Silicon as a Vegetable Crops Modulator—A Review



Abstract
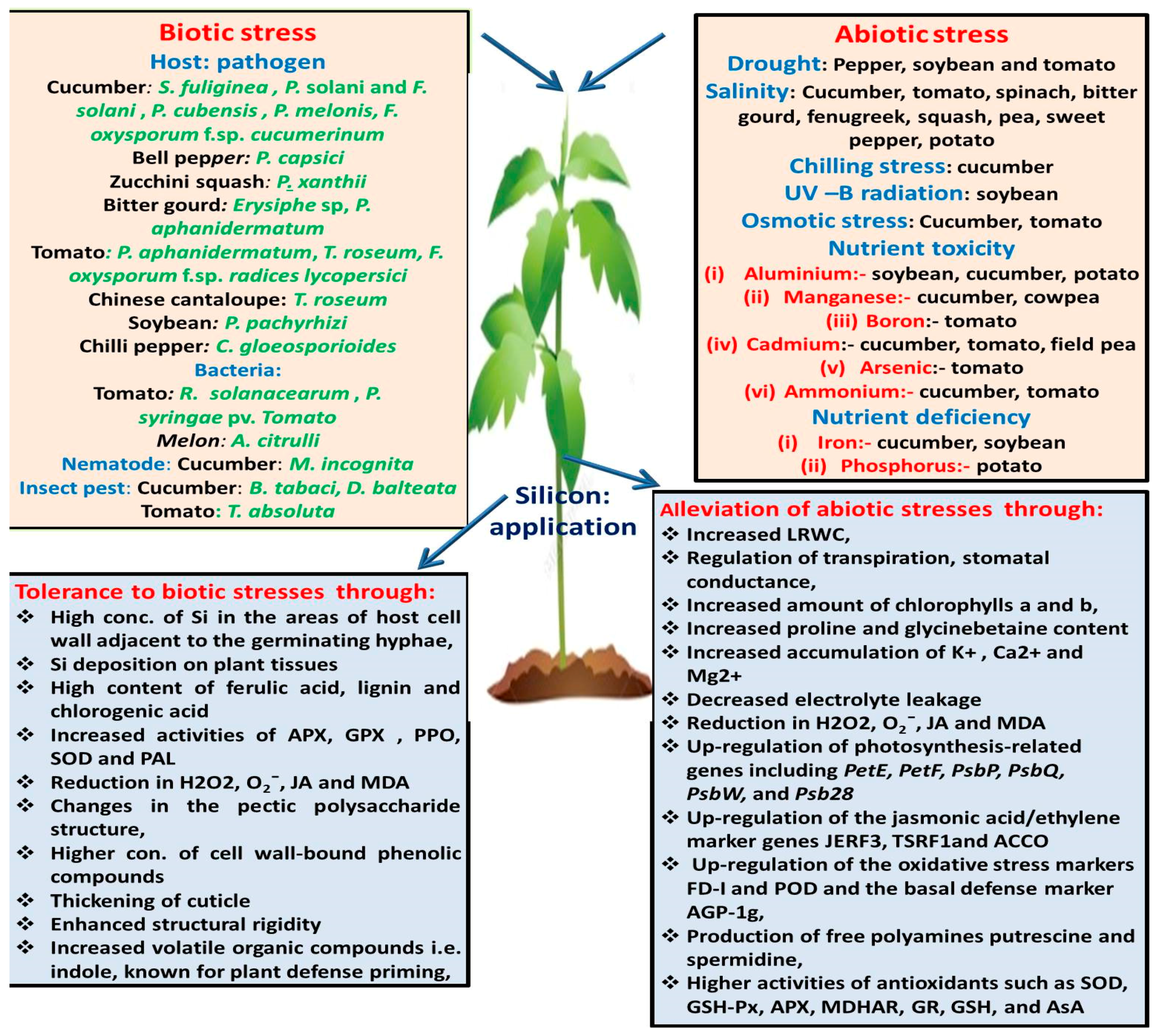
1. Introduction
2. Biotic Stress
2.1. Fungal Pathogens
2.1.1. Cucurbitaceae
2.1.2. Solanaceae
2.1.3. Leguminosae
2.2. Bacterial Pathogens
2.3. Insect Pest and Nematodes
3. Abiotic Stresses
3.1. Salinity
3.2. Drought
3.3. Other stresses
3.4. Mineral Toxicity
3.4.1. Aluminum
3.4.2. Manganese
3.4.3. Cadmium
3.4.4. Ammonium
4. Methods of Silicon Application on Vegetable Crops
5. Omics-Based Studies
6. Silicon Transporters
7. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ben Rejeb, I.; Pastor, V.; Mauch-Mani, B. Plant Responses to Simultaneous Biotic and Abiotic Stress: Molecular Mechanisms. Plants 2014, 3, 458–475. [Google Scholar] [CrossRef] [PubMed]
- Baloch, Q.B.; Chachar, Q.I.; Tareen, M.N. Effect of foliar application of macro and micro nutrients on production of green chilies (Capsicum annuum L.). J. Agric. Tech. 2008, 4, 177–184. [Google Scholar]
- Younis, A.; Riaz, A.; Sajid, M.; Mushtaq, N.; Ahsan, M.; Hameed, M.; Tariq, U.; Nadeem, M. Foliar application of macro-and micronutrients on the yield and quality of Rosa hybrida cvs. Cardinal and Whisky Mac. Afr. Biotech. 2013, 12, 702–708. [Google Scholar]
- Deshmukh, R.K.; Ma, J.F.; Bélanger, R.R. Editorial: Role of Silicon in Plants. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef]
- Carpinteri, A.; Manuello, A. Reply to “Comments on ‘Geomechanical and Geochemical Evidence of Piezonuclear Fission Reactions in the Earth’s Crust’by A. Carpinteri and A. Manuello” by U. Bardi and G. Comoretto. Strain 2013, 49, 548–551. [Google Scholar] [CrossRef]
- Meena, V.D.; Dotaniya, M.L.; Coumar, V.; Rajendiran, S.; Ajay; Kundu, S.; Rao, A.S. A Case for Silicon Fertilization to Improve Crop Yields in Tropical Soils. Proc. Natl. Acad. Sci. India 2014, 84, 505–518. [Google Scholar] [CrossRef]
- Epstein, E. Silicon. Ann Review Plant Biol. 1999, 50, 641–664. [Google Scholar] [CrossRef] [PubMed]
- White, P.J.; Brown, P.H. Plant nutrition for sustainable development and global health. Ann. Bot. 2010, 105, 1073–1080. [Google Scholar] [CrossRef]
- Tamai, K.; Ma, J.F. Characterization of silicon uptake by rice roots. New Phytologist. 2003, 158, 431–436. [Google Scholar] [CrossRef]
- Luyckx, M.; Hausman, J.-F.; Lutts, S.; Guerriero, G. Silicon and Plants: Current Knowledge and Technological Perspectives. Front. Plant Sci. 2017, 8. [Google Scholar] [CrossRef] [PubMed]
- Bakhat, H.F.; Bibi, N.; Zia, Z.; Abbas, S.; Hammad, H.M.; Fahad, S.; Ashraf, M.R.; Shah, G.M.; Rabbani, F.; Saeed, S. Silicon mitigates biotic stresses in crop plants: A review. Crop Prot. 2018, 104, 21–34. [Google Scholar] [CrossRef]
- Dann, E.K.; Muir, S. Peas grown in media with elevated plant-available silicon levels have higher activities of chitinase and β-1, 3-glucanase, are less susceptible to a fungal leaf spot pathogen and accumulate more foliar silicon. Australas. Plant Pathol. 2002, 31, 9–13. [Google Scholar] [CrossRef]
- Heath, M.C.; Stumpf, M.A. Ultrastructural observations of penetration sites of the cowpea rust fungus in untreated and silicon-depleted French bean cells. Physiol. Mol. Plant Pathol. 1986, 29, 27–39. [Google Scholar] [CrossRef]
- Liang, Y.C.; Sun, W.C.; Si, J.; Römheld, V. Effects of foliar-and root-applied silicon on the enhancement of induced resistance to powdery mildew in Cucumis sativus. Plant Pathol. 2005, 54, 678–685. [Google Scholar] [CrossRef]
- Chérif, M.; Asselin, A.; Bélanger, R.R. Defense responses induced by soluble silicon in cucumber roots infected by Pythium spp. Phytopathology 1994, 84, 236–242. [Google Scholar] [CrossRef]
- Barman, K.; Sharma, S.; Siddiqui, M.W. Emerging Postharvest Treatment of Fruits and Vegetables; CRC Press: Boca Raton, FL, USA, 2018; ISBN 978-1-351-04629-9. [Google Scholar]
- James, A.; Zikankuba, V. Postharvest management of fruits and vegetable: A potential for reducing poverty, hidden hunger and malnutrition in sub-Sahara Africa. Cogent Food Agric. 2017, 3, 1312052. [Google Scholar] [CrossRef]
- Reynolds, O.L.; Keeping, M.G.; Meyer, J.H. Silicon-augmented resistance of plants to herbivorous insects: A review. Ann. Appl. Biol. 2009, 155, 171–186. [Google Scholar] [CrossRef]
- Correa, R.S.; Moraes, J.C.; Auad, A.M.; Carvalho, G.A. Silicon and acibenzolar-S-methyl as resistance inducers in cucumber, against the whitefly Bemisia tabaci (Gennadius) (Hemiptera: Aleyrodidae) biotype B. Neotrop. Entomol. 2005, 34, 429–433. [Google Scholar] [CrossRef]
- Cooke, J.; Leishman, M.R. Consistent alleviation of abiotic stress with silicon addition: A meta-analysis. Func. Ecol. 2016, 30, 1340–1357. [Google Scholar] [CrossRef]
- Ashraf, M.; Afzal, M.; Ahmad, R.; Maqsood, M.A.; Shahzad, S.M.; Aziz, A.; Akhtar, N. Silicon management for mitigating abiotic stress effects in plants. Plant Stress 2010, 4, 104–114. [Google Scholar]
- Shen, X.; Li, J.; Duan, L.; Li, Z.; Eneji, A.E. Nutrient acquisition by soybean treated with and without silicon under ultraviolet-B radiation. J. Plant Nut. 2009, 32, 1731–1743. [Google Scholar] [CrossRef]
