S.; Delzon, S.; Choat, B.; Badel, E.; Brodribb, T.J.; Burlett, R.; Cochard, H.; Charra-Vaskou, K.; Lavigne, B.; Li, S.; et al. Are needles of Pinus pinaster more vulnerable to xylem embolism than branches? New insights from X-ray computed tomography. Plant Cell Environ. 2016, 39, 860–870. [Google Scholar] [CrossRef] [PubMed]
- Bouche, P.S.; Jansen, S.; Sabalera, J.C.; Cochard, H.; Burlett, R.; Delzon, S. Low intra-tree variability in resistance to embolism in four Pinaceae species. Ann. For. Sci. 2016, 73, 681–689. [Google Scholar] [CrossRef][Green Version]
- Losso, A.; Andreas, B.; Birgit, D.; Dullin, C.; Ganthaler, A.; Petruzzellis, F.; Savi, T.; Tromba, G.; Nardini, A.; Mayr, S.; et al. Insights from in vivo micro-CT analysis: Testing the hydraulic vulnerability segmentation in Acer pseudoplatanus and Fagus sylvatica seedlings. New Phytol. 2019, 221, 1831–1842. [Google Scholar] [CrossRef] [PubMed]
- Alder, N.N.; Sperry, J.S.; Pockman, W.T. Root and stem xylem embolism, stomatal conductance, and leaf turgor in Acer grandidentatum populations along a soil moisture gradient. Oecologia 1996, 105, 293–301. [Google Scholar] [CrossRef]
- Schulte, P.J.; Hacke, U.; Schoonmaker, A.L. Pit membrane structure is highly variable and accounts for a major resistance to water flow through tracheid pits in stems and roots of two boreal conifer species. New Phytol. 2015, 208, 102–113. [Google Scholar] [CrossRef]
- Tixier, A.; Herbette, S.; Jansen, S.; Capron, M.; Tordjeman, P.; Cochard, H.; Badel, E. Modelling the mechanical behaviour of pit membranes in bordered pits with respect to cavitation resistance in angiosperms. Ann. Bot. 2014, 114, 325–334. [Google Scholar] [CrossRef]
- Schmitz, N.; Koch, G.; Schmitt, U.; Beeckman, H.; Koedam, N. Intervessel pit structure and histochemistry of two mangrove species as revealed by cellular UV microspectrophotometry and electron microscopy: Intraspecific variation and functional significance. Microsc. Microanal. 2008, 14, 387–397. [Google Scholar] [CrossRef]
- Awad, H.; Barigah, T.; Badel, E.; Cochard, H.; Herbette, S. Poplar vulnerability to xylem cavitation acclimates to drier soil conditions. Physiol. Plant. 2010, 139, 280–288. [Google Scholar] [CrossRef]
- Zhang, Y.; Klepsch, M.; Jansen, S. Bordered pits in xylem of vesselless angiosperms and their possible misinterpretation as perforation plates. Plant Cell Environ. 2017, 40, 2133–2146. [Google Scholar] [CrossRef]
- Jansen, S.; Schenk, H.J. On the ascent of sap in the presence of bubbles. Am. J. Bot. 2015, 102, 1561–1563. [Google Scholar] [CrossRef]
- Schenk, H.J.; Espino, S.; Romo, D.M.; Nima, N.; Do, A.Y.T.; Michaud, J.M.; Papahadjopoulos-sternberg, B.; Yang, J.; Zuo, Y.Y.; Steppe, K.; et al. Xylem surfactants introduce a new element to the cohesion-tension theory. Plant Physiol. 2017, 173, 1177–1196. [Google Scholar] [CrossRef] [PubMed]
- Schenk, H.J.; Espino, S.; Sarah, M.R.-C.; Jansen, S. From the sap’s perspective: The nature of vessel surfaces in angiosperm xylem. Am. J. Bot. 2018, 105, 172–185. [Google Scholar] [CrossRef] [PubMed]
- Kaack, L.; Altaner, C.M.; Carmesin, C.; Diaz, A.; Holler, M.; Kranz, C.; Neusser, G.; Odstrcil, M.; Schenk, H.J.; Schmidt, V.; et al. Function and three-dimensional structure of intervessel pit membranes in angiosperms: A review. IAWA J. 2019, 40, 673–702. [Google Scholar] [CrossRef]
