Molecular Characterization of the Dwarf53 Gene Homolog in Dasypyrum Villosum

 , , and
, , and
Abstract
1. Introduction
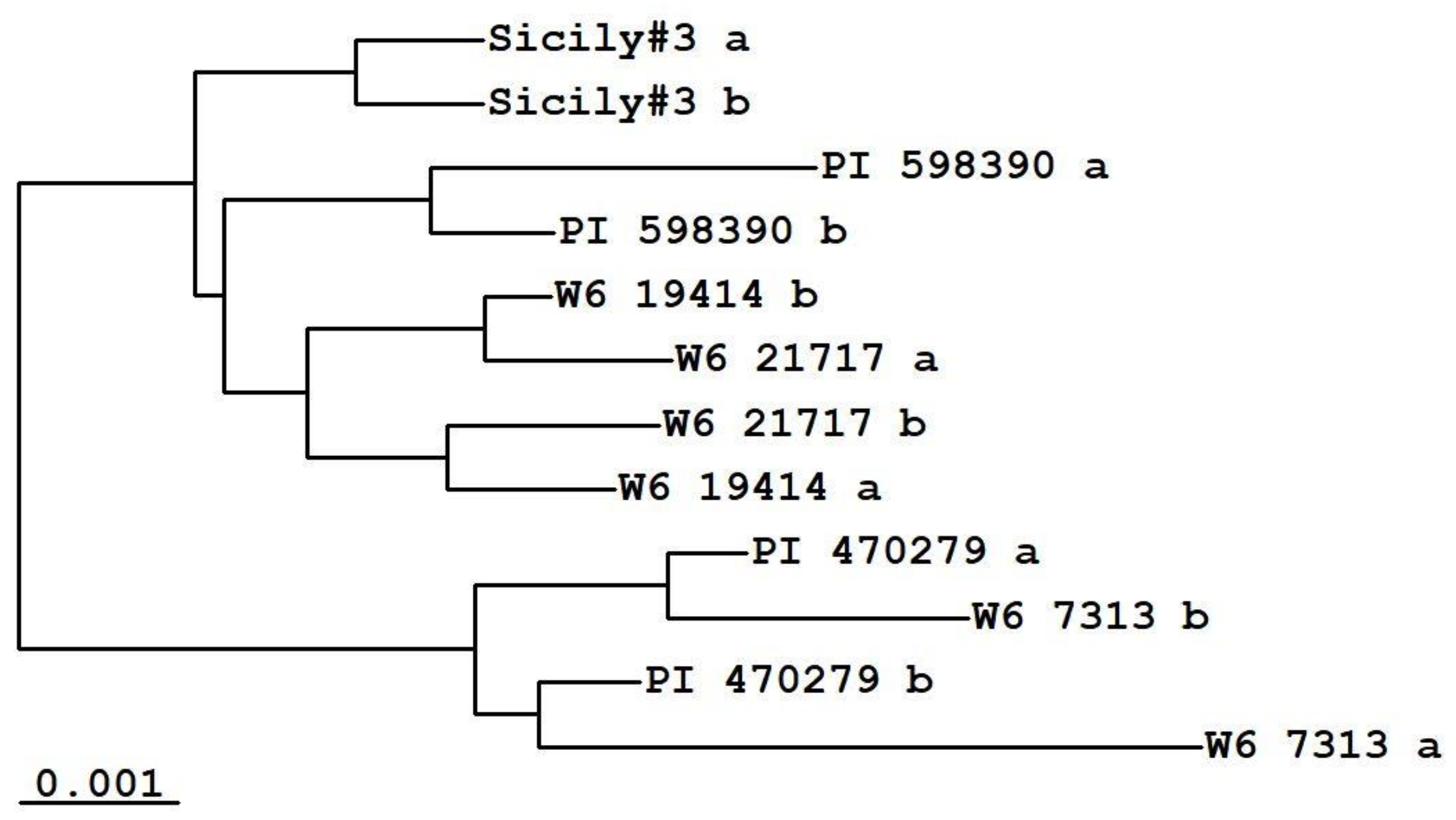
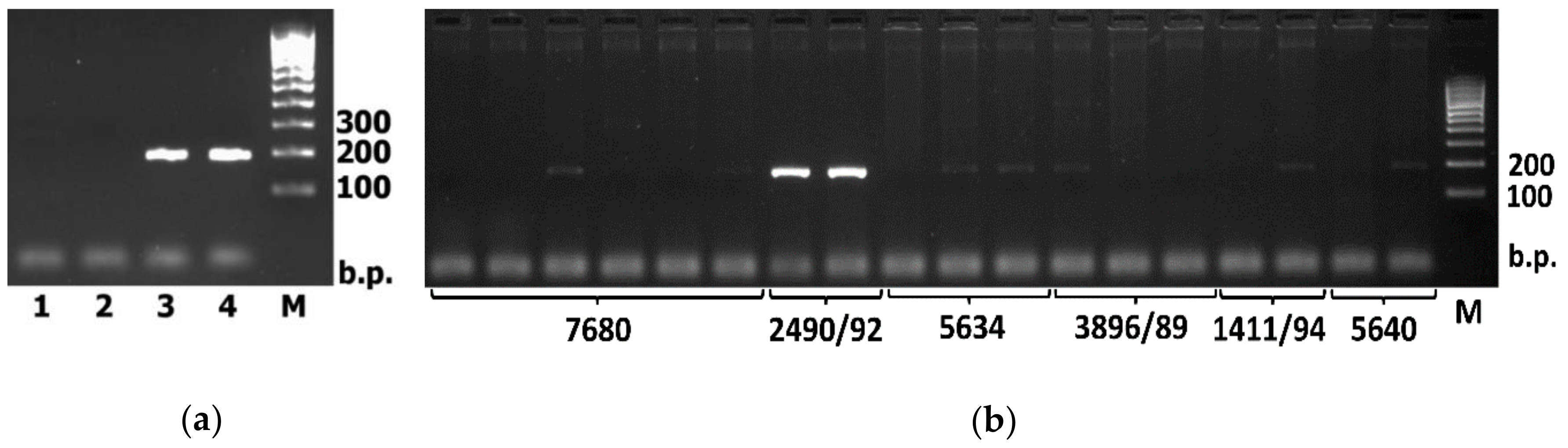
2. Results
3. Discussion
4. Materials and Methods
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Iwata, N. Linkage studies in rice (Oryza sativa L.). Linkage groups for six genes newly described. Jpn. J. Breed. 1977, 27, 250–251. [Google Scholar]
- Wei, L.-R.; Xu, J.-C.; Li, X.-B.; Qian, Q.; Zhu, L.-H. Genetic Analysis and Mapping of the Dominant Dwarfing Gene D-53 in Rice. J. Integr. Plant Biol. 2006, 48, 447–452. [Google Scholar] [CrossRef]
- Jiang, L.; Liu, X.; Xiong, G.; Liu, H.; Chen, F.; Wang, L.; Meng, X.; Liu, G.; Yu, H.; Yuan, Y.; et al. DWARF 53 acts as a repressor of strigolactone signalling in rice. Nature 2013, 504, 401–405. [Google Scholar] [CrossRef] [PubMed]
- Zhou, F.; Lin, Q.; Zhu, L.; Ren, Y.; Zhou, K.; Shabek, N.; Wu, F.; Mao, H.; Dong, W.; Gan, L.; et al. D14-SCF(D3)-dependent degradation of D53 regulates strigolactone signalling. Nature 2013, 504, 406–410. [Google Scholar] [CrossRef]
- Ruyter-Spira, C.; Al-Babili, S.; Van der Krol, S.; Bouwmeester, H. The biology of strigolactones. Trends Plant Sci. 2013, 18, 72–83. [Google Scholar] [CrossRef] [PubMed]
