Inhibition of Digestive Enzymes and Antioxidant Activity of Extracts from Fruits of Cornus alba, Cornus sanguinea subsp. hungarica and Cornus florida–A Comparative Study



Abstract
1. Introduction
2. Results
2.1. Total Phenols Content
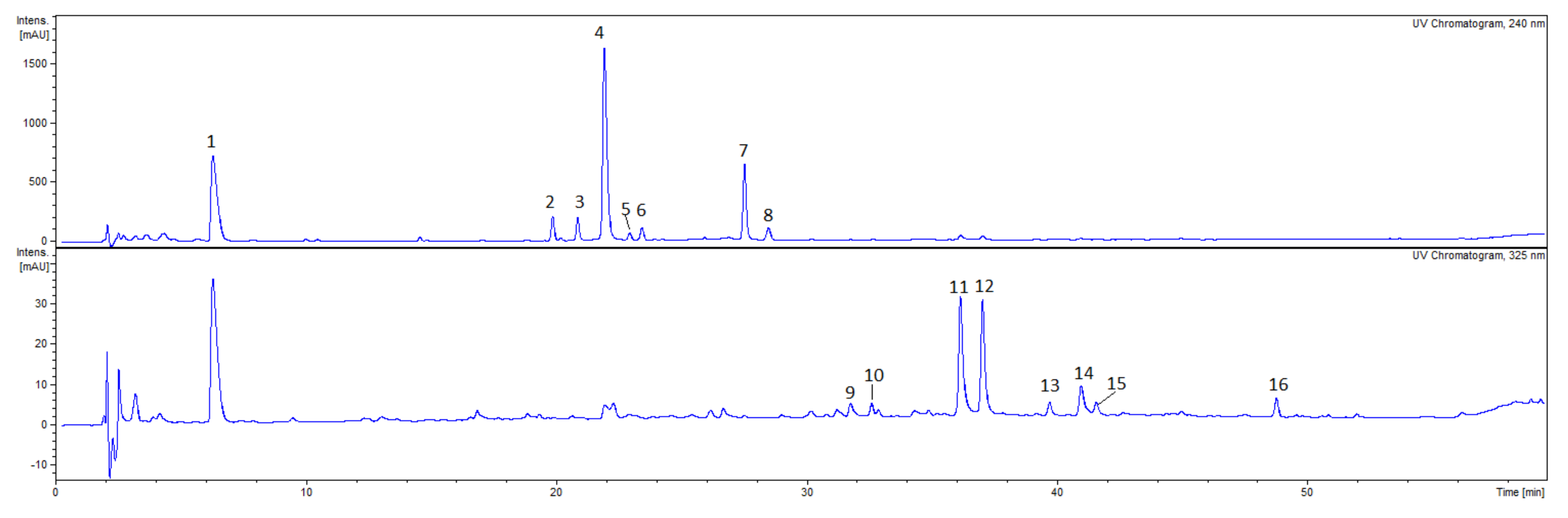
2.2. Phytochemical Analysis of Extracts
2.3. Antioxidant Activity
2.4. Digestive Enzymes Inhibition
3. Discussion
4. Materials and Methods
4.1. General Experimental Procedures
4.2. Plant Material and Extracts Preparation
4.3. Phytochemical Analysis
4.3.1. HPLC-DAD-MS/MS
4.3.2. Total Polyphenols Content
4.3.3. Isolation of Compounds from CA Fruits
4.4. Biological Experiments
4.4.1. DPPH Assay
4.4.2. α-Glucosidase Inhibition
4.4.3. α-Amylase Inhibition
4.4.4. Pancreatic Lipase Activity
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Xiang, Q.Y.; Brunsfeld, S.J.; Soltis, D.E.; Soltis, P.S. Phylogenetic relationships in Cornus based on chloroplast DNA restriction sites: Implications for biogeography and character evolution. Syst. Bot. 1996, 21, 515–534. [Google Scholar] [CrossRef]
- Fan, C.Z.; Xiang, Q.Y. Phylogenetic relationships within Cornus (Cornaceae) based on 26S rDNA sequences. Am. J. Bot. 2001, 88, 1131–1138. [Google Scholar] [CrossRef]
- Czerwińska, M.E.; Melzig, M.F. Cornus mas and Cornus officinalis-analogies and differences of two medicinal plants traditionally used. Front. Pharmacol. 2018, 9, 894. [Google Scholar] [CrossRef] [PubMed]
- Bate-Smith, E.; Ferguson, I.; Hutson, K.; Jensen, S.; Nielsen, B.; Swain, T. Phytochemical interrelationships in the Cornaceae. Biochem. Sys. Ecol. 1975, 3, 79–89. [Google Scholar] [CrossRef]
- Jensen, S.; Kjaer, A.; Nielsen, B. The genus Cornus: Non-flavonoid glucosides as taxonomic markers. Biochem. Sys. Ecol. 1975, 3, 75–78. [Google Scholar] [CrossRef]
- Asgary, S.; Rafieian-Kopaei, M.; Shamsi, F.; Najafi, S.; Sahebkar, A. Biochemical and histopathological study of the anti-hyperglycemic and anti-hyperlipidemic effects of cornelian cherry (Cornus mas L.) in alloxan-induced diabetic rats. J. Complement. Integr. Med. 2014, 11, 63–69. [Google Scholar] [CrossRef] [PubMed]
- Park, C.H.; Noh, J.S.; Tanaka, T.; Uebaba, K.; Cho, E.J.; Yokozawa, T. The effects of Corni fructus extract and its fractions against α-glucosidase inhibitory activities in vitro and sucrose tolerance in normal rats. Am. J. Chin. Med. 2011, 39, 367–380. [Google Scholar] [CrossRef] [PubMed]
- Shishehbor, F.; Azemi, M.E.; Zameni, D.; Saki, A. Inhibitory effects of hydroalcoholic extracts of barberry, sour cherry and Cornelian cherry on α-amylase and α-glucosidase activities. Int. J. Pharm. Res. Allied Sci. 2016, 5, 423–428. [Google Scholar]
- Pyrkosz-Biardzka, K.; Kucharska, A.Z.; Sokol-Letowska, A.; Strugala, P.; Gabrielska, J. A comprehensive study on antioxidant properties of crude extracts from fruits of Berberis vulgaris L., Cornus mas L. and Mahonia aquifolium Nutt. Pol. J. Food Nutr. Sci. 2014, 64, 91–99. [Google Scholar] [CrossRef]
