Significance of Plant Growth Promoting Rhizobacteria in Grain Legumes: Growth Promotion and Crop Production

 ,
,
Abstract
1. Introduction
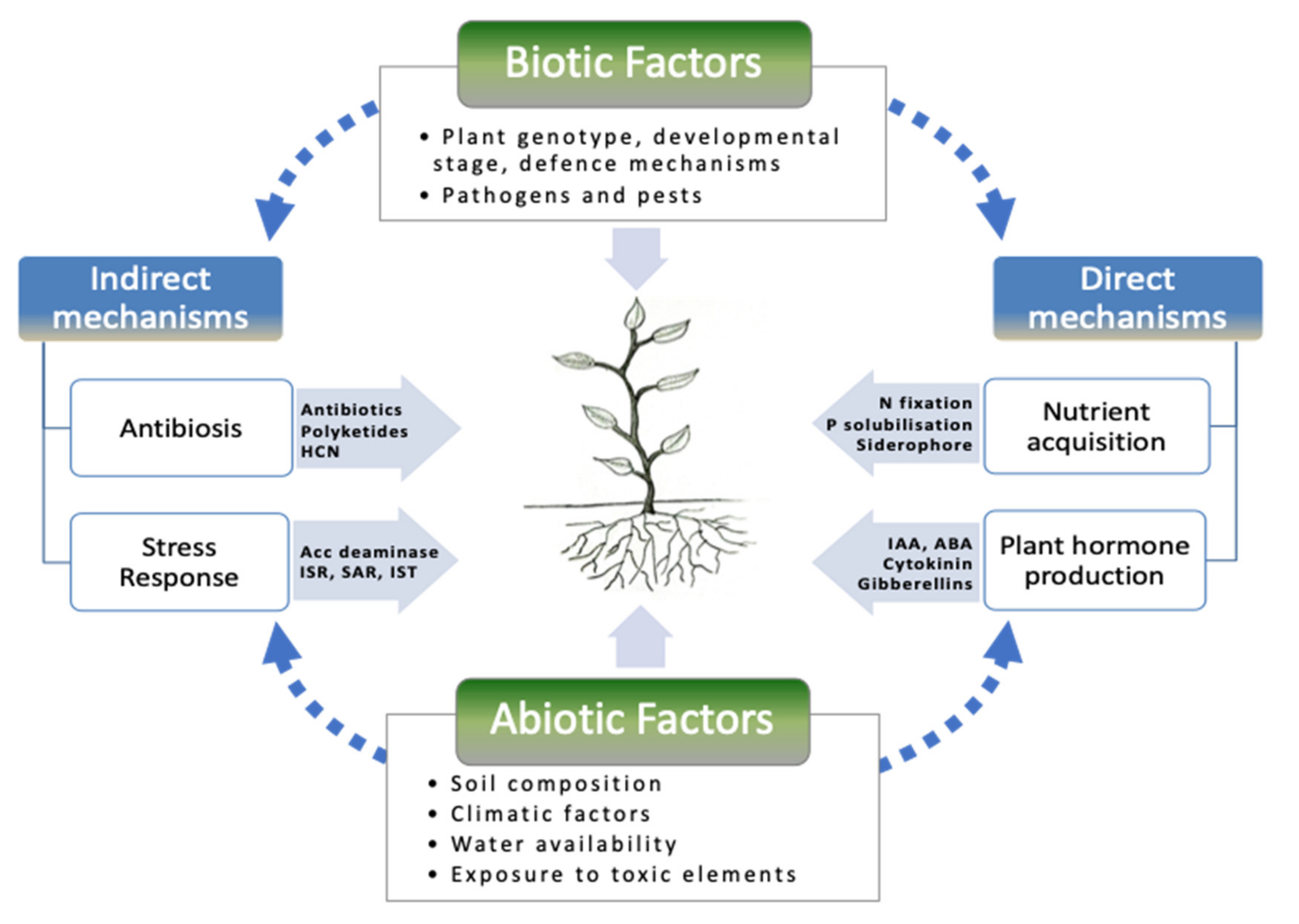
2. Growth-Promoting Mechanisms of PGPR
2.1. Nitrogen Fixation
N2 + 8H+ + 8e− + 16 ATP → 2NH3 + H2 + 16ADP + 16 Pi
2.2. P Solubilization
2.3. Production of Plant Growth Regulators (Hormones)
3. Influence of PGPR Strains on Plant Growth Promotion and Nutrient Uptake
4. Molecular Techniques Used in PGPR Study
5. Conclusions and Prospects
Author Contributions
Funding
Conflicts of Interest
References
- Kloepper, J.W. Plant Growth-Promoting Rhizobacteria on Radishes. In Proceedings of the 4th International Conference on Plant Pathogenic Bacter, Station de Pathologie Vegetale et Phytobacteriologie, INRA, Angers, France, 27 August–2 September 1978; Volume 2, pp. 879–882. [Google Scholar]
- Dobereiner, J.; Day, J.M. Associative Symbioses in Tropical Grasses: Characterization of Microorganisms and Dinitrogen-Fixing Sites. In Proceedings of the 1st International Symposium on Nitrogen Fixation; Washington State University Press: Pullman, WA, USA, 1976; Volume 2, pp. 518–538. [Google Scholar]
- Raaijmakers, J.M.; Paulitz, T.C.; Steinberg, C.; Alabouvette, C.; Moënne-Loccoz, Y. The rhizosphere: A playground and battlefield for soilborne pathogens and beneficial microorganisms. Plant Soil 2009, 321, 341–361. [Google Scholar] [CrossRef]
- Gopalakrishnan, S.; Sathya, A.; Vijayabharathi, R.; Varshney, R.K.; Gowda, C.L.L.; Krishnamurthy, L. Plant growth promoting rhizobia: Challenges and opportunities. 3 Biotech 2015, 5, 355–377. [Google Scholar] [CrossRef] [PubMed]
- Compant, S.; Duffy, B.; Nowak, J.; Clément, C.; Barka, E.A. Use of plant growth-promoting bacteria for biocontrol of plant diseases: Principles, mechanisms of action, and future prospects. Appl. Environ. Microbiol. 2005, 71, 4951–4959. [Google Scholar] [CrossRef] [PubMed]
- Beneduzi, A.; Ambrosini, A.; Passaglia, L.M.P. Plant growth-promoting rhizobacteria (PGPR): Their potential as antagonists and biocontrol agents. Genet. Mol. Biol. 2012, 35, 1044–1051. [Google Scholar] [CrossRef] [PubMed]
- Ali, S.; Hameed, S.; Shahid, M.; Iqbal, M.; Lazarovits, G.; Imran, A. Functional characterization of potential PGPR exhibiting broad-spectrum antifungal activity. Microbiol. Res. 2020, 232, 126389. [Google Scholar] [CrossRef] [PubMed]
- Bais, H.P.; Weir, T.L.; Perry, L.G.; Gilroy, S.; Vivanco, J.M. The role of root exudates in rhizosphere interactions with plants and other organisms. Annu. Rev. Plant Biol. 2006, 57, 233–266. [Google Scholar] [CrossRef]
- Sasse, J.; Martinoia, E.; Northen, T. Feed your friends: Do plant exudates shape the root microbiome? Trends Plant Sci. 2018, 23, 25–41. [Google Scholar] [CrossRef]
- Jones, D.L.; Nguyen, C.; Finlay, R.D. Carbon flow in the rhizosphere: Carbon trading at the soil-root interface. Plant Soil 2009, 321, 5–33. [Google Scholar] [CrossRef]
- Badri, D.V.; Vivanco, J.M. Regulation and function of root exudates. Plant Cell Environ. 2009, 32, 666–681. [Google Scholar] [CrossRef]
- Steinkellner, S.; Lendzemo, V.; Langer, I.; Schweiger, P.; Khaosaad, T.; Toussaint, J.-P.; Vierheilig, H. Flavonoids and strigolactones in root exudates as signals in symbiotic and pathogenic plant-fungus interactions. Molecules 2007, 12, 1290–1306. [Google Scholar] [CrossRef]
- Gamalero, E.; Glick, B.R. Mechanisms used by plant growth-promoting bacteria. In Bacteria in Agrobiology: Plant Nutrient Management; Maheshwari, D., Ed.; Springer: Berlin/Heidelberg, Germany, 2011; pp. 17–46. [Google Scholar] [CrossRef]
- Malik, K.; Bilal, R.; Mehnaz, S.; Rasyl, G.; Mirza, M.S.; Ali, A. Association of nitrogen-fixing, plant-growth-promoting rhizobacteria (PGPR) with kallar grass and rice. Plant Soil 1997, 194, 37–44. [Google Scholar] [CrossRef]
- Alori, E.T.; Glick, B.R.; Babalola, O.O. Microbial phosphorus solubilization and its potential for use in sustainable agriculture. Front. Microbiol. 2017, 8, 971. [Google Scholar] [CrossRef] [PubMed]
- Egamberdieva, D.; Wirth, S.J.; Alqarawi, A.A.; Abd-Allah, E.F.; Hashem, A. Phytohormones and beneficial microbes: Essential components for plants to balance stress and fitness. Front. Microbiol. 2017, 8, 2104. [Google Scholar] [CrossRef]
- Korir, H.; Mungai, N.W.; Thuita, M.; Hamba, Y.; Masso, C. Co-inoculation effect of rhizobia and plant growth promoting rhizobacteria on common bean growth in a low phosphorus soil. Front. Plant Sci. 2017, 8, 141. [Google Scholar] [CrossRef]
- Ngumbi, E.; Kloepper, J. Bacterial-mediated drought tolerance: Current and future prospects. Appl. Soil Ecol. 2016, 105, 109–125. [Google Scholar] [CrossRef]
- El-Esawi, M.A.; Alaraidh, I.A.; Alsahli, A.A.; Alamri, S.A.; Ali, H.M.; Alayafi, A.A. Bacillus firmus (SW5) augments salt tolerance in soybean (Glycine max L.) by modulating root system architecture, antioxidant defense systems and stress-responsive genes expression. Plant Physiol. Biochem. 2018, 132, 375–384. [Google Scholar] [CrossRef]
- Ryu, C.M.; Farag, M.A.; Hu, C.H.; Reddy, M.S.; Wei, H.X.; Paré, P.W.; Kloepper, J.W. Bacterial volatiles promote growth in Arabidopsis. Proc. Natl. Acad. Sci. USA 2003, 100, 4927–4932. [Google Scholar] [CrossRef] [PubMed]
- Sharma, M.; Mishra, V.; Rau, N.; Sharma, R.S. Increased iron-stress resilience of maize through inoculation of siderophore-producing Arthrobacter globiformis from mine. J. Basic Microbiol. 2016, 56, 719–735. [Google Scholar] [CrossRef]
- Kloepper, J.W.; Ryu, C.M. Bacterial endophytes as elicitors of induced systemic resistance. In Microbial Root Endophytes; Springer: Berlin/Heidelberg, Germany, 2006; pp. 33–52. [Google Scholar]
- Valverde, G.; Otabbong, E. Evaluation of N2-fixation measured by the 15N-dilution and N-difference methods in Nicaraguan and Ecuadorian Phaseolus vulgaris L. plants inoculated with Rhizobium leguminosarum biovar. Acta Agric. Scand. 1997, 47, 71–80. [Google Scholar]
- Akhtar, M.S.; Siddiqui, Z.A. Glomus intraradices, Pseudomonas alcaligenes, and Bacillus pumilus: Effective agents for the control of root-rot disease complex of chickpea (Cicer arietinum L.). J. Gen. Plant Pathol. 2008, 74, 53–60. [Google Scholar] [CrossRef]
- Verma, J.P.; Yadav, J.; Tiwari, K.N. Enhancement of nodulation and yield of chickpea by co-inoculation of indigenous Mesorhizobium spp. and plant growth–promoting rhizobacteria in eastern Uttar Pradesh. Commun. Soil Sci. Plant Anal. 2012, 43, 605–621. [Google Scholar] [CrossRef]
- Hamaoui, B.; Abbadi, J.; Burdman, S.; Rashid, A.; Sarig, S.; Okon, Y. Effects of inoculation with Azospirillum brasilense on chickpeas (Cicer arietinum) and faba beans (Vicia faba) under different growth conditions. Agronomy 2001, 21, 553–560. [Google Scholar] [CrossRef]
- Abdiev, A.; Khaitov, B.; Toderich, K.; Park, K.W. Growth, nutrient uptake and yield parameters of chickpea (Cicer arietinum L.) enhance by Rhizobium and Azotobacter inoculations in saline soil. J. Plant Nutr. 2019, 42, 2703–2714. [Google Scholar] [CrossRef]
- Kumar, R.; Chandra, R. Influence of PGPR and PSB on Rhizobium leguminosarum bv. viciae strain competition and symbiotic performance in lentil. World J. Agric. Sci. 2008, 4, 297–301. [Google Scholar]
- Saini, P.; Khanna, V. Evaluation of native rhizobacteria as promoters of plant growth for increased yield in lentil (Lens culinaris). Recent Res. Sci. Technol. 2012, 4, 5–9. [Google Scholar]
- Mishra, P.K.; Bisht, S.C.; Ruwari, P.; Joshi, G.K.; Singh, G.; Bisht, J.K.; Bhatt, J.C. Bioassociative effect of cold tolerant Pseudomonas spp. and Rhizobium leguminosarum-PR1 on iron acquisition, nutrient uptake and growth of lentil (Lens culinaris L.). Eur. J. Soil Biol. 2011, 47, 35–43. [Google Scholar] [CrossRef]
- Tsigie, A.; Tilak, K.V.B.R.; Saxena, A.K. Field response of legumes to inoculation with plant growth-promoting rhizobacteria. Biol. Fertil. Soils 2011, 47, 971–974. [Google Scholar] [CrossRef]
- Zaidi, A.; Khan, M.S. Co-inoculation effects of phosphate solubilizing microorganisms and Glomus fasciculatum on green gram-Bradyrhizobium symbiosis. Turk. J. Agric. For. 2006, 30, 223–230. [Google Scholar]
- Qureshi, M.A.; Shakir, M.A.; Iqbal, A.; Akhtar, N.; Khan, A. Co-inoculation of phosphate solubilizing bacteria and rhizobia for improving growth and yield of mungbean (Vigna radiata L.). J. Anim. Plant Sci. 2011, 21, 491–497. [Google Scholar]
- Sharma, A.; Johri, B.N. Combat of iron-deprivation through a plant growth promoting fluorescent Pseudomonas strain GRP3A in mung bean (Vigna radiata L. Wilzeck). Microbiol. Res. 2003, 158, 77–81. [Google Scholar] [CrossRef]
- Noreen, R.; Ali, S.A.; Hasan, K.A.; Sultana, V.; Ara, J.; Ehteshamul-Haque, S. Evaluation of biocontrol potential of fluorescent Pseudomonas associated with root nodules of mungbean. Crop Prot. 2015, 75, 18–24. [Google Scholar] [CrossRef]
- Desai, G.P.K.S.; Pinisetty, S. Impact of seed bacterization with PGPR on growth and nutrient uptake in different cultivable varieties of green gram. Asian J. Agric. Res. 2015, 9, 113–122. [Google Scholar] [CrossRef]
- Harish, S.; Manjula, K.; Podile, A.R. Fusarium udum is resistant to the mycolytic activity of a biocontrol strain of Bacillus subtilis AF1. FEMS Microbiol. Ecol. 1998, 25, 385–390. [Google Scholar] [CrossRef]
