Generation Times of E. coli Prolong with Increasing Tannin Concentration while the Lag Phase Extends Exponentially



Abstract
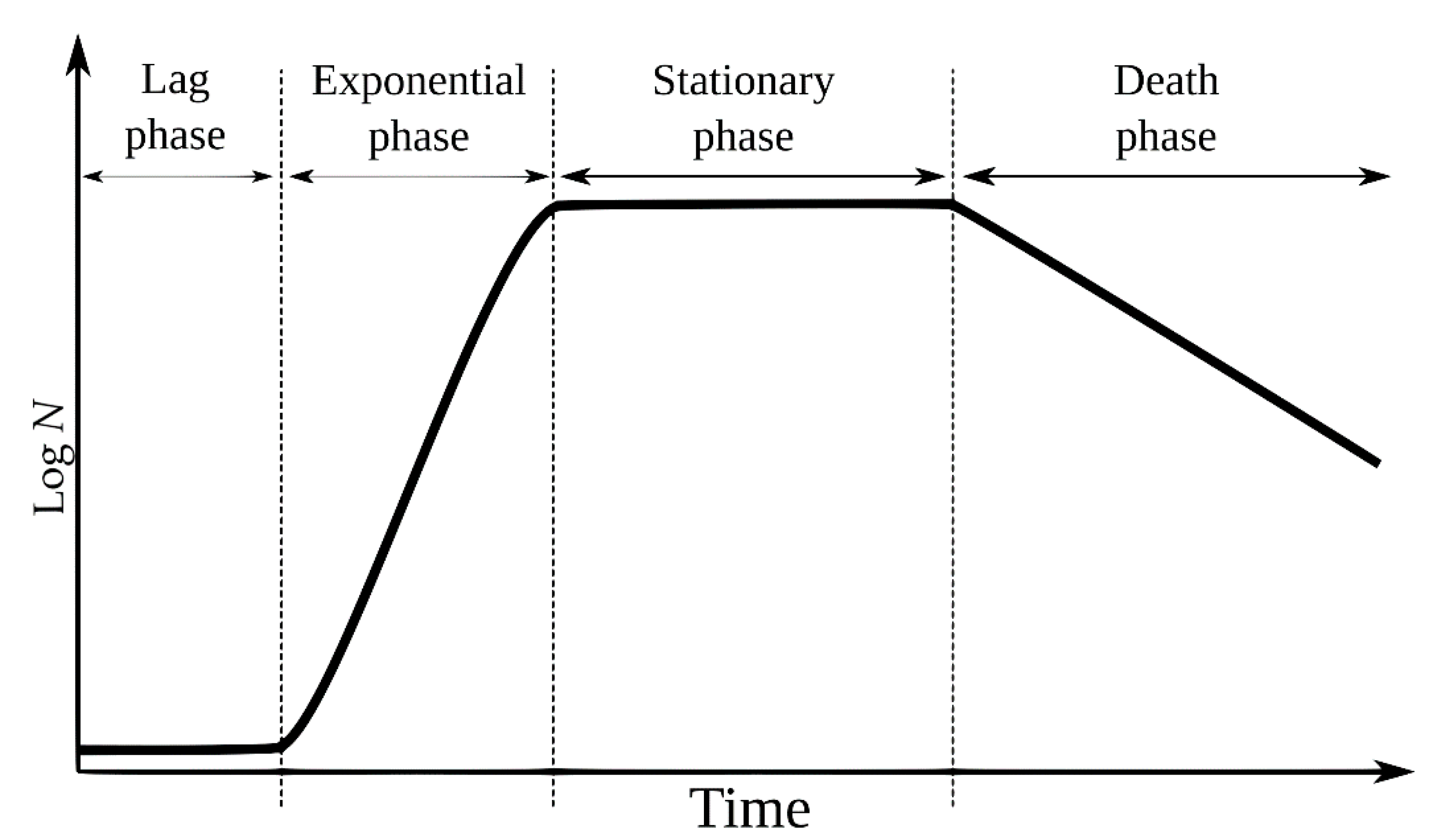
1. Introduction
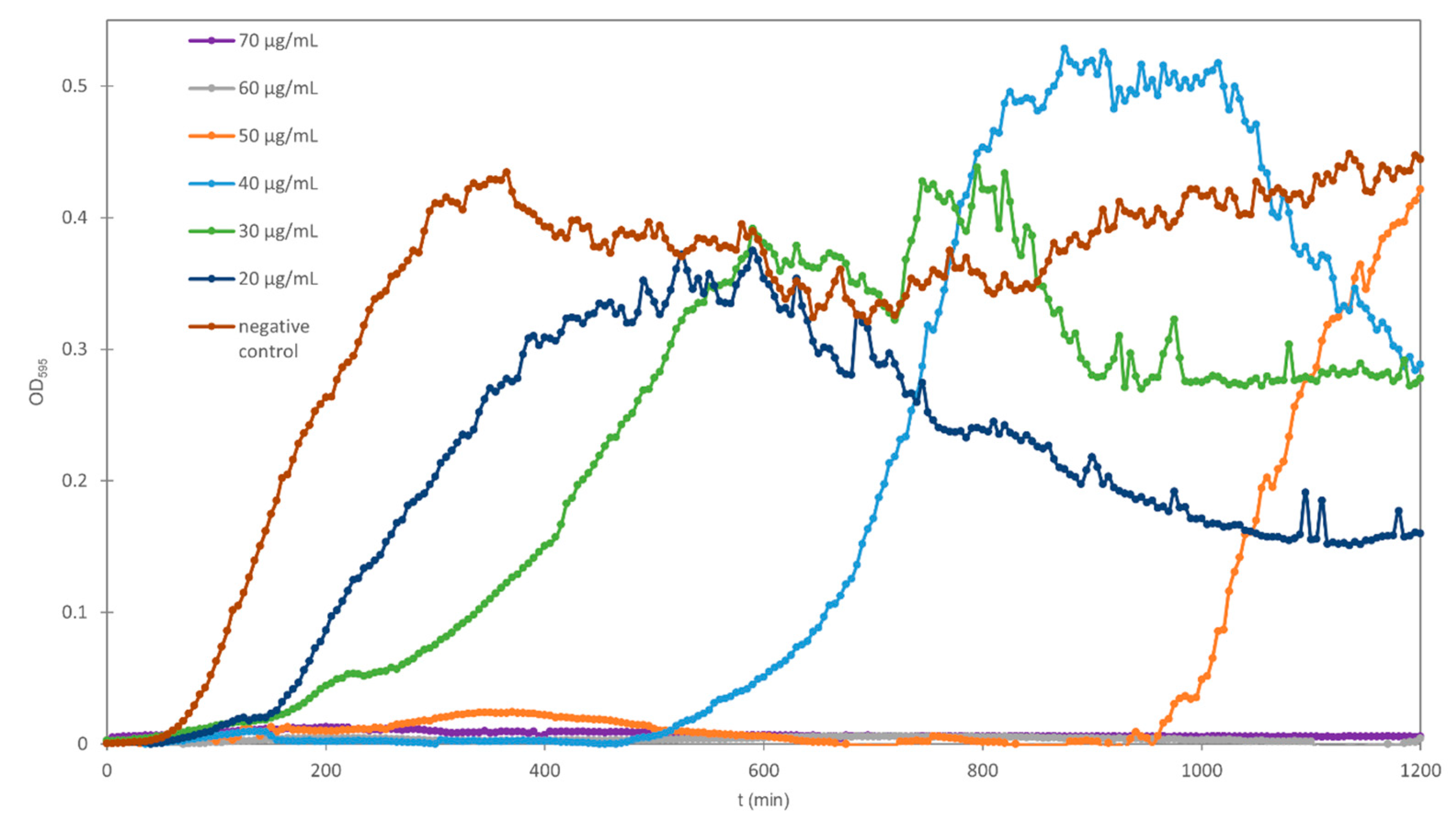
2. Results and Discussion
2.1. Growth Rates and Generation Times
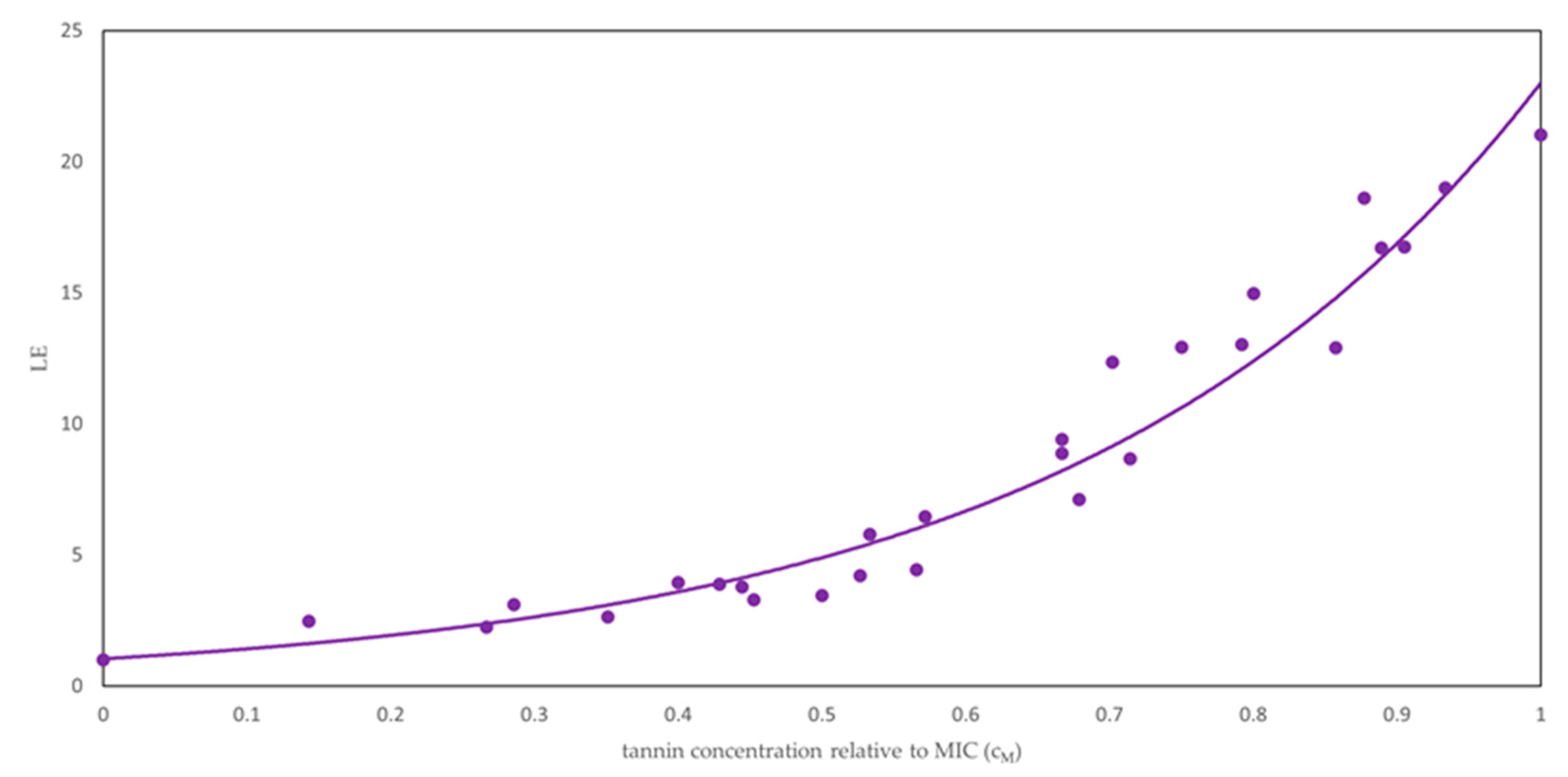
2.2. Lag Times
3. Materials and Methods
3.1. Studied Agents
3.2. Bacterial Strain
3.3. Kinetic Growth Assay
3.4. Determination of Lag Times, Bacterial Growth Rates and Generation Times
3.5. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Cowan, M.M. Plant Products as Antimicrobial Agents. Clin. Microbiol. Rev. 1999, 12, 564–582. [Google Scholar] [CrossRef] [PubMed]
- Mai, P.Y.; Levasseur, M.; Buisson, D.; Touboul, D.; Eparvier, V. Identification of Antimicrobial Compounds from Sandwithia guyanensis-Associated Endophyte Using Molecular Network Approach. Plants 2019, 9, 47. [Google Scholar] [CrossRef] [PubMed]
- Huang, Q.; Liu, X.; Zhao, G.; Hu, T.; Wang, Y. Potential and challenges of tannins as an alternative to in-feed antibiotics for farm animal production. Anim. Nutr. 2018, 4, 137–150. [Google Scholar] [CrossRef]
- Chandra, H.; Bishnoi, P.; Yadav, A.; Patni, B.; Mishra, A.P.; Nautiyal, A.R. Antimicrobial resistance and the alternative resources with special emphasis on plant-based antimicrobials-A review. Plants 2017, 6, 16. [Google Scholar] [CrossRef] [PubMed]
- Serrano, J.; Puupponen-Pimiä, R.; Dauer, A.; Aura, A.-M.; Saura-Calixto, F. Tannins: Current knowledge of food sources, intake, bioavailability and biological effects. Mol. Nutr. Food Resour. 2009, 53, 310–329. [Google Scholar] [CrossRef] [PubMed]
- Khanbabaee, K.; Ree, T.V. Tannins: Classification and definition. R. Soc. Chem. 2001, 18, 641–649. [Google Scholar] [CrossRef]