- Asgary, S.; Rafieian-Kopaei, M.; Adelnia, A.; Kazemi, S.; Shamsi, F. Comparing the effects of lovastatin and cornus MAS fruit on fibrinogen level in hypercholesterolemic rabbits. ARYA Atheroscler. 2010, 6, 1–5. [Google Scholar] [PubMed]
- Jayaprakasam, B.; Vareed, S.K.; Olson, L.K.; Nair, M.G. Insulin secretion by bioactive anthocyanins and anthocyanidins present in fruits. J. Agric. Food Chem. 2005, 53, 28–31. [Google Scholar] [CrossRef] [PubMed]
- Neha, K.; Haider, M.R.; Pathak, A.; Yar, M.S. Medicinal prospects of antioxidants: A review. Eur. J. Med. Chem. 2019, 178, 687–704. [Google Scholar] [CrossRef] [PubMed]
- Świerczewska, A.; Buchholz, T.; Melzig, M.F.; Czerwińska, M.E. In vitro α-amylase and pancreatic lipase inhibitory activity of Cornus mas L. and Cornus alba L. fruit extracts. J. Food Drug Anal. 2019, 27, 249–258. [Google Scholar] [CrossRef] [PubMed]
- Park, K.H.; Yin, J.; Yoon, K.H.; Hwang, Y.J.; Lee, M.W. Antiproliferative effects of new dimeric ellagitannin from Cornus alba in prostate cancer cells including apoptosis-related S-phase arrest. Molecules 2016, 21, 137. [Google Scholar] [CrossRef]
- Bujor, A.; Miron, A.; Luca, S.V.; Skalicka-Wozniak, K.; Silion, M.; Ancuceanu, R.; Dinu, M.; Girard, C.; Demougeot, C.; Totoson, P. Metabolite profiling, arginase inhibition and vasorelaxant activity of Cornus mas, Sorbus aucuparia and Viburnum opulus fruit extracts. Food Chem. Toxicol. 2019, 133, 110764. [Google Scholar] [CrossRef]
- Lu, Y.R.; Sun, Y.; Foo, L.Y.; McNabb, W.C.; Molan, A.L. Phenolic glycosides of forage legume Onobrychis viciifolia. Phytochemistry 2000, 55, 67–75. [Google Scholar] [CrossRef]
- Nakamura, S.; Fujimoto, K.; Matsumoto, T.; Nakashima, S.; Ohta, T.; Ogawa, K.; Matsuda, H.; Yoshikawa, M. Acylated sucroses and acylated quinic acids analogs from the flower buds of Prunus mume and their inhibitory effect on melanogenesis. Phytochemistry 2013, 92, 128–136. [Google Scholar] [CrossRef]
- Nakamura, K.; Ogasawara, Y.; Endou, K.; Fujimori, S.; Koyama, M.; Akano, H. Phenolic compounds responsible for the superoxide dismutase-like activity in high-brix apple vinegar. J. Agric. Food Chem. 2010, 58, 10124–10132. [Google Scholar] [CrossRef]
- Pacifico, S.; D’Abrosca, B.; Scognamiglio, M.; Gallicchio, M.; Galasso, S.; Monaco, P.; Fiorentino, A. Antioxidant polyphenolic constituents of Vitis × labruscana cv. ‘Isabella’ Leaves. Open Nat. Prod. J. 2013, 6, 5–11. [Google Scholar] [CrossRef][Green Version]
- Romero, C.; Brenes, M.; García, P.; Garrido, A. Hydroxytyrosol 4-β-D-glucoside, an important phenolic compound in olive fruits and derived products. J. Agric. Food Chem. 2002, 50, 3835–3839. [Google Scholar] [CrossRef]
