2.1. Taxonomic Remarks
During the phytosociological investigation carried out in the high-mountains of Greece, we have collected several orophytes belonging to the genus Astragalus and Allium, which are very peculiar from the taxonomical point of view and traited as taxa new to science. Moreover, the taxonomic rank in some of them was modified. They are the following:
- (1)
Astragalus corinthiacus Brullo, Giusso & Musarella, sp. nov.
Holotype: Greece, Sterea Hellas, eastern slopes of Mt. Parnassus, on the bottom of carbonatic dolines with deep silt-clay soils, ca. 1800 m a.s.l., 07.VII.2006, S. Brullo, C.M. Musarella & G. Giusso del Galdo s.n. (CAT).
Diagnosis: Astragalo cephalonico affinis sed stipulis coriaceis, uninervatis, sparsim piloso-ciliatis dorsaliter, aristis triangularibus, 3–6 mm longis, foliolis lineari-ellipticis, 1–2.2 mm latis, viridibus, pubescentibus vel laxe lanuginosis, bracteis subulatis vel lineari-subulatis, dense ciliatis dorsaliter, numquam glabris margine, bracteolis praesentibus, tubo calice 4–4.5 mm longo et dentibus subequalibus, 9–10 mm longis, corolla roseo-purpurescenti, vessillo 16.5–18 mm longo, hastato, tubo staminorum 15 mm longo.
Description: Dwarf shrub forming a loose, spiny cushion, 30–60 cm tall. Stems woody, tomentose-lanuginose, with hairs 0.2–1.5 mm long, loosely branched, tough, with persistent stipules and rachis in the old parts of the branches. Stipules coriaceous, straw coloured, 8–12 mm long, usually 1-nervate, adnate to the petiole for 4.5–7 mm, ciliate at the margin, sparsely lanuginose dorsally, free part triangular, acuminate, 3–6 mm long. Leaves paripinnate, 2.5–4 cm long, with ivory rachis, covered by sparsely lanuginous hairs; petiole 8–20 mm long; terminal spine 3–5 mm long. Leaflets linear-elliptical, dark green, acuminate at the apex, 3–8.5 × 1–2.2 mm, more or less paired, covered by sparsely and appressed lanuginose hairs. Leaflet peduncle 0.2–0.4 mm long. Inflorescence crowded in subsessile racemes up to 8–10-flowered. Bracts subulate to linear-subutate, hyaline, usually curved dorsally, exceeding calyx tube, 8–10 mm long, 0.5–2 mm wide, dorsally ciliate-pilose, often glabrous laterally. Bracteoles subulatis up to 8 mm longis, cilate-pilose. Calyx cylindrical, white-hyaline, densely covered by rigid hyaline hairs 1–3 mm long, up to the teeth apex, tube 4–4.5 mm long, teeth subulate, subequal, 9–10 mm long. Corolla pink-purplish: standard hastate, 16.5–18 mm long, minutely emarginate, with blade 9–10 × 5.5–6 mm; wings 13–14 mm long, with blade 6–7 × 1.5–1.7 mm and auricle 0.6 mm long; keel 14–14.5 mm long. Staminal tube 15 mm long and free stamen 13 mm long; anthers o.8 mm long. Pistill 15–16 mm long; ovary 4–4.5 mm long, densely hairy; style hairy at the base. Pod 7 mm long, ellipsoid, densely pilose-appressed.
Etymology: From “Corinthus”, the Latin name of the city of Corinth and its gulf between Sterea Ellas and Peloponnese.
Distribution: The new species occurs in the mountain places of Mt. Parnassus and Mt. Giona where it is localized in the carbonatic dolines on silt-clay soils, mainly on the eastern and northern slopes at 1600–1900 m a.s.l.
Notes: This new species shows close relationships with
A. cephalonicus C. Presl, occurring in the Ionian islands of Cephalonia and Lefkada. In particular,
A. cephalonicus differs from
A. corinthiacus in having stipules membranaceous, linear-triangular, plurinerved, densely ciliate-hirsute, free part 5–10 mm long, leaflets oblong, up to 3 mm wide, greyish-green, densely villose, bracts ovate-lanceolate, long ciliate, 2.5–2.8 mm wide, bracteoles lacking, calyx with tube 5.5–7 mm long, teeth unequal, the three lower teeth 5.5–7 mm long, the upper two 7–9 mm long, corolla whitish to pinkish-white, standard spathulate with blade 13–16 × 5.5–6 mm, staminal tube 14 mm long. Previously Strid [
73] also pointed out that the populations of
A. cephalonicus of Cephalonia differed from those ones occurring in Sterea Ellas.
- (2)
Astragalus rumelicus Bunge, Mém. Acad. Imp. Sci. St.-Pétersbourg, Sér. 7. 15(1): 81 (1868)
- (a)
subsp. euboicus (Širj.) Brullo, Giusso & Musarella comb. et stat nov.
Bas.: Astragalus rumelicus var. euboicus Širj., Repert. Spec. Nov. Regni Veg. 47: 200. 1939.
- (b)
subsp. rumelicus
- (c)
subsp. taygeticus (Širj.) Brullo, Giusso & Musarella comb. et stat. nov.
Bas.: Astragalus rumelicus var. taygeticus Sirj., Repert. Spec. Nov. Regni Veg. 47: 199. 1939.
Syn.: Astracantha rumelica (Bunge) Reer & Podlech subsp. taygetica (Širj.) Reer & Podlech, Mitt. Bot. Staatssaml. Munchen 22: 544. 1986.
Notes: According to Širjaev [
79] the two subspecies differ from the type in some morphological characteristics. In particular, the subsp.
euboicus differs in having leaflets denser, outspread white-villous, calyx with short teeth, and corolla 11 mm long, while the subsp.
taygeticus apart from having leaflets denser outspread white-villous, is differentiated by a calyx with longer teeth, and corolla 13 mm long.
- (3)
Allium hirtovaginatum subsp. samium Brullo, Pavone & Salmeri, subsp. nov.
Holotype: Greece. Samos, Mt. Kerkis, esemplare coltivato, 22 July 1993, S. Brullo s.n. (CAT).
Diagnosis: A typo differt scapo usque ad 35 cm alto, foliis 5–6, pilis subadpressis 0.3–0.4 mm longis, florum pedicellis usque ad 7 cm longis, spatha 3.5–7(−9) cm longa, appendice usque ad 40 mm longa, perigonio 7–8 mm longo, tepalis e purpura superne albo-roseis, exterioribus lineari-lanceolatis, obtusiusculis vel rotundatis apice, 2–2.5 mm latis, staminum filamentibus subulato-triangulis, exterioribus usque ad 1.8 mm longis, interioribus 2–2.5 mm longis, annulo 1.2–1.4 mm alto, capsula 4.2 × 4 mm.
Description: Bulb ovoid, sometimes bulbiliferous, 15–20 × 8–12 mm, with brown tunics, fibrous slightly reticulate, split at the base, covering the stem up to 2 cm. Stem erect, flexuous 15–35 cm high, covered by the leaf sheaths 1/2–2/3 of its length. Leaves 5–6, filiform, semicylindrical, shorter than the inflorescence, 4–20 cm long, hairy with dense subappressed hairs 0.3–0.4 mm long. Inflorescence fastigiate, unilateral, with 5–10(−12) flowers on pedicels 1–5(−7) cm long. Spathe 1-valved, longer than the inflorescence or subequal, persistent, 9–11-nerved, 3.5–7(−9) cm long, with an appendage 15–40 mm long. Bostryces 2. Perigon cylindrical-suburceolate, 7–8 mm long; tepals white-pink, tinged with purple in the upper part, with a brown-purplish mid-vein, the outer linear-lanceolate, entire, subobtuse or rounded at the apex, 2–2.5 mm wide, the inner linear-oblong, rounded and feebly gnawed-undulate at the apex, 1.2–1.8 mm wide. Stamens with white filaments, yellowish below, subulate-triangular, unequal, the outer 0.9–1.8 mm long and 0.8–1 mm wide at the base, the inner 2–2.5 mm long and 1.2–1.5 mm wide at the base, below connate with tepals into an annulus 1.2–1.4 mm high; anthers straw coloured-yellowish, linear-elliptical, apiculate, 1.4 × 0.6 mm. Ovary greenish, subglobose-pyriform, smooth, 1.5–1.8 × 1.3–1.6 mm. Style white, 1–1.8 mm long. Capsule trivalved, subglobose, 4.2 × 4 mm.
Etymology: From Latin “Samius” = of Samos, Greek island of E Aegean area.
Distribution and habitat: It is exclusive of Samos, Aegean island near the Turkish coast. It grows in the semirupestrian stands, where it is frequent within ephemeral meadows placed among the phrygana, from sea level to submountain belt.
- (4)
Allium cremnophilum Brullo, Pavone & Salmeri, sp. nov.
Holotype: Greece. Thassos, Ipsario, 26 June 2003, S. Brullo & G. Giusso s.n. (CAT).
Diagnosis: Allio hirtovaginato simili sed bulbo bulbillifero, scapo flexuoso, prostrato-adscendentis, vaginis foliorum per 1/3–1/2 longitudinis tecto, pilis lanuginosis 0.5–1.4 mm longis, perigonio cylindrico-campanulato, tepalis 7.5–8 mm longis, staminum filamentibus omnino luteolis, exterioribus 1.4–2 mm long, interioribus 2.8–3.5 mm, ovario globoso-ovoideo, rugoso superne, 1–1.1 × 1.1–12 mm, capsula obovoidea, 3.7 × 3.4 mm.
Description: Bulb ovoid, often paired, bulbiliferous, 8–13 × 6–10 mm, with brown tunics, reticulate-fibrous, split at the base, covering the stem up to 2 cm. Stem flexuous, prostrate-ascending, 6–11 cm high, covered by the leaf sheaths 1/3–1/2 of its length, often bearing two inflorescences. Leaves 3, filiform, subcylindrical, longer than the inflorescence or subequal, 4–8 cm long, hairy-woolly with patent hairs 0.5–1.4 mm long. Inflorescence fastigiate, unilateral, with 4–8 flowers on pedicels 10–30 mm long. Spathe 1-valved, longer than the inflorescence or subequal, with 7 nerves of which 4 are incomplete, 8–32 mm long, with an appendage 5–20 mm long. Bostryces 2. Perigon cylindrical-campanulate, 7.5–8 mm long; tepals whitish to white-pinkish, with purplish mid-vein, the outer lanceolate, entire and acute at the apex, 1.8–2 mm wide, the inner linear-oblong, with purple striae above, subobtuse and gnawed-undulate the apex, 1.6–1.8 mm wide. Stamens with yellowish filaments, triangular-subulate, unequal, the outer 1.4–2 mm long and 0.5–0.8-mm wide at the base, the inner 2.8–3.5 mm long and 0.8–1 mm wide at the base, below connate with tepals into an annulus 0.8–1 mm high; anthers straw coloured, elliptical, apiculate, 1.5 × 0.8–0.9 mm. Ovary greenish, globose-ovoid, rugose above, 1–1.1 × 1.1–1.2 mm. Style white, 1.3–1.4 mm long. Capsule trivalved, obovoid, 3.7 × 3.4 mm.
Etymology: From the Greek words “cremnos” = crevice and “philos” = fond of, in reference with its habitat.
Distribution: At present, this species occurs only in Thassos at the top of Mt. Ipsario, a N Aegean island near Kavala, Greece. Usually, it grows in depth of calcareous crevices at c. 1200 m of altitude, mixed with chasmophytic vegetation or more rarely inside the thorny dwarf-shrubs of Astragalus angustifolius subsp. odonianus.
- (5)
Allium cylleneum Brullo, Pavone & Salmeri, sp. nov.
Holotype: GREECE. Peloponnisos, Mount Kyllini, 5 July 2006, S. Brullo, G. Giusso & C. Musarella s.n. (CAT).
Diagnosis: Allio cremnophilo simili sed bulbis aggregatis, tunicis fibrosis leviter reticulatis, usque ad 4 cm scapum tegentibus, scapo e vaginis foliorum per 3/4 longitudinis tecto vel totaliter, foliis usque ad 11 cm longis, pilis curvatis, subappressatis, 0.3–0.6 mm longis, spatha 5–7-nervata, nervis completis, perigonio campanulato-urceolato, tepalis 6.5–7 mm longis, omino eroso-undulatis et rotundatis superne, staminum filamentibus albidis, subulatis, ovario ovoideo, laeve, capsula 3.5 × 4 mm.
Description: Bulb ovoid, clustered, 12–20 × 8–12 mm, with pale brown tunics, fibrous with subparallel fibres feebly reticulate, split at the base, covering the stem up to 4 cm. Stem flexuous, erect or erect-ascending, 4–10 cm high, covered by the leaf sheaths from 3/4 of its lengt to totally. Leaves 3, filiform, subcylindrical, normally longer than the inflorescence, 6–11 cm long, hairy with curved subappressed hairs 0.3–0.6 mm long. Inflorescence fastigiate, unilateral, with 3–6 flowers on pedicels 5–25 mm long. Spathe 1-valved, shorter than the inflorescence or subequal, 5–7-nerved, 18–35 mm long, with an appendage 6–13 mm long. Bostryces 2. Perigon campanulate-urceolate with tepals white-pinkish, with purple striae and mid-vein, gnawed-undulate and rounded at the apex, 6.5–7 × 1.6–1.8 mm, the outer linear-lanceolate, the inner linear-elliptical. Stamens with white filaments, subulate, unequal, the outer 1.2–2 mm long and 0.6–0.8 mm wide at the base, the inner 2.8–3.3 mm long and 0.7–1 mm wide at the base, below connate with tepals into an annulus 1–1.2 mm high; anthers straw coloured, ovate-elliptical, apiculate, 1.3–1.4 × 0.7–0.8 mm. Ovary yellow-greenish, ovoid, smooth, 1–1.1 × 1.1–1.2 mm. Style white, 1.2–1.3 mm long. Capsule trivalved, obovoid, 3.5 × 4 mm.
Etymology: from Latin “cylleneus” = from Mt. Kyllini (N Peloponnese).
Distribution: At present, this species seems confined to the top of Mt. Profitis Ilias, the highest summit of Kyllini massif in N Peloponnese (Greece). It is quite rare and occurs in the orophilous dwarf-shrub communities with Astragalus rumelicus subsp. taygeticus on Mesozoic limestone, at 2200–2400 m of altitude.
- (6)
Allium orosamium Brullo, Giusso & Musarella, sp. nov.
Holotype: Greece, Island of Samos, Mt. Kerkis, near the top at 1100 m a.s.l., 02/07/2003, S. Brullo s.n. (CAT).
Diagnosis: Allio stamineo simili sed tunicis bulborum fibroso-coriaceis, scapo usque ad 27 cm alto, spathis 5–6-nervatis, inflorescentia 20–35 floribus, perigonio campanulato, tepalis brunneo-viridibus, max. 5 mm longis, 2–2.2 mm latis, staminum filamentibus albidis, 3.5–4.5 mm longis, ovario obovoideo, papilloso, 3.2–3.5 × 2.2–2.4 mm, stilo 0.5 mm longo, capsula subglobosa 5 × 5.2 mm.
Description: Bulb ovoid, 12–15 × 7–9 mm, with outer tunics fibrous-coriaceous, dark brown, the inner ones membranous, whitish. Scape glabrous, erect, 9–27 cm high, covered by leaf sheaths for 1/2–2/3 of its length. Leaves 3–5, green, semicylindrical, costate, with blade 10–20 cm long. Spathe persistent, with 2 unequal valves, longer than umbel, the larger 5–6-nerved, 3–8 cm long, the smaller 5–6-nerved, 2–5 cm long. Inflorescence lax, diffuse, 20–35-flowered; pedicels unequal, flexuous, 7–20 mm long. Perigon campanulate, with tepals unequal, brownish green tinged with brown-purplish, oblong, rounded at apex, the outers 4.5–4.8 × 2.2 mm, the inners 4.8–5 × 2–2.1 mm. Stamens simple, exserted, with filaments subulate, 3.5–4.5 mm long, white, connate at base into an annulus 0.6–1 mm high; anthers oblong, straw, rounded at apex, 1.2 × 0.7 mm. Ovary obovoid, yellow-greenish, papillose above, 3.2–3.5 × 2.2–2.4 mm. Style white, 0.5 mm long. Capsule widely subglobose, green, 5 × 5.2 mm.
Etymology: From “oros” Greek name of “mountain” and “Samius” Latin adjective of Samos (Aegean Island).
Distribution: This species is localized in the top of Mt. Kerkis (Samos island), where it grows in the carbonatic rocky stands within the community characterized by Astragalus creticus subsp. samius.
- (7)
Allium karvounis Brullo, Giusso & Musarella, sp. nov.
Greece, Island of Samos, Mt. Ambelos, near the top at 1100 m a.s.l., 11/06/2005, S. Brullo & C.M. Musarella s.n. (CAT).
Diagnosis: Allio stamineo simili sed bulbis maioribus, tunicis fibroso-coriaceis, scapis minoribus e vaginis foliorum per 1/2 longitudinis tectis, lamina foliorum rigida, spathis brevioribus, inflorescentia usque ad 80 floribus, pedicellis usque ad 40 mm longis, tepalis minoribus 3.8–4 × 1.6–1.8 mm, staminum filamentibus brevioribus, ovario obovoideo, laeviter papilloso, maiore, stilo 2–6 mm longo, capsula maiore.
Description: Bulb ovoid, 10–15 × 8–12 mm, with outer tunics fibrous-coriaceous, dark brown, the inner ones membranous, whitish. Scape glabrous, erect, 18–24 cm high, covered by leaf sheaths for 1/2 of its length. Leaves 3–4, green, semicylindrical, costate, with blade rigid, 8–20 cm long. Spathe persistent, with 2 unequal valves, longer than umbel, the larger 7-nerved, 4–7 cm long, the smaller 5–7-nerved, 2–4 cm long. Inflorescence fastigiate, compact, 25–80-flowered; pedicels unequal, flexuous, 8–40 mm long. Perigon conic-campanulate, with tepals equal, greenish yellow tinged with purplish, oblong, rounded at apex, 3.8–4 × 1.6–1.8 mm. Stamens simple, exserted, with filaments subulate, 3.5–5 mm long, white below, purplish above, connate at base into an annulus 0.4–0.5 mm high; anthers oblong, yellow, rounded at apex, 1.2–1.3 × 0.8–0.9 mm. Ovary obovoid slightly throttled, green, slightly papillose above, 1.8–2 × 1.8–2 mm. Style white, 2–6 mm long. Capsule obovoid, green, 4.5–5 × 4.5–5 mm.
Etymology: From “Karvounis” old name of Ambelos mount from Samos (Aegean Island).
Distribution: This species is localized in the top of Mt. Ambelos from Samos island in the Aegean area, where it grows into the orophilous cushion-like vegetation.
- (8)
Allium lefkadensis Brullo, Giusso & Musarella, sp. nov.
Holotype: Greece, Lefkàda, Ionian Islands, Mt. Elati, near the top at 1000 m a.s.l., 16/07/2011, S. Brullo & G. Giacalone s.n. (CAT).