- Santos-Buelga, C.; Scalbert, A. Proanthocyanidins and tannin-like compounds—Nature, occurrence, dietary intake and effects on nutrition and health. J. Sci. Food Agric. 2000, 80, 1094–1117. [Google Scholar] [CrossRef]
- Frutos, P.; Hervás, G.; Giráldez, F.J.; Mantecón, A.R. Review. Tannins and ruminant nutrition. Span. J. Agric. Res. 2004, 2, 191–202. [Google Scholar] [CrossRef]
- Scalbert, A. Antimicrobial properties of tannins. Phytochemistry 1991, 30, 3875–3883. [Google Scholar] [CrossRef]
- Frankel, E.N.; Kanner, J.; German, J.B.; Parks, E.; Kinsella, J.E. Inhibition of oxidation of human low-density lipoprotein by phenolic substances in red wine. Lancet 1993, 341, 454–457. [Google Scholar] [CrossRef]
- Smith, A.H.; Mackie, R.I. Effect of Condensed Tannins on Bacterial Diversity and Metabolic Activity in the Rat Gastrointestinal Tract. Appl. Environ. Microbiol. 2004, 70, 1104–1115. [Google Scholar] [CrossRef] [PubMed]
- Ikigai, H.; Nakae, T.; Hara, Y.; Shimamura, T. Bactericidal catechins damage the lipid bilayer. Biochim. Biophys. Acta 1993, 1147, 132–136. [Google Scholar] [CrossRef]
- Banso, A.; Adeyemo, S.O. Evaluation of antibacterial properties of tannins isolated from Dichrostachys cinerea. Afr. J. Biotechnol. 2007, 6, 1785–1787. [Google Scholar] [CrossRef]
- Fraga-Corral, M.; García-Oliveira, P.; Pereira, A.G.; Lourenço-Lopes, C.; Jimenez-Lopez, C.; Prieto, M.A.; Simal-Gandara, J. Technological application of tannin-based extracts. Molecules 2020, 25. 614. [Google Scholar] [CrossRef] [PubMed]
- Madigan, M.T.; Martinko, J.M.; Bender, K.S.; Buckley, D.H.; Stahl, D.A. Brock Biology of Microorganisms, 4th ed.; Macmillan Magazines Ltd: Stuttgart, Germany, 2015. [Google Scholar]