- Majeed Zargar, S.; Nazir, M.; Kumar Agrawal, G.; Kim, D.-W.; Rakwal, R. Silicon in plant tolerance against environmental stressors: Towards crop improvement using omics approaches. Current Proteom. 2010, 7, 135–143. [Google Scholar] [CrossRef]
- Sivanesan, I.; Park, S.W. The role of silicon in plant tissue culture. Front. Plant Sci. 2014, 5, 571. [Google Scholar] [CrossRef]
- Frew, A.; Weston, L.A.; Reynolds, O.L.; Gurr, G.M. The role of silicon in plant biology: A paradigm shift in research approach. Ann. Bot. 2018, 121, 1265–1273. [Google Scholar] [CrossRef]
- Hall, A.D.; Morison, C.G.T. Armstrong Henry Edward On the function of silica in the nutrition of cereals―Part I. Proceed. Royal Soc. London. 1906, 77, 455–477. [Google Scholar] [CrossRef]
- Samuels, A.L.; Glass, A.D.M.; Ehret, D.L.; Menzies, J.G. Distribution of silicon in cucumber leaves during infection by powdery mildew fungus (Sphaerotheca fuliginea). Canad. J. Bot. 1991, 69, 140–146. [Google Scholar] [CrossRef]
- El-Samman, M.G. The role of soluble silicon in controlling some root rot diseases of cucumber and tomato. Ann. Agric. Sci. (Cairo) 2000, 4, 1411–1419. [Google Scholar]
- Wei, G.; Zhu, Z.; Li, J.; Yao, Q. Effects of silicon supply and Sphaerotheca fuliginea inoculation on resistance of cucumber seedlings against powdery mildew. J. Appl. Ecol. 2004, 15, 2147–2151. [Google Scholar]
- Mohaghegh, P.; Khoshgoftarmanesh, A.H.; Shirvani, M.; Sharifnabi, B.; Nili, N. Effect of silicon nutrition on oxidative stress induced by Phytophthora melonis infection in cucumber. Plant Dis. 2011, 95, 455–460. [Google Scholar] [CrossRef]
- Heine, G.; Tikum, G.; Horst, W.J. The effect of silicon on the infection by and spread of Pythium aphanidermatum in single roots of tomato and bitter gourd. J. Exp. Bot. 2006, 58, 569–577. [Google Scholar] [CrossRef]
- Hasan, A.K.; Samir, S.H. Effect of copper and silicon nutrients and salicylic acid to induce systemic resistance for cucumber plants against Pythium aphanidermatum (Edson) Fitz. Arab J. Plant Prot. 2007, 25, 171–174. [Google Scholar]
- Yu, Y.; Schjoerring, J.K.; Du, X. Effects of silicon on the activities of defense-related enzymes in cucumber inoculated with Pseudoperonospora cubensis. J. of Plant Nut. 2010, 34, 243–257. [Google Scholar] [CrossRef]
- Zhou, X.; Shen, Y.; Fu, X.; Wu, F. Application of Sodium Silicate Enhances Cucumber Resistance to Fusarium Wilt and Alters Soil Microbial Communities. Front. Plant Sci. 2018, 9, 624. [Google Scholar] [CrossRef]
- Yurong, G.; Hua, Z.; Derong, C. Inhibitory mechanisms of two silicon compounds on mildew powder of melon. Sci. Agric. Sin. 2005. [Google Scholar]
- Buttaro, D.; Bonasia, A.; Minuto, A.; Serio, F.; Santamaria, P. Effect of silicon in the nutrient solution on the incidence of powdery mildew and quality traits in carosello and barattiere (Cucumis melo L.) grown in a soilless system. J. Hort. Sci. Biotech. 2009, 84, 300–304. [Google Scholar] [CrossRef]
- Dallagnol, L.J.; Rodrigues, F.A.; Tanaka, F.A.O.; Amorim, L.; Camargo, L.E.A. Effect of potassium silicate on epidemic components of powdery mildew on melon. Plant Pathol. 2012, 61, 323–330. [Google Scholar] [CrossRef]
- Ratnayake, R.; Daundasekera, W.A.M.; Ariyarathne, H.M.; Ganehenege, M.Y.U. Some biochemical defense responses enhanced by soluble silicon in bitter gourd-powdery mildew pathosystem. Austral. Plant Pathol. 2016, 45, 425–433. [Google Scholar] [CrossRef]
- Guo, Y.; Liu, L.; Zhao, J.; Bi, Y. Use of silicon oxide and sodium silicate for controlling Trichothecium roseum postharvest rot in Chinese cantaloupe (Cucumis melo L.). Int. J. Food Sci. Tech. 2007, 42, 1012–1018. [Google Scholar] [CrossRef]
- Savvas, D.; Giotis, D.; Chatzieustratiou, E.; Bakea, M.; Patakioutas, G. Silicon supply in soilless cultivations of zucchini alleviates stress induced by salinity and powdery mildew infections. Environ. Exp. Bot. 2009, 65, 11–17. [Google Scholar] [CrossRef]
- Lepolu Torlon, J.; Heckman, J.; Simon, J.; Wyenandt, C. Silicon soil amendments for suppressing powdery mildew on pumpkin. Sustainability 2016, 8, 293. [Google Scholar] [CrossRef]
- Diogo, R.V.; Wydra, K. Silicon-induced basal resistance in tomato against Ralstonia solanacearum is related to modification of pectic cell wall polysaccharide structure. Physiol. Mol. Plant Pathol. 2007, 70, 120–129. [Google Scholar] [CrossRef]
- Huang, C.-H.; Roberts, P.D.; Datnoff, L.E. Silicon suppresses Fusarium crown and root rot of tomato. J. Phytopathol. 2011, 159, 546–554. [Google Scholar] [CrossRef]
- David, D.; Weerahewa, H.L.D. Silicon Suppresses Anthracnose Diseases in Tomato (Lycopersicon esculentum) by Enhancing Disease Resistance; OUSL: Nugegoda, Sri Lanka, 2012. [Google Scholar]
- French-Monar, R.D.; Rodrigues, F.A.; Korndörfer, G.H.; Datnoff, L.E. Silicon suppresses Phytophthora blight development on bell pepper. J. Phytopathol. 2010, 158, 554–560. [Google Scholar] [CrossRef]
- Jayawardana, H.; Weerahewa, H.L.D.; Saparamadu, M. Enhanced resistance to anthracnose disease in chili pepper (Capsicum annuum L.) by amendment of the nutrient solution with silicon. J. Horticult. Sci. Biotech. 2015, 90, 557–562. [Google Scholar] [CrossRef]
