Drought Yield QTL (qDTY) with Consistent Effects on Morphological and Agronomical Traits of Two Populations of New Rice (Oryza sativa) Lines

 ,
, 
 and
and
Abstract
1. Introduction
2. Results
2.1. Drought Imposition
2.2. Assortment of PLs into qDTY Classes
2.3. Effects of qDTY Classes
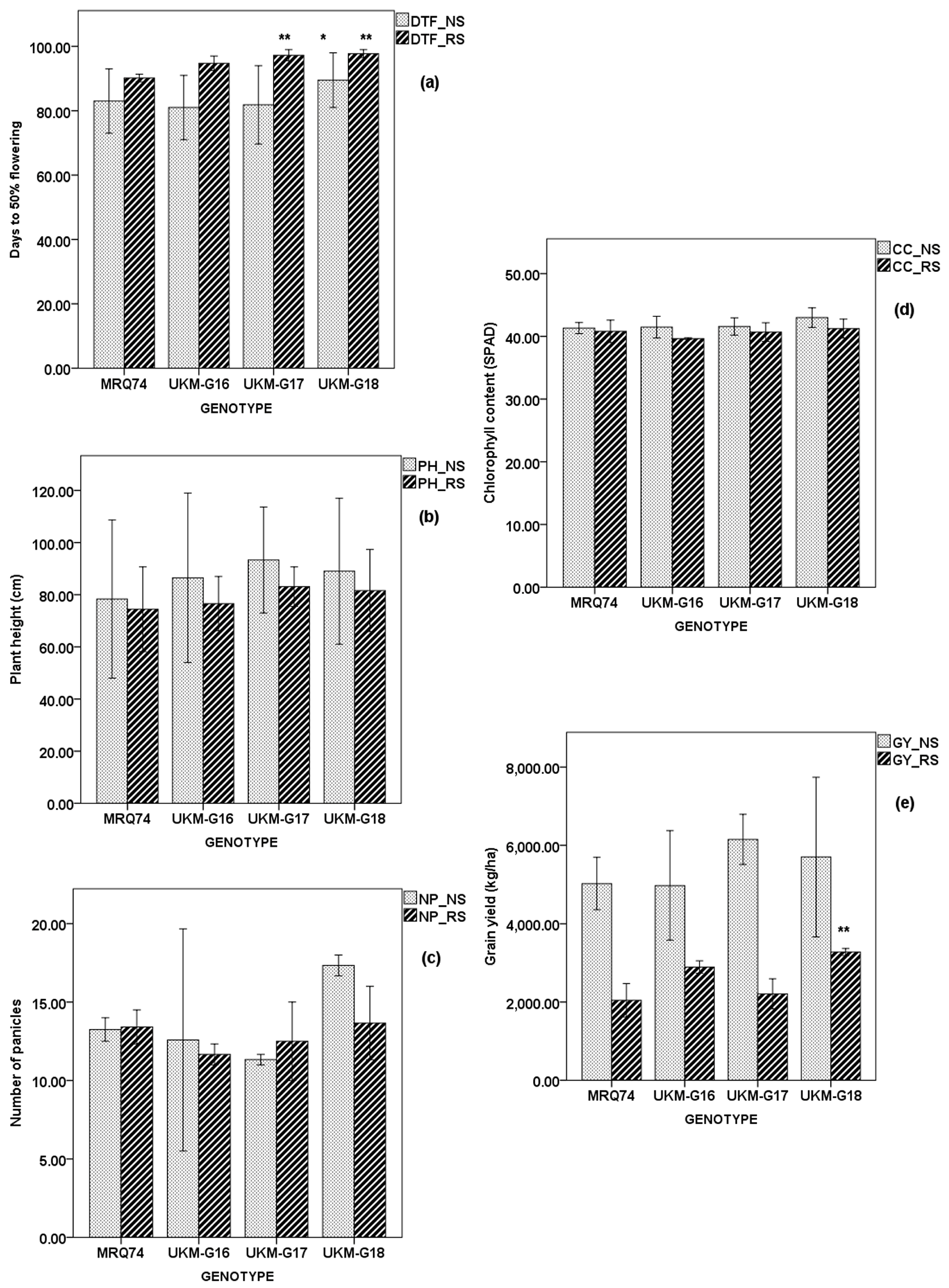
2.4. Performance of Selected MRQ74 PLs and MR219 PLs
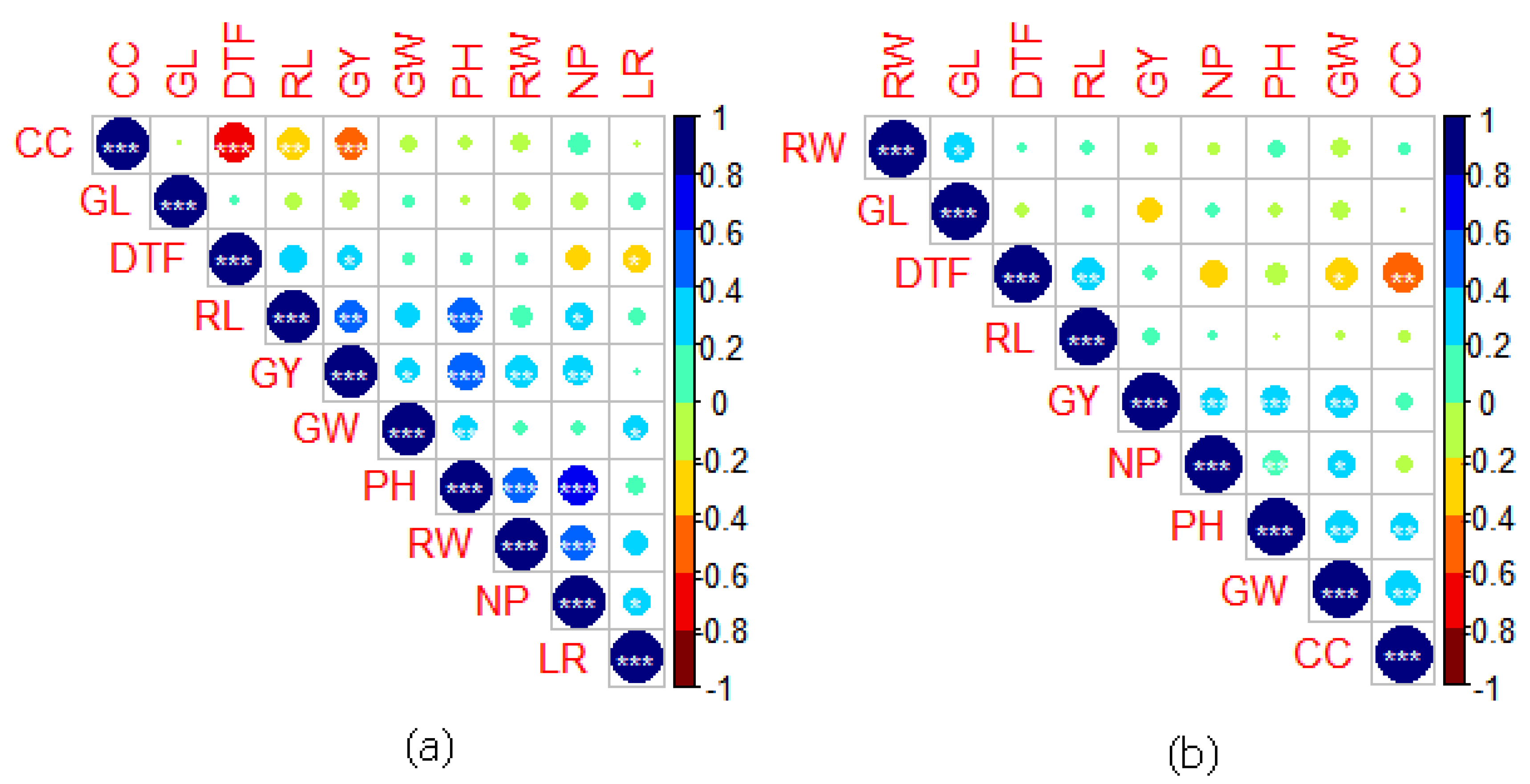
2.5. Correlation between Traits under NS and RS
2.6. Heritability of Traits under NS and RS
3. Discussion
4. Materials and Methods
4.1. Plant Materials
4.2. RS and NS Preparation and Experimental Design
4.3. Collection and Analysis of Morphological and Agronomical Traits
4.4. Genotyping for qDTY Validation
4.5. Analysis of Variances, Heritability, Correlation and qDTY Class Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Khush, G.S. What it will take to Feed 5.0 Billion Rice consumers in 2030. Plant Mol. Biol. 2005, 59, 1–6. [Google Scholar] [CrossRef] [PubMed]
- Swamy, B.P.M.; Kumar, A. Genomics-based precision breeding approaches to improve drought tolerance in rice. Biotechnol. Adv. 2013, 31, 1308–1318. [Google Scholar] [CrossRef] [PubMed]
- Barnabas, B.; Jager, K.; Feher, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef] [PubMed]
- Boyer, J.S.; Westgate, M.E. Grain yields with limited water. J. Exp. Bot. 2004, 55, 2385–2394. [Google Scholar] [CrossRef] [PubMed]
- Kumar, R.; Venuprasad, R.; Atlin, G.N. Genetic analysis of rainfed lowland rice drought tolerance under naturally-occurring stress in Eastern India: Heritability and QTL effects. Field Crop. Res. 2007, 103, 42–52. [Google Scholar] [CrossRef]
- Ouk, M.; Basnayake, J.; Tsubo, M.; Fukai, S.; Fischer, K.S.; Cooper, M.; Nesbitt, H. Use of drought response index for identification of drought tolerant genotypes in rainfed lowland rice. Field Crop. Res. 2006, 99, 48–58. [Google Scholar] [CrossRef]
- Bernier, J.; Serraj, R.; Kumar, A.; Venuprasad, R.; Impa, S.; Veeresh, V.G.; Oane, R.; Spaner, D.; Atlin, G. The large-effect drought-resistance QTL qtl12.1 increases water uptake in upland rice. Field Crop. Res. 2009, 110, 139–146. [Google Scholar] [CrossRef]
- Swamy, B.P.M.; Vikram, P.; Dixit, S.; Ahmed, H.U.; Kumar, A. Meta-analysis of grain yield QTL identified during agricultural drought in grasses showed consensus. BMC Genom. 2011, 12, 319. [Google Scholar] [CrossRef]