- Rolfe, M.D.; Rice, C.J.; Lucchini, S.; Pin, C.; Thompson, A.; Cameron, A.D.S.; Alston, M.; Stringer, M.F.; Betts, R.P.; Baranyi, J.; et al. Lag phase is a distinct growth phase that prepares bacteria for exponential growth and involves transient metal accumulation. J. Bacteriol. 2012, 194, 686–701. [Google Scholar] [CrossRef] [PubMed]
- Besser, M.D.R.E.; Griffin, M.D.P.M.; Slutsker, M.D.M.P.H.L. Escherichia coli O157:H7 gastroenteritis and the hemolytic uremic syndrome: An emerging infectious disease. Annu. Rev. Med. 1999, 50, 355–367. [Google Scholar] [CrossRef] [PubMed]
- Andrews, J.M. Determination of minimum inhibitory concentrations. J. Antimicrob. Chemother. 2001, 48, 5–16. [Google Scholar] [CrossRef]
- Balouiri, M.; Sadiki, M.; Ibnsouda, S.K. Methods for in vitro evaluating antimicrobial activity: A review. J. Pharm. Anal. 2016, 6, 71–79. [Google Scholar] [CrossRef]
- Henis, Y.; Tagari, H.; Volcani, R. Effect of water extracts of carob pods, tannic acid, and their derivatives on the morphology and growth of microorganisms. J. Appl. Microbiol. 1964, 12, 204–209. [Google Scholar] [CrossRef]
- Chung, K.T.; Stevens, S.E., Jr.; Lin, W.F.; Wei, C.I. Growth inhibition of selected food-borne bacteria by tannic acid, propyl gallate and related compounds. Lett. Appl. Microbiol. 1993, 17, 29–32. [Google Scholar] [CrossRef]
- Saxena, G.; McCutcheon, A.R.; Farmer, S.; Towers, G.H.N.; Hancock, R.E.W. Antimicrobial constituents of Rhus glabra. J. Ethnopharmacol. 1994, 42, 95–99. [Google Scholar] [CrossRef]
- Chung, K.T.; Lu, Z.; Chou, M.W. Mechanism of inhibition of tannic acid and related compounds on the growth of intestinal bacteria. Food Chem. Toxicol. 1998, 36, 1053–1060. [Google Scholar] [CrossRef]
- Taguri, T.; Tanaka, T.; Kouno, I. Antimicrobial Activity of 10 Different Plant Polyphenols against Bacteria Causing Food-Borne Disease. Biol. Pharm. Bull. 2004, 27, 1965–1969. [Google Scholar] [CrossRef] [PubMed]
