Comparative Seed Morphology of Tropical and Temperate Orchid Species with Different Growth Habits

 , ,
, , 
Abstract
1. Introduction
2. Results
2.1. Pod and Seed Morphology
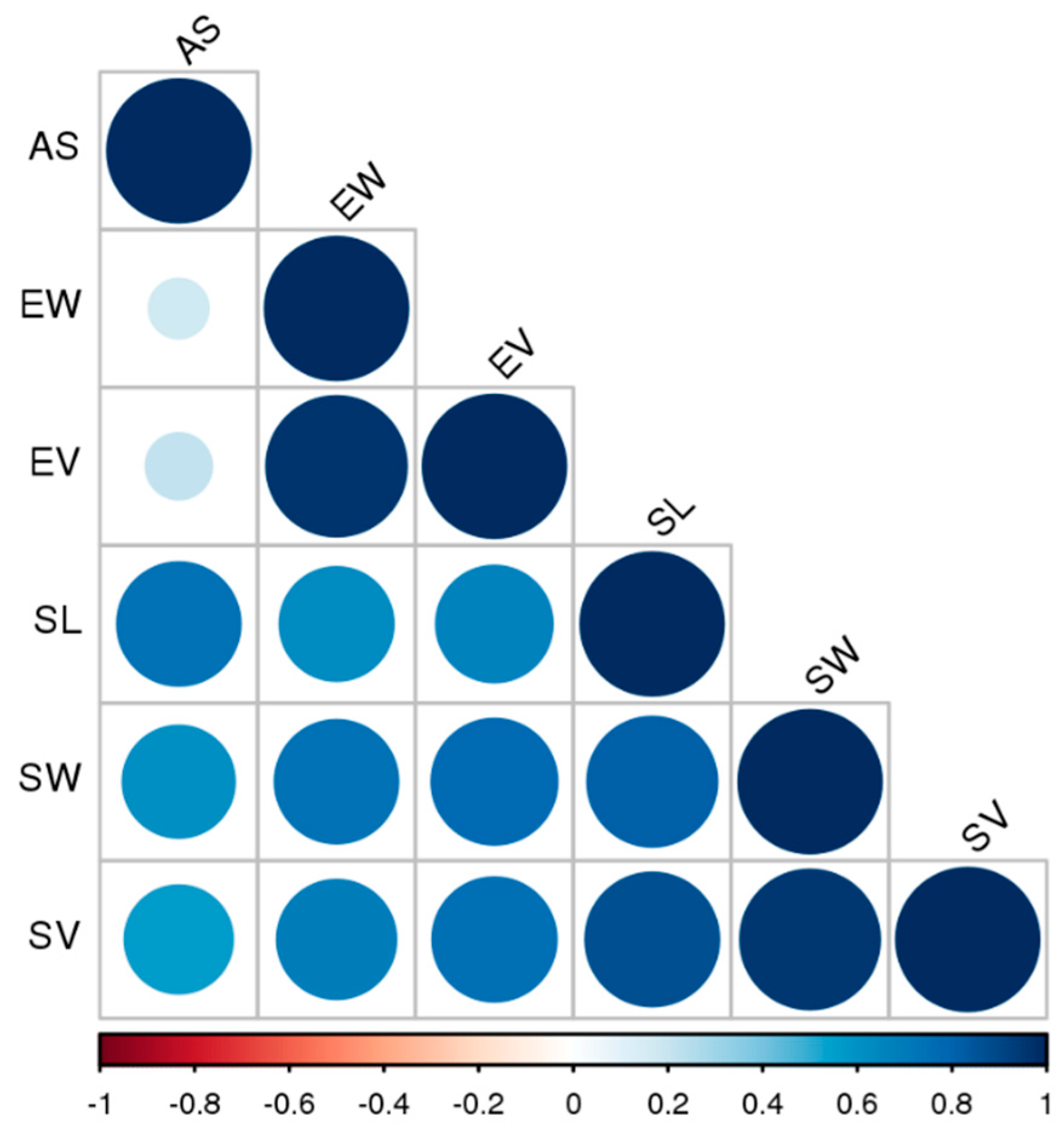
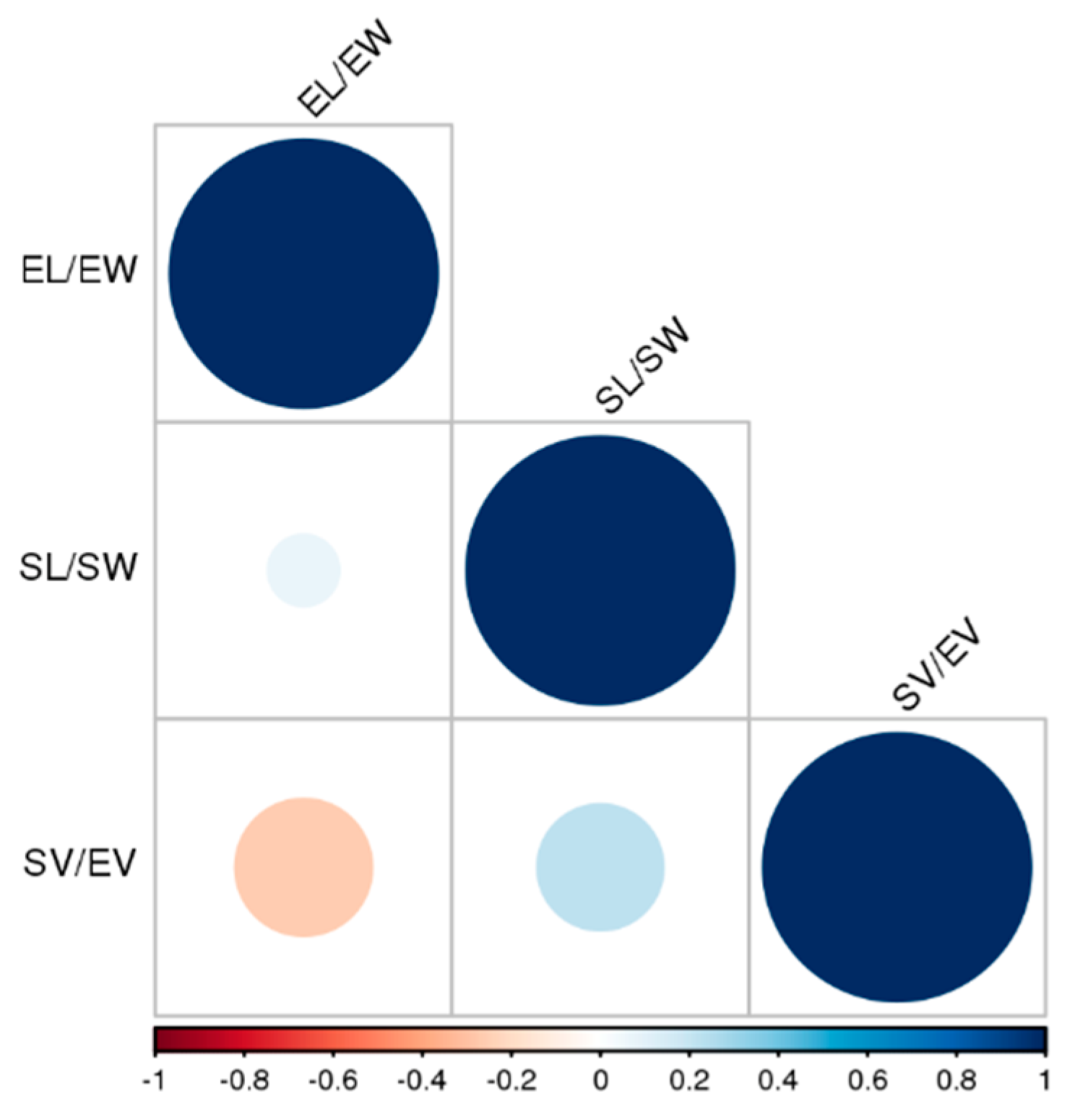
2.2. Seed Micromorphology Assessment
3. Discussion
4. Materials and Methods
4.1. Seed Materials
4.2. Evaluation of Seed Morphological Variability
4.3. Assessment on Seed Micro-Morphometric
4.4. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rubluo, A.; Chavez, V.; Martinez, A.P.; Martinez-Vazques, O. Strategies for the recovery of endangered orchids and cacti through in-vitro culture. Biol. Conserv. 1993, 63, 163–169. [Google Scholar] [CrossRef]
- Prasongsom, S.; Thammasiri, K.; Pritchard, H.W. Seed micromorphology and ex vitro germination of Dendrobium orchids. Acta Hortic. 2017, 1167, 339–344. [Google Scholar] [CrossRef]
- Gallo, F.R.; Souza, L.A.; Milaneze-Gutierre, M.A.; Almeida, O.J.G. Seed structure and in vitro seedling development of certain Laeliinae species (Orchidaceae). Rev. Mex. Biodivers. 2016, 87, 68–73. [Google Scholar] [CrossRef]
- Gamarra, R.; Ortúñez, E.; Cela, P.G.; Merencio, Á. Seed micromorphology of Orchidaceae in the Gulf of Guinea (West Tropical Africa), Plant Syst. Evol. 2018, 304, 665–677. [Google Scholar] [CrossRef]
- Tsutsumi, C.; Yukawa, T.; Lee, N.S.; Lee, C.S.; Kato, M. Phylogeny and comparative seed morphology of epiphytic and terrestrial species of Liparis (Orchidacea) in Japan. J. Plant Res. 2007, 120, 405–412. [Google Scholar] [CrossRef] [PubMed]
- Arditti, J.; Ghani, A.K.A. Tansley Review No. 110. Numerical and physical properties of orchid seeds and their biological implications. New Phytol. 2000, 145, 367–421. [Google Scholar] [CrossRef]
- Healey, P.L.; Michaud, J.D.; Arditti, J. Morphometry of Orchid Seeds. III. Native Claifornia and Related Species of Goodyera, Piperia, Platanthera and Spiranthes. Am. J. Bot. 1980, 67, 508–518. [Google Scholar] [CrossRef]