- Dutta, S.; Morang, P.; Kumar, S.N.; Kumar, B.S.D. Fusarial wilt control and growth promotion of pigeon pea through bioactive metabolites produced by two plant growth promoting rhizobacteria. World J. Microbiol. Biotechnol. 2014, 30, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Kumar, G.P.; Desai, S.; Reddy, G.; Amalraj, E.L.D.; Rasul, A.; Ahmed, S.K.M.H. Seed bacterization with fluorescent Pseudomonas spp. enhances nutrient uptake and growth of Cajanus cajan L. Commun. Soil Sci. Plant Anal. 2015, 46, 652–655. [Google Scholar] [CrossRef]
- Sharma, R.; Paliwal, J.S.; Chopra, P.; Dogra, D.; Pooniya, V.; Bisaria, V.S.; Swarnalakshmi, K.; Sharma, S. Survival, efficacy and rhizospheric effects of bacterial inoculants on Cajanus cajan. Agric. Ecosyst. Environ. 2017, 240, 244–252. [Google Scholar] [CrossRef]
- Dey, R.; Pal, K.K.; Bhatt, D.M.; Chauhan, S.M. Growth promotion and yield enhancement of peanut (Arachis hypogaea L.) by application of plant growth-promoting rhizobacteria. Microbiol. Res. 2004, 159, 371–394. [Google Scholar] [CrossRef] [PubMed]
- Yuttavanichakul, W.; Lawingsa, P.; Wongkaew, S.; Teaumroong, N.; Boonkerd, N.; Nomura, N.; Tittabutr, P. Improvement of peanut rhizobial inoculant by incorporation of plant growth promoting rhizobacteria (PGPR) as biocontrol against the seed borne fungus, Aspergillus niger. Biol. Control 2012, 63, 87–97. [Google Scholar] [CrossRef]
- Masciarelli, O.; Llanes, A.; Luna, V. A new PGPR co-inoculated with Bradyrhizobium japonicum enhances soybean nodulation. Microbiol. Res. 2014, 169, 609–615. [Google Scholar] [CrossRef]
- Molla, A.H.; Shamsuddin, Z.H.; Halimi, M.S.; Morziah, M.; Puteh, A.B. Potential for enhancement of root growth and nodulation of soybean co-inoculated with Azospirillum and Bradyrhizobium in laboratory systems. Soil Biol. Biochem. 2001, 33, 457–463. [Google Scholar] [CrossRef]
- Argaw, A. Evaluation of Co-inoculation of Bradyrhizobium japonicum and phosphate solubilizing Pseudomonas spp. effect on soybean (Glycine max L. (Merr.)) in Assossa Area. J. Agric. Sci. Technol. 2012, 14, 213–224. [Google Scholar]
- Cattelan, A.J.; Hartel, P.G.; Fuhrmann, J.J. Screening for plant growth-promoting rhizobacteria to promote early soybean growth. Soil Sci. Soc. Am. J. 1999, 63, 1670–1680. [Google Scholar] [CrossRef]
- Elkoca, E.; Turan, M.; Dinmez, M.F. Effects of single, dual and triple inoculations with Bacillus subtilis, Bacillus megaterium and Rhizobium leguminosarum bv. Phaseoli on nodulation, nutrient uptake, yield and yield parameters of common bean (Phaseolus vulgaris L. Cv. ’Elkoca-05’). J. Plant Nutr. 2010, 33, 2104–2119. [Google Scholar] [CrossRef]
- Figueiredo, M.V.B.; Martinez, C.R.; Burity, H.A.; Chanway, C.P. Plant growth-promoting rhizobacteria for improving nodulation and nitrogen fixation in the common bean (Phaseolus vulgaris L.). World J. Microbiol. Biotechnol. 2008, 24, 1187–1193. [Google Scholar] [CrossRef]
- Yadegari, M.; Rahmani, H.A.; Noormohammadi, G.; Ayneband, A. Evaluation of bean (Phaseolus vulgaris) seeds inoculation with Rhizobium phaseoli and plant growth promoting rhizobacteria on yield and yield components. Pak. J. Biol. Sci. 2008, 11, 1935–1939. [Google Scholar] [CrossRef]
- Burdman, J.; Kigel, Y.O. Effects of Azospirillum brasilense on nodulation and growth of common bean (Phaseolus vulgaris L.). Soil Biol. Biochem. 1997, 29, 923–929. [Google Scholar] [CrossRef]
- Sánchez, A.C.; Gutiérrez, R.T.; Santana, R.C.; Urrutia, A.R.; Fauvart, M.; Michiels, J.; Vanderleyden, J. Effects of co-inoculation of native Rhizobium and Pseudomonas strains on growth parameters and yield of two contrasting Phaseolus vulgaris L. genotypes under Cuban soil conditions. Eur. J. Soil Biol. 2014, 62, 105–112. [Google Scholar] [CrossRef]
- Samy, A.; El-Azeem, A.; Mehana, T.A.; Shabayek, A. Response of faba bean (Vicia faba L.) to inoculation with plant growth-promoting rhizobacteria. Catrina. Int. J. Environ. Sci. 2007, 2, 67–75. [Google Scholar]
- Rodelas, B.; González-López, J.; Pozo, C.; Salmerón, V.; Martínez-Toledo, M.V. Response of faba bean (Vicia faba L.) to combined inoculation with Azotobacter and Rhizobium leguminosarum bv. viceae. Appl. Soil Ecol. 1999, 12, 51–59. [Google Scholar] [CrossRef]
- Radwan, S.S.; Dashti, N.; El-Nemr, I.M. Enhancing the growth of Vicia faba plants by microbial inoculation to improve their phytoremediation potential for oily desert areas. Int. J. Phytoremediation 2005, 7, 19–32. [Google Scholar] [CrossRef]
- Egamberdiyeva, D.; Höflich, G. Effect of plant growth-promoting bacteria on growth and nutrient uptake of cotton and pea in a semi-arid region of Uzbekistan. J. Arid Environ. 2004, 56, 293–301. [Google Scholar] [CrossRef]
- Zahir, Z.A.; Munir, A.; Asghar, H.N.; Shaharoona, B.; Arshad, M. Effectiveness of rhizobacteria containing ACC deaminase for growth promotion of peas (Pisum sativum) under drought conditions. J. Microbiol. Biotechnol. 2008, 18, 958–963. [Google Scholar] [PubMed]
- Mishraa, P.K.; Bishta, S.C.; Jeevanandana, K.; Kumara, S.; Bisht, J.K.; Bhatt, J.C. Synergistic effect of inoculating plant growth-promoting Pseudomonas spp. and Rhizobium leguminosarum-FB1 on growth and nutrient uptake of rajmash (Phaseolus vulgaris L.). Agron. Soil Sci. 2014, 60, 799–815. [Google Scholar] [CrossRef]
- Dastager, S.G.; Deepa, C.K.; Pandey, A. Plant growth promoting potential of Pontibacter niistensis in cowpea (Vigna unguiculata (L.) Walp.). Appl. Soil Ecol. 2011, 49, 250–255. [Google Scholar] [CrossRef]
- Zhukov, V.A.; Shtark, O.Y.; Borisov, A.Y.; Tikhonovich, I.A. Breeding to improve symbiotic effectiveness of legumes. In Plant Breeding from Laboratories to Fields; Intech: Rijeka, Croatia, 2013; pp. 167–207. [Google Scholar]
- Fred, E.B.; Baldwin, I.L.; McCoy, E. Root Nodule Bacteria and Leguminous Plants; University of Wisconsin: Madison, WI, USA, 1932. [Google Scholar]
- Graham, P.H. Studies on the utilisation of carbohydrates and Krebs cycle intermediates by Rhizobia, using an agar plate method. Antonie Van Leeuwenhoek 1964, 30, 68–72. [Google Scholar] [CrossRef] [PubMed]
- Jarvis, B.D.W.; Van Berkum, P.; Chen, W.X.; Nour, S.M.; Fernandez, M.P.; Cleyet-Marel, J.C.; Gillis, M. Transfer of Rhizobium loti, Rhizobium huakuii, Rhizobium ciceri, Rhizobium mediterraneum, and Rhizobium tianshanense to Mesorhizobium gen. nov. Int. J. Syst. Evol. Microbiol. 1997, 47, 895–898. [Google Scholar] [CrossRef]
- Young, J.P.W. Phylogeny and taxonomy of rhizobia. Plant Soil 1996, 186, 45–52. [Google Scholar] [CrossRef]
- Berrada, H.; Fikri-Benbrahim, K. Taxonomy of the rhizobia: Current perspectives. Microbiol. Res. J. Int. 2014, 4, 616–639. [Google Scholar] [CrossRef]
- Zuanazzi, J.A.S.; Clergeot, P.H.; Quirion, J.-C.; Husson, H.-P.; Kondorosi, A.; Ratet, P. Production of Sinorhizobium meliloti nod gene activator and repressor flavonoids from Medicago sativa roots. Mol. Plant Microbe. Interact. 1998, 11, 784–794. [Google Scholar] [CrossRef]
- Sprent, J.I.; Ardley, J.K.; James, E.K. From North to South: A latitudinal look at legume nodulation processes. S. Afr. J. Bot. 2013, 89, 31–41. [Google Scholar] [CrossRef]
- Fischer, H.-M. Genetic regulation of nitrogen fixation in rhizobia. Microbiol. Rev. 1994, 58, 352–386. [Google Scholar] [CrossRef] [PubMed]
- Ishizuka, J. Trends in biological nitrogen fixation research and application. In Biological Nitrogen Fixation for Sustainable Agriculture; Ladha, J.K., George, T., Bohlool, B.B., Eds.; Developments in Plant and Soil Sciences; Springer: Dordrecht, The Netherlands, 1992; Volume 49, pp. 197–209. [Google Scholar] [CrossRef]
- Brockwell, J.; Bottomley, P.J.; Thies, J.E. Manipulation of rhizobia microflora for improving legume productivity and soil fertility: A critical assessment. Plant Soil 1995, 174, 143–180. [Google Scholar] [CrossRef]
- Kahindi, J.H.P.; Woomer, P.; George, T.; de Souza Moreira, F.M.; Karanja, N.K.; Giller, K.E. Agricultural intensification, soil biodiversity and ecosystem function in the tropics: The role of nitrogen-fixing bacteria. Appl. Soil Ecol. 1997, 6, 55–76. [Google Scholar] [CrossRef]
- Peoples, M.B.; Herridge, D.F.; Ladha, J.K. Biological nitrogen fixation: An efficient source of nitrogen for sustainable agricultural production? In Management of Biological Nitrogen Fixation for the Development of More Productive and Sustainable Agricultural Systems; Springer: Dordrecht, The Netherlands, 1995; pp. 3–28. [Google Scholar]
- Muresu, R.; Polone, E.; Sulas, L.; Baldan, B.; Tondello, A.; Delogu, G.; Cappuccinelli, P.; Alberghini, S.; Benhizia, Y.; Benhizia, H. Coexistence of predominantly nonculturable rhizobia with diverse, endophytic bacterial taxa within nodules of wild legumes. FEMS Microbiol. Ecol. 2008, 63, 383–400. [Google Scholar] [CrossRef] [PubMed]
- Zhao, K.; Tung, C.-W.; Eizenga, G.C.; Wright, M.H.; Ali, M.L.; Price, A.H.; Norton, G.J.; Islam, M.R.; Reynolds, A.; Mezey, J.; et al. Genome-wide association mapping reveals a rich genetic architecture of complex traits in Oryza sativa. Nat. Commun. 2011, 2, 1–10. [Google Scholar] [CrossRef]
- Dudeja, S.S.; Giri, R.; Saini, R.; Suneja-Madan, P.; Kothe, E. Interaction of endophytic microbes with legumes. J. Basic Microbiol. 2012, 52, 248–260. [Google Scholar] [CrossRef]
