The Genus Pratylenchus (Nematoda: Pratylenchidae) in Israel: From Taxonomy to Control Practices


 ,
,
Abstract

1. Introduction
2. Overview of Israeli Agriculture
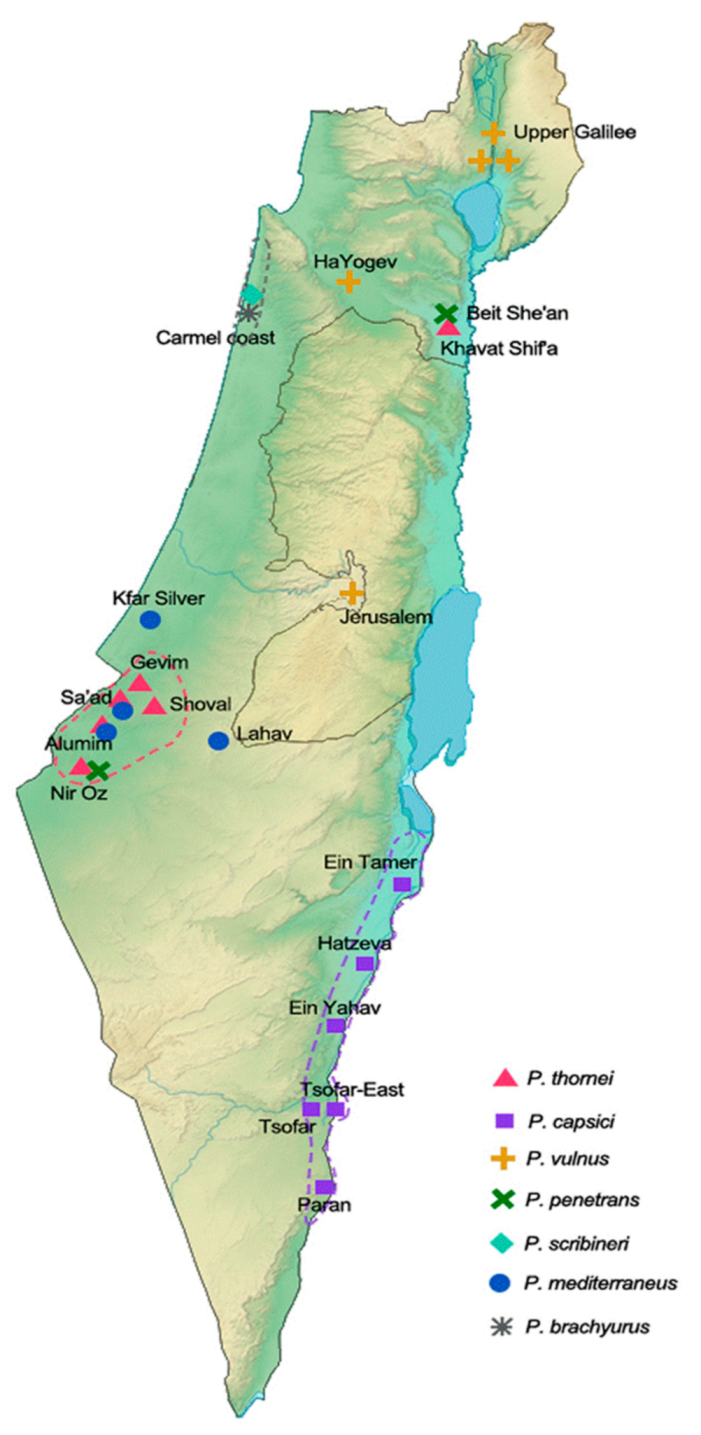
3. Occurrence of Pratylenchus Species in Israel
4. Taxonomy and Diversity of Pratylenchus Species in Israel
4.1. Pratylenchus mediterraneus Corbett, 1983
4.2. Pratylenchus thornei Sher and Allen, 1953
4.3. Pratylenchus neglectus (Rensch, 1924) Filipjev and S. Stekhoven, 1941
4.4. Pratylenchus vulnus Allen and Jensen, 1951
4.5. Pratylenchus pratensis (de Man, 1880) Filipjev, 1936
4.6. Pratylenchus capsici Qing, Bert, Gamliel, Bucki, Duvrinin, Alon, Braun-Miyara, 2019
4.7. Pratylenchus penetrans (Cobb, 1917) Filipjev and Stekhoven, 1941
4.8. Pratylenchus scribneri Steiner in Sherbakoff and Stanley, 1943
4.9. Pratylenchus brachyurus (Godfrey, 1929) Filipjev and Stekhoven, 1941
5. Biology and Pathogenicity of Pratylenchus Species
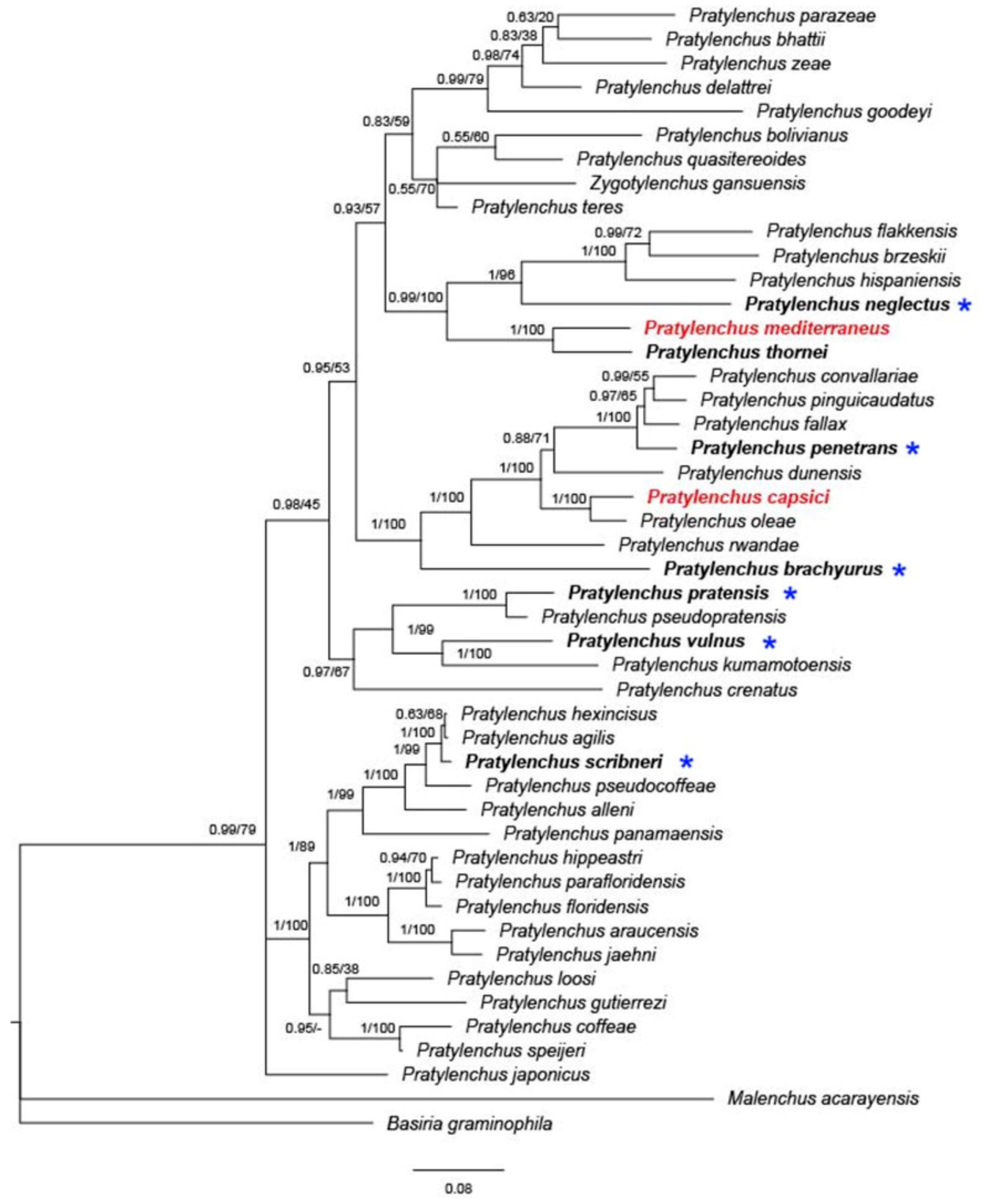
6. Phylogeny and Evolution of Pratylenchus Species Occurring in Israel
7. Control and Management Practices
7.1. The Use of Soil Fumigants
7.2. Common Control Methods in Used to Manage Plant Parasitic Nematodes
7.3. Resistance to Root-Lesion Nematodes
8. Challenges and Perspectives for Pratylenchus Research in Israel
8.1. Taxonomy and Diagnosis of Pratylenchus Species
8.2. Control/Management of Pratylenchus Species
9. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Geraert, E. The Pratylenchidae of the World: Identification of the Family Pratylenchidae (Nematoda: Tylenchida); Academia Press: Ghent, Belgium, 2013. [Google Scholar]
- Hodda, M.; Collins, S.J.; Vanstone, V.A.; Hartley, D.; Wanjura, W.; Kehoe, M. Pratylenchus quasitereoides n. sp. from cereals in Western Australia. Zootaxa 2014, 3866, 277–288. [Google Scholar] [CrossRef]
- Palomares-Rius, J.E.; Guesmi, I.; Horrigue-Raouani, N.; Cantalapiedra-Navarrete, C.; Liébanas, G.; Castillo, P. Morphological and molecular characterisation of Pratylenchus oleae n. sp.(Nematoda: Pratylenchidae) parasitizing wild and cultivated olives in Spain and Tunisia. Eur. J. Plant Pathol. 2014, 140, 53–67. [Google Scholar] [CrossRef]
- Nicol, J.; Turner, S.; Coyne, D.; Den Nijs, L.; Hockland, S.; Maafi, Z.T. Current nematode threats to world agriculture. In Genomics and Molecular Genetics of Plant-Nematode Interactions; Springer: Dordrecht, The Netherlands, 2011; pp. 21–43. [Google Scholar]