- Neto, N.B.M.; Custodio, C.C. Orchid conservation through seed banking: Ins and outs. Selbyana 2005, 26, 229–235. [Google Scholar]
- Barthlott, W.; Grosse-Veldmann, B.; Korotkova, N. Orchid seed diversity: A scanning electron microscopy survey. Englera 2014, 32, 3–245. [Google Scholar]
- Calevo, J.; Giovannini, A.; Cornara, L.; Peccenini, S.; Monroy, F. Orchis patens Desf.: Seed morphology of an endangered Mediterranean orchid. Plant Biosys. Int. J. Deal. Asp. Plant Biol. 2017, 151, 770–774. [Google Scholar] [CrossRef]
- Chase, M.W.; Christenhusz, M.J.M.; Mirenda, T. The Book of Orchids: A Life-Size Guide to Six Hundred Species from Around the World; The University of Chicago Press: Chicago, IL, USA, 2017; 656p. [Google Scholar]
- Bianco, M.L.; Grillo, O.; Garcia, P.E.; Mascia, F.; Venora, G.; Bacchetta, G. Morpho-calorimetric characterisation of Malva alliance taxa by seed image analysis. Plant Biol. 2016, 19, 90–98. [Google Scholar] [CrossRef] [PubMed]
- Verma, J.; Sharma, K.; Thakur, K.; Sembi, J.K.; Vij, S.P. Study on seed morphometry of some threatened Western Himalayan orchids. Turk. J. Bot. 2014, 38, 234–251. [Google Scholar] [CrossRef]
- Akcin, T.A.; Ozdener, Y.; Akcin, A. Taxonomic value of seed characters in orchids from Turkey. Belg. J. Bot. 2009, 142, 124–139. [Google Scholar]
- Seedling Ecology and Evolution; Leck, M., Parker, V., Simpson, R., Eds.; Cambridge University Press: Cambridge, UK, 2008. [Google Scholar] [CrossRef]
- Dressler, R.L. The subfamilies of the Orchidaceae. Selbyana 1979, 5, 197–206. [Google Scholar]
- Lavarack, P.S.; Harris, W.F.; Stocker, G. Dendrobium and Its Relatives; Timber Press: Portland, OR, USA, 2000. [Google Scholar]
- Irawati. Chapter 9: Conservation of orchids the germs of the tropics. In Conservation of Tropical Plant Species; Normah, M.N., Chin, H.F., Reed, B.M., Eds.; Springer: New York, NY, USA, 2013; pp. 171–187. [Google Scholar] [CrossRef]