- Kaushik, P.; Dhaliwal, M.S.; Kumar, S. Usefulness of Three under Exploited Vegetable Legumes- A Review. Int. J. Curr. Microbiol. Appl. Sci. 2018, 7, 861–870. [Google Scholar] [CrossRef]
- Arsenault-Labrecque, G.; Menzies, J.G.; Bélanger, R.R. Effect of silicon absorption on soybean resistance to Phakopsora pachyrhizi in different cultivars. Plant Dis. 2012, 96, 37–42. [Google Scholar] [CrossRef]
- Lemes, E.M.; Mackowiak, C.L.; Blount, A.; Marois, J.J.; Wright, D.L.; Coelho, L.; Datnoff, L.E. Effects of silicon applications on soybean rust development under greenhouse and field conditions. Plant Dis. 2011, 95, 317–324. [Google Scholar] [CrossRef]
- Dannon, E.A.; Wydra, K. Interaction between silicon amendment, bacterial wilt development and phenotype of Ralstonia solanacearum in tomato genotypes. Physiol. Mol. Plant Pathol. 2004, 64, 233–243. [Google Scholar] [CrossRef]
- Wydra, K.; Diogo, R.; Dannon, E.; Semrau, J. Soil amendment with silicon and bacterial antagonists induce resistance against bacterial wilt caused by Ralstonia solanacearum in tomato. Tropentag 2005. [Google Scholar]
- Ghareeb, H.; Bozsó, Z.; Ott, P.G.; Repenning, C.; Stahl, F.; Wydra, K. Transcriptome of silicon-induced resistance against Ralstonia solanacearum in the silicon non-accumulator tomato implicates priming effect. Physiol. Mol. Plant Pathol. 2011, 75, 83–89. [Google Scholar] [CrossRef]
- Wydra, K.; Semrau, J.; Dannon, E.; Diogo, R. Characterization of the interaction of antagonistic bacteria and of silicon (SiO∼ 2) with tomato infected with Ralstonia solanacearum. Mitteilungen-Biolosichen Bundesantalt Fur Land Und Forstwirtschaft 2006, 408, 112. [Google Scholar]
- Wang, L.; Cai, K.; Chen, Y.; Wang, G. Silicon-mediated tomato resistance against Ralstonia solanacearum is associated with modification of soil microbial community structure and activity. Biol. Trace Element Res. 2013, 152, 275–283. [Google Scholar] [CrossRef]
- Wang, L.; Chen, Y.; Cai, K.; Wang, G. Effects of exogenous silicon supply on the activity of antioxidant enzymes of tomato leaves infected by Ralstonia solanacearum. J. South China Agric. Univ. 2014, 35, 74–78. [Google Scholar]
- Ferreira, R.S.; Moraes, J.C. Silicon influence on resistance induction against Bemisia tabaci biotype B (Genn.)(Hemiptera: Aleyrodidae) and on vegetative development in two soybean cultivars. Neotrop. Entomol. 2011, 40, 495–500. [Google Scholar] [CrossRef]
- Callis-Duehl, K.L.; McAuslane, H.J.; Duehl, A.J.; Levey, D.J. The effects of silica fertilizer as an anti-herbivore defense in cucumber. J. Hortic. Res. 2017, 25, 89–98. [Google Scholar] [CrossRef]
- Dos Santos, M.C.; Junqueira, M.R.; de Sá, V.M.; Zanúncio, J.C.; Serrão, J.E. Effect of silicon on the morphology of the midgut and mandible of tomato leafminer Tuta absoluta (Lepidoptera: Gelechiidae) larvae. Invert. Surv. J. 2015, 12, 158–165. [Google Scholar]
- Dugui-Es, C.; Pedroche, N.; Villanueva, L.; Galeng, J.; De, D.W. Management of root knot nematode, Meloidogyne incognita in cucumber (Cucumis sativus) using silicon. Comm. Agric. Appl. Biol. Sci. 2010, 75, 497–505. [Google Scholar]
- Miyake, Y.; Takahashi, E. Effect of silicon on the growth of cucumber plant in soil culture. Soil Sci. Plant Nut. 1983, 29, 463–471. [Google Scholar] [CrossRef]
- Menzies, J.G.; Ehret, D.L.; Glass, A.D.M.; Helmer, T.; Koch, C.; Seywerd, F. Effects of soluble silicon on the parasitic fitness of Sphaerotheca fuliginea on Cucumis sativus. Phytopathology (USA) 1991. [Google Scholar] [CrossRef]
- Schuerger, A.C.; Hammer, W. Suppression of powdery mildew on greenhouse-grown cucumber by addition of silicon to hydroponic nutrient solution is inhibited at high temperature. Plant Dis. 2003, 87, 177–185. [Google Scholar] [CrossRef]
- Yao, Q.; Zhang, X.; Jiang, W.; Wei, G. Influences of silicon on phenylalanine metabolism in cucumber leaves and its relation to resistance to powdery mildew. J. Jilin Agric. Univ. 2009, 31, 16–21. [Google Scholar]
- Wolff, S.A.; Karoliussen, I.; Rohloff, J.; Strimbeck, R. Foliar applications of silicon fertilisers inhibit powdery mildew development in greenhouse cucumber. J. Food Agric. Environ. 2012, 10, 355–359. [Google Scholar]
- Chitarra, W.; Gilardi, G.; Cogliati, E.E.; Pugliese, M.; Gullino, M.L.; Garibaldi, A. Effects of silicon and concentration of nutrient solution on the control of Fusarium wilt of tomato. Protezione delle Colture 2013, 59–60. [Google Scholar]
- Chitarra, W.; Cogliati, E.; Pugliese, M.; Gilardi, G.; Gullino, M.L.; Garibaldi, A. Effect of silicates and electrical conductivity on Fusarium wilt of tomato grown soilless. In Proceedings of the VIII International Symposium on Chemical and Non-Chemical Soil and Substrate Disinfestation 1044, Torino, Italy, 13–17 July 2014; pp. 53–55. [Google Scholar]
- Weerahewa, H.L.D.; David, D. Effect of silicon and potassium on tomato anthracnose and on the postharvest quality of tomato fruit (Lycopersicon esculentum Mill.). J. Nat. Sci. Found. Sri Lanka 2015, 43, 273–280. [Google Scholar] [CrossRef]
- Somapala, K.; Weerahewa, D.; Thrikawala, S. Silicon rich rice hull amended soil enhances anthracnose resistance in tomato. Proced. Food Sci. 2016, 6, 190–193. [Google Scholar] [CrossRef]