- Becker, H.; Scher, J.M.; Speakman, J.-B.; Zapp, J. Bioactivity guided isolation of antimicrobial compounds from Lythrum salicaria. Fitoterapia 2005, 76, 580–584. [Google Scholar] [CrossRef] [PubMed]
- Min, B.R.; Pinchak, W.E.; Anderson, R.C.; Callaway, T.R. Effect of tannins on the in vitro growth of Escherichia coli O157:H7 and in vivo growth of generic Escherichia coli excreted from steers. J. Food Prot. 2007, 70, 543–550. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, F.; Hatano, T.; Ito, H.; Taniguchi, S.; Takanashi, E.; Okamoto, K. Antibacterial effects of guava tannins and related polyphenols on Vibrio and Aeromonas species. Nat. Prod. Commun. 2008, 3, 711–720. [Google Scholar] [CrossRef]
- Shuaibu, M.N.; Wuyep, P.A.; Yanagi, T.; Hirayama, K.; Tanaka, T.; Kouno, I. The use of microfluorometric method for activity-guided isolation of antiplasmodial compound from plant extracts. Parasitol. Res. 2008, 102, 1119–1127. [Google Scholar] [CrossRef]
- Borges, A.; Ferreira, C.; Saavedra, M.J.; Simões, M. Antibacterial activity and mode of action of ferulic and gallic acids against pathogenic bacteria. Microb. Drug Resist. 2013, 19, 256–265. [Google Scholar] [CrossRef]
- Rios, J.L.; Recio, M.C.; Villar, A. Screening methods for natural products with antimicrobial activity: A review of the literature. J. Ethnopharmacol. 1988, 23, 127–149. [Google Scholar] [CrossRef]
- Gehrt, A.; Peter, J.; Pizzo, P.A.; Walsh, T.J. Effect of increasing inoculum sizes of pathogenic filamentous fungi on MICs of antifungal agents by broth microdilution method. J. Clin. Microbiol. 1995, 33, 1302–1307. [Google Scholar] [CrossRef]
- Jiang, L. Comparison of disk diffusion, agar dilution, and broth microdiultion for antimicrobial susceptibility testing of five chitosans. Master’s Thesis, Louisiana State University and Agricultural and Mechanical College, Baton Rouge, LA, USA, 2011; pp. 1–58. [Google Scholar]