- Amiri, A.; Azemi, M.E.; Khodayar, M.J.; Namjoyan, F. In vitro α-amylase and α-glucosidases inhibitory effects of some plant extracts. Int. J. Pharmacognos. Phyochem. Res. 2015, 7, 315–318. [Google Scholar]
- Gholamhoseinian, A.; Shahouzehi, B.; Sharifi-far, F. Inhibitory effect of some plant extracts on pancreatic lipase. Int. J. Pharmacol. 2010, 6, 18–24. [Google Scholar]
- Wang, H.H.; Wang, Z.Z.; Guo, W.B. Comparative determination of ursolic acid and oleanolic acid of Macrocarpium officinalis (Sieb. et Zucc.) Nakai by RP-HPLC. Ind. Crop Prod. 2008, 28, 328–332. [Google Scholar] [CrossRef]
- Yousfbeyk, F.; Esmaiili, T.; Pashna, Z.; Hozori, Z.; Ghohari, A.R.; Ostad, S.N.; Amin, G.R. Antioxidant activity, total phenol and total anthocyanin contents of Cornus sanguinea L. subsp. australis (C.A. Mey.) Jáv. J. Med. Plants 2014, 13, 69–74. [Google Scholar]
- Stanković, M.S.; Topuzović, M.D. In vitro antioxidant activity of extracts from leaves and fruits of common dogwood (Cornus sanguinea L.). Acta Botanica Gallica 2012, 159, 79–83. [Google Scholar] [CrossRef][Green Version]
- Park, H.M.; Hong, J.H. Antioxidant effects of bioactive compounds isolated from pressurized steam-treated Corni fructus and their protective effects against UVB-irradiated HS68 cells. J. Med. Food 2018, 21, 1165–1172. [Google Scholar] [CrossRef]
- Du, C.T.; Wang, P.L.; Francis, F.J. Anthocyanins of Cornaceae, Cornus canadensis. Phytochemistry 1974, 13, 2002. [Google Scholar] [CrossRef]
- Du, C.T.; Wang, P.L.; Francis, F.J. Anthocyanins of Cornus alternifolia, L. and C. alba L. HortScience 1975, 10, 36–37. [Google Scholar]
- Du, C.T.; Wang, P.L. Anthocyanins of Cornaceae, Cornus kousa Hance and Cornus florida L. HortScience 1974, 9, 243–244. [Google Scholar]
- Du, C.T.; Francis, F.J. Anthocyanins from Cornus mas. Phytochemistry 1973, 12, 2487–2489. [Google Scholar] [CrossRef]
- Vareed, S.K.; Reddy, M.K.; Schutzki, R.E.; Nair, M.G. Anthocyanins in Cornus alternifolia, and Cornus controversa, Cornus kousa and Cornus florida fruits with health benefits. Life Sci. 2006, 78, 777–784. [Google Scholar] [CrossRef] [PubMed]
- Bjorøy, O.; Fossen, T.; Andersen, O. Anthocyanin 3-galactosides from Cornus alba ’Sibirica’ with glucosidation of the B-ring. Phytochemistry 2007, 68, 640–645. [Google Scholar] [CrossRef]
- Seeram, N.P.; Schutzki, R.; Chandra, A.; Nair, M.G. Characterization, quantification and bioactivities of anthocyanins in Cornus species. J. Agric. Food Chem. 2002, 50, 2519–2523. [Google Scholar] [CrossRef] [PubMed]
- Tan, Y.Q.; Chang, S.K.C. Digestive enzyme inhibition activity of the phenolic substances in selected fruits, vegetables and tea as compared to black legumes. J. Funct. Foods 2017, 38, 644–655. [Google Scholar] [CrossRef]
- Rutkowski, L. Klucz do Oznaczania Roślin Naczyniowych Polski Niżowej, 2nd ed.; Wydawnictwo Naukowe PWN: Warszawa, Poland, 2011. [Google Scholar]