- Jones, M.; Fosu-Nyarko, J. Molecular biology of root lesion nematodes (Pratylenchus spp.) and their interaction with host plants. Ann. Appl. Biol. 2014, 164, 163–181. [Google Scholar] [CrossRef]
- Minz, G. Free-living plant-parasitic and possible plant-parasitic nematodes in Israel. Plant Dis. Report. 1957, 41, 92–94. [Google Scholar]
- Cohn, E.; Sher, S.; Bell, A.; Minz, G. Soil Nematodes Occurring in Israel; The Volcani Center: Rishon Letzion, Israel, 1973. [Google Scholar]
- Glazer, I.; Orion, D. Studies on anhydrobiosis of Pratylenchus thornei. J. Nematol. 1983, 15, 333. [Google Scholar]
- Krikun, J.; Orion, D. Verticillium wilt of potato: Importance and control. Phytoparasitica 1979, 7, 107. [Google Scholar] [CrossRef]
- Orion, D.; Amir, J.; Krikun, J. Field observations on Pratylenchus thornei and its effects on wheat under arid conditions. Rev. Nématol. 1984, 7, 341–345. [Google Scholar]
- Orion, D.; Glazer, I. Nematicide seed dressing for Pratylenchus mediterraneus control in wheat. Phytoparasitica 1987, 15, 225–228. [Google Scholar] [CrossRef]
- Orion, D.; Lapid, D. Scanning Electron-Microscope Study on the Interaction of Pratylenchus-Mediterraneus and Vicia-Sativa Roots. Nematologica 1993, 39, 322–327. [Google Scholar] [CrossRef]
- Orion, D.; Nachmias, A.; Lapid, D.; Orenstein, J. Observations on the parasitic behavior of Pratylenchus mediterraneus on excised potato roots. Nematropica 1995, 25, 71–74. [Google Scholar]
- Orion, D.; Shlevin, E. Nematicide seed dressing for cyst and lesion nematode control in wheat. J. Nematol. 1989, 21, 629. [Google Scholar] [PubMed]
- Goldreich, Y.; Karni, O. Climate and precipitation regime in the Arava Valley, Israel. Isr. J. Earth Sci. 2001, 50, 53–59. [Google Scholar] [CrossRef]
- Yair, A. Runoff generation in a sandy area—The Nizzana sands, Western Negev, Israel. Earth Surf. Process. Landf. 1990, 15, 597–609. [Google Scholar] [CrossRef]
- Pasternak, D.; De Malach, Y.; Borovic, I.; Shram, M.; Aviram, C. Irrigation with brackish water under desert conditions IV. Salt tolerance studies with lettuce (Lactuca sativa L.). Agric. Water Manag. 1986, 11, 303–311. [Google Scholar] [CrossRef]
- Qing, X.; Bert, W.; Gamliel, A.; Bucki, P.; Duvrinin, S.; Alon, T.; Braun Miyara, S. Phylogeography and Molecular Species Delimitation of Pratylenchus capsici n. sp., a New Root-Lesion Nematode in Israel on Pepper (Capsicum annuum). Phytopathology 2019, 109, 847–858. [Google Scholar] [CrossRef]