- Venuprasad, R.; Zenna, N.; Choi, R.I.; Amante, M.; Virk, P.S.; Kumar, A. Identification of Marker Loci Associated with Tungro and Drought Tolerance in Near-Isogenic Rice Lines Derived from IR64/Aday Sel. Int. Rice Res. Notes 2007, 32, 27–29. [Google Scholar] [CrossRef]
- Bernier, J.; Kumar, A.; Spaner, D.; Verulkar, S.; Mandal, N.P.; Sinha, P.K.; Peeraju, P.; Dongre, P.R.; Mahto, R.N.; Atlin, G.N. Characterization of the effect of rice drought tolerance qtl12.1 over a range of environments in the Philippines and eastern India. Euphytica 2009, 166, 207–217. [Google Scholar] [CrossRef]
- Bernier, J.; Kumar, A.; Ramaiah, V.; Spaner, D.; Atlin, G. A Large-Effect QTL for Grain Yield under Reproductive-Stage Drought Stress in Upland Rice. Crop Sci. 2007, 47, 507–516. [Google Scholar] [CrossRef]
- Ghimire, K.H.; Quiatchon, L.A.; Vikram, P.; Swamy, B.P.M.; Dixit, S.; Ahmed, H.; Hernandez, J.E.; Borromeo, T.H.; Kumar, A. Identification and mapping of a QTL (qDTY1.1) with a consistent effect on grain yield under drought. Field Crop. Res. 2012, 131, 88–96. [Google Scholar] [CrossRef]
- Vikram, P.; Swamy, B.; Dixit, S.; Ahmed, H.; Cruz, M.T.S.; Singh, A.; Kumar, A. qDTY1.1, a major QTL for rice grain yield under reproductive-stage drought stress with a consistent effect in multiple elite genetic backgrounds. BMC Genet. 2011, 12, 89. [Google Scholar] [CrossRef] [PubMed]
- Swamy, B.P.M.; Ahmed, H.U.; Henry, A.; Mauleon, R.; Dixit, S.; Vikram, P.; Tilatto, R.; Verulkar, S.B.; Perraju, P.; Mandal, N.P.; et al. Genetic, Physiological, and Gene Expression Analyses Reveal That Multiple QTL Enhance Yield of Rice Mega-Variety IR64 under Drought. PLoS ONE 2013, 8, e62795. [Google Scholar] [CrossRef] [PubMed]
- Shamsudin, N.A.A.; Swamy, B.P.M.; Ratnam, W.; Sta Cruz, M.T.; Raman, A.; Kumar, A. Marker assisted pyramiding of drought yield QTLs into a popular Malaysian rice cultivar, MR219. BMC Genet. 2016, 17, 30. [Google Scholar] [CrossRef] [PubMed]
- Shamsudin, N.A.A.; Swamy, B.P.M.; Ratnam, W.; Sta Cruz, M.T.; Sandhu, N.; Raman, A.K.; Kumar, A. Pyramiding of drought yield QTLs into a high quality Malaysian rice cultivar MRQ74 improves yield under reproductive stage drought. Rice 2016, 9, 21. [Google Scholar] [CrossRef] [PubMed]