- Singleton, V.L.; Orthofer, R.; Lamuela-Raventós, R.M. Analysis of total phenols and other oxidation substrates and antioxidants by means of Folin-Ciocalteu reagent. Methods Enzymol. 1999, 299, 152–178. [Google Scholar]
- Czerwińska, M.E.; Duszak, K.; Parzonko, A.; Kiss, A.K. Chemical composition and UVA-protecting activity of extracts from Ligustrum vulgare and Olea europaea leaves. Acta Biol. Cracov. Bot. 2016, 58, 45–55. [Google Scholar] [CrossRef][Green Version]
- Choi, C.W.; Kim, S.C.; Hwang, S.S.; Choi, B.K.; Ahn, H.J.; Lee, M.Y.; Park, S.H.; Kim, S.K. Antioxidant activity and free radical scavenging capacity between Korean medicinal plants and flavonoids by assay-guided comparison. Plant Sci. 2002, 163, 1161–1168. [Google Scholar] [CrossRef]
- Ma, L.F.; Yan, J.J.; Lane, H.Y.; Jin, L.C.; Qiu, F.J.; Wang, Y.J.; Xi, Z.F.; Shan, W.G.; Zhan, Z.J.; Ying, Y.M. Bioassay-guided isolation of lanostane-type triterpenoids as alpha-glucosidase inhibitors from Ganoderma hainanense. Phytochem. Lett. 2019, 29, 154–159. [Google Scholar] [CrossRef]
- Buchholz, T.; Melzig, M.F. Medicinal plants traditionally used for treatment of obesity and diabetes mellitus—Screening for pancreatic lipase and α-amylase inhibition. Phytother. Res. 2016, 30, 260–266. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| No. | Tentative Assignement | Rt [min] | UV [nm] | [M-H]− m/z | MS2 Ions m/z |
|---|---|---|---|---|---|
| CA1 | Unidentified | 3.2 | 226 | 491 | 431, 373 |
| CA2 | Unidentified | 4.8 | 217 | 689 | 527, 515, 481, 353, 172 |
| CA3 | Hydroxytyrosol glucoside | 5.3 | 290 | 631 * | 315 |
| CA4 | Unidentified | 8.5 | 219 | 665 | 491, 373 |
| CA5 | Unidentified | 13.6 | 290 | 489 | 314, 173 |
| CA6 | Unidentified | 20.6 | 290 | 663 | 616, 521, 489, 405, 329 |
| CA7 | Coumaroylquinic acid Tetragalloylglucose | 24.3 24.3 | 290, 310sh overlapped | 337 787 | 191, 163, 119 453, 315 |
| CA8 | Quercetin-3-O-(2″-O-galloyl)-hexoside | 33.7 | 270, 360 | 615 | 463, 301 |
| CA9 | Quercetin 3-O-rhamnoglucoside | 35.2 | 270, 350 | 609 | 463, 301 |
| CA10 | Quercetin 3-O-galactoside | 36.2 | 260, 353 | 463 | 301 |
| CA11 | Kaempferol derivative Quercetin hexoside | 37.0 overlapped | 260, 352 | 599 463 | 447, 313, 285, 169 301 |
| CA12 | Quercetin 3-O-glucuronide | 38.0 | 255, 352 | 477 | 301 |
| CA13 | Kaempferol hexoside | 39.7 | 264, 349 | 447 | 285 |
| CA14 | Quercetin (6″-O-malonyl)-3-O-β-d-glucoside | 41.2 | 265, 353 | 549 | 505, 463, 343, 301 |
| CA15 | Kaempferol 3-O-glucoside | 41.6 | 264, 344 | 447 | 327, 285 |
| CA16 | Kaempferol 3-O-glucuronide | 42.5 | 264, 344 | 461 | 285, 175 |
| CA17 | Kaempferol malonyl-hexoside | 44.6 | 264, 344 | 533 | 489, 447, 285 |
| CA18 | Kaempferol malonyl-hexoside | 46.4 | 264, 350 | 533 | 489, 285 |
| No. | Tentative Assignment | Rt [min] | UV [nm] | [M-H]− m/z | MS2 m/z |
|---|---|---|---|---|---|
| CS1 | Unidentified | 3.1 | 230 | 631 | 315, 243, 161 |