- Stumpf, S.; Hostnik, G.; Primožič, M.; Leitgeb, M.; Salminen, J.-P.; Bren, U. The effect of growth medium strength on minimum inhibitory concentrations of tannins and tannin extracts against E. coli. Molecules 2020, 25. 2947. [Google Scholar] [CrossRef] [PubMed]
- Fridman, O.; Goldberg, A.; Ronin, I.; Shoresh, N.; Balaban, N.Q. Optimization of lag time underlies antibiotic tolerance in evolved bacterial populations. Nature 2014, 513, 418–421. [Google Scholar] [CrossRef] [PubMed]
- Theophel, K.; Schacht, V.J.; Schlüter, M.; Schnell, S.; Stingu, C.S.; Schaumann, R.; Bunge, M. The importance of growth kinetic analysis in determining bacterial susceptibility against antibiotics and silver nanoparticles. Front. Microbiol. 2014, 5, 1–10. [Google Scholar] [CrossRef]
- Li, B.; Qiu, Y.; Shi, H.; Yin, H. The importance of lag time extension in determining bacterial resistance to antibiotics. Analyst 2016, 141, 3059–3067. [Google Scholar] [CrossRef] [PubMed]
- O’Donovan, L.; Brooker, J.D. Effect of hydrolysable and condensed tannins on growth, morphology and metabolism of Streptococcus gallolyticus (S. caprinus) and Streptococcus bovis. Microbiology 2001, 147, 1025–1033. [Google Scholar] [CrossRef]
- Widdel, F. Theory and measurement of bacterial growth. Di dalam Grund. Mikrobiol. 2007, 4, 1–11. [Google Scholar]
- Hider, R.C.; Kong, X. Chemistry and biology of siderophores. Nat. Prod. Rep. 2010, 27, 637–657. [Google Scholar] [CrossRef]
- Searle, L.J.; Méric, G.; Porcelli, I.; Sheppard, S.K.; Lucchini, S. Variation in siderophore biosynthetic gene distribution and production across environmental and faecal populations of Escherichia coli. PLoS ONE 2015, 10, 1–14. [Google Scholar] [CrossRef]