- Navabi, A.; Iqbal, M.; Strenzke, K.; Spaner, D. The relationship between lodging and plant height in a diverse wheat population. Can. J. Plant Sci. 2006, 86, 723–726. [Google Scholar] [CrossRef]
- Xie, Q.; Mayes, S.; Sparkes, D.L. Optimizing tiller production and survival for grain yield improvement in a bread wheat × spelt mapping population. Ann. Bot. 2016, 117, 51–66. [Google Scholar] [CrossRef] [PubMed]
- Liao, M.; Palta, J.; Fillery, I. Root characteristics of vigorous wheat improve early nitrogen uptake. Aust. J. Agric. Res. 2006, 57, 1097–1107. [Google Scholar] [CrossRef]
- Qin, L.; Zhao, J.; Li, T.; Hou, J.; Zhang, X.; Hao, C. TaGW2, a Good Reflection of Wheat Polyploidization and Evolution. Front. Plant Sci. 2017, 8, 318. [Google Scholar] [CrossRef]
- Li, S.; Wang, J.; Wang, K.; Chen, J.; Wang, K.; Du, L.; Ni, Z.; Lin, Z.; Ye, X. Development of PCR markers specific to Dasypyrum villosum genome based on transcriptome data and their application in breeding Triticum aestivum-D. villosum#4 alien chromosome lines. BMC Genomics 2019, 20, 289. [Google Scholar]
- Tajima, F.; Nei, M. Estimation of evolutionary distance between nucleotide sequences. Mol. Biol. Evol. 1984, 1, 269–285. [Google Scholar] [PubMed]
- URGI BLAST. Available online: https://urgi.versailles.inra.fr/blast/ (accessed on 22 December 2019).
