Relationship between the Content of β-D-Glucans and Infection with Fusarium Pathogens in Oat (Avena sativa L.) Plants

 , , ,
, , ,
Abstract
1. Introduction
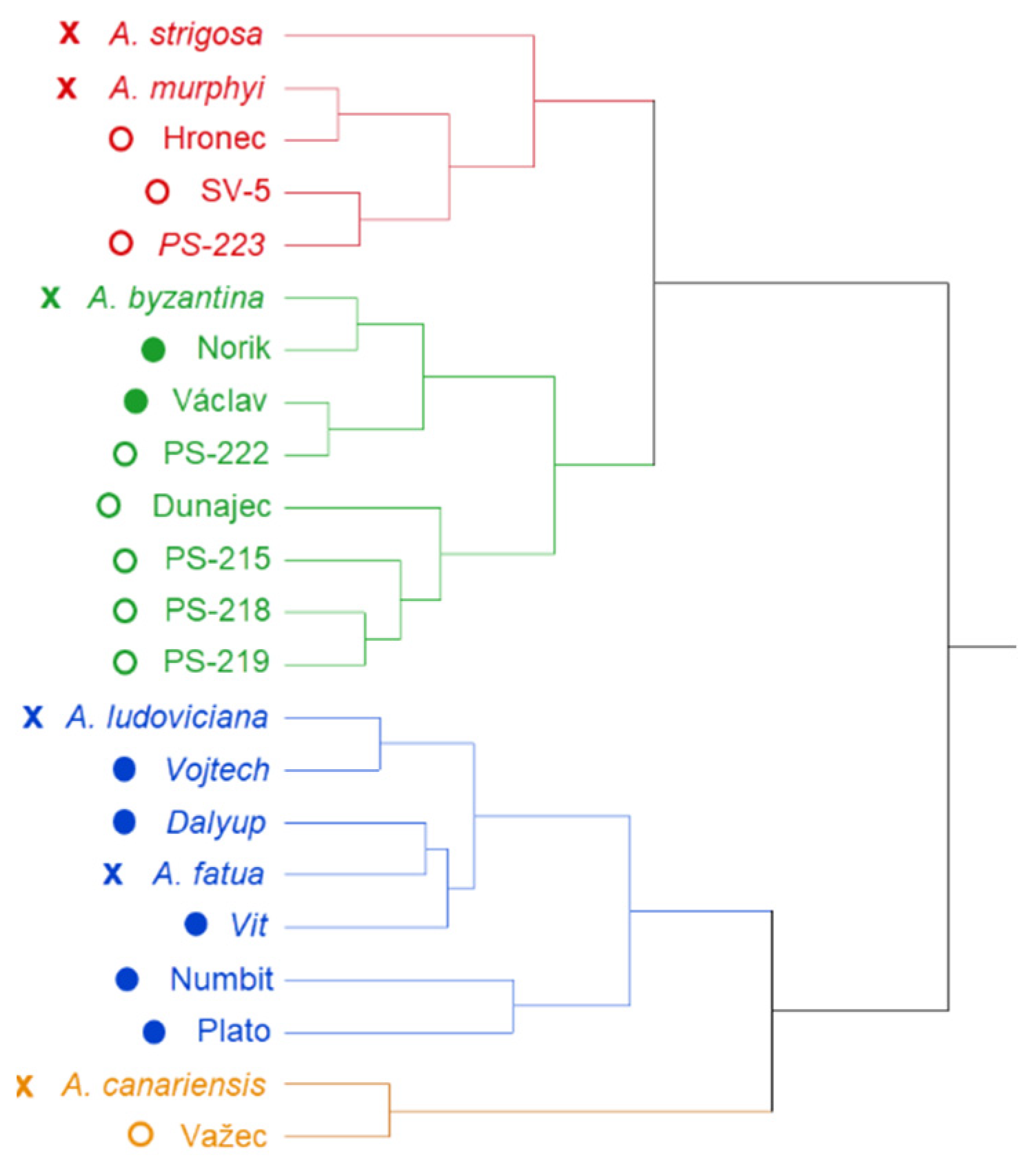
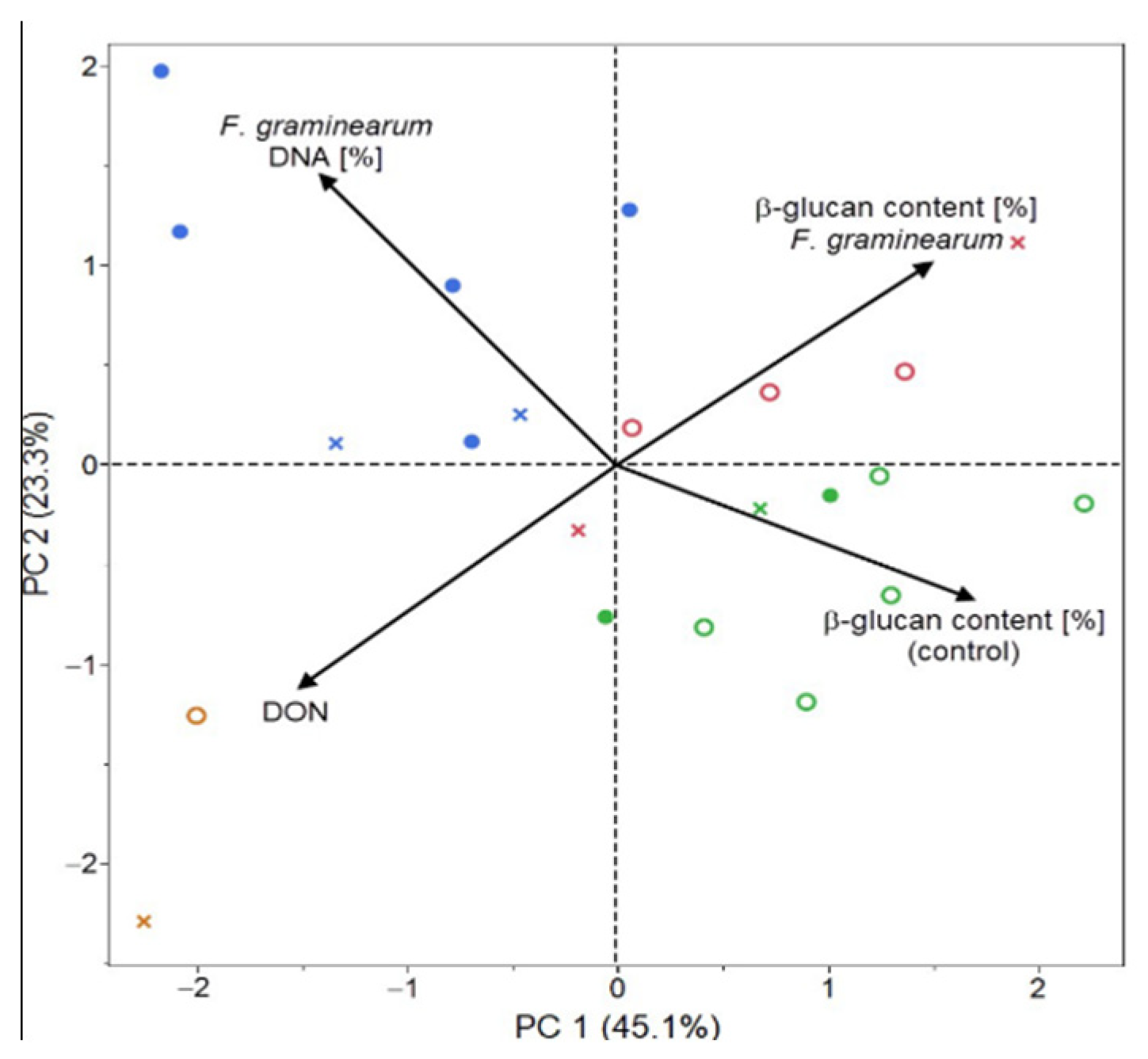
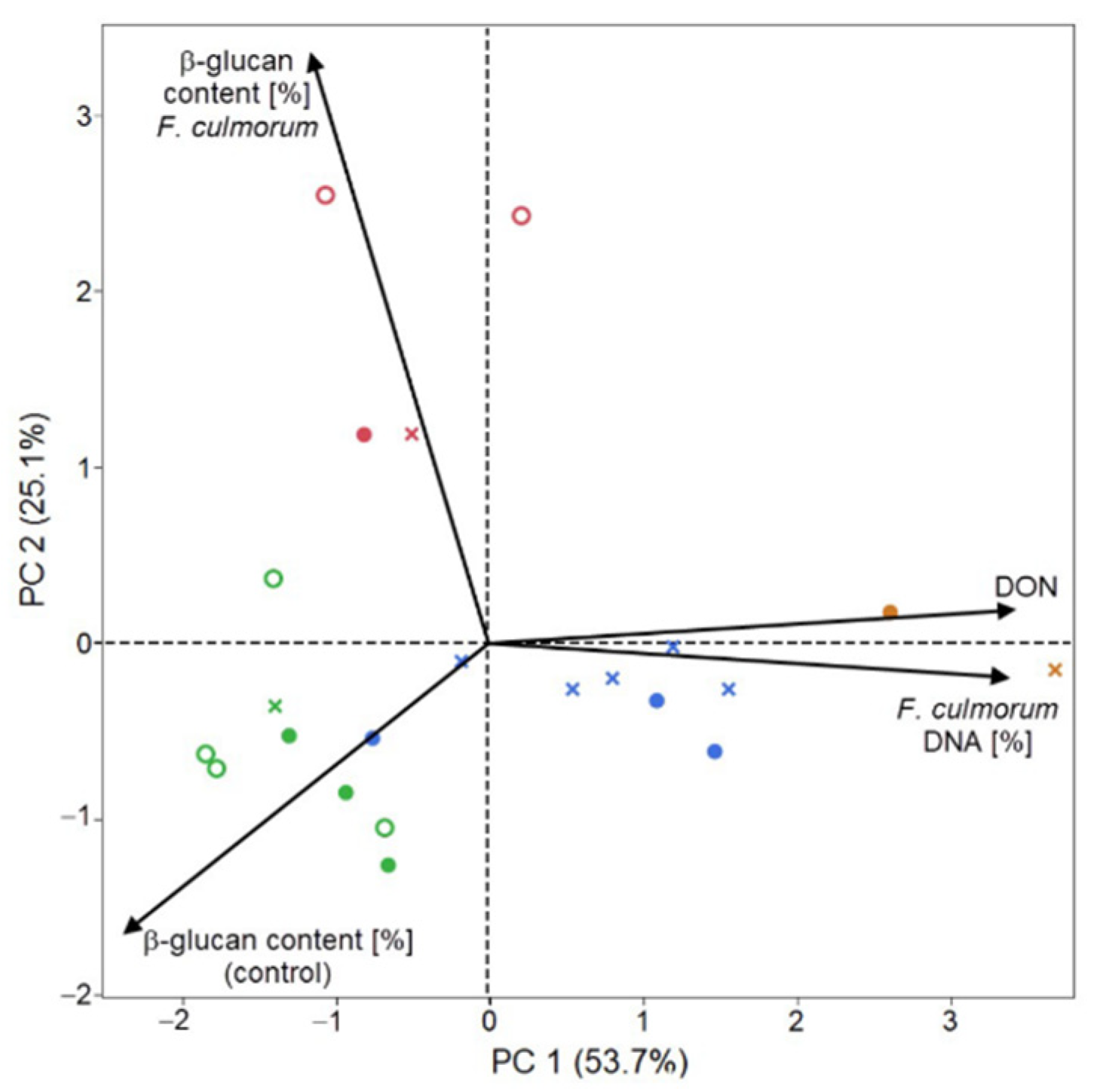
2. Results
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Artificial Inoculation
4.3. Determination of β-D-Glucans
4.4. Quantification of DON
4.5. Extraction and Quantification of DNA
4.6. Statistical Evaluation
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Rasane, P.; Jha, A.; Sabikhi, L.; Kumar, A.; Unnikrishnan, V.S. Nutritional advantages of oats and opportunities for its processing as value added foods—A review. J. Food Sci. Technol. 2015, 52, 662–675. [Google Scholar] [CrossRef] [PubMed]
- Doehlert, D.C.; McMullen, M.S. Genotypic and environmental effects on oat milling characteristics and groat hardness. Cereal Chem. 2000, 77, 148–154. [Google Scholar] [CrossRef]
- FDA. 21 CFR Part 101. Food labeling, health claims: Soluble dietary fiber from certain foods and coronary heart disease. Fed. Regist. 1997, 62, 3584–3601. Available online: https://ecfr.federalregister.gov/current/title-21/chapter-I/subchapter-B/part-101/subpart-E/section-101.81 (accessed on 10 August 2020).