- Carranca, C.; De Varennes, A.; Rolston, D. Biological nitrogen fixation by fababean, pea and chickpea under field conditions estimated by the 15N isotope dilution technique. Eur. J. Agron. 1999, 10, 49–56. [Google Scholar] [CrossRef]
- Horn, C.P.; Dalal, R.C.; Birch, C.J.; Doughton, J.A. Nitrogen fixation in chickpea as affected by planting time and tillage practice. In Proceedings of the 8th Australian Agronomy Conference, Queensland, Australia, 29 January–2 February 1996; pp. 1–5. [Google Scholar]
- Rao, J.V.D.K.K.; Dart, P.J. Nodulation, nitrogen fixation and nitrogen uptake in pigeonpea (Cajanus cajan (L.) Millsp) of different maturity groups. Plant Soil 1987, 99, 255–266. [Google Scholar] [CrossRef]
- Delić, D.; Stajković-Srbinović, O.; Kuzmanović, D.; Rasulić, N.; Mrvić, V.; Andjelović, S.; Knežević-Vukčević, J. Effect of bradyrhizobial inoculation on growth and seed yield of mungbean in Fluvisol and Humofluvisol. Afr. J. Microbiol. Res. 2011, 5, 3946–3957. [Google Scholar]
- Kelstrup, L.; Rowarth, J.S.; Williams, P.H.; Ronson, C. Nitrogen fixation in peas (Pisum sativum L.), lupins (Lupinus angustifolius L.) and lentils (Lens culinaris Medik.). Proc. Agron. Soc. N. Z. 1996, 26, 71–74. [Google Scholar]
- Bremer, E.; Van Kessel, C.; Nelson, L.; Rennie, R.J.; Rennie, D.A. Selection of Rhizobium leguminosarum strains for lentil (Lens culinaris) under growth room and field conditions. Plant Soil 1990, 121, 47–56. [Google Scholar] [CrossRef]
- Usukh, B. The Impact of Lentil and Field Pea Seeding Rates on Dinitrogen Fixation and Subsequent Nitrogen Benefits in an Organic Cropping System. Ph.D. Thesis, University of Saskatchewan, Saskatoon, SK, Canada, 2010. [Google Scholar]
- Sparrowl, S.D.; Cochran, L.; Spanow, E.B. Herbage yield and nitrogen accumulation by seven legume crops on acid and neutral soils in a subarctic environment. Can. J. Plant Sci. 1993, 73, 1037–1045. [Google Scholar] [CrossRef]
- Valverde, A.; Burgos, A.; Fiscella, T.; Rivas, R.; Velazquez, E.; Rodríguez-Barrueco, C.; Cervantes, E.; Chamber, M.; Igual, J.-M. Differential effects of coinoculations with Pseudomonas jessenii ps06 (a phosphate-solubilizing bacterium) and Mesorhizobium ciceri C-2/2 strains on the growth and seed yield of chickpea under greenhouse and field conditions. In Proceedings of the First International Meeting on Microbial Phosphate Solubilization; Springer: Dordrecht, The Netherlands, 2007; pp. 43–50. [Google Scholar]
- Hardarson, G.; Bliss, F.A.; Cigales-Rivero, M.R.; Henson, R.A.; Kipe-Nolt, J.A.; Longeri, L.; Manrique, A.; Pena-Cabriales, J.J.; Pereira, P.A.A.; Sanabria, C.A.; et al. Genotypic variation in biological nitrogen fixation by common bean. Plant Soil 1993, 152, 59–70. [Google Scholar] [CrossRef]
- Smith, D.L.; Hume, D.J. Comparison of assay methods for N2 fixation utilizing white bean and soybean. Can. J. Plant Sci. 1987, 67, 11–19. [Google Scholar] [CrossRef]
- Moawad, H.; El-din, S.M.S.B.; Khalafallah, M. Quantification of nitrogen fixation by the peanutl Rhizobium symbiotic system in a virgin sandy Soil. J. Plant Nutr. Soil Sci. 1986, 149, 668–673. [Google Scholar] [CrossRef]
- Peoples, M.B.; Bell, M.J.; Bushby, H.V.A. Effect of rotation and inoculation with Bradyrhizobium on nitrogen fixation and yield of peanut (Arachis hypogaea L., cv. Virginia Bunch). Aust. J. Agric. Res. 1992, 43, 595–607. [Google Scholar] [CrossRef]
- Toomsan, B.; McDonagh, J.F.; Limpinuntana, V.; Giller, K.E. Nitrogen fixation by groundnut and soyabean and residual nitrogen benefits to rice in farmers’ fields in Northeast Thailand. Plant Soil 1995, 175, 45–56. [Google Scholar] [CrossRef]
- Olivera, M.; Tejera, N.; Iribarne, C.; Ocana, A.; Lluch, C. Growth, nitrogen fixation and ammonium assimilation in common bean (Phaseolus vulgaris): Effect of phosphorus. Physiol. Plant. 2004, 121, 498–505. [Google Scholar] [CrossRef]
- Israel, D.W. Investigation of the role of phosphorus in symbiotic dinitrogen fixation. Plant Physiol. 1987, 84, 835–840. [Google Scholar] [CrossRef]
- Rudresh, D.L.; Shivaprakash, M.K.; Prasad, R.D. Effect of combined application of Rhizobium, phosphate solubilizing bacterium and Trichoderma spp. on growth, nutrient uptake and yield of chickpea (Cicer aritenium L.). Appl. Soil Ecol. 2005, 28, 139–146. [Google Scholar] [CrossRef]
- Swarnalakshmi, K.; Pooniya, V.; Paul, S. Synergistic interaction of Piriformospora indica and microbial inoculants on symbiotic potential, plant nutrition and productivity of chickpea (Cicer arietinum). Indian J. Agron. 2017, 62, 481–488. [Google Scholar]
- Vassilev, N.; Someus, E.; Serrano, M.; Bravo, V.; Garcia Roman, M.; Reyes, A.; Vassileva, M. Novel Approaches in Phosphate-Fertilizer Production Based on Wastes Derived from Rock Phosphate Mining and Food Processing Industry; Nova Science Publishers: Hauppauge, NY, USA, 2009; pp. 387–391. [Google Scholar]
- Alikhani, H.A.; Saleh-Rastin, N.; Antoun, H. Phosphate solubilization activity of rhizobia native to iranian soils. In Proceedings of the First International Meeting on Microbial Phosphate Solubilization; Springer: Dordrecht, The Netherlands, 2007; pp. 35–41. [Google Scholar]
- Miller, S.H.; Browne, P.; Prigent-Combaret, C.; Combes-Meynet, E.; Morrissey, J.P.; O’Gara, F. Biochemical and genomic comparison of inorganic phosphate solubilization in Pseudomonas species. Environ. Microbiol. Rep. 2010, 2, 403–411. [Google Scholar] [CrossRef] [PubMed]
- Goldstein, A.H. Recent progress in understanding the molecular genetics and biochemistry of calcium phosphate solubilization by Gram negative bacteria. Biol. Agric. Hortic. 1995, 12, 185–193. [Google Scholar] [CrossRef]
- Yazdani, M.; Bahmanyar, M.A.; Pirdashti, H.; Esmaili, M.A. Effect of phosphate solubilization microorganisms (PSM) and plant growth promoting rhizobacteria (PGPR) on yield and yield components of corn (Zea mays L.). World Acad. Sci. Eng. Technol. 2009, 49, 90–92. [Google Scholar]
- Gyaneshwar, P.; Kumar, G.N.; Parekh, L.J.; Poole, P.S. Role of soil microorganisms in improving P nutrition of plants. Plant Soil 2002, 245, 83–93. [Google Scholar] [CrossRef]
- Feng, G.; Song, Y.C.; Li, X.L.; Christie, P. Contribution of arbuscular mycorrhizal fungi to utilization of organic sources of phosphorus by red clover in a calcareous soil. Appl. Soil Ecol. 2003, 22, 139–148. [Google Scholar] [CrossRef]
- Verma, J.P.; Yadav, J.; Tiwari, K.N.; Kumar, A. Effect of indigenous Mesorhizobium spp. and plant growth promoting rhizobacteria on yields and nutrients uptake of chickpea (Cicer arietinum L.) under sustainable agriculture. Ecol. Eng. 2013, 51, 282–286. [Google Scholar] [CrossRef]
- Moubayidin, L.; Di Mambro, R.; Sabatini, S. Cytokinin-auxin crosstalk. Trends Plant Sci. 2009, 14, 557–562. [Google Scholar] [CrossRef]
- Overvoorde, P.; Fukaki, H.; Beeckman, T. Auxin control of root development. Cold Spring. Harb. Perspect. Biol. 2010, 2, a001537. [Google Scholar] [CrossRef]
- Yaxley, J.R.; Ross, J.J.; Sherriff, L.J.; Reid, J.B. Gibberellin biosynthesis mutations and root development in pea. Plant Physiol. 2001, 125, 627–633. [Google Scholar] [CrossRef]
- Arshad, M.; Frankenberger, W.T., Jr. Plant growth-regulating substances in the rhizosphere: Microbial production and functions. Adv. Agron. 1997, 62, 45–151. [Google Scholar] [CrossRef]
- Hirsch, A.M.; Fang, Y. Plant hormones and nodulation: What’s the connection? Plant Mol. Biol. 1994, 26, 5–9. [Google Scholar] [CrossRef]
- Teale, W.D.; Paponov, I.A.; Palme, K. Auxin in action: Signalling, transport and the control of plant growth and development. Nat. Rev. Mol. Cell Biol. 2006, 7, 847–859. [Google Scholar] [CrossRef] [PubMed]
- Breakspear, A.; Liu, C.; Roy, S.; Stacey, N.; Rogers, C.; Trick, M.; Morieri, G.; Mysore, K.S.; Wen, J.; Oldroyd, G.E.D. The root hair “infectome” of Medicago truncatula uncovers changes in cell cycle genes and reveals a requirement for auxin signaling in rhizobial infection. Plant Cell 2014, 26, 4680–4701. [Google Scholar] [CrossRef]
- de Billy, F.; Grosjean, C.; May, S.; Bennett, M.; Cullimore, J.V. Expression studies on AUX1-like genes in Medicago truncatula suggest that auxin is required at two steps in early nodule development. Mol. Plant Microbe. Interact. 2001, 14, 267–277. [Google Scholar] [CrossRef]
- Laplaze, L.; Lucas, M.; Champion, A. Rhizobial root hair infection requires auxin signaling. Trends Plant Sci. 2015, 20, 332–334. [Google Scholar] [CrossRef]
- Prinsen, E.; Chauvaux, N.; Schmidt, J.; John, M.; Wieneke, U.; De Greef, J.; Schell, J.; Van Onckelen, H. Stimulation of indole-3-acetic acid production in Rhizobium by flavonoids. FEBS Lett. 1991, 282, 53–55. [Google Scholar] [CrossRef]
- Spaepen, S.; Vanderleyden, J.; Remans, R. Indole-3-acetic acid in microbial and microorganism-plant signaling. FEMS Microbiol. Rev. 2007, 31, 425–448. [Google Scholar] [CrossRef]
- Persello-Cartieaux, F.; David, P.; Sarrobert, C.; Thibaud, M.-C.; Achouak, W.; Robaglia, C.; Nussaume, L. Utilization of mutants to analyze the interaction between Arabidopsis thaliana and its naturally root-associated Pseudomonas. Planta 2001, 212, 190–198. [Google Scholar] [CrossRef]
- Bashan, Y.; Holguin, G.; De-Bashan, L.E. Azospirillum-plant relationships: Physiological, molecular, agricultural, and environmental advances (1997–2003). Can. J. Microbiol. 2004, 50, 521–577. [Google Scholar] [CrossRef]