- Orion, D.; Krikun, J.; Sullami, M. The distribution, pathogenicity and ecology ofPratylenchus Thornei in the northern negev. Phytoparasitica 1979, 7, 3–9. [Google Scholar] [CrossRef]
- Cobertt, D.; Clark, A. Surface features in the taxonomy of Pratylenchus species. Rev. Néinatol 1983, 6, 85–98. [Google Scholar]
- Ryss, A.Y. Root Parasitic Nematodes of the Family Pratylenchidae (Tylenchida) of the World Fauna; Nauka: Leningrad, Russia, 1988. [Google Scholar]
- Café Filho, A.C.; Huang, C. Description of Pratylenchus pseudofallax n. sp. with a key to species of the genus Pratylenchus Filipjev, 1936 (Nematoda: Pratylenchidae). Rev. Nématol. 1989, 12, 7–15. [Google Scholar]
- Castillo, P.; Vovlas, N. Pratylenchus (Nematoda: Pratylenchidae): Diagnosis, biology, pathogenicity and management. In Nematology Monographs and Perspectives; Brill: Leiden, The Netherlands, 2007; Volume 6. [Google Scholar]
- De Luca, F.; Reyes, A.; Troccoli, A.; Castillo, P. Molecular variability and phylogenetic relationships among different species and populations of Pratylenchus (Nematoda: Pratylenchidae) as inferred from the analysis of the ITS rDNA. Eur. J. Plant Pathol. 2011, 130, 415–426. [Google Scholar] [CrossRef]
- Troccoli, A.; De Luca, F.; Handoo, Z.; Di Vito, M. Morphological and molecular characterization of Pratylenchus lentis n. sp.(Nematoda: Pratylenchidae) from Sicily. J. Nematol. 2008, 40, 190. [Google Scholar]
- Waeyenberge, L.; Ryss, A.; Moens, M.; Pinochet, J.; Vrain, T. Molecular characterisation of 18 Pratylenchus species using rDNA restriction fragment length polymorphism. Nematology 2000, 2, 135–142. [Google Scholar] [CrossRef]
- de Luca, F.; Fanelli, E.; Di vito, M.; Reyes, A.; De giorg, C. Comparison of the sequences of the D3 expansion of the 26S ribosomal genes reveals different degrees of heterogeneity in different populations and species of Pratylenchus from the Mediterranean region. Eur. J. Plant Pathol. 2004, 110, 949–957. [Google Scholar] [CrossRef]
- Corbett, D. Three new species of Pratylenchus with a redescription of P. andinus Lordello, Zamith & Boock, 1961 (Nematoda: Pratylenchidae). Nematologica 1983, 29, 390–403. [Google Scholar]
- Greco, N.; Di Vito, M.; Saxena, M.; Reddy, M. Investigation on the root lesion nematode, Pratylenchus thornei, in Syria. Nematol. Mediterr. 1988, 16, 101–105. [Google Scholar]