Diagnosis: Allio stamineo simili sed tunicis interioribus bulborum brunneo-purpurescentibus, scapis minoribus e vaginis foliorum per 1/4–1/3 longitudinis tectis, lamina foliorum 8–16 mm, spathis brevioribus, tepalis minoribus, pruinosis, staminum filamentibus supra roseam suffusis, ovario maiore, stilo longiore.
Description: Bulb ovoid, 15 × 10 mm, with outer tunics coriaceous, dark brown, the inner ones membranous, reddish-brown. Scape glabrous, erect, 10–16 cm high, covered by leaf sheaths for 1/4–1/3 of its length. Leaves 4, green, semicylindrical, costate, 8–16 cm long. Spathe persistent, with 2 unequal valves, longer than umbel, the larger 7-nerved, 3–4 cm long, the smaller 5-nerved, 1.5–2 cm long. Inflorescence lax, diffuse, 20–25-flowered; pedicels unequal, flexuous, 10–25 mm long. Perigon conical-campanulate, with tepals equal, greenish yellow pruinose, oblong, rounded at apex, 4.5 × 2 mm. Stamens simple, exserted, with filaments subulate, 6–7 mm long, white below and slightly tinged with pink above, connate at base into an annulus 0.5–0.6 mm high; anthers oblong, straw, apiculate at apex, 1.2–1.4 × 0.6–0.7 mm. Ovary subglobose, yellow-greenish, slightly rugose-papillose above, 2 × 2–2.1 mm. Style white, 2.5–6 mm long. Capsule not observed.
Etymology: From “Lefkàda”, the Greek Ionian island where this species is confined.
Distribution: The species was observed only on the top of Mt. Elati at Lefkàda Ionian Island.
2.3. Phytosociological Investigation
Previously, the orophilous pulvinate vegetation of central-southern and insular Greece hitherto known in literature were included in
Daphneeto-Festucetea class as described by Quézel [
35]. Within this class, the associations were arranged according to the syntaxonomical scheme proposed by that author, afterwards modified by Quézel et al. [
80]:
DAPHNO OLEOIDIS-FESTUCETEA VARIAE Quézel 1964, corr. Quézel et al. 1992
Syn.: Daphneeto-Festucetea Quézel 1964, Vegetatio, 12:325
Lectotypus: Daphno oleoidis-Festucetalia variae Quézel 1964
DAPHNO OLEOIDIS-FESTUCETALIA VARIAE Quézel 1964, corr. Quézel et al. 1992
Syn.: Daphneeto-Festucetalia Quézel 1964, Vegetatio, 12:325
Lectotypus: Eryngio multifidi-Bromion fibrosi Quézel 1964
STIPO PULCHERRIMAE-MORINION PERSICAE Quézel 1964, corr. Quézel et al. 1992
Syn.: Stipeto-Morinion Quézel 1964, Vegetatio, 12: 326
Lectotypus: Scabioso taygeteae-Onosmetum leptanthae Quézel 1964
Scabioso taygeteae-Onosmetum leptanthae Quézel 1964 Vegetatio, 12:327
Syn.: Ass. à Scabiosa taygetea et Onosma leptanthum Quézel 1964
Galio lucidi-Ribetum uvae-crispae Quézel 1964, Vegetatio, 12:329
Syn.: ass. à Galium lucidum et Ribes uva-crispa Quézel 1964
Onobrychido minoris-Juniperetum foetidissimae Quézel 1973, Biol. Gallo-Hell. 5(1):147
Syn.: ass. à Juniperus foetidissima et Onobrychis ebenoides var. minor Quézel 1973
Juniperetum foetidissimae Georgiadis & Dimopoulos 1993, Bot. Helv. 103:152 (nom. inval.)
scabiosetosum ochroleucae Maroulis & Georgiadis 2005, Fitosociologia 42(1):37
Acer monspessulano-Prunetum mahaleb Georgiadis & Dimopoulos 1993, Bot. Helv. 103:153 (nom. inval.)
Astracantho thracicae-Marrubietum cyllenei Georgiadis & Dimopoulos 1993, Bot. Helv. 103:153 (nom. inval.)
galietosum taygetei Georgiadis & Dimopoulos 1993, Bot. Helv. 103:153 (nom. inval.)
Stipa pennata subsp. pulcherrima-Sesleria vaginalis comm. Maroulis & Georgiadis 2005, Fitosociologia 42(1):42
Hippocrepis comosa-Stipa pennata subsp. pulcherrima comm. Maroulis & Georgiadis 2005, Fitosociologia 42(1):43
ERYNGIO MULTIFIDI-BROMION FIBROSI Quézel 1964, corr. Quézel et al. 1992
Syn.: Eryngieto-Bromion Quézel 1964, Vegetatio, 12:326
Lectotypus: ass. à Astragalus cylleneus et Cirsium cylleneum, Quézel 1964
Sideritetum theezantis Quézel 1964, Vegetatio, 12:331
Syn.: ass. à Sideritis theezans Quézel 1964
Cirsio cyllenei-Astragaletum cyllenei Quézel 1964, Vegetatio, 12:332
Syn.: ass. à Astragalus cylleneus et Cirsium cylleneum Quézel 1964
Marrubio velutini-Astragaletum cretici Quézel 1964, Vegetatio, 12:334
Syn.: ass. à Astragalus creticus ssp. rumelicus et Marrubium velutinum Quézel 1964
Astracantho thracicae-Marrubietum cyllenei Georgiadis & Dimopoulos 1993, Bot. Helv. 103:153, nom. inval.
typicum Georgiadis & Dimopoulos 1993, Bot. Helv. 103: 158 (nom. inval.)
festucetosum cyllenecae Georgiadis & Dimopoulos 1993, Bot. Helv. 103:158 (nom. inval.)
Marrubio cyllenei-Astragaletum rumelici Maroulis & Georgiadis 2005, Fitosociologia 42(1):43
Festuco politae-Festucetum cyllenicae Maroulis & Georgiadis 2005, Fitosociologia 42(1):44
ASTRAGALO ANGUSTIFOLII-SESLERION COERULANTIS Quézel 1964, corr. Quézel et al. 1992.
Syn.: Astragaleto-Seslerion Quézel 1964, Vegetatio, 12:326
Lectotypus: ass. à Minuartia stellata et Erysimum parnassi Quézel 1964, Vegetatio, 12:326
Rindero graecae-Acantholimetum graeci Quézel 1964, Vegetatio, 12:336
Syn.: ass. à Acantholimon echinus et Rindera graeca Quézel 1964
Asteri cyllenei-Globularietum stygiae Quézel 1964, Vegetatio, 12:337
Syn.: ass. à Aster cylleneus et Globularia stygia Quézel 1964
Convolvulo cochlearis-Astragaletum lactei Quézel 1964, Vegetatio, 12:339
Syn.: ass. à Convolvulus cochlearis et Astragalus lacteus Quézel 1964
Erysimo parnassi-Minuartietum stellatae Quézel 1964, Vegetatio, 12:340
Syn.: ass. à Minuartia stellata et Erysimum pusillum ssp. parnassi Quézel 1964
Paronychio chionaeae-Thymetum ciliato-pubescentis Quézel 1964, Vegetatio, 12:341
Syn.: ass. à Paronychia chionaea et Thymus hirsutus ssp. ciliato-pubescens Quézel 1964
Violo-Seslerietum vaginalis Quézel 1973, Biol. Gallo-Hell. 5(1):152
Syn.: ass. à Sesleria coerulans et Viola stojanowii Quézel 1973
Euphrasio salisburgensis-Asperuletum nitidae Quézel 1974, Rev. Biol. Ecol. Medit. 1(1):19
Syn.: ass. à Asperula nitida et Euphrasia salisburgensis Quézel 1974
Festuco cyllenicae-Asperuletum boissieri Georgiadis & Dimopoulos 1993, Bot. Helv. 103: 158, nom. inval.
Within this hierarchical arrangement proposed by Quézel [
35], the most relevant aspect that emerges from this classification was to use only the altitudinal distribution of plant communities as a discriminating criterion for alliance identification. In fact, according to this author, the order
Daphno-Festucetalia includes three alliances which are widespread in all the mountains of Greece and are distributed exclusively at different altitudinal ranges. They are: (a)
Stipeto-Morinion occurring between 1500 and 1700 m; (b)
Eryngieto-Bromion between 1700 and 2200 m; (c)
Astragaleto-Seslerion above 2200 m, sloping down sometimes up to 1700 m. Another important factor to note is that these alliances do not provide any information on the real phytogeographic role of the rich floristic contingent featuring this type of orophilous vegetation. Indeed, Quézel [
35] considered as characteristics of these alliances mainly species having a wide East Mediterranean or even circum-Mediterranean distribution, showing also a wide altitudinal range and not limited to a narrow belt as stated by the author. In particular, the author proposed
Stipa endotricha (=
S. pennata var.
pulcherima),
Melica ciliata, Asphodeline lutea, Ononis pusilla, Morinia persica, Scutellaria rupestris (=
S. peregrina subsp.
rupestris),
Pterocephalus perennis and
Anthemis spruneri (=
A. montana var.
incana) as characteristics of
Stipeto-Morinion; while
Bromus riparius (=
B. fibrosus),
Helictotrichon aetolicum (=
Avena australis),
Eryngium multifidum, Thymus sibthorpii, Galium thymifolium, Campanula spathulata, Podosmermum canum var.
alpinum, and
Carduus tmoleus (=
C. armatus), as characteristics of
Eryngieto-Bromion; finally,
Sesleria tenerrima (=
S. coerulans),
Iberis sempervirens, Astragalus angustifolius, Draba lasiocarpa (=
D. affinis),
Viola graeca (=
Viola heterophylla subsp.
graeca),
Trinia frigida (
Apinella frigida)
, Trinia guicciardii (
Apinella guicciardii)
, Acantholimon graecum (= A. echinus), Lactuca intricata (=
L. graeca)
, Veronica orsiniana subsp.
teucrioides (=
V. austriaca var.
teucrioides)
, V. thessalica, V. thymifolia, Asperula boissieri, and
Tragopogon crocifolius subsp.
samaritani as characteristics of
Astragaleto-Seslerion. On the basis of literature and personal observations, these taxa can not be used to characterize alliances, at most, some of them may be included among the characteristics of order or class, while others are simply accidentals or ubiquitous species. Even, the same author [
35,
36] underlined often some perplexity in the inclusion of a given association in one of the three alliances identified by him, due to the contemporaneous occurrence in the relevés of characteristic species belonging to all three alliances. Therefore, the alliances identified by Quézel [
35] are not being characterized by exclusive species, since they include ubiquitous or species of wider ecological requirements, that are not strictly related to those specific habitats; in this way they do not provide clear information from an ecological and phytogeographical point of view. Based on the above, these alliances do not satisfy the prerequisites required by the sigmatist phytosociological method. They only create a lot of confusion and ambiguity in the syntaxonomical arrangement of this very peculiar kind of orophilous vegetation. In conclusion, these alliances are really ambiguous names that must be rejected (art. 36). Therefore, a new phytosociological framework is necessary to propose. The designation of new alliances must be essentially based on the phytogeographic criteria and such characteristics must include steno-endemic species in order to define unequivocally the geographical boundaries of each syntaxon as well as its syntaxonomical role.
In order to emphasize the distribution of characteristic species within the three alliances and syntaxa of higher rank according to the hierarchic arrangement proposed by Quézel [
35], a synoptic table (
Appendix B,
Table A2) was processed including all the phytosociological relevés published until now on this type of orophilous vegetation in central-southern Greece by Quézel [
35,
38] and Quézel and Katrabassa [
40], as well as other later authors as Georgiadis and Dimopoulos [
42] and Maroulis and Georgiadis [
44]. From the analysis of this table, the floristic comparison among the hitherto recognized associations, which are well differentiated from the phytosociological viewpoint, shows clearly that the species proposed as characteristics of the alliances are distributed indifferently in all three syntaxa, often with high frequency values. Therefore, it can be easily deduced that a single association cannot be clearly and unambiguously attributed to a specific alliance. Quézel [
35] in order to attribute an association to a given alliance, he relied mainly on its altitudinal distribution, rather than considering the information relating to its floristic cortege. Unfortunately, the species selected by the author to define these alliances are not strictly linked to well-defined altitudinal bands, but are widespread almost at all altitudes. From this, it can easily be deduced that, in the case of the orophilous pulvinate vegetation of the Greek mountains, as well as of other geographic territories, this criterion can not be followed. Instead, a purely phytogeographical method must be selected, mainly based on endemic flora, that gives more significant information under phytosociological feature.
On the basis of several unpublished phytosociological relevés carried out by us in the summit stands of most of central and southern Greek mountains as well as in some islands (
Figure 1), it was possible to verify that only a strictly phytogeographic policy can allow for a correct syntaxonomic arrangement of these communities, similar to what has been achieved for other Mediterranean territories [
22,
23,
24,
25,
26,
27,
28,
29,
30,
31,
32,
33,
34,
35,
36,
37,
38,
39,
40,
41,
42,
43,
44,
45,
46,
47,
48]. In fact, it is much more realistic and meaningful to identify alliances based on floristic elements that give clear information on phytogeographic correlations of the various associations, rather than on their altitudinal distribution. In particular, the flora characterizing the orophilous community usually shows a significant richness in relict species, often very isolated, or represented by geographical vicariants of remarkable phytosociological significance. Therefore, for a syntaxonomic arrangement that can best express the floristic and structural organization of the pulvinate-orophilous plant communities currently occurring in the Greek mountains, it has to be based on the choice of species suitable for providing more precise information on their phytogeographical role. Following this viewpoint, this study presents a clearer and more comprehensive syntaxonomical overview of these plant communities, reflecting their origin and diversification. Therefore, for a correct floristic characterization of higher syntaxa (alliances, orders and classes) allowing differentiation of specific alliances, the choice should fall on endemics with restricted distribution, such as those confined to one or few neighbouring or close mountain ranges, and it should gradually move on to those endemics with wider ranges and the other more widespread taxa which should be used for the designation of orders and classes. In addition, the floristic contingent that differentiates the higher syntaxa, and particularly in the case of orophilous vegetation featuring the Mediterranean mountains, provides clearer information about the relationships that the plant communities show among them, since they are the result of paleogeographic vicissitudes of the territories that host them.
Furthermore, it must be emphasized that Quézel et al. [
80] when lectotypified the class
Daphno-Festucetea and the corresponding order
Daphno-Festucetalia, corrected respectively the two names in
Daphno oleoidis-Festucetea variae and
Daphno oleoidis-Festucetalia variae. The use of
Daphne oleoides and
Festuca varia for giving the name to the two syntaxa brings further confusion and ambiguity, since both species are not pertinent to this type of vegetation. In fact,
Daphne oleoides is widespread in all Mediterranean mountains and is considered a typical characteristic species of the class
Junipero-Pinetea sylvestris Rivas-Martínez 1964, as emphasized by Rivas-Martínez [
52], Rivas-Martínez et al. [
53,
54,
55,
80,
81,
82], Stanisci [
56] and Brullo et al. [
57], while in the pulvinate dwarf shrub vegetation it is rather rare and occasional. As concerns
Festuca varia, this species has a properly alpine distribution and is totally absent in Greece [
83], where it is replaced by various other species of this genus. Moreover, it is not possible to identify in a univocal and correct way what is the species of
Festuca to which Quézel [
35] refers in naming these syntaxa.
Besides, among the species proposed by Quézel [
35] as characteristic of the class and order is to be noted that some of them, such as
Juniperus communis var.
hemisphaerica,
Berberis cretica, Prunus prostrata and mainly
Daphne oleoides, are linked to the orophilous communities characterized by phanerophytes and nanophanerophytes belonging to the class
Junipero-Pinetea sylvestris Rivas-Martínez 1965 nom. invers. propos. (=
Pino-Juniperetea Rivas-Martínez 1965). This is in agreement with the literature data concerning this type of orophilous forest vegetation [
55,
56,
57,
63,
84].
In particular as emphasized by Brullo et al. [
57] and Mucina et al. [
84], the woody communities characterized by the dominance of erect or prostrate conifers occurring in Greece and other central-eastern Mediterranenan territories, must be ascribed to syntaxa exclusive to these mountaints, representated by the order
Berberido creticae-Juniperetalia excelsae Mucina in Mucina et al. 2016 and some alliances, such as
Berberido aetnensis-Pinion laricionis (Brullo et al. 2001) Mucina & Theurillat in Mucina 2016,
Juniperion excelso-foetidissimae Em ex Matevski et al. 2010,
Berberido creticae-Juniperion foetidissimae Brullo et al. 2001, etc. These forest communities are relegated to the supra- and oro-Mediterranean belts, as well as supra-temperate belt, where they show a fragmentary distribution, which confirms their relict origin. Usually, they occupy an intermediate position between the typical mountain forests of
Querco-Fagetea and pulvinate orophilous dwarf shrubs linked to cacuminal stands.
Besides as emphasized by Brullo et al. [
57], some associations of
Daphno-Festucetea described by the previous authors must be rather clearly attributed to the class
Junipero-Pinetea sylvestris, since they show a floristic, structural and ecological feature of the last syntaxon. In particular, this is the case of the “ass. à
Galium lucidum et
Ribes uva-crispa Quézel 1964”, “ass. à
Juniperus foetidissima et
Onobrychis ebenoides var.
minor Quézel 1973”, “
Juniperetum foetidissimae Georgiadis & Dimopoulos 1993”, “
Acer monspessulano-Prunetum mahaleb Georgiadis & Dimopoulos 1993”, contributing further to confer a marked ambiguity to the class
Daphno-Festucetea.
For the reasons above mentioned, the names Daphno-Festucetea Quézel 1964 and Daphno-Festucetalia Quézel 1964 must be proposed as nomina ambigua rejicienda (Art. 36), since they are based on very ambiguous alliances, are sources of continuous errors in the univocal and unambiguous designation of the relative associations. The new names proposed here in order to replace those of the two aforesaid syntaxa are Cerastio candidissimi-Astragaletea rumelici and Eryngio multifidi-Armerietalia orphanidis, both having a large distribution in the high mountains of southern Balkans and Aegean area.
The floristic analysis of the investigated plant communities occurring mainly in the high mountains of the Peloponnese and Sterea Ellas, as well as in some Ionian Islands and Euboea, showed the existence of significant sets of endemic species, which have a well-defined geographical distribution that allows the identification of alliances based on a clear phytogeographical role, emphasizing especially the palaeogeographical isolation of the various mountain areas among them.
Based on these criteria, it was possible to distinguish in the aforesaid territories some new alliances, which are well circumscribed from the phytogeographical point of view and allow a very realistic arrangement of the orophilous dwarf shrubby vegetation occurring in these Greek high mountains, emphasizing their floristic affinities. These are: Marrubio velutini-Thymion parnassici, distributed in the mountains of Sterea Hellas and Attica; Festuco achaicae-Marrubion cyllenei, from the North Peloponnese mountains; Sideritido clandestinae-Asperulion mungieri, from South Peloponnese mountains. Moreover, Astragalion cephalonici, from the Ionian islands of Cephalonia and Lefkada, as well as Astragalion euboici from the island of Euboea, must be added to these alliances.