- Pii, Y.; Crimi, M.; Cremonese, G.; Spena, A.; Pandolfini, T. Auxin and nitric oxide control indeterminate nodule formation. BMC Plant Biol. 2007, 7, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Camerini, S.; Senatore, B.; Lonardo, E.; Imperlini, E.; Bianco, C.; Moschetti, G.; Rotino, G.L.; Campion, B.; Defez, R. Introduction of a novel pathway for IAA biosynthesis to rhizobia alters vetch root nodule development. Arch. Microbiol. 2008, 190, 67–77. [Google Scholar] [CrossRef]
- Fukuhara, H.; Minakawa, Y.; Akao, S.; Minamisawa, K. The involvement of indole-3-acetic acid produced by Bradyrhizobium elkanii in nodule formation. Plant Cell Physiol. 1994, 35, 1261–1265. [Google Scholar] [CrossRef]
- Manulis, S.; Haviv-Chesner, A.; Brandl, M.T.; Lindow, S.E.; Barash, I. Differential involvement of indole-3-acetic acid biosynthetic pathways in pathogenicity and epiphytic fitness of Erwinia herbicola pv. gypsophilae. Mol. Plant Microbe. Interact. 1998, 11, 634–642. [Google Scholar] [CrossRef] [PubMed]
- Nieto, K.F.; Frankenberger, W.T., Jr. Biosynthesis of cytokinins by Azotobacter chroococcum. Soil Biol. Biochem. 1989, 21, 967–972. [Google Scholar] [CrossRef]
- Sturtevant, D.B.; Taller, B.J. Cytokinin production by Bradyrhizobium japonicum. Plant Physiol. 1989, 89, 1247–1252. [Google Scholar] [CrossRef] [PubMed]
- Conrad, K.; Bettin, B.; Neumann, S. The Cytokinin production of Azospirillum and Klebsiella possible ecological effects. In Physiology and Biochemistry of Cytokinins in Plants; Academic Publishing: The Hague, The Netherlands, 1992; Available online: https://www.researchgate.net/publication/256111462_The_cytokinin_production_of_Azospirillum_and_Klebsiella_and_its_possible_ecological_effects_In_Physiology_and_Biochemistry_of_Cytokinins_in_Plants_ed_by_M_Kaminek_et_al_1992_pp_401-405 (accessed on 30 May 2014).
- Timmusk, S.; Nicander, B.; Granhall, U.; Tillberg, E. Cytokinin production by Paenibacillus polymyxa. Soil Biol. Biochem. 1999, 31, 1847–1852. [Google Scholar] [CrossRef]
- García de Salamone, I.E.; Hynes, R.K.; Nelson, L.M. Cytokinin production by plant growth promoting rhizobacteria and selected mutants. Can. J. Microbiol. 2001, 47, 404–411. [Google Scholar] [CrossRef]
- Arkhipova, T.N.; Veselov, S.U.; Melentiev, A.I.; Martynenko, E.V.; Kudoyarova, G.R. Ability of bacterium Bacillus subtilis to produce cytokinins and to influence the growth and endogenous hormone content of lettuce plants. Plant Soil 2005, 272, 201–209. [Google Scholar] [CrossRef]
- Aloni, R.; Aloni, E.; Langhans, M.; Ullrich, C.I. Role of cytokinin and auxin in shaping root architecture: Regulating vascular differentiation, lateral root initiation, root apical dominance and root gravitropism. Ann. Bot. 2006, 97, 883–893. [Google Scholar] [CrossRef]
- Noel, T.C.; Sheng, C.; Yost, C.K.; Pharis, R.P.; Hynes, M.F. Rhizobium leguminosarum as a plant growth-promoting rhizobacterium: Direct growth promotion of canola and lettuce. Can. J. Microbiol. 1996, 42, 279–283. [Google Scholar] [CrossRef]
- Frugier, F.; Kosuta, S.; Murray, J.D.; Crespi, M.; Szczyglowski, K. Cytokinin: Secret agent of symbiosis. Trends Plant Sci. 2008, 13, 115–120. [Google Scholar] [CrossRef]
- Cooper, J.B.; Long, S.R. Morphogenetic rescue of Rhizobium meliloti nodulation mutants by trans-zeatin secretion. Plant Cell 1994, 6, 215–225. [Google Scholar] [CrossRef]
- Kisiala, A.; Laffont, C.; Emery, R.J.N.; Frugier, F. Bioactive cytokinins are selectively secreted by Sinorhizobium meliloti nodulating and nonnodulating strains. Mol. Plant Microbe Interact. 2013, 26, 1225–1231. [Google Scholar] [CrossRef]
- Mens, C.; Li, D.; Haaima, L.E.; Gresshoff, P.M.; Ferguson, B.J. Local and systemic effect of cytokinins on soybean nodulation and regulation of their isopentenyl transferase (IPT) biosynthesis genes following rhizobia inoculation. Front. Plant Sci. 2018, 9, 1150. [Google Scholar] [CrossRef]
- Gonzalez-Rizzo, S.; Crespi, M.; Frugier, F. The Medicago truncatula CRE1 cytokinin receptor regulates lateral root development and early symbiotic interaction with Sinorhizobium meliloti. Plant Cell 2006, 18, 2680–2693. [Google Scholar] [CrossRef]
- Murray, J.D.; Karas, B.J.; Sato, S.; Tabata, S.; Amyot, L.; Szczyglowski, K. A cytokinin perception mutant colonized by Rhizobium in the absence of nodule organogenesis. Science 2007, 315, 101–104. [Google Scholar] [CrossRef]
- Dolgikh, E.A.; Kusakin, P.G.; Kitaeva, A.B.; Tsyganova, A.V.; Kirienko, A.N.; Leppyanen, I.V.; Dolgikh, A.V.; Ilina, E.L.; Demchenko, K.N.; Tikhonovich, I.A.; et al. Mutational analysis idicates that abnormalities in rhizobial infection and subsequent plant cell and bacteroid differentiation in pea (Pisum sativum) nodules coincide with abnormal cytokinin responses and localization. Ann. Bot. 2020, 125, 905–923. [Google Scholar] [CrossRef]
- Figueiredo, M.V.B.; Burity, H.A.; Martinez, C.R.; Chanway, C.P. Alleviation of drought stress in the common bean (Phaseolus vulgaris L.) by co-inoculation with Paenibacillus polymyxa and Rhizobium tropici. Appl. Soil Ecol. 2008, 40, 182–188. [Google Scholar] [CrossRef]
- Cowan, A.K.; Cairns, A.L.; Bartels-Rahm, B. Regulation of abscisic acid metabolism: Towards a metabolic basis for abscisic acid-cytokinin antagonism. J. Exp. Bot. 1999, 50, 595–603. [Google Scholar] [CrossRef]
- Cohen, A.C.; Bottini, R.; Piccoli, P.N. Azospirillum brasilense Sp. 245 produces ABA in chemically-defined culture medium and increases ABA content in Arabidopsis plants. Plant Growth Regul. 2008, 54, 97–103. [Google Scholar] [CrossRef]
- Kang, S.M.; Radhakrishnan, R.; Khan, A.L.; Kim, M.J.; Park, J.M.; Kim, B.R.; Shin, D.H.; Lee, I.J. Gibberellin secreting rhizobacterium, Pseudomonas putida H-2-3 modulates the hormonal and stress physiology of soybean to improve the plant growth under saline and drought conditions. Plant Physiol. Biochem. 2014, 84, 115–124. [Google Scholar] [CrossRef]
- Tominaga, A.; Nagata, M.; Futsuki, K.; Abe, H.; Uchiumi, T.; Abe, M.; Kucho, K.; Hashiguchi, M.; Akashi, R.; Hirsch, A.M. Enhanced nodulation and nitrogen fixation in the abscisic acid low-sensitive mutant enhanced nitrogen fixation1 of Lotus japonicus. Plant Physiol. 2009, 151, 1965–1976. [Google Scholar] [CrossRef]
- Suzuki, A.; Akune, M.; Kogiso, M.; Imagama, Y.; Osuki, K.; Uchiumi, T.; Higashi, S.; Han, S.-Y.; Yoshida, S.; Asami, T. Control of nodule number by the phytohormone abscisic acid in the roots of two leguminous species. Plant Cell Physiol. 2004, 45, 914–922. [Google Scholar] [CrossRef]
- Van de Velde, W.; Guerra, J.C.P.; De Keyser, A.; De Rycke, R.; Rombauts, S.; Maunoury, N.; Mergaert, P.; Kondorosi, E.; Holsters, M.; Goormachtig, S. Aging in legume symbiosis. A molecular view on nodule senescence in Medicago truncatula. Plant Physiol. 2006, 141, 711–720. [Google Scholar] [CrossRef]
- Serova, T.A.; Tikhonovich, I.A.; Tsyganov, V.E. Analysis of nodule senescence in pea (Pisum sativum L.) using laser microdissection, real-time PCR, and ACC immunolocalization. J. Plant Physiol. 2017, 212, 29–44. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, S. Gibberellin metabolism and its regulation. Annu. Rev. Plant Biol. 2008, 59, 225–251. [Google Scholar] [CrossRef]
- Gutiérrez-Mañero, F.J.; Ramos-Solano, B.; Probanza, A.N.; Mehouachi, J.; Tadeo, F.R.; Talon, M. The plant-growth-promoting rhizobacteria Bacillus pumilus and Bacillus licheniformis produce high amounts of physiologically active gibberellins. Physiol. Plant. 2001, 111, 206–211. [Google Scholar] [CrossRef]
- Bottini, R.; Cassán, F.; Piccoli, P. Gibberellin production by bacteria and its involvement in plant growth promotion and yield increase. Appl. Microbiol. Biotechnol. 2004, 65, 497–503. [Google Scholar] [CrossRef] [PubMed]
- Dodd, I.C.; Zinovkina, N.Y.; Safronova, V.I.; Belimov, A.A. Rhizobacterial mediation of plant hormone status. Ann. Appl. Biol. 2010, 157, 361–379. [Google Scholar] [CrossRef]
- Salazear-Cerezo, S.; Martinez-Montiel, N.; Garcia –Sanchez, J.; Perezy-Terron, R.; Martinez-Contreras, D. Gibberellin biosynthesis and metabolism: A convergent route for plants, fungi and bacteria. Microbiol. Res. 2018, 208, 85–98. [Google Scholar] [CrossRef] [PubMed]
- Cassan, F.; Bottini, R.; Schneider, G.; Piccoli, P. Azospirillum brasilense and Azospirillum lipoferum hydrolyze conjugates of GA20 and metabolize the resultant aglycones to GA1 in seedlings of rice dwarf mutants. Plant Physiol. 2001, 125, 2053–2058. [Google Scholar] [CrossRef] [PubMed]
- Cassan, F.D.; Lucangeli, C.D.; Bottini, R.; Piccoli, P.N. Azospirillum spp. metabolize (17, 17–2H2) gibberellin A20 to (17, 17–2H2) gibberellin A1 in vivo in dy rice mutant seedlings. Plant Cell Physiol. 2001, 42, 763–767. [Google Scholar] [CrossRef]
- Freiberg, C.; Fellay, R.; Bairoch, A.; Broughton, W.J.; Rosenthal, A.; Perret, X. Molecular basis of symbiosis between Rhizobium and legumes. Nature 1997, 387, 394–401. [Google Scholar] [CrossRef] [PubMed]
- Tully, R.E.; Keister, D.L. Cloning and mutagenesis of a cytochrome P-450 locus from Bradyrhizobium japonicum that is expressed anaerobically and symbiotically. Appl. Environ. Microbiol. 1993, 59, 4136–4142. [Google Scholar] [CrossRef]