- Chozin, M.A.; Lubis, I.; Junaedi, A.; Ehara, H. Some physiological character responses of rice under drought conditions in a paddy system. J. Int. Soc. Southeast Asian Agric. Sci. 2014, 20, 104–114. [Google Scholar]
- Sikuku, P.; Onyango, J.; Netondo, G.W. Physiological and biochemical responses of five nerica rice varieties (Oryza sativa L.) to water deficit at vegetative and reproductive stage. Agric. Biol. J. N. Am. 2012, 3, 93–104. [Google Scholar] [CrossRef]
- Pandey, V.; Shukla, A. Acclimation and Tolerance Strategies of Rice under Drought Stress. Rice Sci. 2015, 22, 147–161. [Google Scholar] [CrossRef]
- Fang, Y.; Xiong, L. General mechanisms of drought response and their application in drought resistance improvement in plants. Cell. Mol. Life Sci. 2015, 72, 673–689. [Google Scholar] [CrossRef] [PubMed]
- Bunnag, S.; Pongthai, P. Selection of Rice (Oryza sativa L.) Cultivars Tolerant to Drought Stress at the Vegetative Stage under Field Conditions. Am. J. Plant Sci. 2013, 4, 1701–1708. [Google Scholar] [CrossRef]
- Ashfaq, M.; Haider, M.S.; Khan, A.S.; Allah, S.U. Breeding potential of the basmati rice germplasm under water stress condition. Afr. J. Biotechnol. 2012, 11, 6647–6657. [Google Scholar]
- Ha, P.T.T. Physiological responses of rice seedlings under drought stress. J. Sci. Dev. 2014, 12, 635–640. [Google Scholar]
- Poorter, L.; Markesteijn, L. Seedling traits determine drought tolerance of tropical tree species. Biotropica 2008, 40, 321–331. [Google Scholar] [CrossRef]
- Kadioglu, A.; Terzi, R. A Dehydration Avoidance Mechanism: Leaf Rolling. Bot. Rev. 2007, 73, 290–302. [Google Scholar] [CrossRef]
- Dixit, S.; Kumar Biswal, A.; Min, A.; Henry, A.; Oane, R.H.; Raorane, M.L.; Longkumer, T.; Pabuayon, I.M.; Mutte, S.K.; Vardarajan, A.R.; et al. Action of multiple intra-QTL genes concerted around a co-localized transcription factor underpins a large effect QTL. Sci. Rep. 2015, 5, 15183. [Google Scholar] [CrossRef]
- Gowda, V.R.P.; Henry, A.; Yamauchi, A.; Shashidhar, H.E.; Serraj, R. Root biology and genetic improvement for drought avoidance in rice. Field Crop. Res. 2011, 122, 1–13. [Google Scholar] [CrossRef]