| CS2 | Unidentified | 4.8 | 236, 315 | 523 | 505, 477, 387, 315, 232, 179 |
| CS3 | Quercetin tri-hexoside | 17.1 | 262, 340 | 787 | 625, 463, 301 |
| CS4 | Quercetin tri-hexoside | 18.2 | 262, 342 | 787 | 625, 463, 301 |
| CS5 | Myricetin tri-hexoside Quercetin-dipentosyl-hexoside | 19.0 overlapped | 263, 342 | 803 757 | 639, 479, 317 625, 595, 463, 301 |
| CS6 | Myricetin tri-hexoside | 19.9 | 264, 330 | 803 | 641, 479, 317 |
| CS7 | Myricetin tri-hexoside Quercetin dihexoside | 22.3 overlapped | 256, 360 | 803 625 | 641, 479, 317 463, 301 |
| CS8 | Isorhamnetin tri-hexoside | 26.8 | 270, 350 | 639 | 477, 315 |
| CS9 | Myricetin dihexoside Quercetin dihexoside | 28.0 overlapped | 250, 360 | 641 625 | 479, 317 463, 301 |
| CS10 | Quercetin dihexoside | 29.0 | 270, 360 | 625 | 463, 301 |
| CS11 | Myricetin dihexoside Quercetin dihexoside | 30.0 overlapped | 264, 342 | 641 625 | 479, 317 463, 301 |
| CS12 | Myrcetin dihexoside | 31.4 | 270, 340 | 641 | 479, 317, 303 |
| CS13 | Myricetin dihexoside Isorhamnetin tri-hexoside | 32.2 overlapped | 260, 340 | 641 639 | 479, 317, 303 477, 396, 272 |
| CS14 | Quercetin dihexoside Quercetin 6-O-α-l-arabinosyl-d-glucoside | 32.8 overlapped | 260, 350 | 625 595 | 463, 301 475, 463, 449, 300, 179 |
| CS15 | Quercetin 3-O-rhamnoglucoside | 36.1 | 260, 352 | 609 | 301 |
| CS16 | Quercetin hexoside | 37.0 | 255, 353 | 463 | 301 |
| CS17 | Quercetin 3-O-glucuronide | 38.0 | 255, 353 | 477 | 301 |
| CS18 | Quercetin hexoside | 42.8 | 260, 350 | 463 | 301 |
| No. | Tentative Assignment | Rt [min] | UV [nm] | [M-H]− m/z | MS2 Ions m/z |
|---|---|---|---|---|---|
| CF1 | Gallic acid glucuronide | 6.3 | 254, 297 | 345 | 169, 123 |
| CF2 | Loganic acid | 19.9 | 233 | 375 | 213, 169 |
| CF3 | Unidentified | 20.9 | 236 | 595 # | 549, 341, 281, 225 |
| CF4 | Dehydrologanin derivative | 22.0 | 238 | 433 # | 387, 225, 179 |
| CF5 | Dehydrologanin derivative | 22.9 | 223, 280 | 433 # | 387, 225, 179, 149 |
| CF6 | Loganin | 23.4 | 234, 280 | 435 # | 389, 227 |
| CF7 | 7-epi-loganin | 27.4 | 232 | 435 # | 389, 227 |
| CF8 | Unidentified | 28.5 | 235 | 539 | 491, 343, 195 |
| CF9 | Quercetin-hexose protocatechuic acid Myricetin hexoside | 31.3 overlapped | 280, 350sh | 599 479 | 483, 465, 405, 301 317, 179 |
| CF10 | Quercetin 6-O-α-l-arabinosyl-d-glucoside | 32.5 | 275, 360 | 595 | 463, 301, 179 |
| CF11 | Tetragalloylglucose Quercetin 3-O-galactoside | 36.1 overlapped | 260, 360 | 787 463 | 635, 465, 319, 249 301 |
| CF12 | Quercetin hexoside | 37.0 | 260, 352 | 463 | 301 |
| CF13 | Tetragalloylglucose Kaempferol hexoside | 39.7 overlapped | 265, 353 | 787 447 | 633, 573, 465, 379, 249 327, 284, 151 |
| CF14 | Quercetin (6″-O-malonyl)-3-O-β-d-glucoside | 41.2 | 280, 360 | 549 | 505 |
| CF15 | Unidentified Kaempferol hexoside | 41.5 overlapped | 280 | 615 447 | 567, 419, 223, 195, 177 285 |