- Kaneko, T.; Nakamura, Y.; Sato, S.; Minamisawa, K.; Uchiumi, T.; Sasamoto, S.; Watanabe, A.; Idesawa, K.; Iriguchi, M.; Kawashima, K. Complete genomic sequence of nitrogen-fixing symbiotic bacterium Bradyrhizobium japonicum USDA110. DNA Res. 2002, 9, 189–197. [Google Scholar] [CrossRef]
- Tatsukami, Y.; Ueda, M. Rhizobial gibberellin negatively regulates host nodule number. Sci. Rep. 2016, 6, 27998. [Google Scholar] [CrossRef]
- Ferguson, B.J.; Ross, J.J.; Reid, J.B. Nodulation phenotypes of gibberellin and brassinosteroid mutants of pea. Plant Physiol. 2005, 138, 2396–2405. [Google Scholar] [CrossRef]
- McAdam, E.L.; Reid, J.B.; Foo, E. Gibberellins promote nodule organogenesis but inhibit the infection stages of nodulation. J. Exp. Bot. 2018, 69, 2117–2130. [Google Scholar] [CrossRef]
- Maekawa, T.; Maekawa-Yoshikawa, M.; Takeda, N.; Imaizumi-Anraku, H.; Murooka, Y.; Hayashi, M. Gibberellin controls the nodulation signaling pathway in Lotus japonicus. Plant J. 2009, 58, 183–194. [Google Scholar] [CrossRef]
- Fonouni-Farde, C.; Tan, S.; Baudin, M.; Brault, M.; Wen, J.; Mysore, K.S.; Niebel, A.; Frugier, F.; Diet, A. DELLA-mediated gibberellin signalling regulates Nod factor signalling and rhizobial infection. Nat. Commun. 2016, 7, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Jin, Y.; Liu, H.; Luo, D.; Yu, N.; Dong, W.; Wang, C.; Zhang, X.; Dai, H.; Yang, J.; Wang, E. DELLA proteins are common components of symbiotic rhizobial and mycorrhizal signalling pathways. Nat. Commun. 2016, 7, 1–14. [Google Scholar] [CrossRef] [PubMed]
- Stracke, S.; Kistner, C.; Yoshida, S.; Mulder, L.; Sato, S.; Kaneko, T.; Tabata, S.; Sandal, N.; Stougaard, J.; Szczyglowski, K. A plant receptor-like kinase required for both bacterial and fungal symbiosis. Nature 2002, 417, 959–962. [Google Scholar] [CrossRef]
- Serova, T.A.; Tsyganova, A.V.; Tikhonovich, I.A.; Tsyganov, V.E. Gibberellins inhibit nodule senescence and stimulate nodule meristem bifurcation in pea (Pisum sativum L.). Front. Plant Sci. 2019, 10, 285. [Google Scholar] [CrossRef] [PubMed]
- Guinel, F.C.; LaRue, T.A. Ethylene inhibitors partly restore nodulation to pea mutant E107 (brz). Plant Physiol. 1992, 99, 515–518. [Google Scholar] [CrossRef] [PubMed]
- Nukui, N.; Ezura, H.; Minamisawa, K. Transgenic Lotus japonicus with an ethylene receptor gene Cm-ERS1/H70A enhances formation of infection threads and nodule primordia. Plant Cell Physiol. 2004, 45, 427–435. [Google Scholar] [CrossRef]
- Shah, S.; Li, J.; Moffatt, B.A.; Glick, B.R. Isolation and characterization of ACC deaminase genes from two different plant growth-promoting rhizobacteria. Can. J. Microbiol. 1998, 44, 833–843. [Google Scholar] [CrossRef]
- Ma, W.; Guinel, F.C.; Glick, B.R. Rhizobium leguminosarum biovar viciae 1-aminocyclopropane-1-carboxylate deaminase promotes nodulation of pea plants. Appl. Environ. Microbiol. 2003, 69, 4396–4402. [Google Scholar] [CrossRef]
- Tittabutr, P.; Awaya, J.D.; Li, Q.X.; Borthakur, D. The cloned 1-aminocyclopropane-1-carboxylate (ACC) deaminase gene from Sinorhizobium sp. strain BL3 in Rhizobium sp. strain TAL1145 promotes nodulation and growth of Leucaena leucocephala. Syst. Appl. Microbiol. 2008, 31, 141–150. [Google Scholar] [CrossRef]
- Nascimento, F.X.; Brígido, C.; Glick, B.R.; Oliveira, S.; Alho, L. Mesorhizobium ciceri LMS-1 expressing an exogenous 1-aminocyclopropane-1-carboxylate (ACC) deaminase increases its nodulation abilities and chickpea plant resistance to soil constraints. Lett. Appl. Microbiol. 2012, 55, 15–21. [Google Scholar] [CrossRef]
- Uchiumi, T.; Ohwada, T.; Itakura, M.; Mitsui, H.; Nukui, N.; Dawadi, P.; Kaneko, T.; Tabata, S.; Yokoyama, T.; Tejima, K. Expression islands clustered on the symbiosis island of the Mesorhizobium loti genome. J. Bacteriol. 2004, 186, 2439–2448. [Google Scholar] [CrossRef] [PubMed]
- Nandasena, K.; Yates, R.; Tiwari, R.; O’Hara, G.; Howieson, J.; Ninawi, M.; Chertkov, O.; Detter, C.; Tapia, R.; Han, S.; et al. Complete genome sequence of Mesorhizobium ciceri bv. biserrulae type strain (WSM1271 T). Stand. Genom. Sci. 2014, 9, 462–472. [Google Scholar] [CrossRef]
- Nukui, N.; Minamisawa, K.; Ayabe, S.-I.; Aoki, T. Expression of the 1-aminocyclopropane-1-carboxylic acid deaminase gene requires symbiotic nitrogen-fixing regulator gene nifA2 in Mesorhizobium loti MAFF303099. Appl. Environ. Microbiol. 2006, 72, 4964–4969. [Google Scholar] [CrossRef] [PubMed]
- Pieterse, C.M.J.; Leon-Reyes, A.; Van der Ent, S.; Van Wees, S.C.M. Networking by small-molecule hormones in plant immunity. Nat. Chem. Biol. 2009, 5, 308–316. [Google Scholar] [CrossRef]
- Shahzad, S.M.; Khalid, A.; Arshad, M.; Tahir, J.; Mahmood, T. Improving nodulation, growth and yield of Cicer arietinum L. through bacterial ACC-deaminase induced changes in root architecture. Eur. J. Soil Biol. 2010, 46, 342–347. [Google Scholar] [CrossRef]
- Chaudhary, D.; Sindhu, S. Inducing salinity tolerance in chickpea (Cicer arietinum L.) by inoculation of 1-aminocyclopropane-1-carboxylic acid deaminase-containing Mesorhizobium strains. Afr. J. Microbiol. Res. 2015, 9, 117–124. [Google Scholar] [CrossRef]
- Sharma, P.; Khanna, V.; Kumari, S. Potential of ACC-deaminase producing plant growth promoting rhizobacteria on water stress mitigation in lentil (Lens culinaris L. Medikus) under axenic conditions. Int. J. Adv. Res. 2015, 3, 59–67. [Google Scholar]
- Shaharoona, B.; Arshad, M.; Zahir, Z.A. Effect of plant growth promoting rhizobacteria containing ACC-deaminase on maize (Zea mays L.) growth under axenic conditions and on nodulation in mung bean (Vigna radiata L.). Lett. Appl. Microbiol. 2006, 42, 155–159. [Google Scholar] [CrossRef]
- Ahmad, M.; Zahir, Z.A.; Asghar, H.N.; Asghar, M. Inducing salt tolerance in mung bean through coinoculation with rhizobia and plant-growth-promoting rhizobacteria containing 1-aminocyclopropane-1-carboxylate deaminase. Can. J. Microbiol. 2011, 57, 578–589. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Maji, D.; Chanotiya, C.S.; Kalra, A. ACC deaminase-containing Arthrobacter protophormiae induces NaCl stress tolerance through reduced ACC oxidase activity and ethylene production resulting in improved nodulation and mycorrhization in Pisum sativum. J. Plant Physiol. 2014, 171, 884–894. [Google Scholar] [CrossRef]
- Safronova, V.I.; Stepanok, V.V.; Engqvist, G.L.; Alekseyev, Y.V.; Belimov, A.A. Root-associated bacteria containing 1-aminocyclopropane-1-carboxylate deaminase improve growth and nutrient uptake by pea genotypes cultivated in cadmium supplemented soil. Biol. Fertil. Soils 2006, 42, 267–272. [Google Scholar] [CrossRef]
- Arshad, M.; Shaharoona, B.; Mahmood, T. Inoculation with Pseudomonas spp. containing ACC-deaminase partially eliminates the effects of drought stress on growth, yield, and ripening of pea (Pisum sativum L.). Pedosphere 2008, 18, 611–620. [Google Scholar] [CrossRef]
- Belimov, A.A.; Dodd, I.C.; Hontzeas, N.; Theobald, J.C.; Safronova, V.I.; Davies, W.J. Rhizosphere bacteria containing 1-aminocyclopropane-1-carboxylate deaminase increase yield of plants grown in drying soil via both local and systemic hormone signalling. New Phytol. 2009, 181, 413–423. [Google Scholar] [CrossRef] [PubMed]
- Husen, E.; Wahyudi, A.T.; Suwanto, A. Growth enhancement and disease reduction of soybean by 1-aminocyclopropane-1-carboxylate deaminase-producing Pseudomonas. Am. J. Appl. Sci. 2011, 8, 1073–1080. [Google Scholar] [CrossRef]
- Saravanakumar, D.; Samiyappan, R. ACC deaminase from Pseudomonas fluorescens mediated saline resistance in groundnut (Arachis hypogea) plants. J. Appl. Microbiol. 2007, 10, 1283–1292. [Google Scholar] [CrossRef] [PubMed]
- Khandelwal, A.; Sindhu, S.S. ACC deaminase containing rhizobacteria enhance nodulation and plant growth in cluster bean (Cyamopsis tetragonoloba L.). J. Microbiol. Res. 2013, 3, 117–123. [Google Scholar] [CrossRef]
- Trung, N.T.; Thuam, N.H. Screening of strong 1-aminocyclopropane-1-carboxylate deaminase producing bacteria for improving the salinity tolerance of cowpea. Appl. Microbiol. 2016, 2, 1000111. [Google Scholar] [CrossRef]
- Silva, E.R.; Zoz, J.; Oliveira, C.E.S.; Zuffo, A.M.; Steiner, F.; Zoz, T.; Vendruscolo, E.P. Can co-inoculation of Bradyrhizobium and Azospirillum alleviate adverse effects of drought stress on soybean (Glycine max L. Merrill.)? Arch. Microbiol. 2019, 201, 325–335. [Google Scholar] [CrossRef]
- Sondergaard, T.E.; Schulz, A.; Palmgren, M.G. Energization of transport processes in plants. Roles of the plasma membrane H+-ATPase. Plant Physiol. 2004, 136, 2475–2482. [Google Scholar] [CrossRef]
- Karlidag, H.; Esitken, A.; Turan, M.; Sahin, F. Effects of root inoculation of plant growth promoting rhizobacteria (PGPR) on yield, growth and nutrient element contents of leaves of apple. Sci. Hortic. 2007, 114, 16–20. [Google Scholar] [CrossRef]
- German, M.A.; Burdman, S.; Okon, Y.; Kigel, J. Effects of Azospirillum brasilense on root morphology of common bean (Phaseolus vulgaris L.) under different water regimes. Biol. Fertil. Soils 2000, 32, 259–264. [Google Scholar] [CrossRef]
- Saikia, S.P.; Dutta, S.P.; Goswami, A.; Bhau, B.S.; Kanjilal, P.B. Role of Azospirillum in the Improvement of Legumes. In Microbes for Legume Improvement; Khan, M.S., Musarrat, J., Zaidi, A., Eds.; Springer: Berlin/Heidelberg, Germany, 2010; pp. 389–408. [Google Scholar] [CrossRef]