- Saxena, M.C.; Singh, K. The Chickpea [Cicer Arietinum]; Commonwealth Agricultural Bureaux International: Wallingford, UK, 1987. [Google Scholar]
- Di Vito, M.; Greco, N.; Ores, G.; Saxena, M.; Singh, K.; Kusmenoglu, I. Plant parasitic nematodes of legumes in Turkey. Nematol. Mediterr. 1994, 22, 245–251. [Google Scholar]
- Kepenekcİ, I. Plant parasitic nematode species of Tylenchida (Nematoda) associated with sesame (Sesamum indicum L.) growing in the Mediterranean region of Turkey. Turk. J. Agric. For. 2002, 26, 323–330. [Google Scholar]
- Greco, N.; Di Vito, M. Nematodes of food legumes in the Mediterranean Basin 1. Eppo Bull. 1994, 24, 393–398. [Google Scholar] [CrossRef]
- Greco, N.; Di Vito, M.; Saxena, M. Plant parasitic nematodes of cool season food legumes in Syria. Nematol. Mediterr. 1992, 20, 37–46. [Google Scholar]
- Troccoli, A.; Di Vito, M. Root lesion and stem nematodes associated with faba bean in North Africa. Nematol. Mediterr. 2002, 30, 79–81. [Google Scholar]
- Choi, D.-R.; Lee, J.-K.; Parte, B.-Y.; Han, H.-R.; Choi, Y.-E. A new and one unrecorded species of Pratylenchus from Korea (Nematoda: Pratylenchidae). J. Asia-Pac. Entomol. 2006, 9, 5–9. [Google Scholar] [CrossRef]
- Orion, D.; Shlevin, E.; Yaniv, A. Controlling the migratory nematode Pratylenchus mediterraneus improves carrot yield quality. Hassadeh 1988, 69, 72–74. [Google Scholar]
- Development MoAaR. Database of Plant Pests in Israel: Ministry of Agriculture and Rural Development; Development MoAaR: Rishon Letzion, Israel, 2020. [Google Scholar]
- Nicol, J. The Distribution, Pathogenicity and Population Dynamics of Pratylenchus Thornei on Wheat in South Australia; Department of Crop Protection, University of Adelaide: Adelaide, Australia, 1996. [Google Scholar]
- Nicol, J.; Rivoal, R.; Taylor, S.; Zaharieva, M. (Eds.) Global importance of cyst (Heterodera spp) and lesion nematode (Pratylenchus spp.) on cereals: Distribution, yield loss, use of host resistance and integration of molecular tools. In Proceedings of the Fourth International Congress of Nematology, Tenerife, Spain, 8–13 June 2002. [Google Scholar]
- Braun Miyara, S. (Nematology Unit, Plant Protection Institute, ARO, The Volcani Center, Rishon Letzion, Israel). Personal communication, 2020.
- Kozodoi, E. (Plant Protection and Inspection Services, Rishon Letzion, Israel). Personal Communication, 2020.
- Pivonia, S. (Central-and Northern-Arava Research and Development, Arava Sapir, Israel). Personal Communication, 2020.