- Smith, A.H.; Zoetendal, E.; Mackie, R.I. Bacterial mechanisms to overcome inhibitory effects of dietary tannins. Microb. Ecol. 2005, 50, 197–205. [Google Scholar] [CrossRef]
- Wiegand, I.; Hilpert, K.; Hancock, R.E.W. Agar and broth dilution methods to determine the minimal inhibitory concentration (MIC) of antimicrobial substances. Nat. Protoc. 2008, 3, 163–175. [Google Scholar] [CrossRef]
- Robinson, T.P.; Ocio, M.J.; Kaloti, A.; MacKey, B.M. The effect of the growth environment on the lag phase of Listeria monocytogenes. Int. J. Food Microbiol. 1998, 44, 83–92. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Sample | csample (µg/mL) | µ (min−1) | td (min) | Lag (min) |
|---|---|---|---|---|
| negative control | 0 | 0.043 ± 0.019 | 20 ± 11 | 53 ± 9 |
| Chestnut | 200 | 0.030 ± 0.008 | 25 ± 6 | 129 ± 19 |
| 300 | 0.024 ± 0.004 | 29 ± 5 | 159 ± 28 | |
| 400 | 0.021 ± 0.004 | 34 ± 5 | 205 ± 35 | |
| 500 | 0.025 ± 0.009 | 30 ± 10 | 419 ± 126 | |
| Quebracho | 200 | 0.024 ± 0.007 | 30 ± 7 | 183 ± 39 |
| 300 | 0.025 ± 0.011 | 34 ± 18 | 685 ± 101 | |
| 400 | 0.010 ± 0.001 | 67 ± 1 | 1115 ± 10 | |
| Colistizer | 200 | 0.034 ± 0.012 | 22 ± 7 | 119 ± 14 |
| 300 | 0.028 ± 0.009 | 27 ± 8 | 209 ± 12 | |
| 400 | 0.031 ± 0.013 | 25 ± 8 | 307 ± 36 | |
| 500 | 0.032 ± 0.015 | 27 ± 12 | 499 ± 86 | |
| 600 | 0.028 ± 0.012 | 29 ± 12 | 794 ± 85 | |
| 700 | 0.034 ± 0.025 | 34 ± 25 | 1007 ± 128 | |
| Mimosa | 200 | 0.045 ± 0.020 | 17 ± 5 | 201 ± 35 |
| 300 | 0.024 ± 0.010 | 34 ± 13 | 470 ± 60 | |
| 400 | 0.014 ± 0.007 | 57 ± 26 | 886 ± 202 | |
| Tannic acid | 20 | 0.026 ± 0.007 | 28 ± 7 | 140 ± 2 |
| 30 | 0.024 ± 0.005 | 30 ± 6 | 223 ± 17 | |
| 40 | 0.043 ± 0.020 | 18 ± 7 | 655 ± 195 | |
| 50 | 0.009 ± 0.005 | 89 ± 52 | 986 ± 10 | |
| Gallic acid | 300 | 0.050 ± 0.023 | 17 ± 8 | 64 ± 5 |