- dos Santos Lima Fagotti, D.; Abrantes, J.L.F.; Cerezini, P.; Fukami, J.; Nogueira, M.A.; del Cerro, P.; Valderrama-Fernández, R.; Ollero, F.J.; Megías, M.; Hungria, M. Quorum sensing communication: Bradyrhizobium-Azospirillum interaction via N-acyl-homoserine lactones in the promotion of soybean symbiosis. J. Basic Microbiol. 2019, 59, 38–53. [Google Scholar] [CrossRef]
- Barnawal, D.; Bharti, N.; Pandey, S.S.; Pandey, A.; Chanotiya, C.S.; Kalra, A. Plant growth-promoting rhizobacteria enhance wheat salt and drought stress tolerance by altering endogenous phytohormone levels and TaCTR1/TaDREB2 expression. Physiol. Plant. 2017, 161, 502–514. [Google Scholar] [CrossRef]
- Sakamoto, K.; Ogiwara, N.; Kaji, T.; Sugimoto, Y.; Ueno, M.; Sonoda, M.; Matsui, A.; Ishida, J.; Tanaka, M.; Totoki, Y. Transcriptome analysis of soybean (Glycine max) root genes differentially expressed in rhizobial, arbuscular mycorrhizal, and dual symbiosis. J. Plant Res. 2019, 132, 541–568. [Google Scholar] [CrossRef] [PubMed]
- Sreevidya, M.; Gopalakrishnan, S.; Kudapa, H.; Varshney, R.K. Exploring plant growth-promotion actinomycetes from vermicompost and rhizosphere soil for yield enhancement in chickpea. Braz. J. Microbiol. 2016, 47, 85–95. [Google Scholar] [CrossRef]
- Tokala, R.K.; Strap, J.L.; Jung, C.M.; Crawford, D.L.; Salove, M.H.; Deobald, L.A.; Bailey, J.F.; Morra, M.J. Novel plant-microbe rhizosphere interaction involving Streptomyces lydicus WYEC108 and the pea plant (Pisum sativum). Appl. Environ. Microbiol. 2002, 68, 2161–2171. [Google Scholar] [CrossRef] [PubMed]
- Khanna, V.; Sharma, P. Potential for enhancing lentil (Lens culinaris) productivity by co-inoculation with PSB, plant growth-promoting rhizobacteria and Rhizobium. Indian J. Agric. Sci. 2011, 81, 932. [Google Scholar]
- Bansal, R.K. Synergistic effect of Rhizobium, PSB and PGPR on nodulation and grain yield of mungbean. J. Food Legum. 2009, 22, 37–39. [Google Scholar]
- Bhattacharjya, S.; Chandra, R. Effect of inoculation methods of Mesorhizobium ciceri and PGPR in chickpea (Cicer areietinum L.) on symbiotic traits, yields, nutrient uptake and soil properties. Legum. Res. Int. J. 2013, 36, 331–337. [Google Scholar]
- Badawi, F.S.F.; Biomy, A.M.M.; Desoky, A.H. Peanut plant growth and yield as influenced by co-inoculation with Bradyrhizobium and some rhizo-microorganisms under sandy loam soil conditions. Ann. Agric. Sci. 2011, 56, 17–25. [Google Scholar] [CrossRef]
- Tilak, K.V.B.R.; Ranganayaki, N.; Manoharachari, C. Synergistic effects of plant-growth promoting rhizobacteria and Rhizobium on nodulation and nitrogen fixation by pigeonpea (Cajanus cajan). Eur. J. Soil Sci. 2006, 57, 67–71. [Google Scholar] [CrossRef]
- Bhattacharjee, S.; Sharma, G.D. Effect of dual inoculation of arbuscular mycorrhiza and Rhizobium on the chlorophyll, nitrogen and phosphorus contents of pigeon pea (Cajanus cajan L.). Adv. Microbiol. 2012, 2, 25945. [Google Scholar] [CrossRef][Green Version]
- Haskett, T.; Wang, P.; Ramsay, J.; O’Hara, G.; Reeve, W.; Howieson, J.; Terpolilli, J. Complete genome sequence of Mesorhizobium ciceri strain CC1192, an efficient nitrogen-fixing microsymbiont of Cicer arietinum. Genome Announc. 2016, 4, e00516-16. [Google Scholar] [CrossRef]
- Weiss, V.A.; Faoro, H.; Tadra-Sfeir, M.Z.; Raittz, R.T.; de Souza, E.M.; Monteiro, R.A.; Cardoso, R.L.A.; Wassem, R.; Chubatsu, L.S.; Huergo, L.F. Draft genome sequence of Herbaspirillum lusitanum P6-12, an endophyte isolated from root nodules of Phaseolus vulgaris. J. Bacteriol. 2012, 194, 4136–4137. [Google Scholar] [CrossRef]
- Simões-Araújo, J.L.; Leite, J.; Passos, S.R.; Xavier, G.R.; Rumjanek, N.G.; Zilli, J.É. Draft genome sequence of Bradyrhizobium sp. strain BR 3267, an elite strain recommended for cowpea inoculation in Brazil. Braz. J. Microbiol. 2016, 47, 781–782. [Google Scholar] [CrossRef][Green Version]
- Schuldes, J.; Orbegoso, M.R.; Schmeisser, C.; Krishnan, H.B.; Daniel, R.; Streit, W.R. Complete genome sequence of the broad-host-range strain Sinorhizobium fredii USDA257. J. Bacteriol. 2012, 194, 4483. [Google Scholar] [CrossRef] [PubMed]
- Siqueira, A.F.; Ormeño-Orrillo, E.; Souza, R.C.; Rodrigues, E.P.; Almeida, L.G.P.; Barcellos, F.G.; Batista, J.S.S.; Nakatani, A.S.; Martínez-Romero, E.; Vasconcelos, A.T.R. Comparative genomics of Bradyrhizobium japonicum CPAC 15 and Bradyrhizobium diazoefficiens CPAC 7: Elite model strains for understanding symbiotic performance with soybean. BMC Genom. 2014, 15, 1–21. [Google Scholar] [CrossRef] [PubMed]
- Zhu, B.; Liu, H.; Tian, W.-X.; Fan, X.-Y.; Li, B.; Zhou, X.-P.; Jin, G.-L.; Xie, G.-L. Genome sequence of Stenotrophomonas maltophilia RR-10, isolated as an endophyte from rice root. Genome Announc. 2012, 194, 1280–1281. [Google Scholar] [CrossRef] [PubMed]
- Mathimaran, N.; Srivastava, R.; Wiemken, A.; Sharma, A.K.; Boller, T. Genome sequences of two plant growth-promoting fluorescent Pseudomonas strains, R62 and R81. J. Bacteriol. 2012, 194, 3272–3273. [Google Scholar] [CrossRef]
- Gamez, R.M.; Rodríguez, F.; Bernal, J.F.; Agarwala, R.; Landsman, D.; Mariño-Ramírez, L. Genome sequence of the banana plant growth-promoting rhizobacterium Bacillus amyloliquefaciens BS006. Genome Announc. 2015, 3, e01391-15. [Google Scholar] [CrossRef]
- Wisniewski-Dyé, F.; Lozano, L.; Acosta-Cruz, E.; Borland, S.; Drogue, B.; Prigent-Combaret, C.; Rouy, Z.; Barbe, V.; Herrera, A.M.; González, V. Genome sequence of Azospirillum brasilense CBG497 and comparative analyses of Azospirillum core and accessory genomes provide insight into niche adaptation. Genes 2012, 3, 576–602. [Google Scholar] [CrossRef] [PubMed]
- Christensen, H.; Kuhnert, P.; Olsen, J.E.; Bisgaard, M. Comparative phylogenies of the housekeeping genes atpD, infB and rpoB and the 16S rRNA gene within the Pasteurellaceae. Int. J. Syst. Evol. Microbiol. 2004, 54 Pt 5, 1601–1609. [Google Scholar] [CrossRef]
- Rivas, R.; Martens, M.; De Lajudie, P.; Willems, A. Multilocus sequence analysis of the genus Bradyrhizobium. Syst. Appl. Microbiol. 2009, 32, 101–110. [Google Scholar] [CrossRef]
- Edwards, U.; Rogall, T.; Blöcker, H.; Emde, M.; Böttger, E.C. Isolation and direct complete nucleotide determination of entire genes. Characterization of a gene coding for 16S ribosomal RNA. Nucleic Acids Res. 1989, 17, 7843–7853. [Google Scholar] [CrossRef] [PubMed]
- Tan, H.W.; Weir, B.S.; Carter, N.; Heenan, P.B.; Ridgway, H.J.; James, E.K.; Sprent, J.I.; Young, J.P.W.; Andrews, M. Rhizobia with 16S rRNA and nifH similar to Mesorhizobium huakuii but novel recA, glnII, nodA and nodC genes are symbionts of New Zealand Carmichaelinae. PLoS ONE 2012, 7, e47677. [Google Scholar] [CrossRef] [PubMed]
- Venkadesaperumal, G.; Amaresan, N.; Kumar, K. Plant growth promoting capability and genetic diversity of bacteria isolated from mud volcano and lime cave of Andaman and Nicobar Islands. Braz. J. Microbiol. 2014, 45, 1271–1281. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Mubeen, S.; Ahmed, I.; Khalid, N.; Suleria, H.A.R.; Bano, A.; Mumtaz, A.S. Identification and characterization of rhizospheric microbial diversity by 16S ribosomal RNA gene sequencing. Braz. J. Microbiol. 2014, 45, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Laguerre, G.; Mavingui, P.; Allard, M.-R.; Charnay, M.-P.; Louvrier, P.; Mazurier, S.-I.; Rigottier-Gois, L.; Amarger, N. Typing of rhizobia by PCR DNA fingerprinting and PCR-restriction fragment length polymorphism analysis of chromosomal and symbiotic gene regions: Application to Rhizobium leguminosarum and its different biovars. Appl. Environ. Microbiol. 1996, 62, 2029–2036. [Google Scholar] [CrossRef]
- Zhang, J.J.; Liu, T.Y.; Chen, W.F.; Wang, E.T.; Sui, X.H.; Zhang, X.X.; Li, Y.; Li, Y.; Chen, W.X. Mesorhizobium muleiense sp. nov., nodulating with Cicer arietinum L. Int. J. Syst. Evol. Microbiol. 2012, 62, 2737–2742. [Google Scholar] [CrossRef]
- Diouf, F.; Diouf, D.; Klonowska, A.; Le Queré, A.; Bakhoum, N.; Fall, D.; Neyra, M.; Parrinello, H.; Diouf, M.; Ndoye, I. Genetic and genomic diversity studies of Acacia symbionts in Senegal reveal new species of Mesorhizobium with a putative geographical pattern. PLoS ONE 2015, 10, e0117667. [Google Scholar] [CrossRef]
- De Bruijn, F.J. Use of repetitive (repetitive extragenic palindromic and enterobacterial repetitive intergeneric consensus) sequences and the polymerase chain reaction to fingerprint the genomes of Rhizobium meliloti isolates and other soil bacteria. Appl. Environ. Microbiol. 1992, 58, 2180–2187. [Google Scholar] [CrossRef] [PubMed]
- De Meyer, S.E.; Van Hoorde, K.; Vekeman, B.; Braeckman, T.; Willems, A. Genetic diversity of rhizobia associated with indigenous legumes in different regions of Flanders (Belgium). Soil Biol. Biochem. 2011, 43, 2384–2396. [Google Scholar] [CrossRef]
- Kaschuk, G.; Hungria, M.; Andrade, D.S.; Campo, R.J. Genetic diversity of rhizobia associated with common bean (Phaseolus vulgaris L.) grown under no-tillage and conventional systems in Southern Brazil. Appl. Soil Ecol. 2006, 32, 210–220. [Google Scholar] [CrossRef]
- Gaunt, M.W.; Turner, S.L.; Rigottier-Gois, L.; Lloyd-Macgilp, S.A.; Young, J.P. Phylogenies of atpD and recA support the small subunit rRNA-based classification of rhizobia. Int. J. Syst. Evol. Microbiol. 2001, 51, 2037–2048. [Google Scholar] [CrossRef] [PubMed]