- Fujii, T.; Lee, S.-J.; Kuroda, N.; Suzuki, Y. Conductive function of intervessel pits through a growth ring boundary of Machilus thunbergii. IAWA J. 2001, 22, 1–14. [Google Scholar] [CrossRef]
- Capron, M.; Tordjeman, P.; Charru, F.; Badel, E.; Cochard, H. Gas flow in plant microfluidic networks controlled by capillary valves. Phys. Rev. E 2014, 89, 033019. [Google Scholar] [CrossRef]
- Maschek, D.; Goodell, B.; Jellison, J.; Lessard, M.; Militz, H. A new approach for the study of the chemical composition of bordered pit membranes: 4Pi and confocal laser scanning microscopy. Am. J. Bot. 2013, 100, 1751–1756. [Google Scholar] [CrossRef]
- Zhang, Y.; Carmesin, C.; Kaack, L.; Klepsch, M.M.; Kotowska, M.; Matei, T.; Schenk, H.J.; Weber, M.; Walther, P.; Schmidt, V.; et al. High porosity with tiny pore constrictions and unbending pathways characterize the 3D structure of intervessel pit membranes in angiosperm xylem. Plant Cell Environ. 2019, 43, 116–130. [Google Scholar] [CrossRef]
- Li, Y.; Yu, Z.; Reese, S.; Simon, J.-W. Evaluation of the out-of-plane response of fiber networks with a representative volume element model. Tappi Journal 2018, 17, 329–339. [Google Scholar] [CrossRef]
- Li, X.; Ji, G.; Chen, X.; Ding, W.; Sun, L.; Xu, W.; Han, H.; Sun, F. Large scale three-dimensional reconstruction of an entire Caenorhabditis elegans larva using Auto CUTS-SEM. J. Struct. Boil. 2017, 200, 87–96. [Google Scholar] [CrossRef]
- Choat, B.; Jansen, S.; Zwieniecki, M.A.; Smets, E.; Holbrook, N.M. Changes in pit membrane porosity due to deflection and stretching: The role of vestured pits. J. Exp. Bot. 2004, 55, 1569–1575. [Google Scholar] [CrossRef]
- Pittermann, J.; Sperry, J.S.; Hacke, U.G.; Wheeler, J.K.; Sikkema, E.H. Torus-margo pits help conifers compete with angiosperms. Science 2005, 310, 1924. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Pittermann, J. New insights into bordered pit structure and cavitation resistance in angiosperms and conifers. New Phytol. 2009, 182, 557–560. [Google Scholar] [CrossRef] [PubMed]
- Pittermann, J.; Choat, B.; Jansen, S.; Stuart, S.A.; Lynn, L.; Dawson, T.E. The relationships between xylem safety and hydraulic efficiency in the Cupressaceae: The evolution of pit membrane form and function. Plant Physiol. 2010, 153, 1919–1931. [Google Scholar] [CrossRef] [PubMed]
- Choat, B.; Jansen, S.; Brodribb, T.J.; Cochard, H.; Delzon, S.; Bhaskar, R.; Bucci, S.J.; Feild, T.S.; Gleason, S.M.; Hacke, U.G.; et al. Global convergence in the vulnerability of forests to drought. Nature 2012, 491, 752–755. [Google Scholar] [CrossRef]
- Christman, M.A.; Sperry, J.S.; Adler, F.R. Testing the ‘rare pit’ hypothesis for xylem cavitation resistance in three species of Acer. New Phytol. 2009, 182, 664–674. [Google Scholar] [CrossRef]
- Christman, M.A.; Sperry, J.S.; Smith, D.D. Rare pits, large vessels and extreme vulnerability to cavitation in a ring-porous tree species. New Phytol. 2011, 193, 713–720. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S. Functional and ecological xylem anatomy. Perspect. Plant Ecol. Evol. Syst. 2001, 4, 97–115. [Google Scholar] [CrossRef]