- Parry, D.W.; Jenkinson, P.; McLeod, L. Fusarium Ear blight (scab) in small-grain cereals—A review. Plant Pathol. 1995, 44, 207–238. [Google Scholar] [CrossRef]
- Noworolnik, K. Yielding and grain quality of oat depending on soil moisture and nitrogen rate. Zywnosc Nauka Technol. Jakosc/Food Sci. Technol. Qual. 2010, 70, 190–196. [Google Scholar] [CrossRef]
- Cyrkler-Degulis, M.; Bulinska-Radomska, Z. Value of old and modern oats cultivars for organic farming. J. Res. Appl. Agric. Eng. (Pol.) 2007, 3, 27–31. Available online: https://www.pimr.eu/wp-content/uploads/2019/05/2007_3_MCZB.pdf (accessed on 12 September 2020).
- Nganje, W.E.; Bangsund, D.A.; Leistritz, F.L.; Wilson, W.W.; Tiapo, N.M. Regional economic impacts of Fusarium Head Blight in wheat and barley. Appl. Econ. Perspect. Policy 2004, 26, 332–347. [Google Scholar] [CrossRef]
- Schöneberg, T.; Kiblera, K.; Sulyokc, M.; Musaa, T.; Thomas, D.B.; Maschere, F.; Bertossa, M.; Voegele, R.T.; Vogelgsang, S. Can plant phenolic compounds reduce Fusarium growth and mycotoxin production in cereals? Food Addit. Contam. 2018, 35, 2455–2470. [Google Scholar] [CrossRef]
- Faltusová, Z.; Vaculová, K.; Pavel, J.; Svobodová, I.; Hajšlová, J.; Ovesná, J. Identification of suitable reference gene for quantitative transcription analysis (RT-qPCR) of Fusarium culmorum genes in infected barley plants. J. Cereal Sci. 2018, 79, 418–423. [Google Scholar] [CrossRef]
- Antonissen, G.; Martel, A.; Pasmans, F.; Ducatelle, R.; Verbrugghe, E.; Vanderbroucke, V.; Li, S.; Haesebrouck, F.; Van Immerseel, F.; Croubels, S. The impact of Fusarium mycotoxins on human and animal host susceptibility to infectious diseases. Toxins 2014, 6, 430–452. [Google Scholar] [CrossRef]
- Pleadin, J.; Vahcic, N.; Persi, N.; Sevelj, D.; Markov, K.; Frece, J. Fusarium mycotoxins’ occurrence in cereals harvested from Croatian. Food Control 2013, 32, 49–54. [Google Scholar] [CrossRef]
- WHO. Evaluation of certain mycotoxins in food. Who Tech. Rep. Ser. 2002, 906, 35–42. [Google Scholar]
- Kosová, K.; Chrpová, J.; Šantrůček, J.; Hynek, R.; Štěrbová, L.; Vítámvás, P.; Bradová, J.; Prášil, I.T. The effect of Fusarium culmorum infection and deoxynivalenol (DON) application on proteome response in barley cultivars chevron and pedant. J. Proteom. 2017, 169, 112–124. [Google Scholar] [CrossRef] [PubMed]
- European Commission. Commission recommendation on the presence of deoxynivalenol, zearalenone, ochratoxin A, T-2 and HT-2 and fumonisins in products intended for animal feeding. Off. J. Eur. Union 2006, L229, 7–9. [Google Scholar]
- European Commission. Commission Recommendation on the presence of T-2 and HT-2 toxin in cereals and cereal products. Off. J. Eur. Union 2013, L91/12, 1–4. [Google Scholar]
- Farrokhi, N.; Burton, R.A.; Brownfield, L.; Hrmova, M.; Wilson, S.M.; Bacic, A.; Fincher, G.B. Plant cell wall biosynthesis: Genetic, biochemical and functional genomics approaches to the identification of key genes. Plant. Biotechnol. J. 2006, 4, 145–167. [Google Scholar] [CrossRef] [PubMed]
- Hoson, T. Physiological functions of plant cell coverings. J. Plant Res. 2002, 115, 277–282. [Google Scholar] [CrossRef]