- Kim, D.H.; Kaashyap, M.; Rathore, A.; Das, R.R.; Parupalli, S.; Upadhyaya, H.D.; Gopalakrishnan, S.; Gaur, P.M.; Singh, S.; Kaur, J. Phylogenetic diversity of Mesorhizobium in chickpea. J. Biosci. 2014, 39, 513–517. [Google Scholar] [CrossRef] [PubMed]
- Payne, G.W.; Vandamme, P.; Morgan, S.H.; LiPuma, J.J.; Coenye, T.; Weightman, A.J.; Jones, T.H.; Mahenthiralingam, E. Development of a recA gene-based identification approach for the entire Burkholderia genus. Appl. Environ. Microbiol. 2005, 71, 3917–3927. [Google Scholar] [CrossRef] [PubMed]
- Blažinkov, M.; Sikora, S.; Uher, D.; Mačešić, D.; Redžepović, S. Genotypic characterisation of indigenous Rhizobium leguminosarum bv. viciae field population in Croatia. Agric. Conspec. Sci. 2007, 72, 153–158. [Google Scholar]
- Marinković, J.; Đorđević, V.; Balešević-Tubić, S.; Bjelić, D.; Vucelić-Radović, B.; Jošić, D. Osmotic stress tolerance, PGP traits and RAPD analysis of Bradyrhizobium japonicum strains. Genetika 2013, 45, 75–86. [Google Scholar] [CrossRef]
- Kasa, P.; Modugapalem, H.; Battini, K. Isolation, screening, and molecular characterization of plant growth promoting rhizobacteria isolates of Azotobacter and Trichoderma and their beneficial activities. J. Nat. Sci. Biol. Med. 2015, 6, 360–363. [Google Scholar] [CrossRef]
- Lerner, A.; Herschkovitz, Y.; Baudoin, E.; Nazaret, S.; Moenne-Loccoz, Y.; Okon, Y.; Jurkevitch, E. Effect of Azospirillum brasilense inoculation on rhizobacterial communities analyzed by denaturing gradient gel electrophoresis and automated ribosomal intergenic spacer analysis. Soil Biol. Biochem. 2006, 38, 1212–1218. [Google Scholar] [CrossRef]
- Sachdev, D.; Nema, P.; Dhakephalkar, P.; Zinjarde, S.; Chopade, B. Assessment of 16S rRNA gene-based phylogenetic diversity and promising plant growth-promoting traits of Acinetobacter community from the rhizosphere of wheat. Microbiol. Res. 2010, 165, 627–638. [Google Scholar] [CrossRef]
- Sharma, R.; Pooniya, V.; Bisaria, V.S.; Swarnalakshmi, K.; Sharma, S. Bioinoculants play a significant role in shaping the rhizospheric microbial community: A field study with Cajanus cajan. World J. Microbiol. Biotechnol. 2020, 36, 1–17. [Google Scholar] [CrossRef]
- Sharma, S.B.; Sayyed, R.Z.; Trivedi, M.H.; Gobi, T.A. Phosphate solubilizing microbes: Sustainable approach for managing phosphorus deficiency in agricultural soils. Springerplus 2013, 2, 587. [Google Scholar] [CrossRef]
- Ding, Y.; Wang, J.; Liu, Y.; Chen, S. Isolation and identification of nitrogen-fixing bacilli from plant rhizospheres in Beijing region. J. Appl. Microbiol. 2005, 99, 1271–1281. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Rasool, M.; Mirza, M.S. Nitrogen fixation and indole acetic acid production potential of bacteria isolated from rhizosphere of sugarcane (Saccharum officinarum L.). Adv. Biol. Res. 2011, 5, 348–355. [Google Scholar]
- Jha, Y.; Subramanian, R.B. Characterization of root-associated bacteria from paddy and its growth-promotion efficacy. 3 Biotech 2014, 4, 325–330. [Google Scholar] [CrossRef] [PubMed][Green Version]
- Naveed, M.; Sohail, Y.; Khalid, N.; Ahmed, I.; Mumtaz, A.S. Evaluation of glucose dehydrogenase and pyrroloquinoline quinine (pqq) mutagenesis that renders functional inadequacies in host plants. J. Microbiol. Biotechnol. 2015, 25, 1349–1360. [Google Scholar] [CrossRef]
- Ovaa, W.; Bitter, W.; Weisbeek, P.; Koster, M. Multiple outer membrane receptors for uptake of ferric pseudobactins in Pseudomonas putida WCS358. Mol. Gen. Genet. 1995, 248, 735–743. [Google Scholar] [CrossRef]
- Woo, S.M.; Lim, J.H.; Jeong, H.Y.; Kim, S.D. Genetic monitoring of plant growth promoting rhizobacterium (PGPR), Bacillus subtilis AH18 using multiplex PCR in field soil. Microbiol. Biotechnol. Lett. 2009, 37, 1–9. [Google Scholar]
- Lim, J.H.; Ahn, C.H.; Jeong, H.Y.; Kim, Y.H.; Kim, S.D. Genetic monitoring of multi-functional plant growth promoting rhizobacteria Bacillus subtilis AH18 and Bacillus licheniformis K11 by multiplex and real-time polymerase chain reaction in a pepper farming field. J. Korean Soc. Appl. Biol. Chem. 2011, 54, 221–228. [Google Scholar] [CrossRef]
- Patten, C.L.; Glick, B.R. Role of Pseudomonas putida indoleacetic acid in development of the host plant root system. Appl. Environ. Microbiol. 2002, 68, 3795–3801. [Google Scholar] [CrossRef] [PubMed]
- Brandao, P.F.B.; Clapp, J.P.; Bull, A.T. Diversity of nitrile hydratase and amidase enzyme genes in Rhodococcus erythropolis recovered from geographically distinct habitats. Appl. Environ. Microbiol. 2003, 69, 5754–5766. [Google Scholar] [CrossRef] [PubMed]


{kind=link}
| Crop | Microbes | Beneficial Effects | References |
|---|---|---|---|
| Cicer arietinum (Chickpea) | Pseudomonas aeruginosa, Pseudomonas alcaligenes, Pseudomonas fluorescens BHUPSB06, Pseudomonas jessenii PS06 | Enhanced acquisition of P and Fe, effective symbiosis with Mesohizobium | [23,24,25] |
| Pseudomonas alcaligenes, Bacillus pumilus | Increase in shoot dry mass, pod number, nodulation, chlorophyll content, N, P and K content | [24,25] | |
| Azospirillum lipoferum FK1, Azospirillum brasilense | Improved nodulation and growth | [19,26] | |
| Azotobacter | Increase in plant–rhizobial symbiosis, biomass, grain yield, N content | [27] | |
| Lens culinaris (Lentil) | Bacillus megaterium Kurthia sp. LK786, Pseudomonas diminuta LK884 | Enhanced symbiotic effect of Rhizobium leguminosarum and improved plant growth | [28,29] |
| Pseudomonas sp. | Enhanced symbiotic effect of Rhizobium leguminosarum and improved plant growth | [30] | |
| Proteus vulgaris | Increased nodulation potential when given in combination with Rhizobium leguminosarum L-12-8 | [31] | |
| Vigna radiata (Green gram) | Bacillus subtilis, Bacillus megaterium | Increase in dry matter and N and P uptake | [32,33] |
| Pseudomonas putida GRP3A | Stimulated iron uptake | [34] | |
| Pseudomonas sp. | Increase in plant height and improved root development | [34,35,36] | |
| Cajanus cajan (Pigeonpea) | Bacillus subtilis AF1, Bacillus cereus BS03 | Increase in shoot, root length, nodulation and biomass | [37,38] |
| Pseudomonas spp., Pseudomonas aeruginosa RRLJ | Significant increase in plant growth and nodulation occupancy of Rhizobium | [38,39] | |
| Azotobacter chroococcum A41, Bacillus megaterium MTCC 453, Pseudomonas fluorescens MTCC9768. | Improved plant growth and yield | [40] | |
| Arachis hypogaea (Groundnut) | Bacillus, Pseudomonas fluorescens | Enhanced synergistic activity of rhizobia for nutrient uptake and plant growth | [41,42] |
| Glycine max (Soybean) | Bacillus amyloliquefaciens LL2012, Bacillus subtilis | Enhanced symbiotic capacity of Bradyrhizobium japonicum | [31,43] |
| Azospirillum brasilense Sp7, Azospirillum lipoferum CCM3863 | Efficient symbiosis with Bradyrhizobium japonicum and enhancement in root growth and shoot dry matter | [44] | |
| Pseudomonas cepacia | Enhanced synergistic activity with Bradyrhizobium japonicum TAL-378 resulted in overall improved plant growth | [45,46] | |
| Phaseolus vulgaris (Common bean) | Bacillus megaterium | Increased nodulation, shoot dry weight, nodule dry weight and chlorophyll content | [47] |
| Paenibacillus polymyxa DSM 36 and Loutit (L) | Increased symbiotic efficiency of Rhizobium tropici | [48] | |
| Azospirillum brasilense, Azospirillum lipoferum S21 | Enhancement of nodulation and N2 fixation activity of Rhizobium | [49,50] | |
| Pseudomonas monteilii, Pseudomonas fluorescens P93 | Synergistic effect of Rhizobium pisi leading to increased nodulation | [48,51] | |
| Vicia faba (Faba bean) | Azospirillum brasilense, Azospirillum lipoferum SM1, Azospirillum brasilense | Increase in growth of root, shoot and improved nodulation | [26,52] |
| Azotobacter chroococcum H23, Azotobacter vinelandii ATCC12837 and Dv42 | Increased nodulation, dry mater and total N content | [53] | |
| Pseudomonas aeruginosa, Pseudomonas putida TK3, Serratia marcescens BM1 Serratia liquefaciens BM4, Xanthobacter autotrophicus BM3 | Increase in the phytoremediation potential Increase in shoot dry weight, number of pods per plant and nodule dry weight | [52,54] | |
| Pseudomonas fluorescens, Pseudomonas alcaligenes PsA15, Pseudomonas denitrificans PsD6 | Increase in fresh and dry weight, root and shoot length, number of leaves per plant | [55,56] | |
| Bacillus polymyxa BcP26, Mycobacterium phlei MbP18, Cellulomonas sp. 32 | Increase in root and shoot growth, nodulation, increase in N and P content | [55] | |
| Phaseolus vulgaris (French bean) | Pseudomonas lurida NPRp15, Pseudomonas putida PGRs4 | Increased root and shoot dry weight, nodulation, nutrient uptake, pod yield | [57] |
| Vigna unguiculata (Cowpea) | Pontibacter niistensis NII-0905 | Increase in root number, root length, shoot length and dry biomass | [58] |
| Legume Crop | Associated Rhizobial Strains | Amount of N Fixed (kg ha−1) | Method of Estimation | Reference |
|---|---|---|---|---|
| Cicer arietinum | Bradyrhizobium ciceri bvs. CP31, CP36 | 19–24 | 15N isotope dilution | [75] |
| Rhizobium sp. | 15–32 | 15N natural abundance | [76] | |
| Cajanus cajan | Rhizobium sp. IHP114 | 13–69 | N Difference | [77] |
| Vigna radiata | Bradyrhizobium japonicum 542 | 116 | N Difference | [78] |