- Choat, B.; Ball, M.; Luly, J.; Holtum, J. Pit membrane porosity and water stress-induced cavitation in four co-existing dry rainforest tree species. Plant Physiol. 2003, 131, 41–48. [Google Scholar] [CrossRef]
- Roskilly, B.; Keeling, E.; Hood, S.; Giuggiola, A.; Sala, A. Conflicting functional effects of xylem pit structure relate to the growth-longevity trade-off in a conifer species. Proc. Natl. Acad. Sci. USA 2019, 116, 15282–15287. [Google Scholar] [CrossRef]
- Valli, A.; Koponen, A.; Vesala, T.; Timonen, J. Simulations of water flow through bordered pits of conifer xylem. J. Stat. Phys. 2002, 107, 121–142. [Google Scholar] [CrossRef]
- Schulte, P.J. Computational fluid dynamics models of conifer bordered pits show how pit structure affects flow. New Phytol. 2012, 193, 721–729. [Google Scholar] [CrossRef] [PubMed]
- Bouche, P.S.; Larter, M.; Domec, J.-C.; Burlett, R.; Gasson, P.; Jansen, S.; Delzon, S. A broad survey of hydraulic and mechanical safety in the xylem of conifers. J. Exp. Bot. 2014, 65, 4419–4431. [Google Scholar] [CrossRef] [PubMed]
- Jansen, S.; McAdam, S. Pits with aspiration explain life expectancy of a conifer species. Proc. Natl. Acad. Sci. USA 2019, 116, 14794–14796. [Google Scholar] [CrossRef]
- Plavcová, L.; Hacke, U. Heterogeneous distribution of pectin epitopes and calcium in different pit types of four angiosperm species. New Phytol. 2011, 192, 885–897. [Google Scholar] [CrossRef] [PubMed]
- Wheeler, J.K.; Sperry, J.S.; Hacke, U.G.; Hoang, N. Inter-vessel pitting and cavitation in woody Rosaceae and other vesselled plants: A basis for a safety versus efficiency trade-off in xylem transport. Plant Cell Environ. 2005, 28, 800–812. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Feild, T.S.; Sano, Y.; Sikkema, E.H.; Pittermann, J. Water transport in vesselless angiosperms: Conducting efficiency and cavitation safety. Int. J. Plant Sci. 2007, 168, 1113–1126. [Google Scholar] [CrossRef]
- Hacke, U.G.; Sperry, J.S.; Wheeler, J.K.; Castro, L. Scaling of angiosperm xylem structure with safety and efficiency. Tree Physiol. 2006, 26, 689–701. [Google Scholar] [CrossRef]
- Lens, F.; Sperry, J.S.; Christman, M.A.; Choat, B.; Rabaey, D.; Jansen, S. Testing hypotheses that link wood anatomy to cavitation resistance and hydraulic conductivity in the genus Acer. New Phytol. 2011, 190, 709–723. [Google Scholar] [CrossRef]
- Dusotoit-Coucaud, A.; Brunel, N.; Tixier, A.; Cochard, H.; Herbette, S. Hydrolase treatments help unravel the function of intervessel pits in xylem hydraulics. Physiol. Plant. 2014, 150, 388–396. [Google Scholar] [CrossRef]
- Boyce, C.K.; Zwieniecki, M.A.; Cody, G.D.; Jacobsen, C.; Wirick, S.; Knoll, A.H.; Holbrook, N.M. Evolution of xylem lignification and hydrogel transport regulation. Proc. Natl. Acad. Sci. USA 2004, 101, 17555–17558. [Google Scholar] [CrossRef]
- Pereira, L.; Domingues-Junior, A.P.; Jansen, S.; Choat, B.; Mazzafera, P. Is embolism resistance in plant xylem associated with quantity and characteristics of lignin? Trees 2017, 32, 349–358. [Google Scholar] [CrossRef]
- Higuchi, T. Biochemistry and Molecular Biology of Wood; Springer Science and Business Media: Berlin/Heidelberg, Germany, 1997; pp. 168–174. [Google Scholar]