In order to highlight that these alliances have a clear phytosociological role with a well-defined phytogeographic boundary than those proposed by Quézel [
35], the associations examined in
Appendix B,
Table A2 were processed according to this new syntaxonomic scheme. As can be clearly observed in the new
Table A3 (
Appendix B), the associations fall within floristically well-differentiated alliances, since they are characterized by endemics exclusive of geographically distinct areas, which are characterized by very similar paleogeographic vicissitudes.
In addition, further phytosociological investigations were carried out in the high mountains of some islands of North Aegean area (Thassos, Lesbos, Chios, and Samos) peaking over 1000 m. a.s.l and hosting this kind of vegetation. Within the orophilous pulvinate dwarf shrubs communities occurring in these islands, some characteristic species of Cerastio candidissimi-Astragaletea rumelici class are still present (although numerically reduced), while species belonging to the Eryngio multifidi-Armerietalia orphanidis order and related alliances are fully missing.
In these insular high-mountain areas, there is a rich set of endemics or eastern Aegean taxa, which allow to differentiate a new vicariant order, namely Noaeo mucronate-Silenetalia urvillei. On essential phytogeographical basis, it is possible to distinguish three floristically well-differentiated alliances, represented by Asperulion samiae, circumscribed to Samos, Festuco pseudosupinae-Astragalion aegeici, distributed to Chios and Lesbos, and Seslerio achtarovii-Anthemidion tenuilobae, from Thassos. Based on the observations above emphasized, a new syntaxonomic scheme is proposed:
CERASTIO CANDIDISSIMI-ASTRAGALETEA RUMELICI Musarella, Brullo & Giusso cl. nov.
ERYNGIO MULTIFIDI-ARMERIETALIA ORPHANIDIS Musarella, Brullo & Giusso ord. nov.
MARRUBIO VELUTINI-THYMION PARNASSICI Musarella, Brullo & Giusso all. nov.
Marrubio velutini-Astragaletum rumelici Quézel 1964
typicum
achilleetosum nobilis Quézel 1964
Astragalo lactei-Convolvuletum cochlearis Quézel 1964
Nepeto epiroticae-Astragaletum corynthiaci (Quézel 1964) Musarella, Brullo & Giusso nom. nov.
Nepeto spruneri-Astragaletum corynthiaci Musarella, Brullo & Giusso ass. nov.
Thymo parnassici-Paronychietum polygonifoliae Quézel 1964
typicum
linetosum angustifolii Quézel 1964
Nepeto sprunerii-Astragaletum tymphrestei Musarella, Brullo & Giusso ass. nov.
Violo stojanowii-Seslerietum vaginalis Quézel 1973
Erysimo parnassi-Minuartietum stellatae Quézel 1964
Aurinio gionae-Minuartietum stellatae Musarella, Brullo & Giusso ass. nov.
Achilleo fraisii-Dianthetum tymphrestei Musarella, Brullo & Giusso ass. nov.
Asperulo luteae-Achilleetum umbellatae Musarella, Brullo & Giusso ass. nov.
Astragalo lactei-Asperuletum apiculatae Musarella, Brullo & Giusso ass. nov.
Diantho minutiflori-Festucetum cyllenicae Musarella, Brullo & Giusso ass. nov.
Scabioso ochroleucae-Sideridetum raeseri Musarella, Brullo & Giusso ass. nov.
Ranunculo psilostachydis-Festucetum cyllenicae Musarella, Brullo & Giusso ass. nov.
Edraiantho parnassici-Globularietum cordifoliae Musarella, Brullo & Giusso ass. nov.
Thymo parnassici-Astragaletum parnassi Musarella, Brullo & Giusso ass. nov.
Chamaecytiso hirsuti-Astragaletum parnassi Musarella, Brullo & Giusso ass. nov.
Onobrychido pentelicae-Genistetum parnassicae Musarella, Brullo & Giusso ass. nov.
Allio cithaeronis-Dianthetum serratifolii Musarella, Brullo & Giusso ass. nov.
Inulo methaneae-Sideritetum atticae Musarella, Brullo & Giusso ass. nov.
ASTRAGALION CEPHALONICI Musarella, Brullo & Giusso all. nov.
Helictotricho convoluti-Thymetum holosericei Musarella, Brullo & Giusso ass. nov.
Saturejo cuneifoliae-Thymetum holosericei Musarella, Brullo & Giusso ass. nov.
Scutellario cephalonicae-Astragaletum cephalonici Musarella, Brullo & Giusso ass. nov.
Paronychio graecae-Astragaletum erinacei Musarella, Brullo & Giusso ass. nov.
ASTRAGALION EUBOICI Musarella, Brullo & Giusso all. nov.
Sideritido euboeae-Astragaletum euboici Musarella, Brullo & Giusso ass. nov.
Scabioso webbianae-Phlomidetum samiae Musarella, Brullo & Giusso ass. nov.
Sideritido euboeae-Festucetum cyllenicae Musarella, Brullo & Giusso ass. nov.
Inulo limonellae-Seslerietum krajinae Musarella, Brullo & Giusso ass. nov.
FESTUCO ACHAICAE-MARRUBION CYLLENEI Musarella, Brullo & Giusso all. nov.
Cirsio hypopsilii-Astragaletum taygetici Quézel 1964 corr.
Astero cyllenei-Globularietum stygiae Quézel 1964
Euphrasio salisburgensis-Asperuletum oetaeae Quézel & Katrabassa 1974 corr.
Marrubio cyllenei-Astragaletum calavrytensis Musarella, Brullo & Giusso ass. nov.
elytrigietosum intermediae Musarella, Brullo & Giusso subass. nov.
hippocrepidetum comosae Musarella, Brullo & Giusso subass. nov.
tulipetosum australis Musarella, Brullo & Giusso subass. nov.
Plantagini graecae-Astragaletum cyllenei Musarella, Brullo & Giusso ass. nov.
Festuco achaicae-Minuartietum stellatae Musarella, Brullo & Giusso ass. nov.
Alysso taygetei-Plantaginetum alpestris Musarella, Brullo & Giusso ass. nov.
Hieracio sartoriani-Seslerietum tenerrimae Musarella, Brullo & Giusso ass. nov.
Asperulo boissieri-Festucetum cyllenicae Georgiadis & Dimopoulos ass. nov.
Ranunculo brevifolii-Seslerietum tenerrimae Musarella, Brullo & Giusso ass. nov.
Astragaletum hellenico-erinacei Musarella, Brullo & Giusso ass. nov.
Festucetum polito-cyllenicae Maroulis & Georgiadis 2005
Arenario filicaulis-Festucetum cyllenicae Musarella, Brullo & Giusso ass. nov.
Aurinio moreanae-Lomelosietum crenatae Musarella, Brullo & Giusso ass. nov.
Onosmo malickyi-Astragaletum hellenici Musarella, Brullo & Giusso ass. nov.
Violo graecae-Festucetum cyllenicae Musarella, Brullo & Giusso ass. nov.
Tripodio graeci-Helictotrichetum heldreichii Musarella, Brullo & Giusso ass. nov.
SIDERITIDO CLANDESTINAE-ASPERULION MUNGIERI Musarella, Brullo & Giusso all. nov.
Scabioso taygeteae-Onosmetum leptanthae Quézel 1964
Danthoniastro compacti-Fumanetum alpinae Musarella, Brullo & Giusso ass. nov.
Sideritido clandestinae-Astragaletum taygetici Musarella, Brullo & Giusso ass. nov.
Rindero graecae-Acantholimetum graeci Quézel 1964
Onosmo heterophyllae-Astragaletum erinacei Musarella, Brullo & Giusso ass. nov.
Astragaletum lacteo-taygetici Musarella, Brullo & Giusso ass. nov.
Violo parnoniae-Astragaletum erinacei Musarella, Brullo & Giusso ass. nov.
astragaletosum erinacei Musarella, Brullo & Giusso subass. nov.
asperuletosum malevonensis Musarella, Brullo & Giusso subass. nov.
NOAEO MUCRONATAE-SILENETALIA URVILLEI Musarella, Brullo & Giusso ord. nov.
ASPERULION SAMIAE Musarella, Brullo & Giusso all. nov.
Astragaletum samii Musarella, Brullo & Giusso ass. nov.
Thymo samii-Astragaletum condensati Musarella, Brullo & Giusso ass. nov.
Campanulo lyratae-Genistetum parnassicae Musarella, Brullo & Giusso ass. nov.
Arenario guicciardii-Seslerietum anatolicae Musarella, Brullo & Giusso ass. nov.
FESTUCO PSEUDOSUPINAE-ASTRAGALION AEGEICI Musarella, Brullo & Giusso all. nov.
Anthemido discoideae-Astragaletum aegeici Musarella, Brullo & Giusso ass. nov.
Diantho zonati-Astragaletum lesbiaci Musarella, Brullo & Giusso ass. nov.
Galio insularis-Thymetum sypilei Musarella, Brullo & Giusso ass. nov.
Acantholimo aegaei-Astragaletum lesbiaci Musarella, Brullo & Giusso ass.nov.
SESLERIO ACHTAROVII-ANTHEMIDION TENUILOBAE Musarella, Brullo & Giusso all.nov.
Paronychio bornmuelleri-Astragaletum odoniani Musarella, Brullo & Giusso ass. nov.
Finally, in order to highlight the phytosociological relantionships among the investigated associations belonging to
Cerastio candidissimi-Astragaletea rumelici, two synoptical tables regarding the orders
Eryngio multifidi-Armerietalia orphanidis (
Appendix B,
Table A4) and
Noaeo mucronatae-Silenetalia urvillei (
Appendix B,
Table A5) are provided.
2.4. Description of the Vegetation
Syn.: Daphneeto-Festucetea Quézel 1964, Vegetatio 12:325, p.p., nom. amb. rejic. propos. (art. 36)
Daphno oleoidis-Festucetea variae Quézel 1964, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18: 82, p.p., nom. amb. rejic. propos. (art. 36)
Holotypus: Eryngio multifidi-Armerietalia orphanidis Musarella, Brullo & Giusso ord. nov. hoc loco.
Characteristic species: Achillea umbellata, Alyssum montanum subsp. graecum Arenaria guicciardii, Asperula boissieri, Asperula lutea, Asperula thessala, Astragalus rumelicus subsp. rumelicus, Beta nana, Campanula radicosa, Centaurea pichleri, Centaurea raphanina subsp. mixta, Cerastium candidissimum, Crepis fraasii subsp. fraasii, Dianthus tymphristeus, Draba lacaitae, Erysimum cephalonicum, Erysimum microstylum, Erysimum pectinatum, Festuca cyllenica subsp. cyllenica, Festuca polita, Fritillaria graeca, Fritillaria guicciardii, Galium citraceum, Galium thymifolium, Helianthemum hymettium, Herniaria parnassica subsp. parnassica, Hieracium lazistanum subsp. Leithneri, Lamium pictum, Leontodon graecus, Lysimachia serpyllifolia, Minuartia confusa, Minuartia attica subsp. attica, Nepeta argolica subsp. argolica, Paronychia albanica subsp. graeca, Poa thessala, Podospermum canum var. alpinum, Pterocephalus perennis subsp. perennis, Scutellaria rupestris subsp. parnassica, Silene radicosa subsp. radicosa, Stipa endotricha, Teucrium montanum var. parnassicum, Trinia frigida, Trinia guicciardi, Trisetum tenuiforme, Verbascum epixanthinum var. epixanthinum, Veronica erinoides, V. thymifolia, Viola chelmea, V. greca.
Differential species: Achillea fraasii, Achillea holosericea, Acinos alpinus subsp. meridionalis, Aethionema saxatile subsp. graecum, Anthemis cretica subsp. cretica, Asyneuma limonifolium, Aubrieta deltoidea var. deltoidea, Aubrieta deltoidea subsp. intermedia, Bromopsis lacmonica, Bromus riparius, Campanula spathulata subsp. spathulata, Carduus tmoleus, Carlina frigida, Carum graecum subsp. graecum, Carum meoides, Dianthus integer subsp. minutiflorus, Dianthus viscidus var. viscidus, Draba lasiocarpa, Euphorbia herniariifolia, Festuca callieri subsp. callieri, Festuca jeanpertii subsp. jeanpertii, Galium incanum subsp. incanum, Geranium macrostylum, Geranium subcaulescens, Helictotrichon aetolicum, Koeleria mitrushii, Linaria peloponnesiaca, Linum elegans, Minuartia juniperina, Minuartia stellata, Morina persica, Myosotis suaveolens, Myosotis sylvatica subsp. canea, Onobrychis alba subsp. pentelica, Pimpinella tragium subsp. polyclada, Pimpinella tragium subsp. tragium, Ranunculus sartorianus, Sedum laconicum, Sempervivum marmoreum, Sesleria tenerrima, Sesleria vaginalis, Silene bupleuroides subsp. staticifolia, Stachys heldreichii, Telephium orientale, Thymus chaubardii, Thymus leucotrichus, Tragopogon crocifolius subsp. samaritanii.
Structure and ecology: The class groups pulvinate orophilous plant communities characterized by dominance of dwarf shrubs, often with tragacanthoid habit, sometimes mixed with caespitose hemicryptophytes, which constitute quite spaced grasslands, where numerous geophytes or rosulate hemicryptophytes play a relevant physiognomic role. The stands colonized by these communities are usually represented by more or less rocky windy ridges and cacuminal surfaces usually with undeveloped soils, as well as more o less stabilized screes. These habitats are distributed mainly in the mountains at 1500–3000 m of altitude, with stands characterized by quite rigid environmental conditions. Sometimes, especially in situations of insularity these plant communities occur also at lower altitudes, sometimes up to 1000 m. From the bioclimatic point of view, these communities are distributed prevalently within the supra- and oro-Mediterranean belts, as well as in supra- and oro-temperate belts, often of sub-Mediterranean type. Downwards, they tend to penetrate into meso-Mediterranean belt, especially due to the degradation processes of the woodlands or when the edaphic conditions are particularly critical, as in the case of blocking of the pedogenetic processes. Dynamically, it is a typically orophilous vegetation showing usually a climatophilous role, even if often it is represented by edaphophilous communities. When these communities are localized within the forest belt, they assume a secondary role, being linked usually to processes of woodland degradation. As concerns its floristic arrangement, this vegetation is characterized by a rich set of endemics, often having a relevant taxonomic and phytogeographic significance. Many of them are relict species belonging to Tertiary elements, often represented by groups taxonomically isolated, segregated in a lot of geographical vicariants. Apart from a contingent of endemic taxa, which are proposed as characteristics of this class, other non-strictly endemic species with a wider distribution are considered as “differential species”, since in Greece they are usually localized in this type of orophilous vegetation.
Distribution: According to literature and unpublished personal data, this class has its greater spread on the mountains of mainland Greece, extending northwards to Albania and Macedonia and eastwards in the north-western and western Anatolia, as well as in Euboea and some Ionian Islands. Moreover, altough floristically rather impoverished, it is represented also in some islands of north-eastern and northern Aegean, such as Samos, Chios, Lesbos, Samothraki and Thassos, where high mountains occur.
Syn.: Daphneeto-Festucetalia Quézel 1964, Vegetatio, 12:325 p.p., nom. amb. rejic. propos. (art. 36).
Daphno oleoidis-Festucetalia variae Quézel 1964, Vegetatio, 12:325, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18:82, p.p., nom. amb. rejic. propos. (art. 36).
Acantholimo-Astragaletalia Voliotis 1973, Sci. Ann. Fac. Phys. Math Univ. Thess. 13:237, p.p., nom. nud.
Holotypus: Sideritido raeseri-Thymion parnassici Musarella, Brullo & Giusso all. nov. hoc loco.
Characteristic species: Acantholimon graecum, Alkanna graeca subsp. boetica, Allium achaium, Allium frigidum, Alyssum repens var. brachyphyllum, Armeria orphanidis, Asperula rigidula, Astragalus angustifolius subsp. erinaceus, Astragalus rumelicus subsp. taygeticus, Avenochloa agropyroides, Centaurea affinis subsp. laconiae, Cirsium hypopsilium, Crepis incana, Dasypyrum hordeaceum, Dianthus androsaceus, Dianthus biflorus, Draba parnassica, Echinops taygeteus, Erodium chrysanthum, Eryngium multifidum, Erysimum asperulum, Erysimum pusillum, Euphorbia deflexa, Festuca janpertii subsp. achaica, Galium taygeteum, Geocaryum parnassicum, Geocaryum peloponnesiacum, Inula candida subsp. limonella, Noccaea graeca, Paronychia albanica subsp. graeca, Rindera graeca, Scutellaria rupestris subsp. rupestris, Verbascum acaule.
Structure and ecology: This order groups the orophilous plant communities, as highlighted in the class, linked mainly to the supra-and oro-temperate belts of sub-Mediterranean type, occurring mainly at above 1700–1800 m of altitude. These plant communities show a climatophilous, or sometimes edaphophilous character, usually are localized in the cacuminal stands of the mountains above the timberline. Within this syntaxon the plant communities distributed also at lower altitudes (1000–1700 m) falling in the meso-and oro-Mediterranean belt can be included. In this case, the vegetation is largely represented by secondary communities, often of edaphophilous type, since linked to degradation processes of the woodlands.
Distribution: On the basis of current knowledge, the order seems to be circumscribed to the mountains of Greece, Peloponnese included, as well as the Ionian Islands (Cephalonia and Lefkas) and Euboea.
Syn.: Eryngieto-Bromion Quézel 1964, Vegetatio, 12:326, p.min.p., nom. amb. rejic. propos. (art. 36).
Eryngio multifidi-Bromion fibrosi Quézel 1964, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18:82 p.min.p., nom. amb. rejic. propos. (art. 36).
Astragaleto-Seslerion Quézel 1964, Vegetatio, 12:326, p.min.p., nom. amb. rejic. propos. (art. 36).
Astragalo angustifolii-Seslerion coerulantis Quézel 1964, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18:82, p.min.p., nom. amb. rejic. propos. (art. 36).
Stipeto-Morinion Quézel 1964, Vegetatio, 12:326, p.min.p, nom. amb. rejic. propos. (art. 36).
Stipo pulcherrimae-Morinion persicae Quézel 1964, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18:82 p.min.p., nom. amb. rejic. propos. (art. 36).
Holotypus: Astragalo lactei-Convolvuletum cochlearis Quézel 1964, hoc loco.
Characteristic species: Alyssum montanum subsp. hymettium, Centaurea affinis subsp. affinis, Centaurea affinis subsp. pallidior Dianthus viscidus var. parnassicus, Erigeron glabratus subsp. graecus, Erysimum parnassi, Festuca graeca subsp. graeca, Galium circae, Geocaryum parnassicum, Lactuca intricata, Linaria parnassica, Marrubium velutinum, Nepeta parnassica, Nepeta spruneri, Satureja parnassica, Sideritis raeseri subsp. raeseri, Thymus leucospermus, Thymus parnassicus, Thymus teucrioides subsp. teucrioides, Verbascum parnassicum.