| Lens culinaris | Rhizobium leguminosarum bv. viciae su391 | 25 | 15N isotope dilution | [79] |
| Rhizobium leguminosarum | 0–105 | 15N isotope dilution | [80] | |
| Rhizobium sp. | 37–55 | 15N natural abundance | [81] | |
| Bradyrhizobium sp. | 82 | N Difference | [82] | |
| Phaseolus vulgaris | Rhizobium leguminosarum bv. phaseoli | 24–39 | 15N isotope dilution | [83] |
| Rhizobium leguminosarum bv. phaseoli | 11–165 | 15N isotope dilution | [84] | |
| Phaseolus vulgaris | Rhizobium phaseoli | 78.7 | 15N isotope dilution | [85] |
| Vicia faba | Rhizobium leguminosarum bv. viciae | 76–125 | 15N isotope dilution | [75] |
| Bradyrhizobium sp. | 210 | N Difference | [82] | |
| Rhizobium phaseoli | 3.8 | acetylene reduction | [85] | |
| Pisum sativum | Rhizobium leguminosarum bv. viciae su-391 | 34–112 | 15N isotope dilution | [79] |
| Bradyrhizobium sp. | 128 | N Difference | [82] | |
| Rhizobium leguminosarum bv. viciae | 31–107 | 15N isotope dilution | [75] | |
| Arachis hypogaea | Rhizobium sp. | 186 | N Difference | [86] |
| Bradyrhizobium sp. | 150–200 | 15N isotope dilution | [87,88] | |
| Glycine max | Bradyrhizobium japonicum | 102.9 | 15N isotope dilution | [85] |
| Bradyrhizobium japonicum | 25.6 | acetylene reduction | [85] | |
| Bradyrhizobium sp. | 108–152 | 15N isotope dilution | [88] |
| PGPR | Crop | Reference |
|---|---|---|
| Pseudomonas aeruginosa | Cicer arietinum | [100] |
| Pseudomonas alcaligenes, Bacillus pumilus | Cicer arietinum | [24] |
| Bacillus megaterium | Lens culinaris | [28] |
| Bacillus megaterium | Phaseolus vulgaris | [47] |
| Pseudomonas fluorescens | Arachis hypogaea | [41] |
| Pseudomonas lurida-NPRp15 and Pseudomonas putida-PGRs4 | Phaseolus vulgaris | [57] |
| Bacillus subtilis | Vigna radiata | [32] |
| Legume Crop | Associated PGPR | Effect | Reference |
|---|---|---|---|
| Cicer arietinum | Serratia proteamaculans J119 | Improved root and shoot growth, nodulation, grain yield | [169] |
| Mesorhizobium ciceri LMS1 | Increase in nodulation and plant growth | [164] | |
| Mesorhizobium | Improved plant growth under salinity stress | [170] | |
| Lens culinaris | Bacillus cereus, Pseudomonas sp. | Promoted plant growth under axenic conditions | [171] |
| Vigna radiata | Pseudomonas putida, Pseudomonas fluorescens, Bradyrhizobium japonicum | Root elongation, increase in nodule number, nodule fresh and dry weight | [172] |
| Pseudomonas fluorescens, Pseudomonas syringae, Rhizobium phaseoli | Significantly reduced salinity stress and increase plant growth | [173] | |
| Pisum sativum | Rhizobium leguminosarum bv. viciae 128C53K | Increased nodulation | [162] |
| Arthrobacter protophormiae | Increased plant tolerance to salt stress and improved plant growth | [174] | |
| Pseudomonas brassicacearum Am3, Pseudomonas marginalis Dp1, | Enhanced nutrient uptake | [175] | |
| Pseudomonas fluorescens, Pseudomonas putida, | Reduced drought stress on plant | [176] | |
| Variovorax paradoxus 5C2 | Improved growth, yield and water use efficiency of drought stressed plants | [177] | |
| Glycine max | Pseudomonas sp. | Increased plant growth and reduced plant fungal disease | [178] |
| Arachis hypogaea | Pseudomonas sp. | Enhanced growth, yield and nutrient uptake | [41] |
| Pseudomonas fluorescens TDK1 | Enhanced resistance to saline stress | [179] | |
| Cyamopsis tetragonoloba (Cluster bean) | Pseudomonas sp. | Improved nodulation and plant growth | [180] |
| Vigna unguiculata | Pseudomonas sp. | Improved plant growth under salt stress | [181] |
| Co-Inoculated Strains | Legume Plant | Positive Effects on Plant Growth Parameters | Reference |
|---|---|---|---|
| Rhizobium leguminosarum bv. viciae, Pseudomonas sp. (PSB), Pseudomonas sp. (PGPR) | Lens culinaris | 81% increase in nodule number, 57% increase in nodule dry weight and 15% improvement in grain yield * | [192] |
| Rhizobium CRM 6, Bacillus polymyxa (PSB), PGPR (KB 133) | Vigna radiata | 110% increase in nodule number, 121% increase in nodule weight and 44% increase in grain yield * | [193] |
| Bacillus amyloliquefaciens LL2012, Bradyrhizobium japonicum | Glycine max | 50% increase in shoot dry weight and 40% increase in root dry weight # | [43] |
| Mesorhizobium ciceri CH-1233, Pseudomonas sp. LK884 | Cicer arietinum | 56% increase in nodule number, 100% increase in nodule dry weight, and 15% increase in grain yield * | [194] |
| Bradyrhizobium sp., Serratia marcescens, Trichoderma harzianum | Arachis hypogaea | 115% increase in nodule number, 94% increase in nodule dry weight and 41% increase in grain yield * | [195] |
| Rhizobium, Azotobacter chroococcum | Cajanus cajan | 248% increase in nodule number, 100% increase in nodule dry weight and 92% increase in N fixation, and 19% increase in grain yield # | [196] |
| Rhizobium, Pseudomonas fluorescens | Cajanus cajan | 388% increase in nodule number, 267% increase in nodule dry weight and 134% increase in N fixation and 66% increase in grain yield # | [196] |
| Rhizobium, Bacillus cereus | Cajanus cajan | 382% increase in nodule number, 196% increase in nodule dry weight and 116% increase in N fixation and 54% increase in grain yield # | [196] |
| Glomus fasciculatum (VAM), Rhizobium | Cajanus cajan | 19% increase in chlorophyll content, 10% increase in N content and 114% increase in P content # | [197] |
| PGPR | Host Plant | Genome Size (Mb) | Reference |
|---|---|---|---|
| Mesorhizobium ciceri CC1192 | Cicer arietinum | 6.94 | [198] |
| Herbaspirillum lusitanum P6-12 | Phaseolus vulgaris | 4.46 | [199] |
| Bradyrhizobium yuanmingense BR 3267 | Vigna unguiculata | 7.90 | [200] |
| Sinorhizobium fredii USDA257 | Glycine max | 6.47 | [201] |
| Bradyrhizobium japonicum CPAC 15, Bradyrhizobium diazoefficiens CPAC 7 | Glycine max | 9.58 | [202] |
| Stenotrophomonas maltophilia RR-10 | Oryza sativa (Rice) | 4.66 | [203] |
| Pseudomonas strain R62 and R81 | Triticum sp. (Wheat) | 6.00 | [204] |
| Bacillus amyloliquefaciens BS006 | Musa sp. (Banana) | 4.17 | [205] |
| Azospirillum brasilense CBG497 | Zea mays (Maize) | 6.50 | [206] |
| Method | PGPR Community/Source Plant | Reference |
|---|---|---|
| 16S rDNA sequencing | Rhizobia, Pantoea agglomerans, Exiguobacterium, Ensifer, Bacillus sp., Pseudomonas and Leclercia | [209,210,211,212] |
| 16S-23S IGS sequencing | Rhizobium leguminosarum bv. viciae, trifolii, and phaseoli, Mesorhizobium populations | [213,214] |
| REP-PCR, ERIC-PCR DNA fingerprinting | Mesorhizobia sp. | [215] |
| Rhizobium meliloti solates | [216] | |
| Rhizobia associated with Belgium legumes | [217] | |
| Box PCR | Mesorhizobium populations associated from Chickpea | [214] |
| Rhizobia associated with common bean | [218] | |
| ARDRA | Mesorhizobium, common bean rhizobia | [214,218] |
| MLSA | gyrB (DNA gyrase), rpoD (RNA polymerase) of Pseudomonas | [217] |
| atpD (ATP synthase) | [219] | |
| gyrB, nifK and nod genes of Mesorhizobium, | [220] | |
| recA of Burkholderia sp. | [221] | |
| RAPD-PCR | Rhizobium leguminosarum bv. viciae strains, Azotobacter and Trichoderma strains, Bradyrhizobium japonicum strains | [222,223,224] |
| DGGE | Azospirillum brasilense in maize | [225] |
| Acinetobacter community from wheat Rhizospheric microbial community in pigeonpea | [226,227] |
| PGP Trait | Related Genes | PGPR Strains | References |
|---|---|---|---|
| Nitrogen fixation | nifH, nifD, nifK (nitrogenase iron protein) | Paenibacillus sp., Klebsiella sp., Azospirillum sp., Burkholderia sp., Bacillus sp., Mesorhizobium sp. | [229,230,231] |
| Phosphate solubilization | pqqC, pqqBCD, pqqAB, pqqE, pqqF (Pyrrolo Quinoline Quinone Synthase) gdh (Glucose Dehydrogenase, cofactor for pqq genes) | Pseudomonas sp., Pseudomonas fluorescens QAU67, Pseudomonas putida QAU90, Bacillus sp. | [232] |
| Siderophores production | pupa (siderophore transporter), sid (siderophore synthesis), dhbF (2,3-Dihydroxy Benzoate synthesis gene) | Pseudomonas putida Bacillus subtilis AH18 Bacillus licheniformis K11 | [233,234,235] |
| IAA synthesis | nhase (nitrile hydratase), amd (amidase), ipdC (indole-3-pyruvate decarboxylase), aec (auxin efflux carrier protein) | Rhodococcus erythropolis, Pseudomonas putida Bacillus subtilis AH18 | [234,236,237] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Swarnalakshmi, K.; Yadav, V.; Tyagi, D.; Dhar, D.W.; Kannepalli, A.; Kumar, S. Significance of Plant Growth Promoting Rhizobacteria in Grain Legumes: Growth Promotion and Crop Production. Plants 2020, 9, 1596. https://doi.org/10.3390/plants9111596
Swarnalakshmi K, Yadav V, Tyagi D, Dhar DW, Kannepalli A, Kumar S. Significance of Plant Growth Promoting Rhizobacteria in Grain Legumes: Growth Promotion and Crop Production. Plants. 2020; 9(11):1596. https://doi.org/10.3390/plants9111596
Chicago/Turabian StyleSwarnalakshmi, Karivaradharajan, Vandana Yadav, Deepti Tyagi, Dolly Wattal Dhar, Annapurna Kannepalli, and Shiv Kumar. 2020. "Significance of Plant Growth Promoting Rhizobacteria in Grain Legumes: Growth Promotion and Crop Production" Plants 9, no. 11: 1596. https://doi.org/10.3390/plants9111596
APA StyleSwarnalakshmi, K., Yadav, V., Tyagi, D., Dhar, D. W., Kannepalli, A., & Kumar, S. (2020). Significance of Plant Growth Promoting Rhizobacteria in Grain Legumes: Growth Promotion and Crop Production. Plants, 9(11), 1596. https://doi.org/10.3390/plants9111596