- Usman, M.; Raheem, Z.; Ahsan, T.; Iqbal, A.; Sarfaraz, Z.N.; Haq, Z. Morphological, Physiological and Biochemical Attributes as Indicators for Drought Tolerance in Rice (Oryza sativa L.). Eur. J. Biol. Sci. 2013, 5, 23–28. [Google Scholar]
- Clark, L.J.; Price, A.H.; Steele, K.A.; Whalley, W.R. Evidence from near-isogenic lines that root penetration increases with root diameter and bending stiffness in rice. Funct. Plant Biol. 2008, 35, 1163–1171. [Google Scholar] [CrossRef]
- Nguyen, H.T.; Babu, R.C.; Blum, A. Breeding for drought resistance in rice: Physiology and molecular genetics considerations. Crop Sci. 1997, 37, 1426–1434. [Google Scholar] [CrossRef]
- Palanog, A.D.; Swamy, B.P.M.; Shamsudin, N.A.A.; Dixit, S.; Hernandez, J.E.; Boromeo, T.H.; Cruz, P.C.S.; Kumar, A. Grain yield QTLs with consistent-effect under reproductive-stage drought stress in rice. Field Crop. Res. 2014, 161, 46–54. [Google Scholar] [CrossRef]
- Henry, A.; Swamy, B.P.M.; Dixit, S.; Torres, R.D.; Batoto, T.C.; Manalili, M.; Anantha, M.S.; Mandal, N.P.; Kumar, A. Physiological mechanisms contributing to the QTL-combination effects on improved performance of IR64 rice NILs under drought. J. Exp. Bot. 2015, 66, 1787–1799. [Google Scholar] [CrossRef]
- Mishra, K.K.; Vikram, P.; Yadaw, R.B.; Swamy, B.P.M.; Dixit, S.; Cruz, M.T.S.; Maturan, P.; Marker, S.; Kumar, A. qDTY12.1: A locus with a consistent effect on grain yield under drought in rice. BMC Genet. 2013, 14, 12. [Google Scholar] [CrossRef]
- Dixit, S.; Swamy, B.P.M.; Vikram, P.; Ahmed, H.U.; Sta Cruz, M.T.; Amante, M.; Atri, D.; Leung, H.; Kumar, A. Fine mapping of QTLs for rice grain yield under drought reveals sub-QTLs conferring a response to variable drought severities. Theor. Appl. Genet. 2012, 125, 155–169. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 3rd ed.; Sinauer Associates: Sunderland, MA, USA, 2002. [Google Scholar]
- Shao, H.; Chu, L.; Shao, M.; Jaleel, C.A.; Hong-mei, M. Higher plant antioxidants and redox signaling under environmental stresses. C. R. Biol. 2008, 331, 433–441. [Google Scholar] [CrossRef]
- Kamoshita, A.; Babu, R.C.; Boopathi, N.M.; Fukai, S. Phenotypic and genotypic analysis of drought-resistance traits for development of rice cultivars adapted to rainfed environments. Field Crop. Res. 2008, 109, 1–23. [Google Scholar] [CrossRef]