- Kurabachew, H.; Wydra, K. Induction of defense related enzymes and gene expression after resistance induction by rhizobacteria and silicon against Ralstonia solanacearum in tomato (Solanum lycopersicum). In Proceedings of the Conference on International Research on Food Security, Natural Resource Management and Rural Development, Zurich, Switzerland, 14–16 September 2010. [Google Scholar]
- Kurabachew, H.; Stahl, F.; Wydra, K. Global gene expression of rhizobacteria-silicon mediated induced systemic resistance in tomato (Solanum lycopersicum) against Ralstonia solanacearum. Physiol. Mol. Plant Pathol. 2013, 84, 44–52. [Google Scholar] [CrossRef]
- Andrade, C.C.L.; Resende, R.S.; Rodrigues, F.Á.; Ferraz, H.G.M.; Moreira, W.R.; Oliveira, J.R.; Mariano, R.L.R. Silicon reduces bacterial speck development on tomato leaves. Trop. Plant Pathol. 2013, 38, 436–442. [Google Scholar] [CrossRef]
- Conceição, C.S.; Felix, K.C.S.; Mariano, R.L.; Medeiros, E.V.; Souza, E.B. Combined effect of yeast and silicon on the control of bacterial fruit blotch in melon. Sci. Hort. 2014, 174, 164–170. [Google Scholar] [CrossRef]
- Kurabachew, H.; Wydra, K. Induction of systemic resistance and defense-related enzymes after elicitation of resistance by rhizobacteria and silicon application against Ralstonia solanacearum in tomato (Solanum lycopersicum). Crop Prot. 2014, 57, 1–7. [Google Scholar] [CrossRef]
- Chen, Y.; Liu, M.; Wang, L.; Lin, W.; Fan, X.; Cai, K. Proteomic characterization of silicon-mediated resistance against Ralstonia solanacearum in tomato. Plant Soil 2015, 387, 425–440. [Google Scholar] [CrossRef]
- Ferreira, H.A.; do Nascimento, C.W.A.; Datnoff, L.E.; de Sousa Nunes, G.H.; Preston, W.; de Souza, E.B.; de Mariano, R.L.R. Effects of silicon on resistance to bacterial fruit blotch and growth of melon. Crop Prot. 2015, 78, 277–283. [Google Scholar] [CrossRef]
- Shahbaz, M.; Ashraf, M.; Al-Qurainy, F.; Harris, P.J.C. Salt tolerance in selected vegetable crops. Crit. Rev. Plant Sci. 2012, 31, 303–320. [Google Scholar] [CrossRef]
- Shannon, M.C.; Grieve, C.M. Tolerance of vegetable crops to salinity. Sci. Hort. 1998, 78, 5–38. [Google Scholar] [CrossRef]
- Munns, R. Genes and salt tolerance: Bringing them together. New phytologist 2005, 167, 645–663. [Google Scholar] [CrossRef]
- Bae, D.; Yong, K.; Chun, S. Effect of salt (NaCl) stress on germination and early seedling growth of four vegetables species. J. Cent. Eur. Agric. 2006, 7, 273–282. [Google Scholar]
- Rizwan, M.; Ali, S.; Ibrahim, M.; Farid, M.; Adrees, M.; Bharwana, S.A.; Zia-ur-Rehman, M.; Qayyum, M.F.; Abbas, F. Mechanisms of silicon-mediated alleviation of drought and salt stress in plants: A review. Environ. Sci. Pollut. Res. 2015, 22, 15416–15431. [Google Scholar] [CrossRef]
- Witzel, K.; Mock, H.P. A proteomic view of the cereal and vegetable crop response to salinity stress. In Agricultural Proteomics Volume 2; Springer: Cham, Switzerland, 2016; pp. 53–69. [Google Scholar]
- Haghighi, M.; Pessarakli, M. Influence of silicon and nano-silicon on salinity tolerance of cherry tomatoes (Solanum lycopersicum L.) at early growth stage. Sci. Hort. 2013, 161, 111–117. [Google Scholar] [CrossRef]
- Zhu, Z.; Wei, G.; Li, J.; Qian, Q.; Yu, J. Silicon alleviates salt stress and increases antioxidant enzymes activity in leaves of salt-stressed cucumber (Cucumis sativus L.). Plant Sci. 2004, 167, 527–533. [Google Scholar] [CrossRef]
- Eraslan, F.; Inal, A.; Pilbeam, D.J.; Gunes, A. Interactive effects of salicylic acid and silicon on oxidative damage and antioxidant activity in spinach (Spinacia oleracea L. cv. Matador) grown under boron toxicity and salinity. Plant Growth Regul. 2008, 55, 207. [Google Scholar] [CrossRef]
- Wang, X.; Ou-yang, C.; Fan, Z.; Gao, S.; Chen, F.; Tang, L. Effects of exogenous silicon on seed germination and antioxidant enzyme activities of Momordica charantia under salt stress. J. Animal Plant Sci. 2010, 6, 700–708. [Google Scholar]
- Khoshgoftarmanesh, A.H.; Khodarahmi, S.; Haghighi, M. Effect of silicon nutrition on lipid peroxidation and antioxidant response of cucumber plants exposed to salinity stress. Arch. Agron. Soil Sci. 2014, 60, 639–653. [Google Scholar] [CrossRef]
- Bu, R.; Xie, J.; Yu, J.; Liao, W.; Xiao, X.; Lv, J.; Wang, C.; Ye, J.; Calderón-Urrea, A. Autotoxicity in cucumber (Cucumis sativus L.) seedlings is alleviated by silicon through an increase in the activity of antioxidant enzymes and by mitigating lipid peroxidation. J. Plant Biol. 2016, 59, 247–259. [Google Scholar] [CrossRef]
- Shen, X.; Li, X.; Li, Z.; Li, J.; Duan, L.; Eneji, A.E. Growth, physiological attributes and antioxidant enzyme activities in soybean seedlings treated with or without silicon under UV-B radiation stress. J. Agron. Crop Sci. 2010, 196, 431–439. [Google Scholar] [CrossRef]
- Romero-Aranda, M.R.; Jurado, O.; Cuartero, J. Silicon alleviates the deleterious salt effect on tomato plant growth by improving plant water status. J. Plant Physiol. 2006, 163, 847–855. [Google Scholar] [CrossRef]