Structure and ecology: Within the order
Eryngio multifidi-Armerietalia orphanidis, this alliance is that one showing more marked characters of continentality. The associations belonging to this syntaxon seem to have greater floristic structural and ecological correlations with those ones occurring in the northern Greece. Clearly, towards to the north of Greece, the bioclimate becomes markedly more mesic with a progressive decrease of its Mediterranean character. This is reflected quite well in the orophilic pulvinate vegetation, which shows a more marked thermophily in the mountains of southern Greece. Therefore, this syntaxon can be considered as the transition term between the southernmost alliances occurring in the Peloponnese and probably the northernmost ones regarding the mountain ranges of Pindus and Mt. Olympus, which is still to be defined under the phytosociological profile including several associations already defined by Quézel [
36]. In particular, the associations falling in the
Marrubio velutini-Thymion parnassici, while maintaining structurally their prerogatives of shrub-pulvinate community, tend to show a certain increase of the hemicriptophytic component. Further, their floristic settlement increases with elements having more relantionships with taxonomic groups having a more northernmost distribution.
Distribution: The alliance is distributed mainly in the massifs of Sterea Ellas, such as Mt. Parnassus, Mt. Giona, Mt. Vardoussia and Mt. Timfristos, as well as of Attica. Probably, plant communities belonging to this syntaxon occur also in other mountains of this continental area of Greece.
Notes: The
Marrubio velutini-Thymion parnassici does not show any clear floristic, ecological and chorological correlation with the three alliances described by Quézel [
35]. In particular, this new syntaxon is floristically differentiated by endemics distributed in the high-mountain belt of the massifs located exclusively in Sterea Ellas and Attica. In addition, this alliance groups associations that are not linked to a well-defined altitudinal belt, but they are distributed from the lower mountain zones (1200–1300 m) up to the high-mountain ones reaching the altitude of 2500 m.
Marrubio velutini-Astragaletum rumelici Quézel 1964, Vegetatio 12:334 (
Appendix C,
Table A6).
Syn.: Association à Astragalus creticus subsp. rumelicus et Marrubium velutinum, Quézel 1964.
Lectotypus: Table 18, rel. 3, Quézel [
35],
hoc loco.Characteristic species: Astragalus rumelicus subsp. rumelicus, A. hellenicus, Nepeta parnassica.
Structure and ecology: The association is located on calcareous and dolomitic substrata, of more or less rocky steep slopes (30°–40°), characterized by eroded or not very deep soils, rich in coarse skeletal component. It assumes a clear climatophilous role in the supra-temperate sub-Mediterranean belt at an elevation of 1800 and 2100 m, while at lower altitudes (examples were found up to 1500 m) shows a clearly secondary pattern, because its spread is linked to the processes of forest degradation, here represented mainly by
Abies cephalonica woods. Physiognomically, this association is dominated by thorny cushion-like of
Astragalus rumelicus subsp.
rumelicus, which often constitues dense populations. Quite significant it is the occurrence, although scattered, in this vegetation of two interesting endemic species, such as
Nepeta parnassica, distributed in Mt. Parnassus and Mt. Chelmos (on the latter, however, is quite rare), and
Astragalus hellenicus, widespread on the mountains of Sterea Ellas. Within this association, as emphatized by Quézel [
35], two subassociations linked to different soil conditions can be distinguished. They are cited by that author as subass.
typicum, localized on carbonatic substrates with no floristic differentiation, and subass.
achilleetosum nobilis Quézel 1964 (lectotypus rel. 12, Table 18, Quézel [
35],
hoc loco) restricted to sandstone or sometimes schist outcrops, differentiated by
Achillea nobilis and
Salvia argentea var.
alpina.
Distribution: This association is well represented on the southernmost massifs of Sterea Ellas, as Mt. Parnassus, Mt. Giona and Mt. Vardoussia. However, its occurrence also in other mountain massifs of this area can not be excluded.
Astragalo lactei-Convolvuletum cochlearis Quézel 1964, Vegetatio 12:339 (
Appendix C,
Table A7).
Syn.: Association à Convolvulus cochlearis et Astragalus lacteus Quézel 1964.
Lectotypus: Table 21, rel. 4, Quézel [
35],
hoc loco.Characteristic species: Astragalus lacteus, Convolvulus cochlearis, Koeleria carniolica.
Structure and ecology: The association is confined to the dolomitic substrates of the ridges that bordered some deep dolines. The surfaces occupied by this association are usually almost flat and are distributed at an altitude of 1650–1800 m, within the supratemperate sub-Mediterranean bioclimatic belt. This vegetation is dominated by small prostrate chamaephytes, among them have a quite significant role Convolvulus cochlearis (=C. parnassicus Boiss. & Orph.), rather rare Balkan endemic. In this association it occurs also Astragalus lacteus, which shows a quite constant frequence, as well as Asperula rigidula and Koeleria carniolica, which are less frequent.
Distribution: Currently it is known only to the Mt. Parnassus, where it is observed near the refuge of the EOS Gherondovrachos.
Notes: As concerns this association, Quézel [
35] highlight that it occupies an intermediate position between the
Astragalo-Seslerion and
Stipo-Morinion alliances, because in its floristic settlement are present characteristic species of both syntaxa. However, the author considers more properly to include it in the
Astragalo-Seslerion, mainly for the occurrence of
Astragalus angustifolius. That is further evidence of the lack of phytosociological value of the alliances proposed by the author.
Nepeto epiroticae-Astragaletum corynthiaci (Quézel 1964) Musarella, Brullo & Giusso nom. nov. (
Appendix C,
Table A8).
Syn.: Association à Astragalus cephalonicus et Nepeta nuda Quézel 1964, Vegetatio 12:357.
Lectotypus: Table 30, rel. 2, Quézel [
35],
hoc loco.Characteristic species: Astragalus corynthiacus, Nepeta nuda var. epirotica.
Structure and ecology: The association is localized on the bottom of dolines and also on slightly inclined surfaces characterized by rather deep silt-clay soils, deposited on carbonate substrata. It is distributed between 1600 and 1900 m of altitude, sometimes reaching 2100 m, having its optimum in the supratemperate sub-Mediterranean belt. Physiognomically, this vegetation is differentiated by the dominance of Astragalus corynthiacus, a new species closely related to A. cephalonicus, which tends to constitute dense and homogeneous populations. Another quite significant species is Nepeta nuda var. epirotica, which seems to have its optimum in these stands. Potentially, this association is linked to the erosion processes and washing away of calcareous rocks that accumulate fine particles into the lower parts of dolines and depressions. These surfaces, in extreme conditions, with very deep soils, are ususally colonized by hemicryptophytic communities of Trifolion parnassi. In fact, in this association, some elements belonging to the latter alliance and related order, Trifolietalia parnassi, are present which clearly have the meaning of transgressiion. In conditions of marked edaphic xericity, such as in the stands with rocky outcrops and superficial soils, the vegetation at issue is replaced by the climatophylous communities of Marrubio velutini-Astragaletum rumelici.
Distribution: The association was currently observed only on Mt. Parnassus, where it is represented mainly in the dolines.
Notes: As regards its phytosociological arrangement, this association was described by Quézel [
35] as Association à
Astragalus cephalonicus et
Nepeta nuda and included into the alliance
Trifolion parnassi, since the author based on its ecological requirements, being linked to deep soils and on the presence of a fair number of species characteristic of this syntaxon. However, it should be noted that the author considered this association structurally very similar to the communities of
Daphno-Festucetalia, especially for the dominance of torny cushion-like shrubs, completely absent in the typical grasslands of
Trifolion parnassi. Moreover, for the presence of a significant settlement of
Daphno-Festucetalia, he considered this association as intermediate between this order and that of
Trifolietalia parnassi. In fact, this perplexity of Quézel [
35] is here shared by us too, but basing on its floristic and structural characterics, it seems to exclude its possible attribution to
Trifolion parnassi. It is to underline that on the whole in this association are well represented many species of
Marrubio velutini-Thymion parnassici and related higher syntaxa. The dominant species was previously identified by Quézel [
35] as
Astragalus cephalonicus, but this attribution was wrong, since it clearly differs from the latter in numerous morphological features and should be treated as a distinct new species named
A. corinthiacus.Nepeto spruneri-Astragaletum corynthiaci Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A9).
Characteristic species: Astragalus corynthiacus, Nepeta spruneri.
Structure and ecology: This association can be considered as a geographical vicariant of Nepeto epiroticae-Astragaletum corynthiaci previously described from Mt Parnassus. It is also caracterized by the dominance of Astragalus corynthiacus, while Nepeta nuda var. epirotica is replaced by N. spruneri. This vegetation shows the same ecology of the above-mentioned association, since it always occurs in the dolines characterized by quite deep soils, usually localized between 1700–1800 m of elevation, sometimes reaching 2000 m. Floristically, it is well differentiated by several species of the alliance and higher ranks, while that ones of Trifolion parnassi are very rare.
Distribution: The association was surveyed in some stands of Mt. Giona.
Thymo parnassici-Paronychietum polygonifoliae Quézel 1964, Vegetatio 12:341 corr. (
Appendix C,
Table A10).
Syn.: Association à Paronychia chionaea et Thymus hirsutus subsp. ciliato-pubescens Quézel 1964.
Lectotypus: Table 23, rel. 3, Quézel [
35],
hoc loco.Characteristic species: Paronychia polygonifolia (=P. chionaea), Edraianthus graminifolius f. minor, Dianthus ventricosus.
Structure and ecology: This association, characterized by dominance of small chamaephytes showing a prostrate or creeping habit, is localized in correspondence to the very windy ridges, usually over 2000 m of altitude. It is possible to observe this vegetation also at lower altitudes (ca. 1800 m), always in cacuminal stands. From the bioclimatic point of view, this association is well represented in the oro-temperate sub-Mediterranean belt extending downward in the supra-temperate sub-Mediterranean one. The surfaces are rather flat with superficial soils rich in minute skeleton, where, due to the action of the winds, the soil evolution is very slow, and the vegetation always keeps a prostrate habit. According to Quézel [
35], this vegetation is dominated by plants showing a small size, such as
Paronychia polygonifolia (as
P. chionaea),
Thymus parnassicus (as
T. hirsutus subsp.
ciliato-pubescens),
Edraianthus graminifolius f.
minor, Dianthus ventricosus. The pulvinated camaephytes and the cespitose grasses are totally absent. The author distinguished two subassociations linked to altitudinal factors, represented at over 2100 m of altitude by the subass.
typicum, which is replaced at lower altitudes from subass.
linetosum angustifolii Quézel 1964 (lectotypus: Table 23 rel. 5,
hoc loco). Floristically, the first subassociation is differentiated by
Euphrasia salisburgensis,
Minuartia condensata, Festuca halleri subsp.
riloensis, Carex kitaibeliana, and
Galium plebeium, while the second one has as differential species
Linum tenuifolium and
Ptilotrichum rupestre.Distribution: The association seems to be exclusive of Mt. Giona, where it is very frequent.
Nepeto sprunerii-Astragaletum tymphrestei Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A11).
Characteristic species: Astragalus tymphresteus.
Structure and ecology: The association was observed in stands at altitudes between 1200 and 1400 m, on slightly inclinated slopes characterized by carbonate rocks within the meso-Mediterranean bioclimatic belt. The soils are poorly developed with many minute skeletons. This vegetation is dominated by Astragalus thymphresteus, thorny dwarf shrub growing with other small chamaephytes, such as Nepeta spruneri, Thymus chaubardii, Chamaecytisus hirsutus, and some cespitose hemicryptophytes.
Distribution: The association was found only on Mt. Giona, where it is circumscribed to stands of lower altitudes, but it probably occurs also in other mountains.
Violo stojanowii-Seslerietum vaginalis Quézel 1973, Biol. Gallo-Hellen. 5(1):152, corr. (
Appendix C,
Table A12).
Syn.: Association à Sesleria coerulans et Viola stojanowii Quézel 1973.
Lectotypus: Table 3, rel. 11, Quézel [
38],
hoc loco.Characteristic species: Viola stojanowii, Thymus teucrioides subsp. teucrioides, Thymus striatus.
Structure and ecology: The association occurs over 2200 m of altitude, where is localized in the small depressions among the cacuminal rocky peaks, where very minute clasts are accumulated and covered by soils rich in clay subject to solifluction. In these stands characterized by an acclivity of 20–30%, the vegetation shows a rather sparse coverage in which Sesleria vaginalis (=S. coerulans) plays an important role. On the whole, it is a floristically quite poor herbaceous vegetation, where Viola stojanowii is physiognomically significant. Usually, this association takes catenal contacts with the scree vegetation belonging to Drypetalia spinosae.
Distribution: This vegetation was described by Quézel [
38] for Mt. Vardoussia, but probably it occurs also in other mountains of Sterea Ellas.
Syn.: Association à Minuartia stellata et Erysimum pusillum subsp. parnassi Quézel 1964.
Lectotypus: Table 22, rel. 3, Quézel [
35],
hoc loco.Characteristic species: Minuartia stellata, Astragalus apollineus, Anthemis spruneri, Allium parnassicum, Anthemis tinctoria var. parnassica, Erigeron alpinus.
Structure and ecology: The association colonizes the rocky outcrops and the stabilized screes at altitudes over 2100 m, within the oro-temperate sub-Mediterranean bioclimatic belt. It is frequent on the prevalently rocky surfaces that, due to the considerable acclivity, the soils are very superficial, accumulating mainly among the rocky crevices and into the bushes. Physiognomically, it is distinguished by the dominance of compact and often voluminous cushion-like shrubs of
Minuartia stellata, that usually grows togheter with
Sesleria vaginalis and several species with prostrate habit. The characteristic species of the alliance
Marrubio velutini-Thymion parnassici are well represented, among them
Erysimum parnassi,
Marrubium velutinum,
Satureja parnassica, which show high coverage value. Within this association Quézel [
35] distinguished two subassociations on phytogeographical base, represented by
saturejetosum parnassicae (=subass.
teucrioides à
Thymus), restricted to Mt. Parnassus, and by
aurinietosum giónae (=subass.
kionae à
Alyssum) for Mt. Giona. The first one corresponds clearly to the type, while the second one must be treated as a distinct association, well differentiated from floristically, also from chorological point of view, named as
Aurinio gionae-Minuartietum stellatae.
Distribution: Actually, this vegetation is distributed only on Mt. Parnassus.
Aurinio gionae-Minuartietum stellatae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A14).
Syn.: Association à Minuartia stellata et Erysimum pusillum subsp. parnassi subass. à Alyssum kionae Quézel 1964.
Holotypus: Table 22, rel. 3, Quézel [
35],
hoc loco.Characteristic species: Minuartia stellata, Aurinia gionae.
Structure and ecology: From the ecological point of view, the association is very similar to Erysimo parnassi-Minuartietum stellatae. In fact, it occurs at altitudes between 2100 and 2450 m, on calcareous substrata, more or less acclive, showing a coverage which not exceeding 70%. Floristically the vegetation differs markedly from the Erysimo parnassi-Minuartietum stellatae, for the almost total absence of Erysimum parnassi, Satureja parnassica, Sesleria vaginalis, all species that in the latter association are fairly common and often dominant. In addition to the absence of all characteristic species, the association at issue differs from the previous one also for the occurrence of the endemic Aurinia gionae. The only common element between the two communities is the dominance of Minuartia stellata.
Distribution: The association is esclusive of some places of Mt. Giona, where it is quite frequent.
Achilleo fraisii-Dianthetum tymphrestei Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A15).
Characteristic species: Dianthus tymphresteus, Valeriana bertiscea.
Structure and ecology: The association is localized on small rocky summits, in the more or less flat places characterized by minute crumbly limestone mixed with a little soil. It has been observed at altitudes of 1700–1800 m of very windy stands, within the supra-temperate sub-Mediterranean belt. Floristically, it is differentiated by the dominance of small pulvinate shrubs of Dianthus tymphresteus, which grows together other cespitose hemicryptophytes and small prostrate chamaephytes, such as Centaurea affinis subsp. affinis, Achillea fraisii, Koeleria mitrushi, Festuca jeanpertii subsp. jeanpertii, Astragalus lacteus, etc.
Distribution: This association was surveyed on Mt. Giona at Liritsa, but it probably occurs also in other neighbouring massifs, such as Vardoussia and Timphristos.
Asperulo luteae-Achilleetum umbellatae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A16).
Characteristic species: Achillea umbellata, Carex caryophyllea.
Structure and ecology: The association colonizes the slopes often rather inclinated with fresh solis, mixed to big size clasts, at altitudes of 1700–1800 m, within the suprat-emperate sub-Mediterranean belt. The surfaces occupied by this vegetation are usually South-facing and are frequent at the base of small rocky ridges. In such habitats, several hemicryptophytes such as Achillea umbellata, Carex caryophyllea, Asperula lutea, Festuca cyllenica subsp. cyllenica, Stipa endotricha, Koeleria mitrushi and Festuca jeanpertii subsp. jeanpertii occur and thrive.
Distribution: This vegetation was surveyed only on Mt. Giona, near Liritsa, where it is very circumscribed.
Astragalo lactei-Asperuletum apiculatae Musarella, Brullo & Giusso ass. nov.
hoc hoco (
Appendix C,
Table A17).
Characteristic species: Asperula purpurea subsp. apiculata, Astragalus lacteus.
Structure and ecology: The association seems exclusive of the calcareous rocky ridges at altitudes between 1500 and 1600 m, where it is linked to slopes with very variable inclination (30–80°), with S-SO exposure. From the biolimatic point of view, it falls between the meso-Mediterranean and supra temperate sub-Mediterranean belts. The vegetation is localized along the large cracks of the rock and is characterized by small chamaephytes and hemicryptophytes. Among them, Asperula purpurea subsp. apiculata, Astragalus lacteus, Achillea holosericea and Thymus chaubardii are dominant togheter with various grasses.
Distribution: The association was observed on Mt. Giona at Mavrikorfi, near Proni, where seems quite localized.
Diantho minutiflori-Festucetum cyllenicae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A18).
Characteristic species: Dianthus integer subsp. minutiflorus, Festuca cyllenica subsp. cyllenica, Silene roemeri subsp. macrocarpa.
Structure and ecology: The association colonizes the more or less stabilized screes with an inclination of 20–30°, at an altitude of around 2000 m. It is found in the orotemperate sub-Mediterranean belt, penetrating downward in the sub-Mediterranean supra.temperate one. Physiognomically, it is differentiated by the dominance of large tuffs of Festuca cyllenica subsp. cyllenica, often associated with Sesleria vaginalis. In particular, this community is characterized by Dianthus integer subsp. minutiflorus and Silene roemeri subsp. macrocarpa. Moreover, Satureja parnassica, Nepeta spruneri, Galium thymifolium, Campanula spathulata subsp. spathulata, and Ranunculus brevifolius are very frequent.
Distribution: The association was surveyed on Mt. Giona at Amfissa, near Pirghakia.
Scabioso ochroleucae-Sideridetum raeseri Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A19).
Characteristic species: Scabiosa ochroleuca, Sideritis raeseri subsp. raeseri, Vincetoxicum hirundinaria subsp. nivale.
Structure and ecology: This association replaces the Diantho minutiflori-Festucetum cyllenicae in the stabilzed screes or, anyway, on the surfaces more compact and richer in soil. Physiognomically, it is differentiated by the dominance of suffruticous shrubs, mainly chamaephytes, such as Scabiosa ochroleuca, Sideritis raeseri subsp. raeseri, Vincetoxicum hirundinaria subsp. nivale, Satureja parnassica, Marrubium velutinum, Asperula lutea, Centaurea affinis subsp. affinis, Nepeta spruneri, etc., while decrease the coverage of the caespitose hemicryptophytes.