- Fromm, J.; Rockel, B.; Lautner, S.; Windeisen, E.; Wanner, G. Lignin distribution in wood cell walls determined by TEM and backscattered SEM techniques. J. Struct. Boil. 2003, 143, 77–84. [Google Scholar] [CrossRef]
- Herbette, S.; Bouchet, B.; Brunel, N.; Bonnin, E.; Guillon, F. Immunolabelling of intervessel pits for polysaccharides and lignin helps in understanding their hydraulic properties in Populus tremula x alba. Ann. Bot. 2015, 115, 187–199. [Google Scholar] [CrossRef] [PubMed]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef]
- van Doorn, W.G.; Hiemstra, T.; Fanourakis, D. Hydrogel regulation of xylem water flow: An alternative hypothesis. Plant Physiol. 2011, 157, 1642–1649. [Google Scholar] [CrossRef]
- West, M.; Vaidya, A.; Singh, A.P. Correlative light and scanning electron microscopy of the same sections gives new insights into the effects of pectin lyase on bordered pit membranes in Pinus radiata wood. Micron 2012, 43, 916–919. [Google Scholar] [CrossRef]
- Mohnen, D. Pectin structure and biosynthesis. Curr. Opin. Plant Boil. 2008, 11, 266–277. [Google Scholar] [CrossRef]
- Allario, T.; Tixier, A.; Awad, H.; Lemaire, C.; Brunel, N.; Badel, E. Ptxtpme1\r and homogalacturonans influence xylem hydraulic. Physiol. Plant. 2018, 163, 502–515. [Google Scholar] [CrossRef]
- Yoneda, A.; Ito, T.; Higaki, T.; Kutsuna, N.; Saito, T.; Ishimizu, T.; Osada, H.; Hasezawa, S.; Matsui, M.; Demura, T. Cobtorin target analysis reveals that pectin functions in the deposition of cellulose microfibrils in parallel with cortical microtubules. Plant J. 2010, 64, 657–667. [Google Scholar] [CrossRef]
- Pereira, L.; Flores-Borges, D.N.; Bittencourt, P.R.; Mayer, J.L.; Kiyota, E.; Araújo, P.; Jansen, S.; Freitas, R.O.; Oliveira, R.S.; Mazzafera, P. Infrared nanospectroscopy reveals the chemical nature of pit membranes in water-conducting cells of the plant xylem. Plant Physiol. 2018, 177, 1629–1638. [Google Scholar] [CrossRef]
- Bauch, J.; Berndt, H. Variability of the chemical composition of pit membranes in bordered pits of gymnosperms. Wood Sci. Technol. 1973, 7, 6–19. [Google Scholar] [CrossRef]
- Wallis, C.M.; Chen, J. Grapevine phenolic compounds in xylem sap and tissues are significantly altered during infection by Xylella fastidiosa. Phytopathol. 2012, 102, 816–826. [Google Scholar] [CrossRef] [PubMed]
- Schenk, H.J.; Steppe, K.; Jansen, S. Nanobubbles: A new paradigm for air-seeding in xylem. Trends Plant Sci. 2015, 20, 199–205. [Google Scholar] [CrossRef] [PubMed]
- Hu, F.; Shi, L.; Min, W. Biological imaging of chemical bonds by stimulated Raman scattering microscopy. Nat. Methods 2019, 16, 830–842. [Google Scholar] [CrossRef]
- Zhao, Y.; Lin, S.; Qiu, Z.; Cao, D.; Wen, J.; Deng, X.; Wang, X.; Lin, J.; Li, X. MicroRNA857 Is involved in the regulation of secondary growth of vascular tissues in Arabidopsis. Plant Physiol. 2015, 169, 2539–2552. [Google Scholar] [CrossRef]
- Zhou, Q.; Ren, Y.; Du, Y.; Hua, D.; Han, W.; Xia, Q. Adhesion energy and related plastic deformation mechanism of Cu/Ru nanostructured multilayer film. J. Alloy. Compd. 2019, 772, 823–827. [Google Scholar] [CrossRef]
- Zhu, N.; Yang, Y.; Ji, M.; Wu, D.; Chen, K. Label-free visualization of lignin deposition in loquats using complementary stimulated and spontaneous Raman microscopy. Hortic. Res. 2019, 6, 72. [Google Scholar] [CrossRef]