- Grądzielewska, A. The genus Dasypyrum—part 2. Dasypyrum villosum—a wild species used in wheat improvement. Euphytica 2006, 152, 441–454. [Google Scholar]
- Sears, E.R. Addition of the Genome of Haynaldia villosa to Triticum aestivum. Am. J. Bot. 1953, 40, 168–174. [Google Scholar] [CrossRef]
- Bie, T.; Zhao, R.; Zhu, S.; Chen, S.; Cen, B.; Zhang, B.; Gao, D.; Jiang, Z.; Chen, T.; Wang, L.; et al. Development and characterization of marker MBH1 simultaneously tagging genes Pm21 and PmV conferring resistance to powdery mildew in wheat. Mol. Breed. 2015, 35, 189. [Google Scholar] [CrossRef]
- Zhang, Q.; Li, Q.; Wang, X.; Wang, H.; Lang, S.; Wang, Y.; Wang, S.; Chen, P.; Liu, D. Development and characterization of a Triticum aestivum-Haynaldia villosa translocation line T4VS⋅4DL conferring resistance to wheat spindle streak mosaic virus. Euphytica 2005, 145, 317–320. [Google Scholar] [CrossRef]
- Zhang, R.; Fan, Y.; Kong, L.; Wang, Z.; Wu, J.; Xing, L.; Cao, A.; Feng, Y. Pm62, an adult-plant powdery mildew resistance gene introgressed from Dasypyrum villosum chromosome arm 2VL into wheat. Theor. Appl. Genet. 2018, 131, 2613–2620. [Google Scholar] [CrossRef]
- Zhang, R.; Sun, B.; Chen, J.; Cao, A.; Xing, L.; Feng, Y.; Lan, C.; Chen, P. Pm55, a developmental-stage and tissue-specific powdery mildew resistance gene introgressed from Dasypyrum villosum into common wheat. Theor. Appl. Genet. 2016, 129, 1975–1984. [Google Scholar] [CrossRef]
- Qi, L.L.; Pumphrey, M.O.; Friebe, B.; Zhang, P.; Qian, C.; Bowden, R.L.; Rouse, M.N.; Jin, Y.; Gill, B.S. A novel Robertsonian translocation event leads to transfer of a stem rust resistance gene (Sr52) effective against race Ug99 from Dasypyrum villosum into bread wheat. Theor. Appl. Genet. 2011, 123, 159–167. [Google Scholar] [CrossRef]
- Zhang, R.; Mingyi, Z.; Xiue, W.; Peidu, C. Introduction of chromosome segment carrying the seed storage protein genes from chromosome 1V of Dasypyrum villosum showed positive effect on bread-making quality of common wheat. Theor. Appl. Genet. 2014, 127, 523–533. [Google Scholar]
- Zhang, R.; Hou, F.; Feng, Y.; Zhang, W.; Zhang, M.; Chen, P. Characterization of a Triticum aestivum-Dasypyrum villosum T2VS·2DL translocation line expressing a longer spike and more kernels traits. Theor. Appl. Genet. 2015, 128, 2415–2425. [Google Scholar] [CrossRef]
- Djanaguiraman, M.; Prasad, P.V.V.; Kumari, J.; Sehgal, S.K.; Friebe, B.; Djalovic, I.; Chen, Y.; Siddique, K.H.M.; Gill, B.S. Alien chromosome segment from Aegilops speltoides and Dasypyrum villosum increases drought tolerance in wheat via profuse and deep root system. BMC Plant Biol. 2019, 19, 242. [Google Scholar] [CrossRef] [PubMed]
- Doyle, P.J. DNA Protocols for Plants. In Molecular Techniques in Taxonomy; Hewitt, G.M., Johnston, A.W.B., Young, J.P.W., Eds.; NATO ASI Series; Springer: Berlin/Heidelberg, Germany, 1991; pp. 283–293. ISBN 978-3-642-83964-1. [Google Scholar]
- BLAST–WHEAT URGI. Available online: https://wheat-urgi.versailles.inra.fr/Seq-Repository/BLAST (accessed on 19 December 2019).
- Primer designing tool. Available online: https://www.ncbi.nlm.nih.gov/tools/primer-blast/ (accessed on 19 December 2019).
- SantaLucia, J. A unified view of polymer, dumbbell and oligonucleotide DNA nearest-neighbor thermodynamics. Proc. Natl. Acad. Sci. USA 1998, 95, 1460–1465. [Google Scholar] [CrossRef] [PubMed]
- Babraham Bioinformatics–FastQC A Quality Control tool for High Throughput Sequence Data. Available online: https://www.bioinformatics.babraham.ac.uk/projects/fastqc/ (accessed on 19 December 2019).