Distribution: The association was surveyed on Mt. Giona, in the same place where the Diantho minutiflori-Festucetum cyllenicae occurs.
Ranunculo psilostachydis-Festucetum cyllenicae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A20).
Characteristic species: Festuca cyllenica subsp. cyllenica, Laserpitium pseudomeum, Ranunculus psilostachys.
Structure and ecology: This association replaces the Diantho minutiflori-Festucetum cyllenicae on the more or less stabilized screes localized at lower altitudes (1700–1750 m) in quite fresh and sheltered stands. Particularly, it is frequent in the supra-temperate sub-Mediterranean belt, on surfaces having an inclination of 25–35°. Physiognomically, this vegetation is dominated by Festuca cyllenica subsp. cyllenica, but in comparison with the prevoius association, in its floristic settlement, a marked decrease of the more orophilous species is observable. Nevertheless, it is well differentiated due to the occurrence of Ranunculus psilostachys, Laserpitium pseudomeum, Galium circae, Avenochloa agropyroides, Trisetum tenuiforme, etc., species linked to stands of lower altitudes.
Distribution: As the two previous associations, this vegetation was surveyed in the same area of Mt. Giona, but at lower altitudes.
Edraiantho parnassici-Globularietum cordifoliae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A21).
Characteristic species: Globularia cordifolia, Anthyllis montana subsp. jacquinii, Edraianthus parnassicus, Silene auricolata.
Structure and ecology: The association is localized in rocky places, generally more or less flat or, however, a little sloped. It shows a wide altimetric range, ranging at altitudes from 1700 to 2150 m, thus affecting the supratemperate and orotemperate sub-Mediterranean bioclimatic belts. It is can be considered as a semi-rupestrian community characterized by prostrate or creeping chamaephytes, such as Globularia cordifolia, Anthyllis montana subsp. jacquinii, Edraianthus parnassicus, Silene auricolata, which grow togheter with other small pulvinate shrubs, among them Paronychia polygonifolia, Satureja parnassica, Thymus leucotrichus, etc.
Distribution: The association is spread on some mountain places of Mt. Giona where, usually, it is localized on small surfaces.
Thymo parnassici-Astragaletum parnassi Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A22).
Characteristic species: Astragalus parnassi.
Structure and ecology: This association is characterized by the dominance of thorny cushion-like shrubs of Astragalus parnassi. This species is linked to very lower altitudes (1000–1300 m) within the meso-Mediterranean bioclimatic belt, characterizing one of the most termophilous communities of the Marrubio velutini-Thymion parnassici. However, the characteristic species of this alliance and the higher syntaxa, are here well represented, among them Thymus parnassicus, Erysimum parnassi, Festuca graeca subsp. graeca, Astragalus angustifolius subsp. erinaceus, Asperula lutea, etc. The vegetation is usually localized in quite fresh and sheltered stands, represented mainly by clearing within the Abies cephalonica woodlands. It colonizes the more or less flat surfaces, showing high coverage values.
Distribution: The association is spread in the southern slopes of Mt. Parnassus.
Chamaecytiso hirsuti-Astragaletum parnassi Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A23).
Characteristic species: Astragalus parnassi, Chamaecytisus hirsutus.
Structure and ecology: This association must be considered as a geographic vicariant of the Thymo parnassici-Astragaletum parnassi. In fact, it occurs on Mt. Giona, where it is localized in habitats very similar to that one occupied by the aforesaid association. This vegetation is always characterized by the dominance of Astragalus parnassi and is ditributed at an altitude of 1250–1500 m, in little inclinate stands localized within the Abies cephalonica woodlands. Floristically, it is characterized by the occurrence of Chamaecytisus hirsutus that forms large creeping cushion-like shrubs, while totally absent are several species of the related alliance, frequent though in the previous association.
Distribution: The association occurs in various localities of Mt. Giona.
Onobrychido pentelicae-Genistetum parnassicae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A24).
Characteristic species: Genista parnassica, Onobrychis alba subsp. pentelica.
Structure and ecology: The association replaces the Thymo parnassici-Astragaletum parnassici on the slopes with northern exposure of the southern part of Mt. Parnassus. This vegetation is very circumscribed and linked to a little inclinated escarpments with very deep and fresh soils at an altitude of 1100–1200 m. It is localized wihin the meso-temperate bioclimatic belt and is characterized by the dominance of the rare Genista parnassica that usually forms large thorny cushion-like shrubs, aften growing with Astragalus angustifolius subsp. erinaceus and, occasionally, with Astragalus rumelicus subsp. rumelicus and A. parnassi. Differential species of this association is Onobrychis alba subsp. pentelica, while as concerns the species of the higher syntaxa are well represented.
Distribution: The association seems circumscribed to a very narrow area of the southern part of Mt. Parnassus.
Allio cithaeronis-Dianthetum serratifolii Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A25).
Characteristic species: Allium cithaeronis, Dianthus serratifolius subsp. serratifolius, Petrorhagia armerioides, Paronychia macedonica, Scabiosa ochroleuca.
Structure and ecology: The association is circumscribed at the cacuminal calcareous plateau of Mt. Kitheronas, at an altitude of 1350–1400 m. It is a very windy place, subjected, unfortunately, to overgrazing, falling in the meso-Mediterranean bioclimatic belt. Physiognomically, it is differentiated by the occurrence of some small chamaephytes as Dianthus serratifolius subsp. serratifolius, Petrorhagia armerioides, Paronychia macedonica, and Scabiosa ochroleuca, growing togheter with the endemic Allium cithaeronis.
Distribution: The association is exclusive of Mt. Kitheronas (Sterea Hellas).
Inulo methaneae-Sideritetum atticae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A26).
Characteristic species: Inula verbascifolia subsp. methanea, Sideritis raeseri subsp. attica, Aethionema saxatile subsp. graecum.
Structure and ecology: The association occurs on the calcareous slopes of Mt. Parnis at an altitude of 1150–1300 m, within an area characterized by a meso-Mediterranean bioclimate and, particularly, affected by a regime of dense fog. It is localized on flat or a little inclinated surfaces with variable exposure. Physiognomically, it is differentiated by the occurrence and often dominace of small shrubs, as Inula verbascifolia subsp. methanea, Sideritis raeseri subsp. attica, Aethionema saxatile subsp. graecum, Alyssum montanum subsp. hymettium, Achillea holosericea, etc.
Distribution: The association was surveyed only in cacuminal stands of Mt. Parnis near Athens.
Holotypus: Scutellario cephalonicae-Astragaletum cephalonici Musarella, Brullo & Giusso ass. nov. hoc loco.
Characteristic species: Astragalus cephalonicus, Centaurea subciliaris subsp. subciliaris, Thymus holosericeus, Petrorhagia fasciculata var. cephallenica, Scutellaria rupestris subsp. cephalonica.
Structure and ecology: The alliance replaces in the Ionian islands of Cephalonia and Lefkada the Marrubio velutini-Thymion parnassici distributed in Sterea Ellas and Attica. The syntaxon at issue is well differentiated from the previous alliance for some floristic and ecological peculiarity due to its geographical isolation. Floristically, it is also differentiated by some insular endemics exclusive of Cephalonia and Lefkada, taxonomically quite significant, such as Astragalus cephalonicus, Centaurea subciliaris subsp. subciliaris, Thymus holosericeus, Scutellaria rupestris subsp. cephalonica, and Petrorhagia fasciculata var. cephallenica. The communities belonging to this alliance are localized on the top of isolated mountain summits at altitudes between 800 and 1400 m, which are markedly affected by moist marine winds.
Distribution: The alliance seems circumscribed to the Ionian Islands of Cephalonia and Lefkada.
Helictotricho convoluti-Thymetum holosericei Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A27, rel. 1–5).
Characteristic species: Helictotrichon convolutum subsp. convolutum, Ononis pusilla, Allium lefkadensis, Aurinia saxatilis subsp. saxatilis, Erysimum linearifolium.
Structure and ecology: The association is localized on the cacuminal plateau more or less windy, characterized by very rocky calcareous substrata with immature soils. This vegetation has its optimum at 800–1000 m of altitude, within the upper meso-Mediterranean belt. Floristically, it is differentiated by the dominance of the endemic
Thymus holosericeus which grows togheter with the tuffs of
Helictotrichon convolutum subsp.
convolutum, an Est-Mediterranean species, and the endemic
Allium lefkadensis. In this association occurs also
Astragalus cephalonicus which was already recorded in this mountain by Hofmann [
85].
Distribution: This association is localized in the Lefkas Island in small places on Mt. Elati (Stravoti).
Saturejo cuneifoliae-Thymetum holosericei Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A27, rel. 6–9).
Characteristic species: Allium cephalonicum, Centaurea spruneri subsp. guicciardi, Satureja cuneifolia.
Structure and ecology: The association, ecologically very similar to the previous one, occurs on calcareous rocky outcrops at 800–1000 m of altitude in the Cephalonia Island. Floristically, it is differentiated from the previous one for the lack of
Helictotrichon convolutum subsp.
convolutum, while
Satureja cuneifolia is frequent, which togheter with
Thymus holosericeus and
Astragalus cephalonicus, characterizes this cushion-like prostrate vegetation. Moreiover, the occurrence of
Allium cephalonicum in this vegetation is significant, as a very rare and isolated endemic species, closely related to
A. callidictyon C. A. Meyer ex Kunth [
86].
Distribution: It is a geographical vicariant of the previous association in Cephalonia Island where it is localized on Mt. Ainos and Mt. Roudhi in open and windy places.
Scutellario cephalonicae-Astragaletum cephalonici Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A27, rel. 10–12).
Characteristic species: Astragalus cephalonicus, Galium ionicum, Erysimum cephalonicum.
Structure and ecology: This association replaces the previous one in the higher stands at altitudes between 1200 and 1400 m, where it is localized in more or less sloping stands characterized by calcareous rocky substrata. Floristically, it is differentiated from the previous association for the dominace of Astragalus cephalonicus which grows togheter with other endemisms as Erysimum cephalonicum and Scutellaria rupestris subsp. cephalonica. This vegetation is localized within supra-Mediterranean bioclimatic belt in the clearing of the Abies cephalonica woodlands that occur in the surfaces with more deep and mature soils.
Distribution: The association occuring in the Cephalonia Island, replaces at higher altitudes the Saturejo cuneifoliae-Thymetum holosericei.
Paronychio graecae-Astragaletum erinacei Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A27, rel. 13–19).
Characteristic species: Astragalus angustifolius subsp. erinaceus, Paronychia albanica subsp. graeca, Galium circae, Trinia glauca subsp. pindica, Aubrieta deltoidea, Viola cephalonica, Astragalus depressus subsp. depressus, Verbascum guicciardii.
Structure and ecology: This association is localized in cacuminal open stands at an altitude of 1600 m, colonizing the calcareous rocks of southern slopes usually quite inclined. These surfaces are strongly affected by winds and daily thermic changes, also subject to long periods of snow cover, with very superficial and eroded soils. Physiognomically it is characterized by small and flattened pulvines of Astragalus angustifolius subsp. erinaceus, growing together with other dwarf orophytes with chamaephytic or hemicryptophytic habit, some of them endemic, such as Paronychia albanica subsp. graeca, Galium circae, Viola cephalonica, Scutellaria rupestris subsp. cephalonica, etc. This vegetation occurs within supra-Mediterranean bioclimatic belt, which is replaced in the northern slopes with not eroded and mature soils by Abies cephalonica woodlands.
Distribution: The association is exclusive of Cephalonia Island it only occurs in the top of Mount Ainos.
Holotypus: Sideritido euboeae-Astragaletum euboici Musarella, Brullo & Giusso ass. nov., hoc loco.
Characteristic species: Astragalus rumelicus subsp. euboicus, Asperula suffruticosa, Hieracium pannosum subsp. euboeum, Nepeta dirphya, Paronychia euboaea, Sideritis euboea, Verbascum delphicum.
Structure and ecology: The alliance can be considered a geographical vicariant on the Euboea mountains of the Marrubio velutini-Thymion parnassici distributed in the continental Central Greece. It is differentiated from the latter alliance for its floristic peculiarities (represented by several endemics), linked to geographical isolation due to its insularity. The plant communities belonging to this syntaxon are surveyed at altitudes between 1000 and 1700 m on prevalently carbonatic substrata.
Distribution: The alliance is circumscribed to the Euboea Island in the Central Egean Sea.
Sideritido euboeae-Astragaletum euboici Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A28).
Characteristic species: Astragalus rumelicus subsp. euboicus, Cytisus supinus.
Structure and ecology: The association is localized on the carbonatic rocky outcrops at 1100–1200 m of altitude, occasionally reaching 1350 m. The surfaces colonized by this vegetation are more or less inclinate and represented by sunny slopes. The vegetation is dominated by pulvinate shrubs of Astragalus rumelicus subsp. euboicus, which covers also very large surfaces. Other shrubs are also very frequent, such as Cytisus supinus, Sideritis euboea, Inula candida subsp. limonella and Nepeta dirphya, species quite important from the physiognomical point of view.
Distribution. The association was surveyed on Mt. Dirfis in the Euboea Island.
Scabioso webbianae-Phlomidetum samiae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A29).
Characteristic species: Phlomis samia, Scabiosa webbiana, Viola euboaea, Helleborus cyclophyllus.
Structure and ecology: The association is circumscribed to the fresh depressions with more deep soils and rich in humus, localized at 1000–1100 m of altitude. Quite significant it is here the occurrence of some mesophilous species with herbaceous habit, such as: Phlomis samia, Scabiosa webbiana, Viola euboaea and Helleborus cyclophyllus.
Distribution: The association was surveyed on Mt. Dirfis in the Euboea Island.
Sideritido euboeae-Festucetum cyllenicae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A30).
Characteristic species: Festuca cyllenica subsp. cyllenica, Sideritis euboea, Bolanthus graecus, Carum graecum subsp. graecum, Arenaria filicaulis subsp. euboica.
Structure and ecology: The cacuminal stands at altitudes over 1550 m are colonized by a herbaceous perennial vegetation dominated by Festuca cyllenica subsp. cyllenica. Usually, this species colonizes the stony soils and the consolidated screes, adapting well to long periods of snow cover. The association is well differentiated from the other communities characterized by the dominace of Festuca cyllenica subsp. cyllenica, distributed in the mountains of continental Greece, due to the occurrence of rare orophytes, some endemic of Euboea, such as Sideritis euboea.
Distribution: The association was surveyed on Mt. Dirfis in the Euboea Island.
Inulo limonellae-Seslerietum vaginalis Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A31).
Characteristic species: Sesleria vaginalis, Inula candida subsp. limonella.
Structure and ecology: The association covers the very inclinate southern rocky slopes of the calcareous summits at 1150–1500 m of altitude. Physiognomically, it is characterized by the dominace, with high coverage values, of Sesleria vaginalis, which grows with small shrubs of Inula candida subsp. limonella, Sideritis euboea and Astragalus rumelicus subsp. euboicus. The association replaces the Sideritido euboeae-Astragaletum euboici in the stands at altitudes over 1150 m of very opened and windy slopes.
Distribution: The association was surveyed on Mt. Dirfis in the Euboea Island.
Syn.: Eryngieto-Bromion Quézel 1964, Vegetatio, 12:326, p.min.p., nom. ambig. rejic. propos. (art. 36).
Eryngio multifidi-Bromion fibrosi Quézel 1964, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18:82 p.min.p.nom. ambig. rejic. propos. (art. 36).
Astragaleto-Seslerion Quézel 1964, Vegetatio, 12:326, p.min.p., nom. ambig. rejic. propos. (art. 36).
Astragalo angustifolii-Seslerion coerulantis Quézel 1964, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18:82, p.min.p., nom. ambig. rejic. propos. (art. 36).
Stipeto-Morinion Quézel 1964, Vegetatio, 12:326, p.min.p., nom ambig. rejic. propos. (art. 36).
Stipo pulcherrimae-Morinion persicae Quézel 1964, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18:82 p.min.p., nom. ambig. rejic. propos. (art. 36).
Holotypus: Festuco achaicae-Minuartietum stellatae Musarella, Brullo & Giusso ass. nov. hoc loco.
Characteristic species: Aster cylleneus, Astragalus calavrytensis, A. cylleneus, Festuca jeanpertii subsp. achaica, Globularia stygia, Marrubium cylleneum, Onobrychis montana subsp. macrocarpa, Sideritis clandestina subsp. peloponnesiaca, Taraxacum cylleneum, Verbascum cylleneum.
Structure and ecology: This alliance represents the southern geographical vicarious of Marrubio velutini-Thymion parnassici, grouping, similarly to the latter, orophilous plant communities structurally characterized by the dominance of chamaephytes and pulvinate nanophanerophytes, sometimes mixed with caespitose hemicryptophytes. Particularly, they differ from those ones occurring in the mountains of Sterea Ellas, apart from the occurrence of a rich set of endemics, also for their ecological requirements. In fact, these communities are subject to climatic conditions characterized by a a more marked thermophily, with higher average annual temperatures and drier rainfall regime, especially in summer. This area falls mainly in the supra-and oro-temperate of sub-Mediterranean type. Moreover, from the phytogeographical point of view, it is possible observe a strong increase of species belonging to taxonomic groups showing a more southern origin.
Distribution: The alliance is distributed in the mountains of northern Peloponnese (Mt. Erimanthos, Mt. Panachaiko, Mt. Chelmos, Mt. Klokos, Mt. Killini and Mt. Menalon).
Notes: The
Festuco achaicae-Marrubion cyllenei has a strictly phytogeographical characterization, since it is floristically differentiated by species confined to the mountains of Achaia, Corinthia and North Arcadia. It groups plant communities occurring in high mountain stands at altitudes from 1200 to 2400 m. This alliance groups, in addition to several new associations, also other ones described by Quézel [
35], Quézel and Katrabassa [
40], Georgiadis and Dimopoulos [
42], Maroulis and Georgiadis [
44], which previously were attributed by these authors in the alliances
Stipo-Morinion,
Eryngio-Bromion and
Astragalus-Seslerion.
Cirsio hypopsilii-Astragaletum taygetici Quézel 1964 corr. (
Table A32)
Syn.: Association à Astragalus cylleneus et Cirsium cylleneum Quézel 1964, Vegetatio 12:332.
Astracantho thracicae-Marrubietum cyllenei Georgiadis & Dimopoulos 1993 Bot. Helv. 103:153, nom. inval. (art. 3 c, 5)
Marrubio cyllenei-Astragaletum rumelici Maroulis & Georgiadis 2005, Fitosociologia 42(1): 43, nom. illeg. (art. 22,23); Holotypus: Table 2, rel. 460, Maroulis & Georgiadis [
44].
Lectotypus: Table 17, rel. 1, Quézel [
35],
hoc loco.Characteristic species: Astragalus rumelicus subsp. taygeticus, Cirsium hypopsilium.