- Trifilò, P.; Gullo, M.A.L.; Salleo, S.; Callea, K.; Nardini, A. Xylem embolism alleviated by ion-mediated increase in hydraulic conductivity of functional xylem: Insights from field measurements. Tree Physiol. 2008, 28, 1505–1512. [Google Scholar] [CrossRef] [PubMed]
- Cochard, H.; Herbette, S.; Hernández, E.; Hölttä, T.; Mencuccini, M. The effects of sap ionic composition on xylem vulnerability to cavitation. J. Exp. Bot. 2010, 61, 275–285. [Google Scholar] [CrossRef] [PubMed]
- Nardini, A.; DiMasi, F.; Klepsch, M.; Jansen, S. Ion-mediated enhancement of xylem hydraulic conductivity in four Acer species: Relationships with ecological and anatomical features. Tree Physiol. 2012, 32, 1434–1441. [Google Scholar] [CrossRef] [PubMed]
- Nardini, A.; Grego, F.; Trifilò, P.; Salleo, S. Changes of xylem sap ionic content and stem hydraulics in response to irradiance in Laurus nobilis. Tree Physiol. 2010, 30, 628–635. [Google Scholar] [CrossRef] [PubMed]
- Nardini, A.; Salleo, S.; Jansen, S. More than just a vulnerable pipeline: Xylem physiology in the light of ion-mediated regulation of plant water transport. J. Exp. Bot. 2011, 62, 4701–4718. [Google Scholar] [CrossRef] [PubMed]
- Jansen, S.; Gortan, E.; Lens, F.; Lo Gullo, M.A.; Salleo, S.; Scholz, A.; Stein, A.; Trifilò, P.; Nardini, A. Do quantitative vessel and pit characters account for ion-mediated changes in the hydraulic conductance of angiosperm xylem? New Phytol. 2011, 189, 218–228. [Google Scholar] [CrossRef] [PubMed]
- Casolo, V.; Tomasella, M.; De Col, V.; Braidot, E.; Savi, T.; Nardini, A. Water relations of an invasive halophyte (Spartina patens): Osmoregulation and ionic effects on xylem hydraulics. Funct. Plant Boil. 2015, 42, 264–273. [Google Scholar] [CrossRef]
- Zwieniecki, M.A. Hydrogel control of xylem hydraulic resistance in plants. Science 2001, 291, 1059–1062. [Google Scholar] [CrossRef]
- Gortan, E.; Nardini, A.; Salleo, S.; Jansen, S. Pit membrane chemistry influences the magnitude of ion-mediated enhancement of xylem hydraulic conductance in four Lauraceae species. Tree Physiol. 2011, 31, 48–58. [Google Scholar] [CrossRef][Green Version]
- Santiago, M.; Pagay, V.; Stroock, A.D. Impact of electroviscosity on the hydraulic conductance of the bordered pit membrane: A theoretical investigation1. Plant Physiol. 2013, 163, 999–1011. [Google Scholar] [CrossRef]
- Gascó, A.; Salleo, S.; Gortan, E.; Nardini, A. Seasonal changes in the ion-mediated increase of xylem hydraulic conductivity in stems of three evergreens: Any functional role? Physiol. Plant. 2007, 129, 597–606. [Google Scholar] [CrossRef]
- Nardini, A.; Antonio, G.; Trifilo, P.; Gullo, M.A.L.; Salleo, S. Ion-mediated enhancement of xylem hydraulic conductivity is not always suppressed by the presence of Ca2+ in the sap. J. Exp. Bot. 2007, 58, 2609–2615. [Google Scholar] [CrossRef]
- Herbette, S.; Cochard, H. Calcium is a major determinant of xylem vulnerability to cavitation. Plant Physiol. 2010, 153, 1932–1939. [Google Scholar] [CrossRef]
- Van Ieperen, W.; Van Meeteren, U.; Van Gelder, H. Fluid ionic composition influences hydraulic conductance of xylem conduits. J. Exp. Bot. 2000, 51, 769–776. [Google Scholar] [CrossRef] [PubMed]