- Hrmova, M.; Fincher, G.B. Structure-function relationship of β-D-glucan endo- and exohydrolases from higher plants. Plant Mol. Biol. 2001, 47, 73–91. [Google Scholar] [CrossRef]
- Luna, E.; Pastor, V.; Robert, J.; Flors, V.; Mauch-Mani, B.; Ton, J. Callose deposition: A multifaceted plant defense response. Mol. Plant. Microbe Interact. J. 2011, 24, 183–193. [Google Scholar] [CrossRef]
- Piršelová, B.; Matušíková, I. Callose: The plant cell wall polysaccharide with multiple biological functions. Acta Physiol. Plant. 2013, 35, 635–644. [Google Scholar] [CrossRef]
- Buckeridge, M.S.; Rayon, C.; Urbanowicz, B.; Tine, M.A.S.; Carpita, N. Mixed linkage (1-3),(1-4)-β-D-glucans of Grasses. Cereal Chem. J. 2004, 81, 115–127. [Google Scholar] [CrossRef]
- Havrlentová, M.; Deáková, L.; Kraic, J.; Žofajová, A. Can β-D-glucan protect oat seeds against a heat stress? Nova Biotechnol. Et Chim. 2016, 15, 107–113. [Google Scholar] [CrossRef]
- Vega- Sanchez, M.E.; Verhertbruggen, Y.; Christensen, U.; Chen, X.; Sharma, V.; Varanasi, P.; Jobling, S.A.; Talbot, M.; White, R.G.; Joo, M.; et al. Loss of Cellulose synthase-like F6 function affects mixed-linkage glucan deposition, cell wall mechanical properties and defense responses in vegetative tissues of rice. Plant Physiol. 2012, 159, 56–69. [Google Scholar] [CrossRef] [PubMed]
- Havrlentova, M.; Kraic, J. Content of β-D-glucan in cereal grains. J. Food Nutr. Res. 2006, 45, 97–103. Available online: https://scholar.google.com/scholar_lookup?title=Content+of+beta-d-glucan+in+cereal+grains&author=Havrlentova%2C+M.&publication_year=2006 (accessed on 12 September 2020).
- Readelli, R.; Sgrulletta, D.; De Stefanis, E. Genetic variability for chemical components in sixty European oat (Avena sativa L.) cultivars. Cereal Res. Commun. 2003, 31, 185–192. [Google Scholar] [CrossRef]
- Zute, S.; Berga, L.; Vicupe, Z. Variability in endosperm β-glucan content of husked and naked oat genotypes. Acta Biol. Univ. Daugavp. 2011, 11, 192–200. Available online: http://sciences.lv/wp-content/uploads/ACTA/2011/11-2/12_Zute.pdf (accessed on 13 September 2020).
- Havrlentová, M.; Bieliková, M.; Mendel, L.; Kraic, J.; Hozlár, P. The correlation of (l-3),(l-4)-beta-D-glucan with some qualitative parameters in the oat grain. Agriculture (Poľnohospodárstvo) 2008, 54, 65–71. Available online: https://www.agriculture.sk/fileadmin/agriculture/files/2006/Issue%204/2_havrl.pdf (accessed on 12 September 2020).
- Tiwari, U.; Cummins, E. Simulation of the factors affecting β-glucan levels during the cultivation of oats. J. Cereal Sci. 2009, 2, 175–183. [Google Scholar] [CrossRef]
- Benlioğlu, B.; Adak, M.S. Importance of crop wild relatives and landraces genetic resources in plant breeding programs. J. Exp. Agric. Int. 2019, 37, 1–8. [Google Scholar] [CrossRef]
- Havrlentová, M.; Petruláková, Z.; Burgárová, A.; Gago, F.; Hlinková, A.; Šturdík, E. Cereal beta-glucans and their significance for the preparation of functional foods—A review. Czech. J. Food Sci. 2011, 29, 11–14. [Google Scholar] [CrossRef]
- Martin, C.; Schöneberg, T.; Vogelgsang, S.; Morisoli, R.; Bertossa, M.; Mauch-Mani, B.; Mascher, F. Resistance against Fusarium graminearum and the relationship to β-glucan in barley grains. Eur. J. Plant Pathol. 2018, 152, 621–634. [Google Scholar] [CrossRef]