Structure and ecology: The association is localized on the slopes more or less inclined with variable exposure, characterized by carbonatic stony substrata with rocky outcrops. The soils are enough evolved, but with a significant component of coarse skeleton. It is widespread at altitudes from 1400 to 2000 m, within the supra-temperate sub-Mediterranean bioclimatic belt, with penetrations upwoard in the oro-temperate submediterranean belt and downward in the meso.Mediterranean one. In fact, examples of this vegetation can be observed up to 2150 m of altitude in places well exposed and sunny, as well as at relative low altitudes (1150 m), limited to cacuminal and very rocky windy stands. Physiognomically, this association is differentiated by large thorny pulvinate individuals, often quite raised from the ground, of
Astragalus rumelicus subsp.
taygeticus, that in the Peloponnese replaces the subsp.
rumelicus, distributed in the central and northern Greece [
87]. Previously, the populations of this
Astragalus occurring in the M. Killini were identified by Quézel [
35] and Georgiadis and Dimopoulos [
42] as
Astragalus cylleneus, quite rare species on this massif, where it is localized in habitat totally different from those ones normally occupied by the association in question. As regards the floristic composition of this pulvinate vegetation, it is observed a rich contingent of characteristic species of the alliance, as well as of higher syntaxa. It assumes usually a climatophilous role especially at altitudes over 1700–1800 m, while at lower altitudes can be considered as an edaphophilous vegetation, limitedly to cacuminal more rocky stands. Within the climatophilous belt relative to
Abies cephalonica woodlands, the association represents usually a substitution aspect, due to degradation of this forest.
Distribution: The association is widespread and well represented in the various mountains of the northern Peloponnese, as Mt. Erimanthos, Mt. Panachaiko, Mt. Klokos and Mt. Killini, where it tends to occupy large surfaces.
Notes: This association was originally described by Quézel [
35] for Mt. Killini as ass. à
Astragalus cylleneus et
Cirsium cylleneum and successively redescribed by Georgiadis and Dimopoulos [
42], but changing its name in
Astracantho thracicae-Marrubietum cyllenei comb. nova, not indicating the holotypus. Therefore, the last syntaxon is an invalid name, according to articles 3 c and 5. In both cases, the authors indicate as characteristic species, physiognomically dominant,
Astragalus cylleneus (=
Astracantha thracica subsp.
cyllenea). Unfortunately, this species was misidentified by these authors, since on the Mt. Killini in the stands where they have carried out the relevés there is exclusively
Astragalus rumelicus subsp.
taygeticus, while the true
A. cylleneus is very rare and confined in depressed areas, such as dolines, characterized by very deep soils rich in silt-clay component, not occurring never on rocky substrata. The association occurs with the same ecological characteristics and floristic composition also on Mt. Erimanthos, where it was correctly described by Maroulis and Georgiadis [
44] as
Marrubio cyllenei-Astragaletum rumelici. However, this syntaxon is an illegitimate name being a synonym of the association described by Quézel [
35], whose name must be corrected in
Cirsio hypopsilii-Astragaletum taygetici. In this association are well represented the characteristic species of the three alliances proposed by Quézel [
35], particularity already evidenced by Georgiadis and Dimopoulos [
42] and also by Maroulis and Georgiadis [
44].
Syn.: Association à Aster cylleneus et Globularia stygia. Quézel 1964, Vegetatio, 12:337.
Lectotypus: Table 20, rel. 5, Quézel [
35],
hoc loco.Characteristic species: Aster cylleneus, Globularia stygia, Macrotoma cephalotes, Taraxacum bythinicum.
Structure and ecology: The association has its best expression between 2000 and 2330 m of altitude, within the orotemperate sub-Mediterranean bioclimatic belt. It can be observed sometimes up to 1800 m in stands representated by rocky ridge. Usually, it is localized on markedly rocky surfaces, constitute by carbonate substrata, as the ridges, saddles and stabilized screes, stends generally very windy with very shallow and undeveloped soils. It is a vegetation dominated by small prostrate dwarf shrubs mixed with several hemicryptophytes. The most important species are Globularia stygia and Aster cylleneus, rare endemics known for Mt. Chelmos and Mt. Killini. Floristically, the association is rather poor with low values of coverage. Dynamically, it can be considered an essentially edaphophilous vegetation.
Distribution: The association is currently known only for Mt. Chelmos and Mt. Killini in the northern Peloponnese.
Notes: This association was previously included by Quézel [
35] within
Astragalo-Seslerion, even though, as evidenced by the same author, the species of this alliance were not well represented in the relevés.
Euphrasio salisburgensis-Asperuletum oetaeae Quézel & Katrabassa 1974, Rev. Biol. Ecol. Medit. 1(1):19, corr. (
Appendix C,
Table A34).
Syn.: Association à Asperula nitida et Euphrasia salisburgensis Quézel & Katrabassa 1974.
Lectotypus: Table 4, rel. 3, Quézel and Katrabassa [
40],
hoc loco.Characteristic species: Asperula oetaea, Euphrasia salisburgensis, Iberis saxatilis subsp. saxatilis.
Structure and ecology: The association is localized at 2000 and 2200 m of altitude in the windy crests, with flat surfaces formed by eroded limestone cracked and free of soil. It is linked to the oro-temperate sub-Mediterranean bioclimatic belt, where it assumes a role clearly edaphophilous. Physiognomically, it is dominated by small prostrate chamaephytes mixed to rosulate hemicryptophytes with coverage values not too high. It is significant the occurrence of some orophytes that find in this vegetation type their optimal growth conditions, such as
Asperula oetaea (by Quèzel and Katrabassa [
40] quoted as
A. nitida),
Paronychia albanica subsp.
graeca (as
P. chionaea),
Euphrasia salisburgensis and
Iberis saxatilis subsp.
saxatilis. These authors distinguish within this association two sub-associations proposed as
erodietosum chrysanthi, located on compact limestone, and
minuartietosum confusae, occurring on calcareous substrata that flake on plakes.
Distribution: The association occurs only on Mt. Chelmos in northern Peloponnese.
Marrubio cyllenei-Astragaletum calavrytensis Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A35).
Syn.: Association à Astragalus cylleneus et Cirsium cylleneum subass. à Astragalus cylleneus Quézel & Katrabassa 1974, Rev. Biol. Ecol. Medit. 1(1):16, non Quézel 1964.
Characteristic species: Astragalus calavrytensis.
Structure and ecology: The association is localized on the little inclined slopes with rocky outcrops and more or less developed soils, rich in coarse skeletal component. It is developped in the bioclimatic belts between the supra-temperate sub-Mediterranean and the oro-temperate sub-Mediterranean, at altitudes of 1800 and 2200 m. Sometimes examples of this vegetation are found up to 1500 m of altitude in the markedly rocky stands. It is a plant community dominated by thorny pulvini of
Astragalus calavrytensis, by Quézel and Katrabassa [
40] mistakenly attributed to
A. cylleneus. This species showing often high values of coverage, it is usually associated with
Marrubium cylleneum, which assumes also a significant physiognomical role. The association has usually a clear climatophilous role, although sometimes, especially at lower altitudes, it represents an aspect of substitution, or at most edaphophilous. It can be considered as a geographical vicariant of the
Cirsio hypopsilii-Astragaletum taygetici occurring in other mountains of the Peloponnese. Whitin this association, three subassociations linked to altitudinal ranges can be distinguished: (a)
elytrigietosum intermediae subass. nov. (holotypus: rel. 6,
hoc loco), distributed at lower altitudes (1250–1650 m), differentiated by
Elytrigia intermedia and
Silene italica subsp.
peloponnesiaca; (b)
hippocrepidetum comosae subass. nov. (holotypus: rel. 10,
hoc loco), distributed between 1650 and 2000 m of altitude, which is differentiated by
Hippocrepis comosa; (c)
tulipetosum australis subass. nov. (holotypus: rel. 21,
hoc loco), localized at 2000–2200 m of altitude, characterized by
Tulipa australis,
Ornithogalum oligophyllum and
Gagea villosa.
Distribution: On the basis of current knowledge, this association seems to be exclusive of Mt. Chelmos in the northern Peloponnese.
Notes: Previously Quézel and Katrabassa [
40] attributed this vegetation to the Ass. à
Astragalus cylleneus et
Cirsium cylleneum described by Quézel [
35] for Mt. Killini. Effectively as previously emphasized, the aforesaid authors mistakenly attributed these populations of
A. calavrytensis to
A. cylleneus. As clearly can be observed from floristic composition and ecology, the vegetation of Mt Chelmos is well differentiated from that one of Mt. Killini and therefore they must be treated as two distinct associations.
Plantagini graecae-Astragaletum cyllenei Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A36).
Characteristic species: Astragalus cylleneus, Alopecurus gerardii, Plantago atrata subsp. graeca, Potentilla recta.
Structure and ecology: The association is localized in small depressions, similar to dolines, in the middle of the carbonatic rock outcrops, where there is a fairly deep soil rich in silt and clay, accumulated as a result of processes of washing away of the surrounding surfaces more or less sloping. It was surveyed in the supra-temperate sub-Mediterranean bioclimatic belt at 1800–2000 m of altitude. Physiognomically, it is dominated by Astragalus cylleneus, usually associated with numerous other orophytes of the alliance and higher syntaxa. The deep and compact soil justifies the occurrence of mesic species of the Trifolion parnassi, such as Alopecurus gerardii, Plantago atrata subsp. graeca and Potentilla recta. The arrangment of this association in the Festuco achaicae-Marrubion cyllenei rather than in the Trifolion parnassi is justified by the fact that from the structural point of view it is a shrub vegetation of tragacanthoid type, as most of the community of the alliance in question and not of a meadow with prevalence of small herbaceous hemicryptophytes. In addition, the floristic contingent of species of the Cerastio candidissimi-Astragaletea rumelici as well as the related alliance is clearly prevalent respect to that one of Trifolietalia and Trifolion parnassi.
Distribution: This association was observed on Mt. Killini, where is localized exclusively on Mt. Simios.
Notes: The Plantagini graecae-Astragaletum cyllenei is floristically and ecologically quite related to the Nepeto epiroticae-Astragaletum corynthiaci occurring on M. Parnassuss. In fact, both associations are characterized by the dominance of vicariant tragacantoidi species of Astragalus and by the occurrence of species of Trifolion parnassi. In addition, they are localized exclusively in stands more or less depressed with very thick and compact soils, poor in skeleton.
Festuco achaicae-Minuartietum stellatae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A37).
Syn.: Aggr. à Minuartia stellata Quézel & Katrabassa 1974, Rev. Biol. Ecol. Medit. 1(1):18.
Comm. à Minuartia stellata Georgiadis & Dimopoulos 1993, Bot. Helv. 103:160.
Characteristic species: Minuartia stellata, Festuca jeanpertii subsp. achaica, Allium cylleneum.
Structure and ecology: The association is linked to rocky stands with calcareous outcrops or to compact rocky surfaces, more or less sloping at the foot of vertical walls. It is a habitat of semirupestrian type, with soils present only in rocky crevices or in small ledges. This vegetation seems to have its optimum in the oro-temperate sub-Mediterranean bioclimatic belt, at 2000–2250 m of altitude; examples can be observed also at lower altitudes (up to 1800 m) within the supra-temperate sub-Mediterranean belt. Physiognomically, the association is differentiated by the occurrence of compact and large pulvini of
Minuartia stellata, sometimes mixed with smaller other ones of
Asperula boissieri. The characteristics of the alliance of higher syntaxa are here well represented; among these show a greater diffusion and coverage
Astragalus angustifolius subsp.
erinaceus, Festuca janpertii subsp.
achaica and
Festuca cyllenica subsp.
cyllenica. The association is a typical edaphophilous aspect, colonizing small areas scattered in midst of the tragacanthoid community of
Cirsio hypopsilii-Astragaletum taygetici or
Plantagini graecae-Astragaletum cyllenei. Previously, it was described by Quézel and Katrabassa [
40] as. aggr. à
Minuartia stellata and by Georgiadis and Dimopoulos (ref. [
42] as comm. à
Minuartia stellata).
Distribution: This association is well represented on Mt. Chelmos, Mt. Killini and Mt Klokos in the northern Peloponnese.
Notes: The
Festuco achaicae-Minuartietum stellatae can be considered as a southern vicariant of the
Erysimo parnassi-Minuartietum stellatae, association described by Quézel [
35] for Mt. Parnassus.
Alysso taygetei-Plantaginetum alpestris Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A38).
Characteristic species: Alyssum taygeteum, Plantago holosteum var. alpestris, Scorzonera mollis.
Structure and ecology: The association is linked to cacuminal stations and very windy stands, localizing on carbonate substrata flaking in platelets, with primitive or very immature soils. It is distributed within the oro-temperate sub-Mediterranean bioclimatic belt, at 2000–2100, where it has its optimum on surfaces strongly eroded and subject to gelifluxion. It is a low pulvinar vegetation characterized by small, often prostrate, chamaephytes, in which play a significant role Alyssum taygeteum and Plantago holosteum var. alpestris, growing usually together with Astragalus angustifolius subsp. erinaceus and Astragalus rumelicus subsp. taygeticus.
Distribution: The association was observed only on Mt. Chelmos in the northern Peloponnese.
Hieracio sartoriani-Seslerietum tenerrimae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A39).
Syn.: Ass. à Astragalus cylleneus et Cirsium cylleneum subass. à Festuca varia facies à Sesleria coerulans Quézel & Katrabassa 1974, Rev. Biol. Ecol. Medit. 1(1):18.
Characteristic species: Sesleria tenerrima, Hieracium sartorianum, Arenaria cretica var. stygia, Galium incanum subsp. incanum, Silene auriculata.
Structure and ecology: The association is localized on the rocky ridges, sometimes cacuminal, on substrates consisting of compact craked limestone, with soil present only in the rocky ravines and ledges. It was observed at 1900–2350 m of altitude, within the oro-temperate sub-Mediterranean bioclimatic belt, in ecologically very specialized contexts. In fact, in these stands there are very rigid environmental conditions, as strong winds, soil erosion, marked acclivity, gelifluction, etc. This vegetation represents a typical orophilous thinned out grassland, dominated by Sesleria tenerrima. In the middle of the tuffs of this grass grow several hemicryptophytes and chasmophyte, that highlight the semirupestrian characteristics of the habitat. This association clearly constitutes an edaphophilous aspect, replaced in typically rocky habitats by casmophilous comminities of Asplenietea trichomanis.
Distribution: Based on current knowledge, the association is known only for Mt. Chelmos in northern Peloponnese.
Notes: Within this association some relevés carried out by Quézel and Katrabassa [
40] and considered by them as a facies à
Sesleria caerulans of the subass. à
Festuca varia of the ass. à
Astragalus cylleneus et
Cirsium cyllenneum can be included.
Asperulo boissieri-Festucetum cyllenicae Georgiadis & Dimopoulos ex Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A40).
Syn.: Festuco cyllenicae-Asperuletum boissieri Georgiadis & Dimopoulos 1993, Bot. Helv. 103(2):158, nom. inval. (art. 5).
Holotypus: Table 3, rel. 1, Georgiadis and Dimopoulos [
42],
hoc loco.Characteristic species: Festuca cyllenica subsp. cyllenica, Dianthus integer subsp. minutiflorus.
Structure and ecology: The association colonizes the rocky calcareous substrata and stabilized screens, more or less sloping with shallow undeveloped and heavily skeletal soils. It is usually distributed at 2000 and 2200 m of altitude, coming down sometimes up to 1800 m, within the oro-temperate sub-Mediterranean bioclimatic belt, penetrating marginally also in that one supra temperate sub-Mediterranean belt. Physiognomically, it is differentiated by the dominance of large tufts of Festuca cyllenica subsp. cyllenica, that, sometimes, are mixed with those ones of Sesleria vaginalis. Scattered with these grasses there are some low prostrate pulvini of Asperula boissieri and Astragalus angustifolius subsp. erinaceus. It usually assumes a climatophilous role in the higher cacuminal places of the mountains.
Distribution: The association is known only for Mt. Killini in the northern Peloponnese.
Notes: This association was described by Georgiadis and Dimopoulos [
42] from various stands of Mt. Killini and included by them with some perplexity within the
Astragalo-Seslerion, due to the occurrence of a relevant number of characteristics of the
Eryngio-Bromion. However, this syntaxon is an invalid name, because the authors do not indicate the relevé type of the association.
Ranunculo brevifolii-Seslerietum tenerrimae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A41).
Characteristic species: Sesleria tenerrima, Ranunculus brevifolius, Ranunculus sartorianus, Dianthus serratifolius subsp. abbreviatus.
Structure and ecology: The association is linked to stabilized screes characterized by a marked acclivity and occurrence of undeveloped soils with a high percentage of skeleton. It is located at 2000–2100 m of altitude, within the oro-temperate sub-Mediterranean bioclimatic belt. It is a typical grassland characterized by the dominance of Sesleria tenerrima, showing a very scattered coverage, interspersed with small bare sufaces. Mixed with this grass there are tuffs of Festuca cyllenica subsp. cyllenica, which often show a high coverage, and several quite significant rosulate hemicryptophytes, such as Ranunculus brevifolius, Ranunculus sartorianus, Dianthus serratifolius subsp. abbreviatus. For its peculiar ecology, the association must to be considered as an edaphophilous aspect, which tends due to the natural evolution of the soil, towards pulvinate communities, structurally more evolved.
Distribution: The association occurs only in Killini massif on Mt. Simios (northern Peloponnese).
Astragaletum hellenico-erinacei Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A42).
Characteristic species: Astragalus angustifolius subsp. erinaceus, A. hellenicus.
Structure and Ecology: The association is localized on fairly inclined rocky slopes, with soils more or less deep and rich in coarse skeleton. From the structural point of view, the vegetation is differentiated by the dominance of tragacanthoid pulvini of Astragalus angustifolius subsp. erinaceus, which grows togheter with several chamaephytes and hemicryptophytes, among them Astragalus hellenicus, endemic species, rare in the Peloponnese. This association, showing usually a climatophilous character, at least in the rocky cacuminal stands, represents often a substitution aspect, replacing the forests of Abies cephalonica as a result of soil degradation processes.
Distribution: The association was suveyed only on Mt. Menalon in the Central Peloponnese.
Notes: On the whole, it can be considered as a thermophilous vicariant of the community with Astragalus rumelicus subsp. taygeticus occurring in the other massifs of the northern Peloponnese.
Festucetum polito-cyllenicae Maroulis & Georgiadis 2005, Fitosociologia 42(1):44, corr. (
Appendix C,
Table A43).
Syn.: Festuco politae-Festucetum cyllenicae Maroulis & Georgiadis 2005, Fitosociologia 42(1):44.
Holotypus: Table 2, rel. 509, Maroulis and Georgiadis [
44].
Characteristic species: Festuca cyllenica subsp. cyllenica, Festuca polita, Campanula albanica subsp. albanica and Taraxacum delphicum.
Structure and ecology: The association is localized along the very sloped surfaces on stabilized screes or rocky stands with undevelopped calcareous soils rich in scheleton. It is widespread at 1750–2200 m of altitude, within the supra-temperate and oro-temperate sub-Mediterranean bioclimatic belts, where it plays a climatophilous role. This vegetation constitutes dense orophilous grasslands dominated by Festuca cyllenica subsp. cyllenica, Festuca polita and Sesleria vaginalis, where are frequent several other hemicryptophytes and small chamaephytes.
Distribution: According to literature, it is widespread on various mountains of the Erimanthos massif in the Northern Peloponnese.
Notes: The association described by Maroulis and Georgiadis [
44] as
Festuco politae-Festucetum cyllenicae, was included by the authors in the
Eryngio-Bromion although there is a significant contingent of characteristics of
Astragalo-Seslerion. For its structure and ecology, as well as for its floristic composition, this association is quite related to
Asperulo boissieri-Festucetum cyllenicae from Mt. Killini, from which differs mainly for its floristic set.