- Shahid, M.A.; Balal, R.M.; Pervez, M.A.; Abbas, T.; Aqueel, M.A.; Javaid, M.M.; Garcia-Sanchez, F. Foliar spray of phyto-extracts supplemented with silicon: An efficacious strategy to alleviate the salinity-induced deleterious effects in pea (Pisum sativum L.). Turk. J. Bot. 2015, 39, 408–419. [Google Scholar] [CrossRef]
- Tantawy, A.S.; Salama, Y.A.M.; El-Nemr, M.A.; Abdel-Mawgoud, A.M.R. Nano silicon application improves salinity tolerance of sweet pepper plants. Int. J. ChemTech Res. 2015, 8, 11–17. [Google Scholar]
- Siddiqui, M.H.; Al-Whaibi, M.H.; Faisal, M.; Al Sahli, A.A. Nano-silicon dioxide mitigates the adverse effects of salt stress on Cucurbita pepo L. Env. Toxicol. Chem. 2014, 33, 2429–2437. [Google Scholar] [CrossRef]
- Zhu, Y.-X.; Xu, X.-B.; Hu, Y.-H.; Han, W.-H.; Yin, J.-L.; Li, H.-L.; Gong, H.-J. Silicon improves salt tolerance by increasing root water uptake in Cucumis sativus L. Plant Cell Rep. 2015, 34, 1629–1646. [Google Scholar] [CrossRef]
- Manivannan, A.; Soundararajan, P.; Muneer, S.; Ko, C.H.; Jeong, B.R. Silicon mitigates salinity stress by regulating the physiology, antioxidant enzyme activities, and protein expression in Capsicum annuum ‘Bugwang’. BioMed Res. Int. 2016. [Google Scholar] [CrossRef]
- Yin, J.; Jia, J.; Lian, Z.; Hu, Y.; Guo, J.; Huo, H.; Zhu, Y.; Gong, H. Silicon enhances the salt tolerance of cucumber through increasing polyamine accumulation and decreasing oxidative damage. Ecotoxicol. Environ. Safety 2019, 169, 8–17. [Google Scholar] [CrossRef]
- Lobato, A.K.S.; Coimbra, G.K.; Neto, M.A.M.; Costa, R.C.L.; Santos, F.; Oliveira, N.; Luz, L.M.; Barreto, A.G.T.; Pereira, B.W.F.; Alves, G.A.R. Protective action of silicon on water relations and photosynthetic pigments in pepper plants induced to water deficit. Res. J. Biol. Sci. 2009, 4, 617–623. [Google Scholar]
- Lobato, A.K.S.; Luz, L.M.; Costa, R.C.L.; Santos Filho, B.G.; Meirelles, A.C.S.; Oliveira Neto, C.F.; Laughinghouse, I.V.; Neto, M.A.M.; Alves, G.A.R.; Lopes, M.J. Silicon exercises influence on nitrogen compounds in pepper subjected to water deficit. Res. J. Biol. Sci. 2009, 4, 1048–1055. [Google Scholar]
- Hamayun, M.; Sohn, E.-Y.; Khan, S.A.; Shinwari, Z.K.; Khan, A.L.; Lee, I.-J. Silicon alleviates the adverse effects of salinity and drought stress on growth and endogenous plant growth hormones of soybean (Glycine max L.). Pak. J. Bot. 2010, 42, 1713–1722. [Google Scholar]
- Shi, Y.; Zhang, Y.; Yao, H.; Wu, J.; Sun, H.; Gong, H. Silicon improves seed germination and alleviates oxidative stress of bud seedlings in tomato under water deficit stress. Plant Physiol. Biochem. 2014, 78, 27–36. [Google Scholar] [CrossRef]
- Cao, B.; Ma, Q.; Zhao, Q.; Wang, L.; Xu, K. Effects of silicon on absorbed light allocation, antioxidant enzymes and ultrastructure of chloroplasts in tomato leaves under simulated drought stress. Sci. Hort. 2015, 194, 53–62. [Google Scholar] [CrossRef]
- Ullah, U.; Ashraf, M.; Shahzad, S.M.; Siddiqui, A.R.; Piracha, M.A.; Suleman, M. Growth behavior of tomato (Solanum lycopersicum L.) under drought stress in the presence of silicon and plant growth promoting rhizobacteria. Soil Environ. 2016, 35, 65–75. [Google Scholar]
- Shi, Y.; Zhang, Y.; Han, W.; Feng, R.; Hu, Y.; Guo, J.; Gong, H. Silicon enhances water stress tolerance by improving root hydraulic conductance in Solanum lycopersicum L. Front. Plant Sci. 2016, 7, 196. [Google Scholar] [CrossRef]
- Cao, B.-L.; Wang, L.; Gao, S.; Xia, J.; Xu, K. Silicon-mediated changes in radial hydraulic conductivity and cell wall stability are involved in silicon-induced drought resistance in tomato. Protoplasma 2017, 254, 2295–2304. [Google Scholar] [CrossRef]
- Hattori, T.; Sonobe, K.; Inanaga, S.; An, P.; Morita, S. Effects of silicon on photosynthesis of young cucumber seedlings under osmotic stress. J. Plant Nut. 2008, 31, 1046–1058. [Google Scholar] [CrossRef]
- Zhang, P.Y.; Gao, R.G.; Yang, F.J.; Wang, X.F.; Wei, M.; Shi, Q.H.; Li, Y. Effects of silicon on photosynthetic characteristics and activity of antioxidant enzymes in continuous-cropped cucumber seedlings. J. Appl. Ecol. 2014, 25, 1733–1738. [Google Scholar]
- Jafari, S.R.; Arvin, S.M.J.; Kalantari, K.M. Response of cucumber (Cucumis sativus L.) seedlings to exogenous silicon and salicylic acid under osmotic stress. Acta Biologica Szegediensis 2015, 59, 25–33. [Google Scholar]
- Liu, J.-J.; Lin, S.-H.; Xu, P.-L.; Wang, X.-J.; Bai, J.-G. Effects of exogenous silicon on the activities of antioxidant enzymes and lipid peroxidation in chilling-stressed cucumber leaves. Agric. Sci. China 2009, 8, 1075–1086. [Google Scholar] [CrossRef]
- Baylis, A.D.; Gragopoulou, C.; Davidson, K.J.; Birchall, J.D. Effects of silicon on the toxicity of aluminum to soybean. Comm. Soil Sci. Plant Anal. 1994, 25, 537–546. [Google Scholar] [CrossRef]
- Bityutskii, N.P.; Yakkonen, K.L.; Petrova, A.I.; Shavarda, A.L. Interactions between aluminum, iron and silicon in Cucumber sativus L. grown under acidic conditions. J. Plant Physiol. 2017, 218, 100–108. [Google Scholar] [CrossRef]
- Dorneles, A.O.S.; Pereira, A.S.; Sasso, V.M.; Possebom, G.; Tarouco, C.P.; Schorr, M.R.W.; Rossato, L.; Ferreira, P.A.A.; Tabaldi, L.A. Aluminum stress tolerance in potato genotypes grown with silicon. Bragantia 2019. [Google Scholar] [CrossRef]