- Kofuji, K.; Aoki, A.; Tsubaki, K.; Konishi, M.; Isobe, T.; Murata, Y. Antioxidant Activity of β-Glucan. ISRN Pharm. 2012, 2012, 125864. [Google Scholar] [CrossRef] [PubMed]
- Zhou, K.; Hao, J.; Griffey, C.; Chung, H.; O’Keefe, S.F.; Chen, J.; Hogan, S. Antioxidant properties of Fusarium head blight-resistant and -susceptible soft red winter wheat grains grown in Virginia. J. Agric. Food Chem. 2007, 55, 3729–3736. [Google Scholar] [CrossRef] [PubMed]
- Boutigny, A.-L.; Atanasova-Pénichon, V.; Benet, M.; Barreau, C.; Richard-Forget, F. Natural phenolic acids from wheat bran inhibit Fusarium culmorum trichothecene biosynthesis in vitro by repressing Tri gene expression. Eur. J. Plant. Pathol. 2010, 127, 275–286. [Google Scholar] [CrossRef]
- Pani, G.; Scherm, B.; Azara, E.; Balmas, V.; Jahanshiri, Z.; Carta, P.; Fabbri, D.; Dettori, M.A.; Fadda, A.; Dessì, A.; et al. Natural and natural-like phenolic inhibitors of type B trichothecene in vitro production by the wheat (Triticum spp.) Pathogen Fusarium culmorum. J. Agric. Food Chem. 2014, 62, 4969–4978. [Google Scholar] [CrossRef]
- Yiannikouris, A.; André, G.; Poughon, L.; François, J.; Dussap, C.G.; Jeminet, G.; Bertin, G.; Jouany, J.P. Chemical and conformational study of the interactions involved in mycotoxin complex at ion with β-D-glucans. Biomacromolecules 2006, 7, 1147–1155. Available online: https://pubs.acs.org/doi/10.1021/bm050968t (accessed on 30 September 2020). [CrossRef]
- Yiannikouris, A.; Francois, J.; Poughon, L.; Dussap, C.G.; Bertin, G.; Jeminet, G.; Jouany, J.P. Adsorption of zearalenone by β-D-glucans in the Saccharomyces cerevisiae cell wall. J. Food Prot. 2004, 67, 1195–1200. [Google Scholar] [CrossRef]
- Schwarz, P.B.; Jones, B.L.; Steffensen, B. Enzymes associated with Fusarium infection of barley. J. Am. Soc. Brew. Chem. 2002, 60, 130–134. [Google Scholar] [CrossRef]
- Wang, J.; Wieser, H.; Pawelzik, E.; Weinert, J.; Keutgen, A.J.; Wolf, G.A. Impact of the fungal protease produced by Fusarium culmorum on the protein quality and breadmaking properties of winter wheat. Eur. Food Res. Technol. 2005, 220, 552–559. [Google Scholar] [CrossRef]
- Oliveira, P.M.; Mauch, A.; Jacob, F.; Arendt, E.K. Impact of Fusarium culmorum-infected barley malt grains on brewing and beer quality. J. Am. Soc. Brew. Chem. 2012, 70, 186–194. [Google Scholar] [CrossRef]
- Oliveira, P.M.; Mauch, A.; Jacob, F.; Waters, D.M.; Arendt, E.K. Fundamental study on the influence of Fusarium infection on quality and ultrastructure of barley malt. Int. J. Food Microbiol. 2012, 156, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Fincher, G.B. Morphology and chemical composition of barley endosperm cell walls. J. Inst. Brew. 1975, 81, 116–122. [Google Scholar] [CrossRef]
- Becker, M.; Vincent, C.; Reid, J.S.G. Biosynthesis of (1,3)(1,4)-beta-glucan and (1,3)-beta-glucan in barley (Hordeum vulgare L.). Planta 1995, 10, 331–338. [Google Scholar] [CrossRef]
- Wilson, S.M.; Burton, R.A.; Collins, H.M.; Doblin, M.S.; Pettolino, F.A.; Shirley, N.; Fincher, G.B.; Bacic, A. Pattern of deposition of cell wall polysaccharides and transcript abundance of related cell wall synthesis genes during differentiation in barley endosperm. Plant. Physiol. 2012, 159, 655–670. [Google Scholar] [CrossRef] [PubMed]