Arenario filicaulis-Festucetum cyllenicae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A44).
Characteristic species: Festuca cyllenica subsp. cyllenica, Arenaria filicaulis subsp. filicaulis, Ranunculus psilostachys.
Structure and ecology: The association occurs mainly on stabilized screes or on quite inclined slopes covered by calcareous stones, mixed with scarce humus. It is surveyed at 1500 and 1600 m of altitude on northern slopes, it colonizes large surfaces. Physiognomically, it is characterized by the dominance of large tufts of Festuca cyllenica subsp. cyllenica, which grows very well on inclined slopes subject to long periods of snow coverage. This vegetation, where it is frequent also Festuca jeanpertii subsp. achaica, results well differentiated from the other associations with Festuca cyllenica subsp. cyllenica for the occurrence of Arenaria filicaulis subsp. filicaulis and Ranunculus psilostachys.
Distribution: The association is frequent on Mt. Panachaiko in the northern Peloponnese.
Aurinio moreanae-Lomelosietum crenatae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A45).
Characteristic species: Aurinia moreana, Lomelosia crenata subsp. crenata.
Structure and ecology: The association is localized in semirupestrian habits on very sloped (70–80°) limestone outcrops, usually showing a northern exposure. This vegetation dominated by Lomelosia crenata subsp. crenata and Aurinia moreana, is distributed at 1600 and 1700 m of altitude. It is an edaphophilous community, replacing in this rocky stand the Cirsio hypopsilii-Astragaletum taygetici. On the whole, the species of the alliance and higher syntaxa are here well represented, among them there are Festuca jeanpertii subsp. achaica, Astragalus angustifolius subsp. erinaceus, Astragalus rumelicus subsp. taygeticus, Achillea umbellata, etc.
Distribution: The association was surveyed only on Mt. Klokos in northern Peloponnese.
Onosmo malickyi-Astragaletum hellenici Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A46).
Characteristic species: Onosma erectum subsp. malickyi, Astragalus hellenicus, Alyssum murale.
Structure and ecology: The association occurs at 1300–1400 m of altitude, in the clearing within the Abies cephalonica woodlands. The surfaces are slightly inclined, and the soils are covered with a bed of fir needles. Floristically, it is characterized by hemicryptophytes and small chamaephytes, among them Onosma erectum subsp. malickyi, Astragalus hellenicus, Alyssum murale, Helianthemum hymettium and Festuca jeanpertii subsp. achaica.
Distribution: This community was surveyed only on Mt. Chelmos near Mavros Logos.
Violo graecae-Festucetum cyllenicae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A47).
Characteristic species: Festuca cyllenica subsp. cyllenica, Viola graeca, Ornithogalum oligophyllum.
Structure and ecology: The association occurs on calcareous stabilized screes and stony slopes at 2000 and 2500 m of altitude. It is a pioneer vegetation linked to slightly inclined surfaces and poor in soil. Physiognomically, it is characterized by the dominance of Festuca cyllenica subsp. cyllenica, which constitute wide grasslands where occur several orophytes of higher syntaxa. Small hemicryptophytes and geophytes found often refuge among the tuffs of this plant, among them Viola graeca, Ornithogalum oligophyllum, Allium frigidum, Galium taygeteum, Geocaryum peloponnesiacum, etc.
Distribution: This association is widespread on Mt. Chelmos.
Tripodio graeci-Helictotrichetum heldreichii Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A48).
Characteristic species: Helictotrichon convolutum subsp. heldreichii, Tripodion graecum.
Structure and ecology: The association occurs at 1400 and 1600 m of altitude, in the large rock clearing within the Abies cephalonica woodlands. Normally it is frequent along the more or less inclined slopes characterized by rocky outcrops with very shallow and immature soils. The occurrence of Tripodion graecum is significant, i.e., since it is a species known only from a few places in the Peloponnese and Anatolia (mainly in the Taurus region). It is usually associated with Helictotrichon convolutum subsp. heldreichii, generally with high values of coverage, and Festuca jeanpertii subsp. achaica. In this vegetation the species of higher syntaxa are overall well represented.
Distribution: The association is widespread in the lower montane belt of Mt. Menalon, Central Peloponnese.
Syn.: Eryngieto-Bromion Quézel 1964, Vegetatio, 12:326, p.min.p., nom. ambig. rejic. propos. (art. 36).
Eryngio multifidi-Bromion fibrosi Quézel 1964, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18:82 p.min.p., nom. ambig. rejic. propos. (art. 36).
Astragaleto-Seslerion Quézel 1964, Vegetatio, 12:326, p.min.p., nom. ambig. rejic. propos. (art. 36).
Astragalo angustifolii-Seslerion coerulantis Quézel 1964, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18:82, p.min.p., nom. ambig. rejic. propos. (art. 36).
Stipeto-Morinion Quézel 1964, Vegetatio, 12: 26, p.min.p., nom. ambig. rejic. propos. (art. 36).
Stipo pulcherrimae-Morinion persicae Quézel 1964, corr. Quézel, Barbero & Akman 1992, Ecol. Medit. 18:82, p.min.p., nom. ambig. rejic. propos. (art. 36).
Holotypus: Sideritido clandestinae-Astragaletum taygetici Musarella, Brullo & Giusso ass. nov.
Characteristic species: Achillea setacea, Achillea taygetea, Allium pycnotrichum, Anthemis laconica, Asperula boryana, Asperula mungieri, Astragalus taygeteus, Asyneuma psaridis, Crepis heldreichiana, Nepeta camphorata, Phitosia crocifolia, Sideritis clandestina subsp. clandestina, Viola sfikasiana.
Structure and ecology: It gathers, likewise to the previous alliances included in order Eryngio multifidi-Armerietalia orphanidis, the orophilous plant communities rich in chamaephytes and nanophanerophytes, often with pulvinate habit, as well as in hemicryptophytes, while rarer are the geophytes. On the whole, the associations belonging to this alliance show a more marked thermophily than those ones of the other two alliances. In addition, the considerable contingent of endemics that characterizes this syntaxon is represented mainly by species taxonomically quite isolated or otherwise of remarkable phytogeographical significance. From the bioclimatic point of view, this alliance falls in an area affected by termotypes referring to supra-and oro-Mediterranean, since one detects a long period of high summer dryness enough, although there is a certain tendency towards the supra- and oro-temperate sub-Mediterranean type, with ombrotypes characterized by scarce rainfall, especially during the summertime.
Distribution: The alliance is confined to the southern Peloponnese including the massifs of the Taygetos and Parnon.
Notes: Into this alliance, analogously to the other two previously described, fall within part of the alliances described by Quézel [
35], namely
Eryngio-Bromion,
Stipo-Morinion, and
Astragalo-Seslerion.
Scabioso taygeteae-Onosmetum leptanthae Quézel 1964, Vegetatio, 12:327 (
Appendix C,
Table A49).
Syn.: Association à Scabiosa taygetea et Onosma leptanthum Quézel 1964, Vegetatio, 12: 327.
Lectotypus: Table 15, rel. 2, Quézel [
35],
hoc loco.Characteristic species: Onosma leptantha, Scabiosa taygetea subsp. taygetea, Calamintha suaveolens, Tripodion graecum.
Structure and ecology: The association is located on rocky outcrops or however more or less rocky surfaces consisting of compact limestone subject to heavy erosion and washing away. The soils are very superficial and localized in crevices and ledges. It is widespread within the meso-mediterranean and supra-Mediterranean bioclimatic belt, at 1250–1800 m of altitude. This vegetation is dominated by chamaephytes and nanophanerophytes of small and medium size, mixed to several caespitose hemicryptophytesche, and among them there are Onosma leptantha, Scabiosa taygetea subsp. taygetea, Pterocephalus perennis spp. perennis, Stipa endotricha, Dasypyrum hordeaceum, Koeleria mitrushii, Bromus riparius, Festuca jeanpertii subsp. jeanpertii. The association has a purely edaphophilous role, although it represents a secondary aspect too, as a result of degradation processes of Abies cephalonica woodlands.
Distribution: The association is distributed on Mt. Taygetos in the southern Peloponnese.
Notes: This association was considered by Quézel et al. [
80] as the nomenclatural type of the alliance
Stipo pulcherrimae-Morinion persicae, although in the relative phytosociological table there are several characteristic species of the other two alliances described by Quézel [
3].
Danthoniastro compacti-Fumanetum alpinae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A50).
Characteristic species: Fumana paphlagonica subsp. alpina, Danthoniastrum compactum.
Structure and ecology: The association is localized on the slightly sloping limestone slabs, especially with an eastern exposure. It is developped within the supra-Mediterranean bioclimatic belt at an altitude of about a 1700 m. The surfaces are free of soil except in the cracks and small depressions, that allow the establishment of a rather sparse vegetation. It is a vegetation rich in small prostrate chamaephytes, sometimes creeping, among them particularly significant are Fumana paphlagonica subsp. alpina, Helianthemum hymettium, Teucrium montanum var. parnassicum and Asperula mungieri. Several hemicryptophytes, such as Danthoniastrum compactum, Festuca jeanpertii subsp. jeanpertii, Koeleria mitrushii and Stipa endotricha are also well represented. This association plays a clearly edaphophilous role replacing the Scabioso taygeteae-Onosmetum leptanthae in the above-mentioned habitats.
Distribution: Basing on the current knowledge, the association is confined to a small area of Mt. Taygetos in the southern Peloponnese.
Sideritido clandestinae-Astragaletum taygetici Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A51).
Syn.: Association à Sideritis theezans, Quézel 1964, Vegetatio, 12:331, nom. illeg. (art. 29).
Characteristic species: Astragalus rumelicus subsp. taygeticus, A. taygeteus, Plantago holosteum var. alpestris, Hypericum olympicum and Arabis subflava.
Structure and ecology: The association is widely distributed on flat and sometimes more or less sloped surfaces, characterized by not very deep soils, rich in minute skeletal component of carbonatic nature. It grows at 1700 and 2100 m of altitude, within the supra-Mediterranean bioclimatic belt with penetrations in oro-Mediterranean that one. This vegetation is physiognomically differentiated by the dominance of flashy tragacanthoid pulvini of Astragalus rumelicus subsp. taygeticus, A. taygeteus and, more rarely, by A. angustifolius subsp. erinaceus. Many other small shrubs are also quite frequent, such as Plantago holosteum var. alpestris, Sideritis clandestina subsp. clandestina, Cerastium candidissimum and several hemicryptophytes. Overall, the association, which usually shows a high value of coverage, must be considered as a climatophilous aspect, spread on all slopes regardless of exposure. From the physiognomic-structural point of view, it is quite related with the other pulvinate tragacanthoid associations dominated by Astragalus rumelicus s.l., such as Cirsio hypopsilii-Astragaletum taygetici, Astragaletum lacteo-taygetici and Marrubio velutini-Astragaletum rumelici.
Distribution: The association is exclusive of Mt Taygetos, in the southern Peloponnese.
Notes: This vegetation was described by Quézel [
35] with the name “Association à
Sideritis theezans”, which actually is not very significant from the floristic-structural viewpoint. In fact,
Sideritis theezans (whose correct name is
S. clandestina subsp.
clandestina) is a small camaephyte with a secondary physiognomically role in the context of this shrubby association, dominated by some tragacanthoid nanophanerophytes, such as
Astragalus rumelicus subsp.
taygeticus and
A. taygeteus. According to art. 29, the syntaxon is an illegitimate name and therefore should be replaced by a new name that expresses in clear way its floristic and physiognomic-structural peculiarities. It is therefore proposed the new name
Sideritido clandestinae-Astragaletum taygetici with a better floristic characterization. In this regard it should be noted that
S. clandestina subsp.
clandestina is not exclusive to this association, but it is an endemic chamaephyte widespread in various orophilous communities of southern Peloponnese and therefore it has been proposed as characteristics of the alliance
Sideritido clandestinae-Asperulion mungieri.
Rindero graecae-Acantholimetum graeci Quézel 1964, Vegetatio, 12:336 (
Appendix C,
Table A52), corr.
Syn.: Association à Acantholimon echinus et Rindera graeca Quézel 1964, Vegetatio, 12:336.
Lectotypus: Table 19, rel. 2, Quézel [
35],
hoc loco.Characteristic species: Sesleria vaginalis, Jurinea taygetea, Minuartia condensata, Campanula papillosa, Erigeron epiroticus, Aethionema carlsbergii, Alyssum taygeteum, Bupleurum sibthorpianum.
Structure and ecology: The association is localized in cacuminal stands of high altitude, about 2200–2400 m, within the oro-Mediterranean bioclimatic belt. It prefers quite acclive surfaces, where it shows a coverage of 40–70%, which decreases significantly at higher altitudes. The substrata are represented by carbonatic rocks that break up into plaquettes or sometimes by semi-stabilized screes. Physiognomically the vegetation is characterized by thorny pulvini of
Astragalus angustifolius subsp.
erinaceus and
Acantholimon graecum, mixed to several caespitose hemicryptophytes, such as
Sesleria vaginalis. In addition, it is very significant the occurrence of some rare endemics exclusive of this vegetation, as
Jurinea taygetea and
Aethionema carlsbergii. This community, showing a clear climatophilous role, is linked to winterproof environmental features, such as the prolonged snow cover, the accentuated phenomena of gelifluction, exposure to cold winds and the occurrence of rocky substrata with shallow and immature soils. The name of this association must be corrected in
Rindero graecae-Acantholimetum graeci since
Acantholimon echinus subsp.
echinus used by Quézel [
35] is taxonomically incorrect and should be attributed to
Acantholimon graecum (see Dimopoulos et al. [
71].
Distribution: The association is confined to the cacuminal higher part of Mt. Taygetos, in the southern Peloponnese.
Notes: This association was included by Quézel [
35] in the
Astragalo-Seslerion and afterwards indicated by Quézel et al. [
80] as the lectotype of this alliance.
Onosmo heterophyllae-Astragaletum erinacei Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A53).
Characteristic species: Onosma heterophylla.
Structure and ecology: The association was surveyed on carbonatic rocky slopes more or less inclined occurring at relatively low altitudes (1300–1500 m), characterized by coarse material mixed with immature soils. It is localized within the meso-Mediterranean bioclimatic belt, usually occupied by Abies cephalonica woodlands. From the physiognomic-structural point of view, this vegetation is differentiated by thorny pulvini of Astragalus angustifolius subsp. erinaceus, that grow together with other shrubs and several caespitose hemicryptophytes; among the latter there are Onosma heterophylla, Stipa endotricha, Festuca jeanpertii subsp. jeanpertii, Koeleria mitrushii, Bromus riparius, Stipa holosericea. It is a substitution community linked to the degradation processes of the woody vegetation, although in strictly rocky conditions it tends to have an edaphophilous role.
Distribution: The association has been surveyed on M. Parnon, exclusively at Prophitis M. Ilias, near Agriani in the Southern Peloponnese.
Notes: This association is closely related to Scabioso taygeteae-Onosmetum leptanthi from Mt. Taygetos, of which it can be considered a geograpical vicariant.
Astragaletum lacteo-taygetici Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A54).
Characteristic species: Astragalus rumelicus subsp. taygeticus, A. lacteus, Cynoglossum pustulatum subsp. parviflorum.
Structure and ecology: The association is linked to rocky slopes characterized by very compact limestone with soil accumulating only in crevices and depressions of the rocks. It is well developed between 1400 and 1800 m of altitude, within the meso-Mediterranean and supra-Mediterranean bioclimatic belts, constituting usually a climatophilous vegetation which tends to expand towards lower elevations as a result of the degration processes of Abies cephalonica forest. This vegetation is physiognomically characterized by tragacanthoid pulvini of Astragalus rumelicus subsp. taygeticus and A. angustifolius subsp. erinaceus, in the midst of which grow several small chamaephytes and caespitose or rosulate hemicryptophytes.
Distribution: The association occurs only on the massif of Parnon, in the southern Peloponnese, where it is common in several mountains.
Notes: From the physiognomic-structural and partially floristic viewpoint, this association is quite similar to Sideritido clandestinae-Astragaletum taygetici from Mt. Taygetos, differing, however especially for the dynamic role, since the latter association is distributed in a higher altitudinal belt.
Violo parnoniae-Astragaletum erinacei Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A55).
Characteristic species: Viola parnonia, Astragalus agraniotii, Centaurea parnonia.
Structure and ecology: The association covers the rather inclined rocky slopes characterized by generally undeveloped calcareous soils, sometimes represented by lithosols. It is widespread within the bioclimatic supra-Mediterranean bioclimatic belt at 1700–1900 m of altitude, regardless of exposure. In this community plays a significant physiognomic role Astragalus angustifolius subsp. erinaceus, which tends to constitute with its characteristic compact thorny pulvini extensive populations. Furthermore, the occurrence of several chamaephytes and hemicryptophytes, including in particular some rare endemics such as Viola parnonia, Astragalus agraniotii and Centaurea parnonia, differentiate vey well this vegetation from other ones of this alliance. Based on the edaphic characteristics, it is possible to distinguish two subassociations, indicated as astragaletosum erinacei and asperuletosum malevonensis, which will be examined below.
Distribution: The association was surveyd exclusively on Megali Tourla, which is the highest mountain of the Parnon Massif in the southern Peloponnese.
- (a)
astragaletosum erinacei Musarella, Brullo & Giusso subass. nov.
hoc loco (
Appendix C,
Table A55, rel. 1–3).
Characteristic species: Astragalus angustifolius subsp. erinaceus (dominant).
Structure and ecology: It represents the typical aspect of the association and is localized on the inclined slopes of the summit, where it colonizes surfaces rich in clastic stabilized material with more or less deep and rich in coarse skeleton soils. Physiognomically, it is differentiated by the dominance of Astragalus angustifolius subsp. erinaceus, which tends to cover most of the surface occupied by the association. This subassociation plays a prevalently climatophylous role, although currently it is also widespread in stands in the past occupied by Abies cephalonica forests, where has a secondary meaning as a result of degradation processes of soil.
Distribution: See association.
- (b)
asperuletosum malevonensis Musarella, Brullo & Giusso subass. nov.
hoc loco (
Appendix C,
Table A55, rel. 4–11).
Characteristic species: Achillea umbellata, Asperula malevonensis, Helianthemum canum subsp. canum.
Structure and ecology: It replaces the typical aspect in correspondence of compact outcrops, consisting of more or less cracked limestones. In these stands, some species which show markedly chasmophytic habit often occur, such as Achillea umbellata, Helianthemum canum subsp. canum and the local endemic Asperula malevonensis, thus providing a distinct physiognomy. It is an edaphophilous aspect showing a scarce coverage, forming small patches in the middle of the previous subassociation.
Distribution: See association.
Holotypus: Asperulion samiae Musarella, Brullo & Giusso all. nov. hoc loco.