- Serraj, R.; McNally, K.L.; Slamet-Loedin, I.; Kohli, A.; Haefele, S.M.; Atlin, G.; Kumar, A. Drought Resistance Improvement in Rice: An Integrated Genetic and Resource Management Strategy. Plant Prod. Sci. 2011, 14, 1–14. [Google Scholar] [CrossRef]
- Vikram, P.; Swamy, B.P.M.; Dixit, S.; Singh, R.; Singh, B.P.; Miro, B.; Kohli, A.; Henry, A.; Singh, N.K.; Kumar, A. Drought susceptibility of modern rice varieties: An effect of linkage of drought tolerance with undesirable traits. Sci. Rep. 2015, 5, 14799. [Google Scholar] [CrossRef]
- Fukai, S.; Pantuwan, G.; Jongdee, B.; Cooper, M. Screening for drought resistance in rainfed lowland rice. Field Crop. Res. 1999, 64, 61–74. [Google Scholar] [CrossRef]
- Pantuwan, G.; Fukai, S.; Cooper, M.; Rajataserreekul, S.; O’Toole, J.C. Yield response of rice (Oryza sativa L.) genotypes to different types of drought under rainfed lowlands Part 1. Grain yield and yield components. Field Crop. Res. 2002, 73, 153–168. [Google Scholar] [CrossRef]
- Romyen, P.; Hanviriyapant, P.; Rajatasereekul, S.; Khunthasuvon, S.; Fukai, S.; Basnayake, J.; Skulkhu, E. Lowland rice improvement in northern and northeast Thailand: 2. Cultivar differences. Field Crop. Res. 1998, 59, 109–119. [Google Scholar] [CrossRef]
- Lafitte, H.R.; Price, A.H.; Courtois, B. Yield response to water deficit in an upland rice mapping population: Associations among traits and genetic markers. Theor. Appl. Genet. 2004, 109, 1237–1246. [Google Scholar] [CrossRef]
- Chang, T.T.; Somrith, B.; O’Toole, J.C. Rainfed Lowland Rice: Selected Papers from 1978; IRRI: Los Banos, Philippines, 1979. [Google Scholar]
- Venuprasad, R.; Bool, M.E.; Quiatchon, L.; Atlin, G.N. A QTL for rice grain yield in aerobic environments with large effects in three genetic backgrounds. Theor. Appl. Genet. 2012, 124, 323–332. [Google Scholar] [CrossRef]
- Zhao, X.; Qin, Y.; Sohn, J.K. Identification of main effects, epistatic effects and their environmental interactions of QTLs for yield traits in rice. Genes Genom. 2010, 32, 37–45. [Google Scholar] [CrossRef]
- Kanjoo, V.; Punyawaew, K.; Siangliw, J.L.; Jearakongman, S.; Vanavichit, A.; Toojinda, T. Evaluation of Agronomic Traits in Chromosome Segment Substitution Lines of KDML105 Containing Drought Tolerance QTL under Drought Stress. Rice Sci. 2012, 19, 117–124. [Google Scholar] [CrossRef]