- Oka, Y. Nematicidal activity of fluensulfone against some migratory nematodes under laboratory conditions. Pest Manag. Sci. 2014, 70, 1850–1858. [Google Scholar] [CrossRef]
- Agrios, G.N. Introduction to Plant Pathology; Academic Press: New York, NY, USA, 1988; pp. 3–39. [Google Scholar]
- Rotenberg, D.; MacGuidwin, A.; Saeed, I.; Rouse, D. Interaction of spatially separated Pratylenchus penetrans and Verticillium dahliae on potato measured by impaired photosynthesis. Plant Pathol. 2004, 53, 294–302. [Google Scholar] [CrossRef]
- Rowe, R.C.; Powelson, M.L. Potato early dying: Management challenges in a changing production environment. Plant Dis. 2002, 86, 1184–1193. [Google Scholar] [CrossRef]
- Siti, E. The interrelationships between Pratylenchus tornei and Verticilium daliae, and their effect on potatoes. In Jerusalem HUo; Faculty of Agriculture—Hebrew University of Jerusalem: Rehovot, Israel, 1978. [Google Scholar]
- Talavera, M.; Valor, H.; Tobar, A. Post-anhydrobiotic viability of Pratylenchus thornei and Merlinius brevidens. Phytoparasitica 1998, 26, 293. [Google Scholar] [CrossRef]
- Orion, D. Nematodes of agricultural importance in Israel. Nematology 2000, 2, 735–736. [Google Scholar] [CrossRef]
- Castillo, P.; Trapero-Casas, J.; Jiménez-Díaz, R. Effect of time, temperature, and inoculum density on reproduction of Pratylenchus thornei in carrot disk cultures. J. Nematol. 1995, 27, 120. [Google Scholar]
- Thompson, J.; Clewett, T.; O’Reilly, M. Temperature response of root-lesion nematode (Pratylenchus thornei) reproduction on wheat cultivars has implications for resistance screening and wheat production. Ann. Appl. Biol. 2015, 167, 1–10. [Google Scholar] [CrossRef]
- Kurppa, S.; Vrain, T.C. Penetration and feeding behavior of Pratylenchus penetrans in strawberry roots. Rev. Nématol. 1985, 8, 273–276. [Google Scholar]
- Zunke, U. Observations on the invasion and endoparasitic behavior of the root lesion nematode Pratylenchus penetrans. J. Nematol. 1990, 22, 309. [Google Scholar] [PubMed]
- Inserra, R.; Vovlas, N. Effects of Pratylenchus vulnus on the growth of sour orange. J. Nematol. 1977, 9, 154. [Google Scholar]
- Vovlas, N.; Troccoli, A. Histopathology of broad bean roots infected by the lesion nematode Pratylenchus penetrans. Nematol. Mediterr. 1990, 18, 239–242. [Google Scholar]
- De Waele, D.; Jordaan, E. Plant-parasitic nematodes on field crops in South Africa. 1. Maize. Rev. Nématol. 1988, 11, 65–74. [Google Scholar]
- Coosemans, J. The influence of Pratylenchus penetrans on growth of Impatiens balsamina L. Daucus carota L., Linum usitatissimum L. and Crysanthemum indicum L. Symp. Int. Phytopharm Gent Belg. 1975, 27, 465–471. [Google Scholar]
- Hay, F.; Pethybridge, J. Nematodes associated with carrot production in Tasmania, Australia, and the effect of Pratylenchus crenatus on yield and quality of Kuroda-type carrot. Plant Dis. 2005, 89, 1175–1180. [Google Scholar] [CrossRef]
- Potter, J.; Olthof, T.H. Nematode pests of vegetable crops. In Plant Parasitic Nematodes in Temperate Agriculture; CABI: Wallinford, UK, 1993; pp. 171–207. [Google Scholar]
- Vrain, T.; Belair, G. Symptoms induced by the lesion nematode, Pratylenchus penetrans on carrot taproots in organic soil. Phytoprotection 1981, 62, 79–81. [Google Scholar]
- Gamliel, A. (ARO-Volcani Center, Rishon LeTsiyon, Israel). Personal communication, 2020.