- Jupa, R.; Doubková, P.; Gloser, V. Ion-mediated increases in xylem hydraulic conductivity: Seasonal differences between coexisting ring- and diffuse-porous temperate tree species. Tree Physiol. 2019, 39, 1313–1328. [Google Scholar] [CrossRef] [PubMed]
- Jansen, S.; Choat, B.; Vinckier, S.; Lens, F.; Schols, P.; Smets, E. Intervascular pit membranes with a torus in the wood of Ulmus (Ulmaceae) and related genera. New Phytol. 2004, 163, 51–59. [Google Scholar] [CrossRef]
- Klepsch, M.M.; Schmitt, M.; Knox, J.P.; Jansen, S. The chemical identity of intervessel pit membranes in Acer challenges hydrogel control of xylem hydraulic conductivity. AoB Plants 2016, 8, 1–14. [Google Scholar] [CrossRef]
- Kulasinski, K.; Guyer, R.; Keten, S.; Derome, D.; Carmeliet, J. Impact of moisture adsorption on structure and physical properties of amorphous biopolymers. Macromolecules 2015, 48, 2793–2800. [Google Scholar] [CrossRef]
- Zelinka, S.L.; Bourne, K.J.; Hermanson, J.C.; Glass, S.V.; Costa, A.; Wiedenhoeft, A.C. Force-displacement measurements of earlywood bordered pits using a mesomechanical tester. Plant Cell Environ. 2015, 38, 2088–2097. [Google Scholar] [CrossRef]
- Cox, H.L. The elasticity and strength of paper and other fibrous materials. Br. J. Appl. Phys. 1952, 3, 72–79. [Google Scholar] [CrossRef]
- Mayr, S.; Schmid, P.; Laur, J.; Rosner, S.; Charra-Vaskou, K.; Dämon, B.; Hacke, U.G. Uptake of water via branches helps timberline conifers refill embolized xylem in late winter. Plant Physiol. 2014, 164, 1731–1740. [Google Scholar] [CrossRef]
- Brodersen, C.R.; Roddy, A.B.; Wason, J.W.; McElrone, A.J. Functional status of xylem through time. Annu. Rev. Plant Boil. 2019, 70, 407–433. [Google Scholar] [CrossRef]





© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, S.; Wang, J.; Yin, Y.; Li, X.; Deng, L.; Jiang, X.; Chen, Z.; Li, Y. Investigating Effects of Bordered Pit Membrane Morphology and Properties on Plant Xylem Hydraulic Functions—A Case Study from 3D Reconstruction and Microflow Modelling of Pit Membranes in Angiosperm Xylem. Plants 2020, 9, 231. https://doi.org/10.3390/plants9020231
Li S, Wang J, Yin Y, Li X, Deng L, Jiang X, Chen Z, Li Y. Investigating Effects of Bordered Pit Membrane Morphology and Properties on Plant Xylem Hydraulic Functions—A Case Study from 3D Reconstruction and Microflow Modelling of Pit Membranes in Angiosperm Xylem. Plants. 2020; 9(2):231. https://doi.org/10.3390/plants9020231
Chicago/Turabian StyleLi, Shan, Jie Wang, Yafang Yin, Xin Li, Liping Deng, Xiaomei Jiang, Zhicheng Chen, and Yujun Li. 2020. "Investigating Effects of Bordered Pit Membrane Morphology and Properties on Plant Xylem Hydraulic Functions—A Case Study from 3D Reconstruction and Microflow Modelling of Pit Membranes in Angiosperm Xylem" Plants 9, no. 2: 231. https://doi.org/10.3390/plants9020231
APA StyleLi, S., Wang, J., Yin, Y., Li, X., Deng, L., Jiang, X., Chen, Z., & Li, Y. (2020). Investigating Effects of Bordered Pit Membrane Morphology and Properties on Plant Xylem Hydraulic Functions—A Case Study from 3D Reconstruction and Microflow Modelling of Pit Membranes in Angiosperm Xylem. Plants, 9(2), 231. https://doi.org/10.3390/plants9020231