- Linsel, K.J.; Keiper, F.J.; Forgan, A.; Oldach, K.H. New insights into the infection process of Rhynchosporium secalis in barley using GFP. Fungal Genet. Biol. 2011, 48, 124–131. [Google Scholar] [CrossRef] [PubMed]
- Mesterházy, Á.; Bartók, T.; Kászonyi, G.; Varga, M.; Tóth, B.; Varga, J. Common resistance to different Fusarium spp. causing Fusarium Head Blight in wheat. Eur. J. Plant Pathol. 2005, 112, 267–281. [Google Scholar] [CrossRef]
- Tóth, B.; Kászonyi, G.; Bartók, T.; Varga, M.; Mesterházy, Á. Common resistance of wheat to members of the Fusarium graminearum species complex and F. culmorum. Plant. Breed. 2008, 127, 1–8. [Google Scholar] [CrossRef]
- Tekle, S.; Dill-Macky, R.; Skinnes, H.; Tronsmo, A.M.; Bjørnstad, Å. Infection process of Fusarium graminearum in oats (Avena sativa L.). Eur. J. Plant Pathol. 2012, 132, 431–442. [Google Scholar] [CrossRef]
- Martin, C.; Schöneberg, M.C.; Vogelgsang, S.; Mendes Ferreira, C.S.; Morisoli, R.; Bertossa, M.; Bucheli, T.D.; Mauch-Mani, B.; Mascher, F. Responses of Oat Grains to Fusarium poae and F. langsethiae infections and mycotoxin contaminations. Toxins 2018, 10, 47. [Google Scholar] [CrossRef]
- Miller, S.S.; Fulcher, R.G.; Sen, A.; Arnason, J.T. Oat endosperm cell walls. Isolation, composition and comparison with other tissues. Cereal Chem. 1995, 72, 421–427. Available online: https://scholar.google.com/scholar_lookup?title=Oat+endosperm+cell+walls.+Isolation,+composition,+and+comparison+with+other+tissues&author=Miller,+S.S.&author=Fulcher,+R.G.&author=Sen,+A.&author=Arnason,+J.T.&publication_year=1995&journal=Cereal+Chem.&volume=72&pages=421%E2%80%93427 (accessed on 12 September 2020).
- Tvarůžek, L.; Matušinský, P.; Vyšohlídová, M. Metodika pro zakládání a hodnocení pokusů s umělou inokulací obilnin fuzáriózami klasů. 2012. Available online: https://www.vukrom.cz/userfiles/files/vysledky_vyzkumu/Metodiky/2012_Metodika_pro_zakladani_a_hodnoceni_pokusu_s_umelou_inokulaci_obilnin_fuzariozami_klasu.pdf (accessed on 10 May 2019).
- Šliková, S.; Hozlár, P.; Gregová, E. Ovos: Infekcia Hubami Fusarium spp. v Poľných Podmienkach; Edičné Stredisko NPPC, Výskumný Ústav Pôdoznalectva a Ochrany Pôdy: Bratislava, Slovakia, 2017; p. 91. [Google Scholar]
- Nirenberg, H. Untersuchungen über die morphologische und biologische Differenzierung in der Fusarium Sektion Liseola. Mitt. Aus Der Biol. Bundesanst. Für Land Und Forstwirtsch. 1976, 169, 1–117. [Google Scholar] [CrossRef]
- McCleary, B.V.; Codd, R. Measurement of (1 → 3),(1 → 4)-β-D-glucan in barley and oats: A streamlined enzymic procedure. J. Sci. Food Agric. 1991, 55, 303–312. [Google Scholar] [CrossRef]
- Leišová, L.; Kučera, L.; Chrpová, J.; Sýkorová, S.; Šíp, V.; Ovesná, J. Quantification of Fusarium culmorum in wheat and barley tissues using real_time PCR in comparison with DON content. J. Phytopathol. 2006, 154, 603–611. [Google Scholar] [CrossRef]