- Townshend, J.; Davidson, T. Some weed hosts of Pratylenchus penetrans in premier strawberry plantations. Can. J. Bot. 1960, 38, 267–273. [Google Scholar] [CrossRef]
- Hogger, C.; Bird, G. Weed and indicator hosts of plant-parasitis nematodes in georgia cotton and soybean fields. Plant Dis. Rep. 1976, 60, 223–226. [Google Scholar]
- Koen, H. Notes on the host range, ecology and population dynamics of Pratylenchus brachyurus. Nematologica 1967, 13, 118–124. [Google Scholar] [CrossRef]
- Edwards, D.; Wehunt, E. Hosts of Pratylenchus coffeae with additions from central american banana-producing areas. Plant Dis. Rep. 1973, 57, 47. [Google Scholar]
- Ayoub, S. Pratylenchus zeae found on corn, milo, and three suspected new hosts in California. Plant Dis. Rep. 1961, 45, 940. [Google Scholar]
- Manuel, J.S.; Reynolds, D.; Bendixen, L.; Riedel, R. Weeds as Hosts of Pratylenchus; Ohio Agricultural Research and Development Center: Wooster, OH, USA, 1980. [Google Scholar]
- Vanstone, V.A.; Russ, M.H. Ability of weeds to host the root lesion nematodes Pratylenchus neglectus and P. thornei I. Grass weeds. Australas. Plant Pathol. 2001, 30, 245–250. [Google Scholar] [CrossRef]
- Katoh, K.; Standley, D.M. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A.; Hoover, P.; Rougemont, J. A rapid bootstrap algorithm for the RAxML web servers. Syst. Biol. 2008, 57, 758–771. [Google Scholar] [CrossRef]
- Ronquist, F.; Teslenko, M.; Van Der Mark, P.; Ayres, D.L.; Darling, A.; Höhna, S.; Larget, B.; Liu, L.; Suchard, M.A.; Huelsenbeck, J.P. MrBayes 3.2: Efficient Bayesian phylogenetic inference and model choice across a large model space. Syst. Biol. 2012, 61, 539–542. [Google Scholar] [CrossRef]
- Gamliel, A.; Katan, J. Control of plant disease through soil solarization. In Disease Control in Crops; Wiley-Blackwell: Hoboken, NJ, USA, 2009. [Google Scholar]
- Gamliel, A.; Katan, J. Soil Solarization: Theory and Practice; APS Press/The American Phytopathological Society: St. Paul, MN, USA, 2012. [Google Scholar]
- Katan, J. Cultural approaches for disease management: Present status and future prospects. J. Plant Pathol. 2010, 92, S7–S9. [Google Scholar]
- Katan, J.; Gamliel, A. Soilborne diseases, control by physical methods. In Encyclopedia of Agrophysics; Springer: Dordrecht, The Netherlands, 2011; pp. 813–816. [Google Scholar]
- Triky-Dotan, S.; Austerweil, M.; Steiner, B.; Peretz-Alon, Y.; Katan, J.; Gamliel, A. Accelerated degradation of metam-sodium in soil and consequences for root-disease management. Phytopathology 2009, 99, 362–368. [Google Scholar] [CrossRef]
- Di Primo, P.; Gamliel, A.; Austerweil, M.; Steiner, B.; Beniches, M.; Peretz-Alon, I.; Katan, J. Accelerated degradation of metam-sodium and dazomet in soil: Characterization and consequences for pathogen control. Crop Prot. 2003, 22, 635–646. [Google Scholar] [CrossRef]
- Triky-Dotan, S.; Austerweil, M.; Steiner, B.; Peretz-Alon, Y.; Katan, J.; Gamliel, A. Generation and dissipation of methyl isothiocyanate in soils following metam sodium fumigation: Impact on verticillium control and potato yield. Plant Dis. 2007, 91, 497–503. [Google Scholar] [CrossRef]
- Hafez, S.; Sundararaj, P. Evaluation of chemical nematicides for the suppression of Meloidogyne chitwoodi and M. hapla on potato. Int. J. Nematol. 2001, 11, 192–194. [Google Scholar]
- Ajwa, H.; Trout, T.; Mueller, J.; Wilhelm, S.; Nelson, S.; Soppe, R.; Shatley, D. Application of alternative fumigants through drip irrigation systems. Phytopathology 2002, 92, 1349–1355. [Google Scholar] [CrossRef]
- Ruzo, L.O. Physical, chemical and environmental properties of selected chemical alternatives for the pre-plant use of methyl bromide as soil fumigant. Pest Manag. Sci. Former. Pestic. Sci. 2006, 62, 99–113. [Google Scholar] [CrossRef]
- Klose, S.; Ajwa, H.A.; Browne, G.T.; Subbarao, K.V.; Martin, F.N.; Fennimore, S.A.; Westerdahl, B.B. Dose response of weed seeds, plant-parasitic nematodes, and pathogens to twelve rates of metam sodium in a California soil. Plant Dis. 2008, 92, 1537–1546. [Google Scholar] [CrossRef] [PubMed]