| CF16 | Unidentified Unidentified | 48.8 overlapped | 280 | 533 712 | 307, 225, 163 666, 605, 504, 442, 377, 322 |
| Aqueous-Ethanolic Extract | DPPH Scavenging (µg/mL) |
|---|---|
| Cornus alba | 91.47 ± 3.66 a,b |
| Cornus florida | 17.10 ± 6.89 |
| Cornus sanguinea | 130.03 ± 31.61 a,b |
| Ascorbic acid | 3.73 ± 0.83 |
| Cornus alba | Cornus florida | Cornus sanguinea | |
|---|---|---|---|
| α -glucosidase | |||
| Aqueous-ethanolic extract | 50.38 ± 14.05 | 38.87 ± 2.65 | 70.07 ± 16.62 † |
| Acarbose | 150.43 ± 0.04 *,# 231.91 ± 0.06 (µM) | ||
| α-amylase | |||
| Aqueous-ethanolic extract | 115.20 ± 14.31 a,c | 5018.43 ± 14.70 b,c | 651.44 ± 12.99 a,b |
| Acarbose | 2.4 ± 0.06 § 3.7 ± 0.6 (µM) | ||
| No. | Fraction | UV Max | [M-H]− m/z | MS2 Fragment Ions (m/z) | Identified Compound | IC50 [mM] | References |
|---|---|---|---|---|---|---|---|
| 1 | D2.1 | 217, 324 | 353 | 191 | chlorogenic acid | 0.80 ± 0.32 | [18] |
| 2 | D2.3 | 223, 311 | 675 * | 337, 191 | 5-O-coumaroylquinic acid | 0.84 ± 0.14 | [16] |
| 3 | D2.4 | 226, 312 | 351 | 163, 145, 119 | 5-O-(E)-p-coumaroylquinic acid methyl ester | 0.16 ± 0.01 | [17] |
| 4 | D2.5 | 265, 344 | 475 | 327, 299, 285, 255, 227 | kaempferol 3-O-glucuronide 6″-methylester | 0.21 ± 0.05 | [19] |
| 5 | H1.1 | 196, 290 | 631 * | 315, 179, 143 | hydroxytyrosol glucoside | 1.46 ± 0.18 | [20] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Truba, J.; Stanisławska, I.; Walasek, M.; Wieczorkowska, W.; Woliński, K.; Buchholz, T.; Melzig, M.F.; Czerwińska, M.E. Inhibition of Digestive Enzymes and Antioxidant Activity of Extracts from Fruits of Cornus alba, Cornus sanguinea subsp. hungarica and Cornus florida–A Comparative Study. Plants 2020, 9, 122. https://doi.org/10.3390/plants9010122
Truba J, Stanisławska I, Walasek M, Wieczorkowska W, Woliński K, Buchholz T, Melzig MF, Czerwińska ME. Inhibition of Digestive Enzymes and Antioxidant Activity of Extracts from Fruits of Cornus alba, Cornus sanguinea subsp. hungarica and Cornus florida–A Comparative Study. Plants. 2020; 9(1):122. https://doi.org/10.3390/plants9010122
Chicago/Turabian StyleTruba, Joanna, Iwona Stanisławska, Marta Walasek, Wioleta Wieczorkowska, Konrad Woliński, Tina Buchholz, Matthias F. Melzig, and Monika E. Czerwińska. 2020. "Inhibition of Digestive Enzymes and Antioxidant Activity of Extracts from Fruits of Cornus alba, Cornus sanguinea subsp. hungarica and Cornus florida–A Comparative Study" Plants 9, no. 1: 122. https://doi.org/10.3390/plants9010122
APA StyleTruba, J., Stanisławska, I., Walasek, M., Wieczorkowska, W., Woliński, K., Buchholz, T., Melzig, M. F., & Czerwińska, M. E. (2020). Inhibition of Digestive Enzymes and Antioxidant Activity of Extracts from Fruits of Cornus alba, Cornus sanguinea subsp. hungarica and Cornus florida–A Comparative Study. Plants, 9(1), 122. https://doi.org/10.3390/plants9010122