| 450 | 0.043 ± 0.016 | 18 ± 8 | 85 ± 17 | |
| 600 | 0.031 ± 0.013 | 27 ± 13 | 116 ± 36 | |
| 750 | 0.034 ± 0.014 | 22 ± 7 | 139 ± 57 | |
| 900 | 0.024 ± 0.009 | 31 ± 9 | 280 ± 129 | |
| Vescalagin | 50 | 0.038 ± 0.024 | 22 ± 7 | 131 ± 11 |
| 100 | 0.021 ± 0.006 | 35 ± 11 | 165 ± 23 | |
| 150 | 0.020 ± 0.005 | 37 ± 12 | 206 ± 56 | |
| 200 | 0.017 ± 0.004 | 45 ± 17 | 343 ± 127 | |
| 250 | 0.017 ± 0.008 | 48 ± 23 | 460 ± 150 | |
| 300 | 0.017 ± 0.006 | 46 ± 19 | 684 ± 161 | |
| Castalagin | 200 | 0.029 ± 0.005 | 24 ± 5 | 175 ± 23 |
| 250 | 0.016 ± 0.004 | 45 ± 11 | 235 ± 46 | |
| 300 | 0.013 ± 0.003 | 58 ± 23 | 377 ± 144 | |
| 350 | 0.008 ± 0.003 | 95 ± 35 | 691 ± 290 | |
| 400 | 0.005 ± 0.003 | 199 ± 114 | 888 ± 279 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Štumpf, S.; Hostnik, G.; Primožič, M.; Leitgeb, M.; Bren, U. Generation Times of E. coli Prolong with Increasing Tannin Concentration while the Lag Phase Extends Exponentially. Plants 2020, 9, 1680. https://doi.org/10.3390/plants9121680
Štumpf S, Hostnik G, Primožič M, Leitgeb M, Bren U. Generation Times of E. coli Prolong with Increasing Tannin Concentration while the Lag Phase Extends Exponentially. Plants. 2020; 9(12):1680. https://doi.org/10.3390/plants9121680
Chicago/Turabian StyleŠtumpf, Sara, Gregor Hostnik, Mateja Primožič, Maja Leitgeb, and Urban Bren. 2020. "Generation Times of E. coli Prolong with Increasing Tannin Concentration while the Lag Phase Extends Exponentially" Plants 9, no. 12: 1680. https://doi.org/10.3390/plants9121680
APA StyleŠtumpf, S., Hostnik, G., Primožič, M., Leitgeb, M., & Bren, U. (2020). Generation Times of E. coli Prolong with Increasing Tannin Concentration while the Lag Phase Extends Exponentially. Plants, 9(12), 1680. https://doi.org/10.3390/plants9121680