- De Lange, P.J.; Norton, D.A.; Courtney, S.P.; Heenan, P.B.; Barkla, J.W.; Cameron, E.K.; Cameron, R.; Townsend, A.J. Threatened and uncommon plants of New Zealand (2008 revision). N. Z. J. Bot. 2009, 47, 61–96. [Google Scholar] [CrossRef]
- George, S.I. The Natural Guide to New Zealand Orchids; Random House: Auckland, New Zealand, 1999; 176p. [Google Scholar]
- Wang, S.; Xiao, F. Seed morphology of 17 Dendrobium species from Yunnan. China J. Chin. Mater. Med. 2010, 5, 423–426. [Google Scholar] [CrossRef]
- Brzosko, E.; Ostrowiecka, B.; Kotowicz, J.; Bolesta, M.; Gromotowicz, A.; Gromotowicz, M.; Orzechowska, A.; Orzolek, J.; Wojdalska, M. Seed dispersal in six species of terrestrial orchids in Biebrza National Park (NE Poland). Acta Soc. Bot. Pol. 2017, 86, 3557. [Google Scholar] [CrossRef]
- Howe, H.F.; Smallwood, J. Ecology of seed dispersal. Ann. Rev. Ecol. Syst. 1982, 13, 201–228. [Google Scholar] [CrossRef]
- Eriksson, O.; Kainulainen, K. The evolutionary ecology of dust seeds. Perspect. Plant Ecol. Evol. Syst. 2011, 13, 73–87. [Google Scholar] [CrossRef]
- Russel, P.G.; Musil, A.F. Plants must disperse their seeds. In Seeds: The Yearbook of Agriculture; U.S. Gov. Print. Office: Washington, DC, USA, 1961; pp. 80–88. [Google Scholar]
- Chaudhary, B.; Chattopadhyay, P.; Banerjee, N. Modulations in seed micromorphology reveal signature of adaptive species-diversification in Dendrobium (Orchidaceae). Open J. Ecol. 2014, 4, 33–42. [Google Scholar] [CrossRef][Green Version]
- Dangat, B.T.; Gurav, R.V. Studies on seed morphometry of Habenaria species from Western Ghats, India. Richardiana 2016, 16, 174–188. [Google Scholar]
- Emadzade, K.; Lechnebach, C.; Lockhart, P.; Hörandl, E. A molecular phylogeny, morphology and classification of genera of Ranunculeae (Ranunculaceae). Taxon 2010, 59, 809–828. [Google Scholar] [CrossRef]
- Croat, T.B. Flora of Barro Colorado Island; Stanford University Press: Stanford, CA, USA, 1978. [Google Scholar]