- Suralta, R.R.; Inukai, Y.; Yamauchi, A. Genotypic Variations in Responses of Lateral Root Development to Transient Moisture Stresses in Rice Cultivars. Plant Prod. Sci. 2008, 11, 324–335. [Google Scholar] [CrossRef]
- Venuprasad, R.; Shashidhar, H.E.; Hittalmani, S.; Hemamalini, G.S. Tagging quantitative trait loci associated with grain yield and root morphological traits in rice (Oryza sativa L.) under contrasting moisture regimes. Euphytica 2002, 128, 293–300. [Google Scholar] [CrossRef]
- Blum, A. Plant Water Relations, Plant Stress and Plant Production. In Plant Breeding for Water-Limited Environments; Springer: New York, NY, USA, 2011; pp. 11–52. [Google Scholar]
- Asfaliza, R.; Omar, O.; Abdullah, S.; Baka, A.; Harun, M. MRQ74—A new aromatic rice variety. Bul. Teknol. Tanam 2008, 5, 15–20. [Google Scholar]
- Zainuddin, H.; Mokhtar, A.; Amzah, B.; Hashim, M.; Abd Ghafar, M.B. Enam Varieti Padi Popular MARDI. Bul. Teknol. MARDI 2012, 1, 1–10. [Google Scholar]
- IRRI. Standard Evaluation System (SES) for Rice, 5th ed.; International Rice Research Institute: Manila, Philippines, 2013. [Google Scholar]
- O’Toole, J.C.; Cruz, R.T. Response of leaf water potential, stomatal resistance, and leaf rolling to water stress. Plant Physiol. 1980, 65, 428–432. [Google Scholar] [CrossRef]
- Murray, M.G.; Thompson, W.F. Rapid isolation of high molecular weight plant DNA. Nucleic Acids Res. 1980, 8, 4321–4326. [Google Scholar] [CrossRef]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Traits | Environment | qDTY Classes | |||||||
|---|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | G | MRQ74 | ||
| DTF | NS | 87.17 c | 91.00 ab | 89.67 b | 84.63 d | 88.75 b | 76.20 e | 75.67 e | 93.00 a |
| RS | 90.92 a | 91.50 ab | 86.33 abc | 84.06 bc | 91.00 a | 79.70 c | 80.00 c | 89.00 abc | |
| PH | NS | 54.17 bc | 48.00 c | 63.33 a | 54.00 bc | 55.63 b | 52.00 bc | 54.67 bc | 48.00 c |
| RS | 58.14 bc | 48.32 de | 65.75 a | 55.49 cd | 60.43 ab | 47.57 c | 55.95 bcd | 58.17 abcd | |
| NP | NS | 10.63 ab | 6.50 ab | 8.50 ab | 11.81 a | 9.75 ab | 7.10 b | 7.33 ab | 12.50 ab |
| RS | 10.75 a | 6.50 a | 8.17 a | 11.19 a | 11.63 a | 8.80 a | 10.00 a | 14.50 a | |
| CC | NS | 43.65 c | 43.55 bc | 43.70 bc | 44.08 bc | 43.28 c | 47.16 a | 45.53 ab | 42.20 c |