- Gamliel, A. Soil and substrate health. In Integrated Pest and Disease Management in Greenhouse Crops; Springer: Dordrecht, The Netherlands, 2020; pp. 355–383. [Google Scholar]
- Talavera, M.; Miranda, L.; Gómez-Mora, J.A.; Vela, M.D.; Verdejo-Lucas, S. Nematode Management in the Strawberry Fields of Southern Spain. Agronomy 2019, 9, 252. [Google Scholar] [CrossRef]
- Oka, Y.; Pivonia, S. Use of ammonia-releasing compounds for control of the root-knot nematode Meloidogyne javanica. Nematology 2002, 4, 65–71. [Google Scholar] [CrossRef]
- Rodriguez-Kabana, R.; Morgan-Jones, G.; Chet, I. Biological control of nematodes: Soil amendments and microbial antagonists. Plant Soil 1987, 100, 237–247. [Google Scholar] [CrossRef]
- Gamliel, A.; Stapleton, J. Effect of chicken compost or ammonium phosphate and solarization on pathogen control, rhizosphere microorganisms, and lettuce growth. Plant Dis. 1993, 77, 886–891. [Google Scholar] [CrossRef]
- Oka, Y.; Tkachi, N.; Shuker, S.; Yerumiyahu, U. Enhanced nematicidal activity of organic and inorganic ammonia-releasing amendments by Azadirachta indica extracts. J. Nematol. 2007, 39, 9. [Google Scholar]
- Eleckcioglu, I.; Avci, M.; Nicol, J.; Meyveci, K.; Bolat, N.; Yorgancilar, A.; Sahim, E.; Kaplan, A. The Use of Crop Rotation as a Means to Control the Cyst and Lesion Nematode Under Rainfed Wheat Production Systems. In Proceedings of the Plant Protection Congress of Turkey, Samsun, Turkey, 8–10 September 2004. [Google Scholar]
- Storey, R.; Glazer, I.; Orion, D. Lipid Utilisation By Starved and Anhydrobiotic Individuals of Pratylenchus Thornei. Nematologica 1982, 28, 373–378. [Google Scholar]
- Abdel-Rahman, F.H.; Alaniz, N.M.; Saleh, M.A. Nematicidal activity of terpenoids. J. Environ. Sci. Health Part B 2013, 48, 16–22. [Google Scholar] [CrossRef]
- Thompson, J.; Brennan, P.; Clewett, T.; Sheedy, J.; Seymour, N. Progress in breeding wheat for tolerance and resistance to root-lesion nematode (Pratylenchus thornei). Australas. Plant Pathol. 1999, 28, 45–52. [Google Scholar] [CrossRef]
- Talavera, M.; Van stone, V.A. Monitoring Pratylenchus thornei densities in SOCL and roots under resistant (Triticum turgidum durum) and susceptible (Triticum aestivum) wheat cultivars. Phytoparasitica 2001, 29, 29–35. [Google Scholar] [CrossRef]
- Farsi, M. Genetic Variation for Tolerance and Resistance to Pratylenchusneglectus/by Mohammed Farsi. Ph.D. Thesis, University of Adelaide, Dept. of Plant Science, Adelaide, Australia, 1996. [Google Scholar]
- Linsell, K.J.; Riley, I.T.; Davies, K.A.; Oldach, K.H. Characterization of resistance to Pratylenchus thornei (Nematoda) in wheat (Triticum aestivum): Attraction, penetration, motility, and reproduction. Phytopathology 2014, 104, 174–187. [Google Scholar] [CrossRef]
- Leibson, T.; Lifshitz, M. Organophosphate and carbamate poisoning: Review of the current literature and summary of clinical and laboratory experience in southern Israel. Isr. Med. Assoc. J. 2008, 10, 767. [Google Scholar]
- Hague, N.; Gowen, S. Chemical control of nematodes. In Principles and Practice of Nematode Control in Crops; Brown, H., Kerry, B., Eds.; Academic Press: New York, NY, USA; Sydney, Australia, 1987; pp. 131–178. [Google Scholar]
- Oka, Y. Nematicidal activity of fluensulfone compared to that of organophosphate and carbamate nematicides against Xiphinema index and Longidorus vineacola. Eur. J. Plant Pathol. 2019, 154, 565–574. [Google Scholar] [CrossRef]
- Al-Banna, L.; Williamson, V.; Gardner, S.L. Phylogenetic Analysis of Nematodes of the GenusPratylenchusUsing Nuclear 26S rDNA. Mol. Phylogenet. Evol. 1997, 7, 94–102. [Google Scholar] [CrossRef]
- Andrés, M.; Pinochet, J.; Hernández-Dorrego, A.; Delibes, A. Detection and analysis of inter-and intraspecific diversity of Pratylenchus spp. using isozyme markers. Plant Pathol. 2000, 49, 640–649. [Google Scholar] [CrossRef]