- Lehnebach, C.A.; Robertson, A.W. Pollination ecology of four epiphytic orchids of New Zealand. Ann. Bot. 2004, 93, 773–781. [Google Scholar] [CrossRef] [PubMed][Green Version]
- New Zealand Native Orchid Group. A Pocket Guide to the New Zealand Native Orchids; The New Zealand Native Orchid Group Inc.: Wellington, New Zealand, 2015. [Google Scholar]
- Hosomi, S.T.; Santos, R.B.; Custodio, C.C.; Seaton, P.T.; Marks, T.R.; Machado-Neto, N.B. Preconditioning Cattleya seeds to improve the efficacy of the tetrazolium test for viability. Seed Sci. Technol. 2011, 39, 178–189. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Species | Seed Pod Length (cm) | Mature Pod Colour | Seed Colour |
|---|---|---|---|
| Tropical epiphytic | |||
| Dendrobium strebloceras | 6–7 | Yellowish green | Whitish yellow |
| Dendrobium lineale | 3.5–4 | Yellowish green | Yellowish golden |
| Temperate epiphytic | |||
| Dendrobium cunninghamii | 1–1.5 | Yellowish green | Brownish yellow |
| Temperate terrestrial | |||
| Gastrodia cunninghamii | 2–2.5 | (Dark) brown | Brownish (light brown) |
| Pterostylis banksii | 2–2.5 | (Whitish) brown | Ochre (deep orange-brown) |
| Thelymitra nevosa | 1.5–2 | (Purplish) brown | Dark brown |
| Embryo Traits | ||||
|---|---|---|---|---|
| Species | Length (mm) | Width (mm) | Volume *(mm3) | |
| D. strebloceras | 0.25 ± 0.007 a | 0.16 ± 0.005 a | 0.004 ± 0.0003 a | |
| D. lineale | 0.20 ± 0.004 d | 0.097 ± 0.001 d | 0.001 ± 0.00003 d | |
| D. cunninghamii | 0.21 ± 0.004 c | 0.099 ± 0.002 d | 0.001 ± 0.00004 d | |
| G. cunninghamii | 0.23 ± 0.005 b | 0.097 ± 0.001 d | 0.001 ± 0.00004 d | |
| P. banksii | 0.21 ± 0.004 c | 0.13 ± 0.002 c | 0.002 ± 0.0001 c | |
| T. nervosa | 0.24 ± 0.003 a, b | 0.15 ± 0.002 b | 0.003 ± 0.0001 b | |
| Seed Traits | ||||
| Length (mm) | Width (mm) | Volume *(mm3) | Air-space (%) | |
| D. strebloceras | 1.76 ± 0.04 a | 0.32 ± 0.009 a | 0.052 ± 0.003 a | 92.4 ± 0.4 a |