- Burlakoti, R.R.; Estrada, R.; Rivera, V.V.; Boddeda, A.; Secor, G.A.; Adhikari, T.B. Real-time PCR quantification and mycotoxin production of Fusarium graminearum in wheat inoculated with isolates collected from potato, sugar beet and wheat. Phytopathology 2007, 97, 835–841. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| No. | Sample | β-D-Glucans in Control (%) | FC Inoculation | FG Inoculation | |||||
|---|---|---|---|---|---|---|---|---|---|
| β-D-Glucans after Inoculation (%) | DON (mg/kg) | Pathogen DNA (%) | β-D-Glucans after Inoculation (%) | DON (mg/kg) | Pathogen DNA (%) | ||||
| 1. | A. strigosa | 2.55 | 3.30 | 19.39 | 0.28 | 2.90 | 3.17 | 0.28 | |
| 2. | A. ludoviciana | 1.67 | 1.40 | 28.19 | 1.23 | 1.64 | 6.52 | 1.20 | |
| 3. | A. byzantina | 2.52 | 1.41 | 2.32 | 0.04 | 1.23 | 1.16 | 0.04 | |
| 4. | A. fatua | 0.71 | 0.89 | 15.29 | 1.23 | 0.93 | 4.78 | 1.23 | |
| 5. | A. canariensis | 1.30 | 0.89 | 23.33 | 0.03 | 0.72 | 14.93 | 0.03 | |
| 6. | A. murphyi | 1.29 | 1.21 | 14.14 | 0.08 | 1.33 | 4.63 | 0.08 | |
| 7. | A. sativa/hulled | Vaclav | 3.90 | 3.25 | 15.43 | 0.59 | 0.94 | 4.59 | 0.59 |
| 8. | Vojtech | 1.54 | 0.78 | 22.33 | 2.16 | 1.89 | 7.51 | 2.16 | |
| 9. | Dalyup | 2.77 | 1.48 | 25.96 | 1.98 | 1.9 | 5.32 | 1.98 | |
| 10. | Numbit | 1.16 | 1.55 | 29.20 | 3.55 | 1.68 | 10.47 | 3.55 | |
| 11. | Vit | 1.62 | 1.14 | 28.84 | 1.71 | 1.63 | 1.22 | 1.73 | |
| 12. | Plato | 0.96 | 1.11 | 37.20 | 4.29 | 0.94 | 3.22 | 4.29 | |
| 13. | Norik | 3.39 | 1.13 | 10.27 | 0.32 | 1.48 | 2.12 | 0.33 | |
| 14. | A. sativa/naked | Hronec | 1.90 | 4.60 | 7.47 | 0.23 | 1.55 | 3.24 | 0.23 |
| 15. | Važec | 3.15 | 3.94 | 21.68 | 0.92 | 1.00 | 12.77 | 0.92 | |
| 16. | SV-5 | 2.54 | 2.20 | 4.33 | 0.11 | 2.22 | 2.84 | 0.12 | |
| 17. | PS-223 | 1.61 | 0.88 | 5.93 | 0.07 | 1.89 | 2.76 | 0.08 | |
| 18. | PS-222 | 3.73 | 0.77 | 13.77 | 0.46 | 0.99 | 3.72 | 0.46 | |
| 19. | Dunajec | 4.98 | 2.00 | 14.20 | 0.44 | 1.63 | 8.34 | 0.41 | |
| 20. | PS-215 | 5.06 | 1.84 | 9.70 | 0.11 | 2.10 | 2.94 | 0.10 | |
| 21. | PS-218 | 4.43 | 1.75 | 4.39 | 0.18 | 1.54 | 3.85 | 0.17 | |
| 22. | PS-219 | 4.72 | 1.36 | 15.25 | 1.30 | 1.64 | 3.24 | 1.30 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Havrlentová, M.; Gregusová, V.; Šliková, S.; Nemeček, P.; Hudcovicová, M.; Kuzmová, D. Relationship between the Content of β-D-Glucans and Infection with Fusarium Pathogens in Oat (Avena sativa L.) Plants. Plants 2020, 9, 1776. https://doi.org/10.3390/plants9121776
Havrlentová M, Gregusová V, Šliková S, Nemeček P, Hudcovicová M, Kuzmová D. Relationship between the Content of β-D-Glucans and Infection with Fusarium Pathogens in Oat (Avena sativa L.) Plants. Plants. 2020; 9(12):1776. https://doi.org/10.3390/plants9121776
Chicago/Turabian StyleHavrlentová, Michaela, Veronika Gregusová, Svetlana Šliková, Peter Nemeček, Martina Hudcovicová, and Dominika Kuzmová. 2020. "Relationship between the Content of β-D-Glucans and Infection with Fusarium Pathogens in Oat (Avena sativa L.) Plants" Plants 9, no. 12: 1776. https://doi.org/10.3390/plants9121776
APA StyleHavrlentová, M., Gregusová, V., Šliková, S., Nemeček, P., Hudcovicová, M., & Kuzmová, D. (2020). Relationship between the Content of β-D-Glucans and Infection with Fusarium Pathogens in Oat (Avena sativa L.) Plants. Plants, 9(12), 1776. https://doi.org/10.3390/plants9121776