Silicon Affects Root Development, Tissue Mineral Content, and Expression of Silicon Transporter Genes in Poinsettia (Euphorbia pulcherrima Willd.) Cultivars



Abstract
1. Introduction
2. Materials and Methods
2.1. Plant Materials and Growth Conditions
2.2. Supplementary Si Treatments
2.3. Measurement of Root Development and Sampling
2.4. Determination of Tissue Mineral Content
2.5. Quantitative RT-PCR
2.6. Statistical Analysis
3. Results
3.1. Survival Ratio
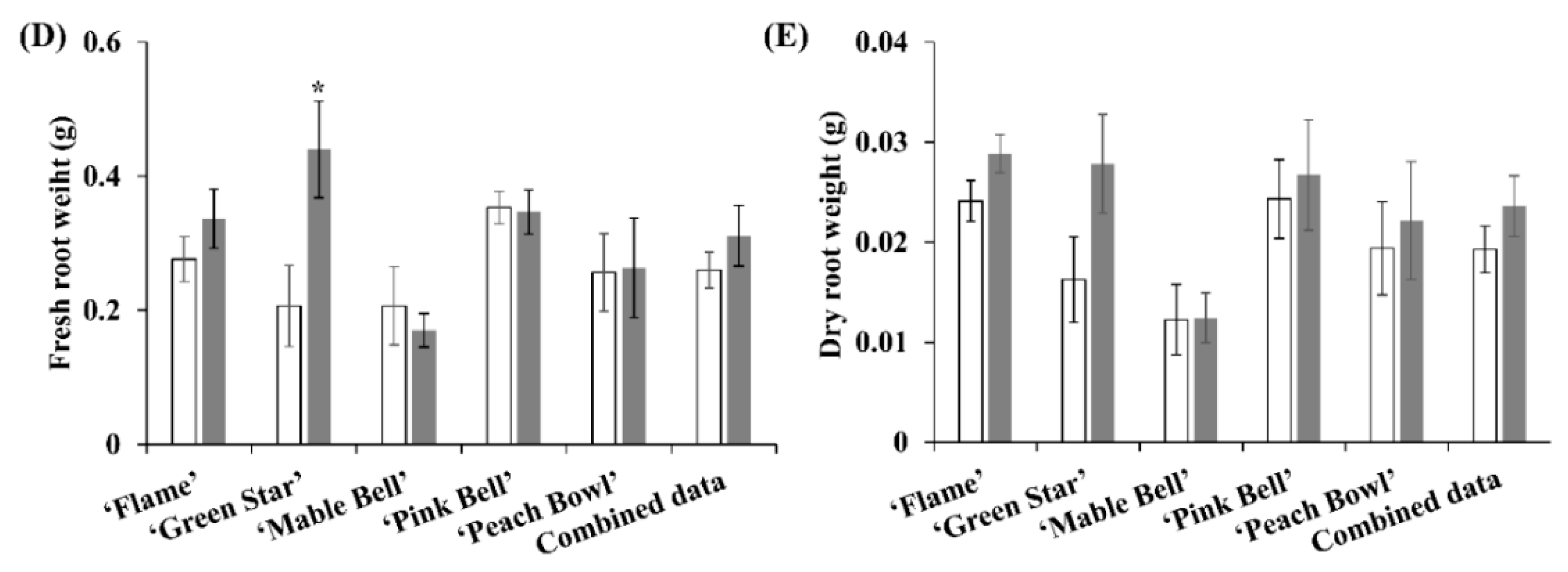
3.2. Root Development
3.3. Tissue Mineral Content
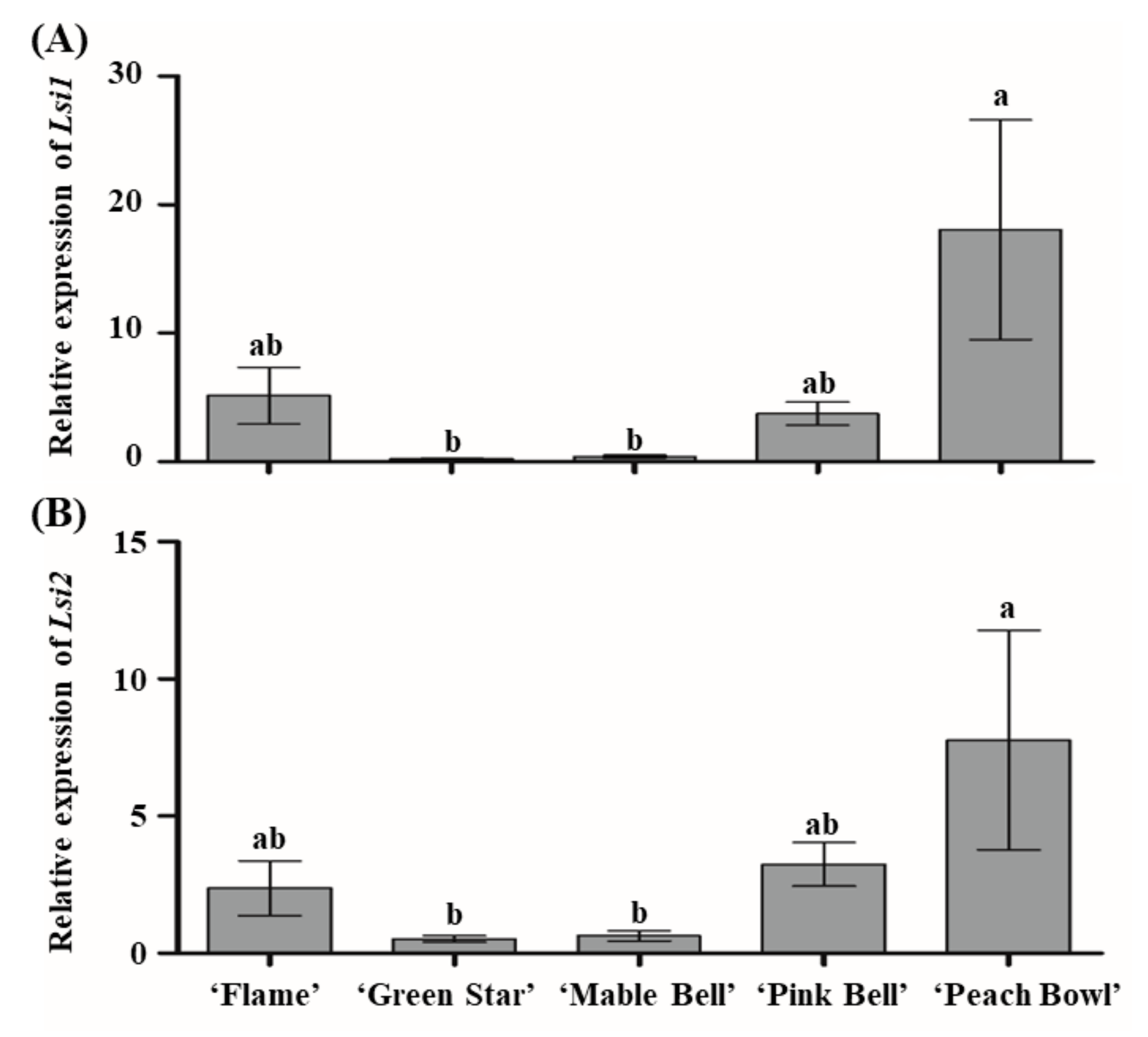
3.4. Expression of Si Transporter Genes
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Anonymous. Council of scientific and industrial research. In The Wealth of India; Council of Scientific and Industrial Research: New Delhi, India, 1978; Volume 3, pp. 224–230. [Google Scholar]
- Doyle, B.; Roycroft, J.; Cassells, A. The effects of stress on the quality of pelargonium propagules during shipment as measured using chlorophyll fluorescence. Available online: http://citeseerx.ist.psu.edu/viewdoc/download?doi=10.1.1.399.2546&rep=rep1&type=pdf (accessed on 16 June 2019).