- Bankevich, A.; Nurk, S.; Antipov, D.; Gurevich, A.A.; Dvorkin, M.; Kulikov, A.S.; Lesin, V.M.; Nikolenko, S.I.; Pham, S.; Prjibelski, A.D.; et al. SPAdes: a new genome assembly algorithm and its applications to single-cell sequencing. J. Comput. Biol. 2012, 19, 455–477. [Google Scholar] [CrossRef] [PubMed]
- GeneStudio.com | Free molecular biology software. Available online: http://genestudio.com/ (accessed on 19 December 2019).
- GitHub–MikhailBazhenov/Transform-RTF-to-FASTA. Available online: https://github.com/MikhailBazhenov/Transform-RTF-to-FASTA (accessed on 19 December 2019).
- Nicholas, K.B.; Nikolas, H.B. Jr. GeneDoc: A tool for editing and annotating multiple sequence alignments. 1997. Available online: https://genedoc.software.informer.com/download/ (accessed on 19 December 2019).
- Zaharia, M.; Bolosky, W.J.; Curtis, K.; Fox, A.; Patterson, D.; Shenker, S.; Stoica, I.; Karp, R.M.; Sittler, T. Faster and More Accurate Sequence Alignment with SNAP. arXiv 2011, arXiv:1111.5572. Available online: https://arxiv.org/abs/1111.5572 (accessed on 19 December 2019).
- Garrison, E.; Marth, G. Haplotype-based variant detection from short-read sequencing. arXiv 2012, arXiv:1207.3907. Available online: https://arxiv.org/abs/1207.3907 (accessed on 19 December 2019).
- GitHub–samtools/bcftools. Available online: https://github.com/samtools/bcftools (accessed on 19 December 2019).
- PHYLIP Home Page. Available online: http://evolution.genetics.washington.edu/phylip.html (accessed on 19 December 2019).
- Stanke, M. The AUGUSTUS gene prediction tool. Available online: http://bioinf.uni-greifswald.de/augustus/ (accessed on 19 December 2019).
- Choi, Y.; Chan, A.P. PROVEAN web server: A tool to predict the functional effect of amino acid substitutions and indels. Bioinformatics 2015, 31, 2745–2747. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Accession | Place of Collection | Source |
|---|---|---|
| W6 19414 | Bulgaria | W6 |
| W6 7313 | Greece | W6 |
| W6 21717 | Crimea | W6 |
| PI 598390 | former USSR | W6 |
| PI 470279 | Turkey | W6 |
| Sicily#3 | Italy, Sicily | AJL |
| Primer Pairs, Sequence Direction is 5’ -> 3’ | Tm | Expected Amplicon Size for Wheat Subgenomes | ||
|---|---|---|---|---|
| A | B | D | ||
| D53-F1: CGTGGTTTATAAGCAAGCAATCCA D53-R1: GGGGAACTTGGACAGGAAGG | 60 | 1239 | 1281 | 1263 |
| D53-F2: TACCTCACCTTCCTGTCCAAGTT D53-R2: TCTTACCCTTTTCATCAAGCTGT | 58 | 1619 | 1459 | 1248 |
| D53-F3: ACTTACTGCATCTGGGTTGATAA D53-R3: ATAGCTTCACACCTTGATTGCAT | 58 | 810 | 1013 | 811 |
| D53-F4: CGGTGTCAACAGTGCAATGAT D53-R4: GCCTCCTGAAGCTGGTGAAT | 60 | 959 | 958 | 959 |
| D53-F5: ATTCACCAGCTTCAGGAGGC D53-R5: ACTACCGTGGACTAGCTACC | 60 | 1403 | 1397 | 1398 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bazhenov, M.; Chernook, A.; Kroupin, P.; Karlov, G.; Divashuk, M. Molecular Characterization of the Dwarf53 Gene Homolog in Dasypyrum Villosum. Plants 2020, 9, 186. https://doi.org/10.3390/plants9020186
Bazhenov M, Chernook A, Kroupin P, Karlov G, Divashuk M. Molecular Characterization of the Dwarf53 Gene Homolog in Dasypyrum Villosum. Plants. 2020; 9(2):186. https://doi.org/10.3390/plants9020186
Chicago/Turabian StyleBazhenov, Mikhail, Anastasiya Chernook, Pavel Kroupin, Gennady Karlov, and Mikhail Divashuk. 2020. "Molecular Characterization of the Dwarf53 Gene Homolog in Dasypyrum Villosum" Plants 9, no. 2: 186. https://doi.org/10.3390/plants9020186
APA StyleBazhenov, M., Chernook, A., Kroupin, P., Karlov, G., & Divashuk, M. (2020). Molecular Characterization of the Dwarf53 Gene Homolog in Dasypyrum Villosum. Plants, 9(2), 186. https://doi.org/10.3390/plants9020186