- Duncan, L.; Inserra, R.; Thomas, W.; Dunn, D.; Mustika, I.; Frisse, L.; Mendes, M.; Morris, K.; Kaplan, D. Molecular and morphological analysis of isolates of Pratylenchus coffeae and closely related species. Nematropica 1999, 29, 61–80. [Google Scholar]
- Loof, P.A. Taxonomic studies on the genus Pratylenchus (Nematoda). Tijdschr. Over Plantenziekten 1960, 66, 29–90. [Google Scholar]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Natural Vegetation | |||||
|---|---|---|---|---|---|
| Fruit Trees (n = 34) | Forest Trees (n = 20) | Herbaceous Plants (n = 12) | |||
| Tylenchorhynchus | 65 | Tylenchus | 70 | Helicotylenchus | 58 |
| Xiphinema | 62 | Xiphinema | 65 | Tylenchus | 58 |
| Helicotylenchus | 53 | Helicotylenchus | 60 | Meloidogyne | 50 |
| Tylenchus | 50 | Tylenchorhynchus | 40 | Tylenchorhynchus | 42 |
| Pratylenchus | 35 | Pratylenchus | 30 | Xiphinema | 33 |
| Meloidogyne | 29 | Rotylenchulus | 20 | ||
| Rotylenchulus | 21 | ||||
| Cultivated crops | |||||
| Deciduous Fruit Trees (n = 38) | Subtropical and Tropical Fruit Trees (n = 20) | Grapevines (n = 17) | |||
| Xiphinema | 76 | Xiphinema | 60 | Helicotylenchus | 65 |
| Tylenchorhynchus | 58 | Tylenchorhynchus | 55 | Xiphinema | 59 |
| Pratylenchus | 47 | Helicotylenchus | 45 | Tylenchorhynchus | 53 |
| Helicotylenchus | 38 | Tylenchus | 25 | Longidorus | 47 |
| Pratylenchus | 20 | Meloidogyne | 47 | ||
| Criconemoides | 20 | Pratylenchus | 29 | ||
| Vegetable crops (n = 41) | Cereal crops and pasture grasses (n = 50) | Pasture Legumes (n = 27) | |||
| Tylenchorhynchus | 59 | Tylenchorhynchus | 84 | Tylenchorhynchus | 85 |
| Pratylenchus | 49 | Pratylenchus | 68 | Tylenchus | 52 |
| Tylenchus | 39 | Tylenchus | 58 | Pratylenchus | 48 |
| Helicotylenchus | 27 | Helicotylenchus | 26 | Helicotylenchus | 30 |
| Longidorus | 22 | Xiphinema | 24 | Ditylenchus | 20 |
| Lawns (n = 11) | |||||
| Helicotylenchus | 82 | Criconemoides | 36 | Pratylenchus | 27 |
| Tylenchorhynchus | 64 | Xiphinema | 36 | ||
| Tylenchus | 55 | Trichodorus | 27 | ||
| Cultivated Crops | |||||
|---|---|---|---|---|---|
| Grapevines | |||||
| Helicotylenchus pseudorobustus | Tomer | ||||
| Xiphinema index | Tomer | ||||
| Aphelenchoides sp. | Tomer | ||||
| Vegetable Crops | Cereal Crops and Pasture Grasses | Pasture Legumes | |||
| Pratylenchus mediterraneus | Kfar Silver | Pratylenchus thornei | Gevim | Pratylenchus thornei | Shif’a Gevim, NirOz |
| Pratylenchus nanus | Shoval | Merlinius nanus | Sde Eliyahu, Tirat Zvi | Merlinius nanus | Shif’a |
| Pratylenchus thornei | Shoval, Alumim | Heterodera avenae | Nirim | Tylenchorhynchus clarus | Nir David |
| Neodolichorhynchus sulcatus | Arava | Heterodera sp. | Nirim | Rotylenchus macrosoma | Shif’a |
| Pratylenchus capsici | Arava | Geocenamus brevidens | Nirim | Tylenchorhynchus zeae | Shif’a |
| Meloidogyne javanica | Mivtachim | ||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Bucki, P.; Qing, X.; Castillo, P.; Gamliel, A.; Dobrinin, S.; Alon, T.; Braun Miyara, S. The Genus Pratylenchus (Nematoda: Pratylenchidae) in Israel: From Taxonomy to Control Practices. Plants 2020, 9, 1475. https://doi.org/10.3390/plants9111475
Bucki P, Qing X, Castillo P, Gamliel A, Dobrinin S, Alon T, Braun Miyara S. The Genus Pratylenchus (Nematoda: Pratylenchidae) in Israel: From Taxonomy to Control Practices. Plants. 2020; 9(11):1475. https://doi.org/10.3390/plants9111475
Chicago/Turabian StyleBucki, Patricia, Xue Qing, Pablo Castillo, Abraham Gamliel, Svetlana Dobrinin, Tamar Alon, and Sigal Braun Miyara. 2020. "The Genus Pratylenchus (Nematoda: Pratylenchidae) in Israel: From Taxonomy to Control Practices" Plants 9, no. 11: 1475. https://doi.org/10.3390/plants9111475
APA StyleBucki, P., Qing, X., Castillo, P., Gamliel, A., Dobrinin, S., Alon, T., & Braun Miyara, S. (2020). The Genus Pratylenchus (Nematoda: Pratylenchidae) in Israel: From Taxonomy to Control Practices. Plants, 9(11), 1475. https://doi.org/10.3390/plants9111475