- Faust, J.; Enfield, A. Effect of temperature and storage duration on quality and rooting performance of poinsettia (Euphorbia pulcherrima ‘Prestige Red’) cuttings. In Proceedings of the VI International Postharvest Symposium, Antalya, Turkey, 8–12 April 2009; pp. 1799–1807. [Google Scholar]
- Salehi Sardoei, A.; Shahmoradzadeh Fahraji, S.; Ghasemi, H. Effect of salicylic acid on rooting of poinsettia (Euphorbia pulcherrima). Int. J. Adv. Biol. Biomed. Res. 2014, 2, 1883–1886. [Google Scholar]
- Ramtin, A.; Khalighi, A.; Hadavi, E.; Hekmati, J. Effect of different IBA concentrations and types of cuttings on rooting and flowering poinsettia pulcherrima L. Int. J. Agr. Sci. 2011, 1, 303–310. [Google Scholar]
- Chowdhuri, T.; Sadhukhan, R.; Mondal, T. Effect of different growth regulators on propagation of poinsettia (Euphorbia pulcherrima Willd.) in subtropical zone of west Bengal. Contemp. Res. India 2017, 7, 161–165. [Google Scholar]
- Fereshtian, M.; Khandan-Mirkohi, A.; Schenk, M. Study on phosphorus supply management of poinsettia grown in peat-based substrate. J. Agr. Sci. Technol. 2018, 17, 179–188. [Google Scholar]
- Meharg, C.; Meharg, A.A. Silicon, the silver bullet for mitigating biotic and abiotic stress, and improving grain quality, in rice? Environ. Exp. Bot. 2015, 120, 8–17. [Google Scholar] [CrossRef]
- Ma, J.F.; Tamai, K.; Yamaji, N.; Mitani, N.; Konishi, S.; Katsuhara, M.; Ishiguro, M.; Murata, Y.; Yano, M. A silicon transporter in rice. Nature 2006, 440, 688–691. [Google Scholar] [CrossRef] [PubMed]
- Deshmukh, R.K.; Vivancos, J.; Ramakrishnan, G.; Guérin, V.; Carpentier, G.; Sonah, H.; Labbé, C.; Isenring, P.; Belzile, F.J.; Bélanger, R.R. A precise spacing between the npa domains of aquaporins is essential for silicon permeability in plants. Plant J. 2015, 83, 489–500. [Google Scholar] [CrossRef]
- Vatansever, R.; Ozyigit, I.I.; Filiz, E.; Gozukirmizi, N. Genome-wide exploration of silicon (Si) transporter genes, Lsi1 and Lsi2 in plants; insights into Si-accumulation status/capacity of plants. Biometals 2017, 30, 185–200. [Google Scholar] [CrossRef]
- Epstein, E. The anomaly of silicon in plant biology. Proc. Natl. Acad. Sci. USA 1994, 91, 11–17. [Google Scholar] [CrossRef]
- Bae, M.J.; Park, Y.G.; Jeong, B.R. Effect of a silicate fertilizer supplemented to the medium on rooting and subsequent growth of potted plants. Hortic. Environ. Biotechnol. 2010, 51, 355–359. [Google Scholar]
- Sivanesan, I.; Jeong, B.R. Silicon promotes adventitious shoot regeneration and enhances salinity tolerance of Ajuga multiflora Bunge by altering activity of antioxidant enzyme. Sci. World J. 2014, 2014, 521703. [Google Scholar] [CrossRef] [PubMed]
- Park, Y.G.; Muneer, S.; Kim, S.; Hwang, S.J.; Jeong, B.R. Foliar or subirrigational silicon supply modulates salt stress in strawberry during vegetative propagation. Hortic. Environ. Biotechnol. 2018, 59, 11–18. [Google Scholar] [CrossRef]
- Park, Y.G.; Muneer, S.; Kim, S.; Hwang, S.J.; Jeong, B.R. Silicon application during vegetative propagation affects photosynthetic protein expression in strawberry. Hortic. Environ. Biotechnol. 2018, 59, 167–177. [Google Scholar] [CrossRef]
- Soundararajan, P.; Manivannan, A.; Ko, C.H.; Muneer, S.; Jeong, B.R. Leaf physiological and proteomic analysis to elucidate silicon induced adaptive response under salt stress in Rosa hybrida ‘Rock Fire’. Int. J. Mol. Sci. 2017, 18, 1768. [Google Scholar] [CrossRef] [PubMed]
- Soundararajan, P.; Manivannan, A.; Ko, C.H.; Jeong, B.R. Silicon enhanced redox homeostasis and protein expression to mitigate the salinity stress in Rosa hybrida ‘Rock Fire’. J. Plant Growth Regul. 2018, 37, 16–34. [Google Scholar] [CrossRef]