Characteristic species: Acantholimon aegaeum, Aethionema saxatile subsp. creticum, Alopecurus davisii, Astragalus angustifolius subsp. aegeicus, Atraphaxis billardierei, Bunium microcarpum subsp. microcarpum, Centaurea urvillei subsp. urvillei, Dianthus zonatus, Draba heterocoma subsp. archipelagi, Erysimum hayekii, Galium heldreichii, Inula heterolepis, Jurinea cadmea, Koeleria lobata, Minuartia anatolica var. polymorpha, Noaea mucronata, Paracaryum aucheri, Paronychia chionaea, Pterocephalus pinardii, Sesleria anatolica, Sideritis sipylea, Silene urvillei, Stachys cretica subsp. smyrnaea, Verbascum pycnostachyum.
Structure and ecology: This order groups the orophilous pulvinate plant communities dominated by chamaephytes and nanophanerophytes with tragacanthoid habit linked to cacuminal very sunny and windy stands at altitudes higher than 900–1000 m. The substrates are prevalently carbonatic with immature soils rich in coarse skeleton occurring mainly in the rocky crevices. The vegetation belonging to this syntaxon colonize mainly rocky surfaces more or less denuded, often quite sloping, affected by moist marine winds or a regime of mists. During the winter period these stations are usually covered for quite short periods by snow. On the basis of investigations carried out in the Aegean area, the habitats colonized by this type of vegetation are represented mainly by the summit areas of island mountains, where the peculiar environmental conditions above emphasized can be found. From the bioclimatic point of view, the order is linked to mountain or high mountain habitats falling into meso- and supra-Mediterranean belts, extending marginally also in the oro-Mediterranean one. Floristically, the order is characterized by a rich contingent of species having mainly an East Aegean-Anatolian distribution, including also several rare endemics.
Distribution: Basing on the current knowledge, the order seems distributed on the mountains of some north-eastern Aegean islands, such as Samos, Chios, Lesvos and Thassos. It is likely that plant communities related to this syntaxon are also present on the island of Samothraki, Mt. Athos, and some coastal mountains of western Anatolia.
Notes: The Noaeo mucronatae-Silenetalia urvillei must be considered as the eastern vicariant of Eryngio multifidi-Armerietalia orphanidis, distributed in the mainland of central-southern Greece, as well as in some Ionian Islands and Euboea.
Holotypus: Astragaletum samii Musarella, Brullo & Giusso ass. nov. hoc loco.
Characteristic species: Allium hirtovaginatum subsp. samium, A. orosamium, Alyssum samium, Anthemis samia, Asperula samia, Erodium sibthorpianum. subsp. vetteri, Satureja spinosa var. glabra, Thymus samius.
Structure and ecology: This alliance gathers the plant communities occurring in cacuminal stands of island mountains, localized at 900–1400 m of altitute. It is an essentially calcicolous syntaxon linked to a meso-Mediterranean bioclimatic belt, extending towards the supra-Mediterranean one. It has its best expression in very peculiar ecological conditions, where some environmental factors play an important role, such as the marine moist winds, rather cold during the autumn and winter, the erosive action of weathering on rocky surfaces, the mists, and the marked summer dryness. Floristically, it is differentiated by a rich endemic and rare species contingent, having a considerable taxonomical and phytogeographical significance.
Distribution: The alliance is confined to the mountains of the island of Samos in the eastern Aegean.
Characteristic species: Astragalus creticus subsp. samius and Allium orosamium.
Structure and ecology: The association is localized on calcareous slopes of cacuminal areas at 1000–1400 m of altitude, where it tends to cover wide surfaces. It has its best expression on compact rocky substrates, often very sloping, represented by calcareous outcrops and buttresses, with soils present almost exclusively in the crevices and ledges. From the bioclimatic viewpoint, this vegetation grows within the meso-Mediterranean belt, extending marginally also in the oro-Mediterranean one, showing a role, not strictly climatophilous, but rather of edaphophilous vegetation. However, it is spread in an area located above the limit of the forests, consisting mainly of Pinus brutia and Quercus calliprinos woodlands. In the tracts with deeper and mature soils, this pulvinate vegetation is mixed to relict of orophilous conifer forest of Junipero-Pinetea sylvestris, where Juniperus foetidissima and J. oxycedrus play an important role. Floristically, the association is characterized by the dominance of thorny pulvini of Astragalus creticus subsp. samius, punctiform endemic of considerable phytogeographical interest. Several relevant endemic orophytes, such as Allium orosamium, Alyssum samium, Anthemis samia, Asperula samia, Erodium sibthorpianum. subsp. vetteri, and Thymus samius occur in this association and probably also Centaurea xylobasis, a rare endemic exclusive of these cacuminal stands.
Distribution: The association is exclusive of the cacuminal area of Mt. Kerkis in the island of Samos (East Aegean).
Thymo samii-Astragaletum condensati Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A57).
Characteristic species: Astragalus condensatus (=A. ptilodes), Valeriana dioscoridis, Phlomis grandiflora Fritillaria carica, Centaurea cariensis subsp. maculiceps, Petrorhagia armeriodes, Vincetoxicum canescens subsp. peduncolatum, Allium karvounis, Lomelosia polykratis, Ranunculus rumelicus, Crocus oliveri subsp. balansae.
Structure and ecology: The association is localized in a cacuminal area characterized by outcrops of compact crystalline limestone (marble), with surfaces flat or slightly sloping. The soils are very shallow and fill the depressions and cracks of the rock. The area in which it is developed, localized at 1100–1200 m of altitude, falls within the meso-Mediterranean bioclimatic belt. From the structural point of view, it is observed the dominance of low pulvinate or creeping shrubs, among them Astragalus condensatus, tragacanthoid species, playing a relevant role, and various other small shrubs, such as Asperula samia, Noaea mucronata, Satureja spinosa var. glabra, Thymus samius, etc. In this association are found numerous endemic species rather rare exclusive of these cacuminal stands. Outside of the limestone outcrops, in correspondence of the schistose substrata, this pulvinate vegetation is replaced by Pinus pallasiana woodlands, adaphically much more exigent. On the whole, this vegetation has a clear edaphophilous role.
Distribution: The association is exclusive of cacuminal area of Mt. Ambelos in the island of Samos (East Aegean).
Notes: The Thymo samii-Astragaletum condensatis can be considered a vicariant of the Astragaletum samii, occurring on different substrata in another mountain of Samos.
Campanulo lyratae-Genistetum parnassicae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A58).
Holotypus: Table 11, rel. 5, Christodoulakis and Georgiadis [
41],
hoc loco.Characteristic species: Genista parnassica, Campanula lyrata subsp. lyrata.
Structure and ecology: Based on the relevés published by Christodoulakis and Georgiadis [
41], at altitude lower than 1000 m always in calcareous rocky stands, more or less sloping, the
Astragaletum samii is replaced by another type of shrub pulvinate vegetation, differentiated by the dominance of
Genista parnassica. In the places occupied by this vegetation,
Astragalus creticus subsp.
samius is wholly absent, as well as the species most significant of the alliance and order decrease and become quite rare. This community, which is proposed as
Campanulo lyratae-Genistetum parnassicae, therefore, can be considered as a vicariant of low altitude of the
Astragaletum samii. From the bioclimatic point of view, the association is distributed in the meso-Mediterranean belt.
Distribution: The association is known only to Mt. Kerkis in the island of Samos (East Aegean).
Arenario guicciardii-Seslerietum anatolicae Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A59).
Characteristic species: Sesleria anatolica, Arenaria guicciardii, Pimpinella peregrina.
Structure and ecology: The association is localized in calcareous markedly sloping rocky places with soils occurring only in the cracks and the crevices. It seems to have its optimum at 900–1000 m of altitude on a little sunshine surface especially with northern exposure, within the meso-Mediterranean bioclimatic belt. This vegetation is dominated by Sesleria anatolica which grows togheter with a rich contingent of small pulvini or creeping shrubs, such as Anthemis samia, Inula heterolepis, Noaea mucronata, Satureja spinosa var. glabra, Sideritis sipylea, etc. As concerns its dynamic role, it is a community prevalently edaphophilous, occurring within the climatophilous Pinus brutia forests, which is linked to surfaces with mature and more or less deep soils.
Distribution: The association was surveyed on Mt. Kerkis in the island of Samos (East Aegean).
Notes: The Arenario guicciardii-Seslerietum anatolicae tends to replace the Astragaletum samii at altitudes lower than 1000 m, limited to shady and fresh stands.
Holotypus: Anthemido discoideae-Astragaletum aegeici Musarella, Brullo & Giusso ass. nov. hoc loco.
Characteristic species: Anthemis cretica subsp. leucanthemoides, Astragalus lesbiacus, Crepis sancta subsp. nemausensis, Erysimum hayekii, Festuca pseudosupina.
Structure and ecology: The alliance gathers pulvinate plant communities dominated by small tragacanthoid shrubs occurring in the mountain cacuminal stands of insular mountains. They are localized at 900–1300 m of altitude, mainly within the meso-Mediterranean bioclimatic belt. The associations falling in this sintaxon are very specialized and linked to very peculiar edaphic and bioclimatic conditions. They are circumscribed to carbonatic substrata represented by ridges and rocky outcrops, with soils present only in the cracks and crevices. The alliance is floristically differentiated by endemic species exclusive to these summit stands, that emphasized their marked geographical isolation.
Distribution: The alliance is circumscribed to the East Aegean islands of Lesvos and Chios.
Notes: This syntaxon can be considered as a geographical vicariant of the Asperulion samiae.
Anthemido discoideae-Astragaletum aegeici Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A60, rel. 1–6).
Characteristic species: Astragalus angustifolius subsp. aegeicus, Anthemis aciphylla subsp. discoidea, Allium stamineum s.l., Silene lesbiaca, Paronychia macrosepala.
Structure and ecology: The association is restricted to cacuminal stands at 900–967 m of altitude, on compact limestone with very superficial soils confined to the crevices of the rock. It is developed within the meso-Mediterranean bioclimatic belt on rocky surfaces usually well exposed and windy. In this vegetation small often thorny pulvini occur, among which a relevant physiognomic role is played by Astragalus angustifolius subsp. aegeicus, Inula heterolepis, Noaea mucronata, Silene urvillei, Sideritis sipylea, Anthemis aciphylla subsp. discoidea, mixed to which there are some caespitose grasses, such as Festuca pseudosupina and Koeleria lobata. It is clearly an edaphophilous vegetation closely related to very peculiar environmental conditions, such as eroded soils, marked winds and mist regime. These factors taken together do not allow a natural evolution of the vegetation towards more mature forms, such as Pinus brutia pine forest widespread in the surrounding areas.
Distribution: The association is exclusive of Mt. Olymbos in the island of Lesbos (Eastern Aegean).
Diantho zonati-Astragaletum lesbiaci Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A60, rel. 7–11).
Characteristic species: Astragalus lesbiacus, Dianthus zonatus, Petrorhagia armerioides.
Structure and ecology: The association is localized on outcrops of calcareous rocks characterized by very shallow and immature soils. Usually it grows on fairly flat surfaces at altitudes between 700 and 800 m. Physiognomically, it shows a coverage rather scattered characterized by small prostrate shrubs, representated mainly by Astragalus lesbiacus, A. angustifolius subsp. aegeicus and Dianthus zonatus. The association usually covers small surfaces interspersed with uncultivated or reforested areas.
Distribution: The association was observed only on Mt. Marathovounos in Chios island.
Galio insularis-Thymetum sypilei Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A60, rel. 12–16).
Characteristic species: Thymus sipyleus, Minuartia attica subsp. idaea, Galium brevifolium subsp. insulare, Minuartia mesogitana subsp. kotschyana, Asyneuma virgatum subsp. cichoriforme.
Structure and ecology: The association colonizes the rocky ridges very windy and washed away at altitudes above 1100 m. The vegetation is rather thinned out with small dwarf shrubs that grow in the cracks of rocks. The more frequent species in this vegetation are Thymus sipyleus, Minuartia attica subsp. idaea, Galium brevifolium subsp. insulare, Minuartia mesogitana subsp. kotschyana, Festuca pseudosupina, Centaurea urvillei subsp. urvillei, Pterocephalus pinardii and Euphorbia herniariifolia. This vegetation is very degraded and floristically impoverished mainly due to heavy grazing
Distribution: The association occurs on Mt. Pelineon in Chios.
Acantholimo aegaei-Astragaletum lesbiaci Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A60, rel. 17–21).
Characterstics species: Astragalus lesbiacum, Acantholimon aegaeum, Thymus zygioides.
Structure and ecology: The association occurs on the cacuminal limestone plateaux, where it colonizes washed rocky surfaces at altitudes between 800 and 900 m. It is characterized by the dominance of pulvinate tragacanthoid shrubs, such as Astragalus lesbiacus, A. angustifolius subsp. aegeicus, Acantholimon aegaeum, and Silene urvillei.
Distribution: The association was surveyed exclusively on Mt. Plakes in Chios island.
Holotypus: Paronychio bornmuelleri-Astragaletum odoniani Musarella, Brullo & Giusso ass. nov. hoc loco.
Characteristic species: Anthemis tenuiloba, Festuca hirtovaginata, Galium insulare, Inula aschersoniana var. athoa, Satureja montana subsp. macedonica, Sesleria achtarovii.
Structure and ecology: This alliance brings plant communities linked to carbonatic substrates of mountain and high-mountain stands, dominated by thorny pulvini. This vegetation is distributed at altitudes above 900 m, where it is localized in places usually represented by summit rocky plateaux, ridges and cacuminal areas with very superficial and immature soils, present mainly in small depressions and crevices. From the bioclimatic viewpoint, this alliance is distributed within the meso-Mediterranean belt, with ombrotype more or less humid, even during the summer, penetrating probably in that supra-Mediterranean one. Floristically, it is differentiated by a set of endemic species with North Aegean distibution.
Distribution: The alliance is currently known only for the island of Thassos in the northen Aegean, but based on the geographic distribution of characteristic species, problably it occurs also in the coastal mountains of North Greece.
Notes: This syntaxon can be considered as a northern vicariant of the other two alliances included in the Noaeo mucronatae-Silenetalia urvillei previously described.
Paronychio bornmuelleri-Astragaletum odoniani Musarella, Brullo & Giusso ass. nov.
hoc loco (
Appendix C,
Table A61).
Characteristic species: Astragalus angustifolius subsp. odonianus, Paronychia bornmuelleri, Cerastium moesiacum subsp. glutinosum, Allium cremnophilum, Dianthus gracilis subsp. xanthianus, Minuartia verna var. thasia.
Structure and ecology: The association is confined to the summit very windy and sunny plateaux, consisting of crystalline limestones, located at altitudes between 900 and 1000 m, falling within the bioclimatic meso-Mediterranean belt. It occurs on rocky substrates with very superficial and immature soils, reaching its maximum expression in situations of ridge. The vegetation is dominated by thorny pulvini of Astragalus angustifolius subsp. odonianus, which forms large populations mixed with small prostrate chamaephytes (Dianthus gracilis subsp. xanthianus, Minuartia verna var. thasia, Paronychia bornmuelleri, Cerastium moesiacum subsp. glutinosum) and several hemicriptophytes represented mainly by caespitose grasses (Festuca hirtovaginata, Sesleria achtarovii, Koeleria lobata, Stipa endotricha). This vegetation is typically edaphophile, since it is linked to peculiar environmental conditions that do not allow the normal development of the soil. In edaphic more mature situations, the association is usually replaced by Juniperus excelsa woodlands.
Distribution: The association is confined to rocky outcrops of the cacuminal stands of Mt. Ipsario in the island of Thassos (northern Aegean).






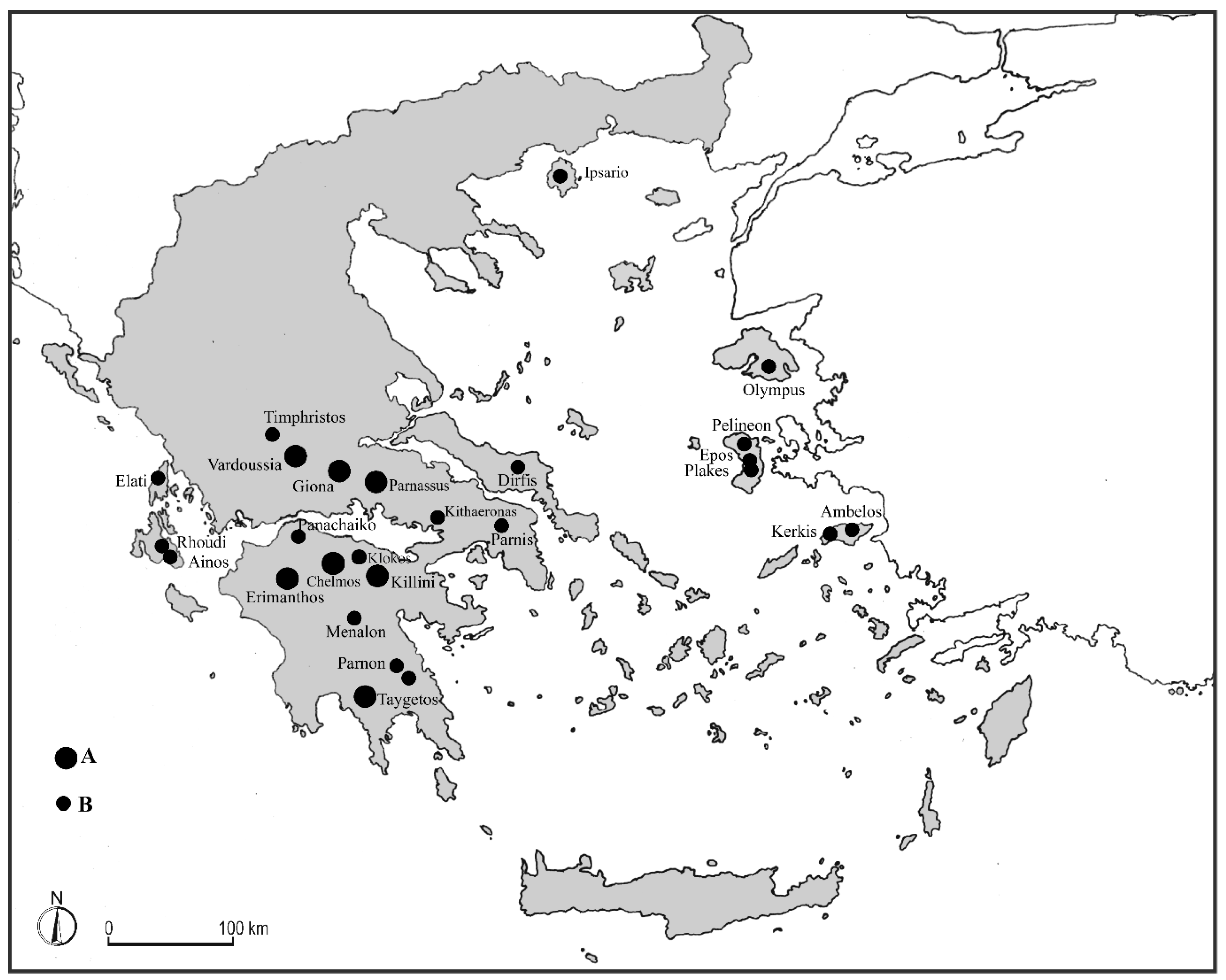{kind=link}
{kind=link}






































