| RS | 42.23 c | 41.27 bc | 42.23 bc | 42.98 bc | 42.09 bc | 44.51 a | 43.92 ab | 42.60 abc | |
| LR | RS | 1.79 bc | 1.00 cd | 1.17 d | 1.94 b | 1.50 bcd | 2.00 b | 2.00 b | 3.00 a |
| RW | NS | 2.05 c | 1.66 f | 1.66 f | 1.95 d | 2.27 b | 1.67 f | 1.86 e | 2.81 a |
| RS | 2.24 b | 1.53 bc | 1.94 bc | 2.20 b | 2.31 b | 1.50 c | 1.83 bc | 3.66 a | |
| RL | NS | 9.72 c | 7.72 f | 7.40 g | 9.35 d | 11.08 b | 7.65 f | 8.53 e | 14.17 a |
| RS | 12.08 a | 9.73 ab | 10.95 ab | 11.61 d | 12.78 a | 9.25 b | 13.02 a | 13.88 a | |
| GW | NS | 20.30 b | 15.50 d | 21.80 a | 18.80 c | 19.30 c | 20.60 b | 22.50 a | 18.40 c |
| RS | 19.20 cd | 16.90 d | 24.10 a | 19.60 c | 20.10 bc | 21.30 b | 23.30 a | 17.2 d | |
| GL | NS | 0.90 a | 0.80 a | 0.92 a | 1.21 a | 0.89 a | 0.89 a | 0.95 a | 0.85 a |
| RS | 0.90 b | 0.90 ab | 0.96 a | 0.89 b | 0.92 ab | 0.94 a | 0.93 ab | 0.89 ab | |
| GY | NS | 2715.36 c | 1446.93 e | 2316.93 d | 3381.47 b | 2460.30 d | 1882.03 e | 1711.60 e | 4361.20 a |
| RS | 920.81 c | 521.85 cd | 1104.75 bc | 976.90 bc | 1826.58 a | 445.85 d | 915.88 bc | 1618.06 ab | |
| Traits | Environment | qDTY Classes | ||||||
|---|---|---|---|---|---|---|---|---|
| A | B | C | D | E | F | MR219 | ||
| DTF | NS | 91.33 d | 85.90 e | 94.00 c | 83.80 f | 86.00 e | 95.50 b | 107.00 a |
| RS | 92.50 bc | 85.85 de | 88.50 cd | 84.20 e | 86.00 de | 95.63 ab | 98.00 a | |
| PH | NS | 62.00 bc | 60.30 c | 50.75 e | 57.00 d | 69.00 a | 55.00 de | 68.00 ab |
| RS | 63.29 b | 59.33 c | 58.17 cd | 55.08 d | 64.00 bc | 55.38 d | 76.67 a | |
| NP | NS | 7.33 a | 7.50 a | 5.00 bc | 5.77 b | 8.33 a | 4.50 c | 5.67 abc |
| RS | 8.03 ab | 8.17 a | 9.42 a | 6.47 b | 6.67 ab | 6.08 b | 10.33 a | |
| CC | NS | 40.38 a | 41.00 a | 40.90 a | 41.83 a | 39.43 a | 39.53 a | 39.00 a |
| RS | 37.58 c | 39.80 ab | 39.42 abc | 40.78 a | 39.00 abc | 36.30 c | 36.17 bc | |
| LR | RS | 0.58 abc | 0.39 bc | 0.50 abc | 0.68 a | 0.67 abc | 0.71 ab | 0.00 c |
| RW | NS | 0.58 b | 0.76 ab | 0.80 ab | 0.68 ab | 0.67 ab | 0.66 ab | 1.36 a |
| RS | 0.58 ab | 0.38 bc | 0.41 c | 0.34 c | 0.90 a | 0.56 abc | 0.82 ab | |
| RL | NS | 15.53 ab | 14.52 abc | 13.63 abc | 13.38 c | 12.55 bc | 13.95 bc | 18.00 a |
| RS | 13.41 ab | 11.16 bc | 12.63 abc | 9.88 c | 16.78 a | 12.22 abc | 13.38 abc | |
| GW | NS | 19.30 a | 18.80 a | 14.50 c | 19.40 a | 19.10 ab | 17.40 b | 16.70 bc |