- Santos, K.M.; Fisher, P.R.; Yeager, T.; Simonne, E.H.; Carter, H.S.; Argo, W.R. Timing of macronutrient supply during cutting propagation of petunia. HortScience 2011, 46, 475–480. [Google Scholar] [CrossRef]
- Santos, K.M.; Fisher, P.R.; Argo, W.R. Stem versus foliar uptake during propagation of Petunia × hybrida vegetative cuttings. HortScience 2009, 44, 1974–1977. [Google Scholar] [CrossRef]
- Gao, X.; Zou, C.; Wang, L.; Zhang, F. Silicon decreases transpiration rate and conductance from stomata of maize plants. J. Plant Nutr. 2006, 29, 1637–1647. [Google Scholar] [CrossRef]
- Ma, C.C.; Li, Q.F.; Gao, Y.B.; Xin, T.R. Effects of silicon application on drought resistance of cucumber plants. Soil Sci. Plant Nutr. 2004, 50, 623–632. [Google Scholar] [CrossRef]
- Chen, W.; Yao, X.; Cai, K.; Chen, J. Silicon alleviates drought stress of rice plants by improving plant water status, photosynthesis and mineral nutrient absorption. Biol. Trace. Elem. Res. 2011, 142, 67–76. [Google Scholar] [CrossRef]
- Gillman, J.H.; Zlesak, D.C. Mist applications of sodium silicate to rose (Rosa L. × ‘Nearly Wild’) cuttings decrease leaflet drop and increase rooting. HortScience 2000, 35, 773. [Google Scholar] [CrossRef]
- Whitcomb, C. Potentials of antitranspirants in plant propagation. Proc. Intl. Plant Prop. Soc. 1974, 24, 342–348. [Google Scholar]
- Guntzer, F.; Keller, C.; Meunier, J.-D. Benefits of plant silicon for crops: A review. Agron. Sustain. Dev. 2012, 32, 201–213. [Google Scholar] [CrossRef]
- Adrees, M.; Ali, S.; Rizwan, M.; Zia-Ur-Rehman, M.; Ibrahim, M.; Abbas, F.; Farid, M.; Qayyum, M.F.; Irshad, M.K. Mechanisms of silicon-mediated alleviation of heavy metal toxicity in plants: A review. Ecotox. Environ. Safe. 2015, 119, 186–197. [Google Scholar] [CrossRef] [PubMed]
- Ouellette, S.; Goyette, M.H.; Labbe, C.; Laur, J.; Gaudreau, L.; Gosselin, A.; Dorais, M.; Deshmukh, R.K.; Belanger, R.R. Silicon transporters and effects of silicon amendments in strawberry under high tunnel and field conditions. Front. Plant Sci. 2017, 8, 949. [Google Scholar] [CrossRef] [PubMed]
- Kamenidou, S.; Cavins, T.J.; Marek, S. Silicon supplements affect floricultural quality traits and elemental nutrient concentrations of greenhouse produced gerbera. Sci. Hortic. 2010, 123, 390–394. [Google Scholar] [CrossRef]
- Ma, J.F.; Yamaji, N.; Tamai, K.; Mitani, N. Genotypic difference in silicon uptake and expression of silicon transporter genes in rice. Plant Physiol. 2007, 145, 919–924. [Google Scholar] [CrossRef] [PubMed]
- Mitani-Ueno, N.; Yamaji, N.; Ma, J.F. High silicon accumulation in the shoot is required for down-regulating the expression of Si transporter genes in rice. Plant Cell Physiol. 2016, 57, 2510–2518. [Google Scholar] [CrossRef]
- Mitani, N.; Yamaji, N.; Ago, Y.; Iwasaki, K.; Ma, J.F. Isolation and functional characterization of an influx silicon transporter in two pumpkin cultivars contrasting in silicon accumulation. Plant J. 2011, 66, 231–240. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Cultivar (A) | Si (mg·L−1) (B) | Si (mg·g−1 Dry Weight) | Macronutrients (mg·g−1 Dry Weight) | Micronutrients (µg·g−1 Dry Weight) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| K | Ca | Mg | P | S | B | Cu | Fe | Mn | Zn | |||
| ‘Flame’ | 0 | 0.32 | 9.74 | 1.65 | 0.40 | 1.04 | 0.84 | 63.19 | 28.08 | 48.69 | 0.69 | 94.50 |
| 50 | 0.47*z | 14.01* | 2.71* | 0.65* | 1.51* | 0.77* | 49.65* | 42.15* | 47.54 | 5.20* | 41.28* | |