- Iwasaki, K.; Maier, P.; Fecht, M.; Horst, W.J. Effects of silicon supply on apoplastic manganese concentrations in leaves and their relation to manganese tolerance in cowpea (Vigna unguiculata (L.) Walp.). Plant Soil 2002, 238, 281–288. [Google Scholar] [CrossRef]
- Shi, Q.; Bao, Z.; Zhu, Z.; He, Y.; Qian, Q.; Yu, J. Silicon-mediated alleviation of Mn toxicity in Cucumis sativus in relation to activities of superoxide dismutase and ascorbate peroxidase. Phytochemistry 2005, 66, 1551–1559. [Google Scholar] [CrossRef]
- Dragišić Maksimović, J.; Bogdanović, J.; Maksimović, V.; Nikolic, M. Silicon modulates the metabolism and utilization of phenolic compounds in cucumber (Cucumis sativus L.) grown at excess manganese. J. Plant Nut. Soil Sci. 2007, 170, 739–744. [Google Scholar] [CrossRef]
- Feng, J.-P.; Shi, Q.-H.; Wang, X.-F. Effects of exogenous silicon on photosynthetic capacity and antioxidant enzyme activities in chloroplast of cucumber seedlings under excess manganese. Agric. Sci. China 2009, 8, 40–50. [Google Scholar] [CrossRef]
- Dragišić Maksimović, J.; Mojović, M.; Maksimović, V.; Römheld, V.; Nikolic, M. Silicon ameliorates manganese toxicity in cucumber by decreasing hydroxyl radical accumulation in the leaf apoplast. J. Exp. Bot. 2012, 63, 2411–2420. [Google Scholar] [CrossRef]
- Dragišić-Maksimović, J.; Mojović, M.; Maksimović, V. Silicon facilitates manganese phytoextraction by cucumber (Cucumis sativus L.). Zaštita materijala 2016, 57, 424–429. [Google Scholar] [CrossRef]
- Farooq, M.A.; Detterbeck, A.; Clemens, S.; Dietz, K.J. Silicon-induced reversibility of cadmium toxicity in rice. J. Exp. Bo.t 2016, 67, 3573–3585. [Google Scholar] [CrossRef]
- Feng, J.; Shi, Q.; Wang, X.; Wei, M.; Yang, F.; Xu, H. Silicon supplementation ameliorated the inhibition of photosynthesis and nitrate metabolism by cadmium (Cd) toxicity in Cucumis sativus L. Sci. Hort. 2010, 123, 521–530. [Google Scholar] [CrossRef]
- Wu, J.; Guo, J.; Hu, Y.; Gong, H. Distinct physiological responses of tomato and cucumber plants in silicon-mediated alleviation of cadmium stress. Front. Plant Sci. 2015, 6, 453. [Google Scholar] [CrossRef]
- Campos, C.N.S.; de Mello Prado, R.; Caione, G.; de Lima Neto, A.J.; Mingotte, F.L.C. Silicon and excess ammonium and nitrate in cucumber plants. Afr. J. Agric. Res. 2016, 11, 276–283. [Google Scholar]
- Barreto, R.F.; Prado, R.M.; Leal, A.J.F.; Troleis, M.J.B.; Junior, G.S.; Monteiro, C.C.; Santos, L.C.N.; Carvalho, R.F. Mitigation of ammonium toxicity by silicon in tomato depends on the ammonium concentration. Acta Agric. Scand. 2016, 66, 483–488. [Google Scholar] [CrossRef][Green Version]
- Al-aghabary, K.; Zhu, Z.; Shi, Q. Influence of silicon supply on chlorophyll content, chlorophyll fluorescence, and antioxidative enzyme activities in tomato plants under salt stress. J. Plant Nut. 2005, 27, 2101–2115. [Google Scholar] [CrossRef]
- Qian, Q.Q.; Zai, W.S.; Zhu, Z.J.; Yu, J.Q. Effects of exogenous silicon on active oxygen scavenging systems in chloroplasts of cucumber (Cucumis sativus L.) seedlings under salt stress. J. Plant Physiol. Mol. Biol. 2006, 32, 107–112. [Google Scholar]
- Qiongqiu, Q.; Wenshan, Z.; Yong, H. Protection of exogenous silicon and CoQ10 on mitochondria in cucumber (Cucumis sativus L.) roots under salt stress. Sci. Agric. Sinica 2006, 39, 1208–1214. [Google Scholar]
- Sun, Y.; Luo, W.; Zhang, W.; Zhou, X. Effects of exogenous silicon on germination characteristics of cucumber seeds under NaHCO3 stress. In Proceedings of the 2010 International Conference on Challenges in Environmental Science and Computer Engineering (CESCE), Wuhan, China, 6–7 March 2010; IEEE: Piscataway, NJ, USA, 2010; Volume 1, pp. 471–474. [Google Scholar]
- Lee, S.K.; Sohn, E.Y.; Hamayun, M.; Yoon, J.Y.; Lee, I.J. Effect of silicon on growth and salinity stress of soybean plant grown under hydroponic system. Agrofor. Syst. 2010, 80, 333–340. [Google Scholar] [CrossRef]
- Amirossadat, Z.; Ghehsareh, A.M.; Mojiri, A. Impact of silicon on decreasing of salinity stress in greenhouse cucumber (Cucumis sativus L.) in soilless culture. J. Biol. Environ. Sci. 2012, 6, 171–174. [Google Scholar]
- Arouiee, H.; Kafi, M.; Neamati, S.H. Effect of Silicon on Growth and Physiological Parameters in Fenugreek (Trigonella foenumgraceum L.) Under Salt Stress. Int. J. Agric. Crop Sci. 2012, 4, 1554–1558. [Google Scholar]
- Haghighi, M.; Afifipour, Z.; Mozafarian, M. The alleviation effect of silicon on seed germination and seedling growth of tomato under salinity stress. Veget. Crops Res. Bull. 2012, 76, 119–126. [Google Scholar] [CrossRef]
- Avcu, S.; Akhoundnejad, Y.; Dașgan, H.Y. Effects of selenium and silicon on salt-stressed tomato. TABAD Tarım Bilimleri Araștırma Dergisi 2013, 6, 183–188. [Google Scholar]
- Wang, H.-S.; Yu, C.; Fan, P.-P.; Bao, B.-F.; Li, T.; Zhu, Z.-J. Identification of two cucumber putative silicon transporter genes in Cucumis sativus. J. Plant Growth Regul. 2015, 34, 332–338. [Google Scholar] [CrossRef]