| RS | 19.60 a | 19.20 a | 19.50 a | 18.60 a | 19.60 ab | 20.00 a | 13.50 b | |
| GL | NS | 0.93 a | 0.97 a | 0.98 a | 0.95 a | 0.97 a | 0.95 a | 1.02 a |
| RS | 0.92 a | 0.96 a | 0.97 a | 0.94 a | 0.93 a | 0.92 a | 0.97 a | |
| GY | NS | 2702.31 a | 2269.58 b | 1811.84 bc | 2269.58 b | 2364.53 abc | 1708.23 c | 2286.23 abc |
| RS | 2753.22 a | 1992.53 b | 1353.31 c | 1438.75 c | 2788.62 a | 1323.47 c | 2005.71 b | |
| Traits | MRQ74 PLs | MR219 PLs | ||
|---|---|---|---|---|
| Environment | NS | RS | NS | RS |
| DTF | 0.99 | 0.66 | 0.99 | 0.83 |
| PH | 0.89 | 0.86 | 0.80 | 0.88 |
| NP | 0.68 | 0.34 | 0.88 | 0.48 |
| CC | 0.69 | 0.55 | 0.67 | 0.53 |
| LR | NA | 0.63 | NA | 0.49 |
| RW | 0.99 | 0.14 | 0.00 | 0.10 |
| RL | 1.00 | 0.33 | 0.69 | 0.39 |
| GW | 0.93 | 0.85 | 0.92 | 0.00 |
| GL | 0.05 | 0.53 | 0.99 | 0.99 |
| GY | 0.98 | 0.87 | 0.91 | 0.96 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Mohd Ikmal, A.; Nurasyikin, Z.; Tuan Nur Aqlili Riana, T.A.; Puteri Dinie Ellina, Z.; Wickneswari, R.; Noraziyah, A.A.S. Drought Yield QTL (qDTY) with Consistent Effects on Morphological and Agronomical Traits of Two Populations of New Rice (Oryza sativa) Lines. Plants 2019, 8, 186. https://doi.org/10.3390/plants8060186
Mohd Ikmal A, Nurasyikin Z, Tuan Nur Aqlili Riana TA, Puteri Dinie Ellina Z, Wickneswari R, Noraziyah AAS. Drought Yield QTL (qDTY) with Consistent Effects on Morphological and Agronomical Traits of Two Populations of New Rice (Oryza sativa) Lines. Plants. 2019; 8(6):186. https://doi.org/10.3390/plants8060186
Chicago/Turabian StyleMohd Ikmal, Asmuni, Zainuddin Nurasyikin, Tuan Ali Tuan Nur Aqlili Riana, Zulkafli Puteri Dinie Ellina, Ratnam Wickneswari, and Abd Aziz Shamsudin Noraziyah. 2019. "Drought Yield QTL (qDTY) with Consistent Effects on Morphological and Agronomical Traits of Two Populations of New Rice (Oryza sativa) Lines" Plants 8, no. 6: 186. https://doi.org/10.3390/plants8060186
APA StyleMohd Ikmal, A., Nurasyikin, Z., Tuan Nur Aqlili Riana, T. A., Puteri Dinie Ellina, Z., Wickneswari, R., & Noraziyah, A. A. S. (2019). Drought Yield QTL (qDTY) with Consistent Effects on Morphological and Agronomical Traits of Two Populations of New Rice (Oryza sativa) Lines. Plants, 8(6), 186. https://doi.org/10.3390/plants8060186