| ‘Green Star’ | 0 | 0.58 | 15.43 | 3.92 | 0.80 | 2.10 | 1.31 | 75.31 | 46.62 | 64.04 | 17.76 | 40.13 |
| 50 | 0.68* | 13.29* | 3.03* | 0.84* | 1.80* | 1.15* | 40.92* | 39.12* | 95.81* | 7.58* | 28.24* | |
| ‘Mable Bell’ | 0 | 0.78 | 13.03 | 3.96 | 0.72 | 1.26 | 1.21 | 92.62 | 62.05 | 81.52 | 3.17 | 35.55 |
| 50 | 0.59* | 13.07 | 3.30* | 0.61* | 1.63* | 1.56* | 84.00 | 55.85* | 81.99 | 0.89* | 58.98* | |
| ‘Pink Bell’ | 0 | 0.26 | 11.56 | 2.58 | 0.85 | 1.82 | 1.20 | 36.19 | 36.42 | 47.03 | 2.40 | 36.53 |
| 50 | 0.22* | 9.75* | 2.81* | 0.90* | 1.47* | 0.98* | 28.26* | 32.32* | 49.36* | 4.68* | 21.30* | |
| ‘Peach Bowl’ | 0 | 0.23 | 9.83 | 2.74 | 0.70 | 1.93 | 1.11 | 29.32 | 30.95 | 44.74 | 1.88 | 46.87 |
| 50 | 0.36* | 10.45* | 3.14* | 0.78* | 1.67* | 1.10 | 21.76 | 34.08* | 58.71* | 6.01* | 29.93* | |
| F-test | A | ***y | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| B | *** | ** | NS | *** | NS | ** | *** | NS | *** | *** | *** | |
| A×B | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
| Cultivar(A) | Si (mg·L−1) (B) | Si (mg·g−1 dry weight) | Macronutrients (mg·g−1 Dry Weight) | Micronutrients (µg·g−1 Dry Weight) | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| K | Ca | Mg | P | S | B | Cu | Fe | Mn | Zn | |||
| ‘Flame’ | 0 | 0.10 | 6.19 | 2.69 | 1.00 | 1.78 | 0.39 | 14.42 | 3.15 | 25.89 | 12.88 | 21.01 |
| 50 | 0.26*z | 6.66* | 2.61 | 1.00 | 1.68* | 0.49* | 11.68* | 4.06* | 30.13* | 10.33* | 16.34* | |
| ‘Green Star’ | 0 | 0.18 | 8.21 | 2.81 | 1.33 | 2.89 | 0.63 | 19.19 | 6.71 | 38.68 | 19.53 | 22.93 |
| 50 | 0.19* | 7.09* | 3.58* | 1.61* | 2.25* | 0.73* | 17.05* | 5.20* | 37.12* | 19.57 | 22.18* | |
| ‘Mable Bell’ | 0 | 0.29 | 7.27 | 3.31 | 1.48 | 2.04 | 0.67 | 15.11 | 8.43 | 36.07 | 17.36 | 19.90 |
| 50 | 0.11* | 9.45* | 4.52* | 2.20* | 3.10* | 0.81* | 15.31 | 7.11* | 51.09* | 27.85* | 25.58* | |
| ‘Pink Bell’ | 0 | 0.09 | 5.63 | 2.91 | 1.13 | 1.97 | 0.41 | 9.57 | 10.45 | 29.82 | 10.90 | 17.98 |
| 50 | 0.11* | 5.30* | 3.28* | 1.50* | 1.86 | 0.48* | 10.86* | 9.14* | 36.12* | 18.49* | 18.65 | |
| ‘Peach Bowl’ | 0 | 0.08 | 4.79 | 2.99 | 1.11 | 1.50 | 0.43 | 7.98 | 8.06 | 37.35 | 11.91 | 19.59 |
| 50 | 0.11* | 6.58* | 4.02* | 1.40* | 2.18* | 0.57* | 9.56* | 11.43* | 42.74* | 20.68* | 23.24* | |
| F-test | A | ***y | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** |
| B | *** | *** | *** | *** | *** | *** | *** | NS | *** | *** | *** | |
| A × B | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | *** | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hu, J.; Cai, X.; Jeong, B.R. Silicon Affects Root Development, Tissue Mineral Content, and Expression of Silicon Transporter Genes in Poinsettia (Euphorbia pulcherrima Willd.) Cultivars. Plants 2019, 8, 180. https://doi.org/10.3390/plants8060180
Hu J, Cai X, Jeong BR. Silicon Affects Root Development, Tissue Mineral Content, and Expression of Silicon Transporter Genes in Poinsettia (Euphorbia pulcherrima Willd.) Cultivars. Plants. 2019; 8(6):180. https://doi.org/10.3390/plants8060180
Chicago/Turabian StyleHu, Jiangtao, Xuan Cai, and Byoung Ryong Jeong. 2019. "Silicon Affects Root Development, Tissue Mineral Content, and Expression of Silicon Transporter Genes in Poinsettia (Euphorbia pulcherrima Willd.) Cultivars" Plants 8, no. 6: 180. https://doi.org/10.3390/plants8060180
APA StyleHu, J., Cai, X., & Jeong, B. R. (2019). Silicon Affects Root Development, Tissue Mineral Content, and Expression of Silicon Transporter Genes in Poinsettia (Euphorbia pulcherrima Willd.) Cultivars. Plants, 8(6), 180. https://doi.org/10.3390/plants8060180