- Li, H.; Zhu, Y.; Hu, Y.; Han, W.; Gong, H. Beneficial effects of silicon in alleviating salinity stress of tomato seedlings grown under sand culture. Acta Physiol. Plant. 2015, 37, 71. [Google Scholar] [CrossRef]
- Zhu, Y.; Guo, J.; Feng, R.; Jia, J.; Han, W.; Gong, H. The regulatory role of silicon on carbohydrate metabolism in Cucumis sativus L. under salt stress. Plant Soil 2016, 406, 231–249. [Google Scholar] [CrossRef]
- Almutairi, Z.M. Effect of nano-silicon application on the expression of salt tolerance genes in germinating tomato (’Solanum lycopersicum’L.) seedlings under salt stress. Plant Omics 2016, 9, 106. [Google Scholar]
- Salah, M.H.; Al-Zahrani, H.S.; Metwali, E.M. Improving the Salinity Tolerance in Potato (Solanum tuberosum) by Exogenous Application of Silicon Dioxide Nanoparticles. Int. J. Agric. Biol. 2017, 19. [Google Scholar]
- Yunus, Q.; Zari, M. Effect of Exogenous Silicon on Ion Distribution of Tomato Plants Under Salt Stress. Commun. Soil Sci. Plant Anal. 2017, 48, 1843–1851. [Google Scholar] [CrossRef]
- Korkmaz, A.; Karagöl, A.; Akınoğlu, G.; Korkmaz, H. The effects of silicon on nutrient levels and yields of tomatoes under saline stress in artificial medium culture. J. Plant Nut. 2018, 41, 123–135. [Google Scholar] [CrossRef]
- Isfahani, F.M.; Tahmourespour, A.; Hoodaji, M.; Ataabadi, M.; Mohammadi, A. Influence of Exopolysaccharide-Producing Bacteria and SiO2 Nanoparticles on Proline Content and Antioxidant Enzyme Activities of Tomato Seedlings (Solanum lycopersicum L.) under Salinity Stress. Pol. J. Environ. Stud. 2018, 28, 153–163. [Google Scholar] [CrossRef]
- Silva, O.N.; Lobato, A.K.S.; Avila, F.W.; Costa, R.C.L.; Oliveira Neto, C.F.; Santos Filho, B.G.; Martins Filho, A.P.; Lemos, R.P.; Pinho, J.M.; Medeiros, M. Silicon-induced increase in chlorophyll is modulated by the leaf water potential in two water-deficient tomato cultivars. Plant Soil Environ. 2012, 58, 481–486. [Google Scholar] [CrossRef]
- Zhang, Y.; Yu, S.H.I.; Gong, H.; Zhao, H.; Li, H.; Hu, Y.; Wang, Y. Beneficial effects of silicon on photosynthesis of tomato seedlings under water stress. J. Integ. Agric. 2018, 17, 2151–2159. [Google Scholar] [CrossRef]
- Khodarahmi, S.; Khoshgoftarmanesh, A.H.; Mobli, M. Effect of silicon nutrition on alleviating cadmium toxicity-induced damage on cucumber (Cucumis sativus L.) at vegetative stage. J. Sci. Tech. Greenhouse Cult. 2012, 3, 103–110. [Google Scholar]
- Marmiroli, M.; Pigoni, V.; Savo-Sardaro, M.L.; Marmiroli, N. The effect of silicon on the uptake and translocation of arsenic in tomato (Solanum lycopersicum L.). Env. Exp. Bot. 2014, 99, 9–17. [Google Scholar] [CrossRef]
- Rahman, M.F.; Ghosal, A.; Alam, M.F.; Kabir, A.H. Remediation of cadmium toxicity in field peas (Pisum sativum L.) through exogenous silicon. Ecotoxicol. Environ. Safety 2017, 135, 165–172. [Google Scholar] [CrossRef]
- Ali, N.; Schwarzenberg, A.; Yvin, J.-C.; Hosseini, S.A. Regulatory Role of Silicon in Mediating Differential Stress Tolerance Responses in Two Contrasting Tomato Genotypes Under Osmotic Stress. Front. Plant Sci. 2018, 9, 1475. [Google Scholar] [CrossRef]
- Mitani, N.; Ma, J.F. Uptake system of silicon in different plant species. J. Exp. Bot. 2005, 56, 1255–1261. [Google Scholar] [CrossRef]
- Mitani, N.; Yamaji, N.; Ago, Y.; Iwasaki, K.; Ma, J.F. Isolation and functional characterization of an influx silicon transporter in two pumpkin cultivars contrasting in silicon accumulation. Plant J. 2011, 66, 231–240. [Google Scholar] [CrossRef]
- Mitani-Ueno, N.; Yamaji, N.; Ma, J.F. Silicon efflux transporters isolated from two pumpkin cultivars contrasting in Si uptake. Plant Signal. Behav. 2011, 6, 991–994. [Google Scholar] [CrossRef]
- Deshmukh, R.K.; Vivancos, J.; Guérin, V.; Sonah, H.; Labbé, C.; Belzile, F.; Bélanger, R.R. Identification and functional characterization of silicon transporters in soybean using comparative genomics of major intrinsic proteins in Arabidopsis and rice. Plant Mol. Biol. 2013, 83, 303–315. [Google Scholar] [CrossRef]
- Sun, H.; Guo, J.; Duan, Y.; Zhang, T.; Huo, H.; Gong, H. Isolation and functional characterization of CsLsi1, a silicon transporter gene in Cucumis sativus. Physiol. Plant. 2017, 159, 201–214. [Google Scholar] [CrossRef]


{kind=link}
| Vegetable Crop | Form of Silicon Applied | Observed Effect of Silicon | Reference |
|---|---|---|---|
| Fungal Pathogen | |||
| C. sativus | Silicate fertilizer | Promoted the growth and yield and also reduced the damage caused by wilt disease | [60] |
| Cucumis sativus | Soluble silicates | Reduced the size of fungal colonies (S. fuliginea) | [27] |
| C. sativus | Soluble silicon | Decreased the receptivity of plants to mildew infection caused by S. fuliginea | [61] |
| C. sativus. and Solanum lycopersicon | Potassium silicate | Reduced the infection caused by Pythium and F.solani | [28] |
| C. sativus | Potassium silicate added to hydroponic nutrient solutions | Suppressed powdery mildew (PM) caused by S.fuliginea | [62] |
| C. sativus | Soluble silicon | Significantly decreased the powdery mildew disease (caused by S. fuliginea) index | [29] |