| D. lineale | 0.42 ± 0.006 f | 0.12 ± 0.002 d | 0.001 ± 0.00006 d | 32.2 ± 2 d |
| D. cunninghamii | 0.48 ± 0.008 e | 0.16 ± 0.003 b | 0.003 ± 0.0002 c | 66.2 ± 1.7 c |
| G. cunninghamii | 0.85 ± 0.02 c | 0.13 ± 0.002 c | 0.004 ± 0.002 c | 66.7 ± 1.2 c |
| P. banksii | 1.09 ± 0.03 b | 0.17 ± 0.003 b | 0.009 ± 0.0004 b | 75.3 ± 1.2 b |
| T. nervosa | 0.54 ± 0.006 d | 0.17 ± 0.003 b | 0.004 ± 0.0001 c | 29.3 ± 1.5 d |
| Species | EL/EW | SL/SW | SV/EV |
|---|---|---|---|
| D. strebloceras | 1.55 ± 0.3 d | 5.6 ± 0.1 c | 15.7 ± 1.2 a |
| D. lineale | 2.02 ± 0.4 c | 3.7 ± 0.8 d | 1.58 ± 0.08 c |
| D. cunninghamii | 2.13 ± 0.4 b | 2.9 ± 0.7 f | 3.4 ± 0.2 b |
| G. cunninghamii | 2.36 ± 0.5 a | 6.7 ± 0.2 a | 3.2 ± 0.1 b |
| P. banksii | 1.61 ± 0.3 d | 6.4 ± 0.2 b | 4.9 ± 0.5 b |
| T. nervosa | 1.64 ± 0.3 d | 3.29 ± 0.5 e | 1.4 ±0.03 c |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Diantina, S.; McGill, C.; Millner, J.; Nadarajan, J.; W. Pritchard, H.; Clavijo McCormick, A. Comparative Seed Morphology of Tropical and Temperate Orchid Species with Different Growth Habits. Plants 2020, 9, 161. https://doi.org/10.3390/plants9020161
Diantina S, McGill C, Millner J, Nadarajan J, W. Pritchard H, Clavijo McCormick A. Comparative Seed Morphology of Tropical and Temperate Orchid Species with Different Growth Habits. Plants. 2020; 9(2):161. https://doi.org/10.3390/plants9020161
Chicago/Turabian StyleDiantina, Surya, Craig McGill, James Millner, Jayanthi Nadarajan, Hugh W. Pritchard, and Andrea Clavijo McCormick. 2020. "Comparative Seed Morphology of Tropical and Temperate Orchid Species with Different Growth Habits" Plants 9, no. 2: 161. https://doi.org/10.3390/plants9020161
APA StyleDiantina, S., McGill, C., Millner, J., Nadarajan, J., W. Pritchard, H., & Clavijo McCormick, A. (2020). Comparative Seed Morphology of Tropical and Temperate Orchid Species with Different Growth Habits. Plants, 9(2), 161. https://doi.org/10.3390/plants9020161