| C. sativus | Potassium metasilicate | Significantly suppressed powdery mildew (P. xanthi) | [14] |
| C. melo | Sodium silicate and nanosized silicon | Significantly decreased the severity of mildew powder | [35] |
| S. lycopersicon and M. charantia | Silicic acid | Symplastic Si was associated with the reduction of the spread of the fungus (P. aphanidermatum) in roots | [31] |
| C. sativus | Significantly reduced the incidence of damping-off (P. aphanidermatum) | [32] | |
| C. melo | Sodium silicate | Reduced the postharvest rot (T. roseum) | [39] |
| C. sativus | Significantly decreased the disease index (S. fuliginea) | [63] | |
| C. melo | Potassium silicate | Reduced the severity and incidence of powdery mildew (S. fuliginea) | [36] |
| C. pepo | Potassium silicate | Enhanced the tolerance to salinity and resistance to powdery mildew (P. xanthii) | [40] |
| C. sativus | Sodium silicate | Reduced downy mildew (P. cubensis) disease index | [33] |
| C. annuum | Calcium silicate | Potentially reduce the severity of Phytophthora blight | [45] |
| C. sativus | Sodium silicate | Enhanced crop resistance to oxidative stress induced by P. melonis infection | [30] |
| S. lycopersicon | Sodium metasilicate nonahydrate | Reduced the disease severity of Fusarium crown and root rot (F. oxysporum f.sp. radicis-lycopersici) | [43] |
| G. max | Wollastonite | Controlled the soybean rust (P. pachyrhizi) | [49] |
| C. melo | Potassium silicate | Controlled the powdery mildew (P. xanthi) | [37] |
| C. sativus | Carbon Silpower solution | Inhibited powdery mildew (P. xanthi) development | [64] |
| G. max | Potassium silicate | Protected plants against soybean rust (P. pachyrhizi) | [48] |
| S. lycopersicon | Sodium silicate | Suppressed anthracnose disease (C. gloeosporiodes) | [44] |
| S. lycopersicon | Potassium silicate | Reduced the severity and incidence of Fusarium wilt (F. oxysporum f. sp. lycopersici) | [65] |
| S. lycopersicon | Potassium silicate | Reduced Fusarium wilt (F. oxysporum f. sp. lycopersici) | [66] |
| C. annuum | Potassium silicate | Enhanced resistance to anthracnose (Colletotrichum gloeosporioides) | [46] |
| S. lycopersicon | Sodium silicate | Controlled anthracanose disaese (C. gloeosporioides)and improved postharvest quality of fruits | [67] |
| M. charantia | Potassium silicate | Strengthened resistance in plants against powdery mildew (Erysiphe sp.) | [38] |
| C. pepo | Calcium silicate, CaMg silicate slag, wollastonite and MontanaGrowTM | Suppressed Powdery mildew (P. xanthii) | [41] |
| S. lycopersicon | Silicon rich rice hull | Enhanced anthracnose resistance (C. dematium) | [68] |
| C. sativus | Sodium silicate | Enhanced resistance to Fusarium wilt (F. oxysporum f. sp. cucumerinum Owen) and altered soil microbial communities | [34] |
| Bacterial pathogen | |||
| S. lycopersicon | Monosilicic acid | Acted as an inducer of resistance against R. solanacearum | [50] |
| S. lycopersicon | Significantly reduced the incidence of bacterial wilt (R. solanacearum) | [51] | |
| S. lycopersicon | Soluble silicon | Reduced wilt incidence ( R. solanacearum) | [53] |
| S. lycopersicon | Monosilicic acid and aerosol powder | Induced basal resistance against R. solanacearum | [42] |
| S. lycopersicon | Reduced severity and incidence of bacterial wilt (R. solanacearum) | [69] | |
| S. lycopersicon | Monosilicic acid | Induced resistance against R. solanacearum | [52] |
| S. lycopersicon | Monosilicilic acid | Induced resistance against bacterial wilt (R. solanacearum) | [70] |
| S. lycopersicon | Supa Sílica and calcium silicate | Reduced the symptoms of bacterial speck ( P. syringae pv. Tomato) | [71] |
| S. lycopersicon | Potassium silicate | Controlled R. solanacearum incidence by changing the soil microorganism amount and enzyme activity | [54] |
| Cucumis melo L. | Calcium silicate | Induced resistance against bacterial fruit blotch (A. citrulli) | [72] |
| S. lycopersicon | Monosilicic acid | Induced systemic resistance against bacterial wilt (R. solanacearum) | [73] |
| S. lycopersicon | Suppressed bacterial wilt (R. solanacearum) | [55] | |
| S. lycopersicon | Potassium silicate | Induced resistance against bacterial wilt (R. solanacearum) | [74] |
| Cucumis melo L. | Calcium silicate | Enhanced resistance to bacterial fruit blotch (A. citrulli) | [75] |
| Insect pest and nematodes | |||
| C. sativus | Calcium silicate | Acted as resistance Inducers against the Whitefly (B. tabaci) | [19] |
| G. max | Silicic acid | significantly decreased the Silverleaf whitefly populations | [56] |
| S. lycopersicon | AgrosilícioTM | Controlled leafminer (T. absoluta) owing to toxic and anti-feeding effect to the larval stage | [58] |
| C. sativus | Potassium silicate | Acted as an anti-herbivore defense | [57] |
| C. sativus | sodium metasilicate | Significantly reduced the activity of root-knot nematode (M. incognita) | [59] |
| Vegetable Crop | Form of Silicon Applied | Observed Effect of Silicon | Reference |
|---|---|---|---|
| Salinity | |||
| C. sativus | Potassium silicate | Alleviated salt stress and increased antioxidant enzymes activity | [83] |
| S. lycopersicon | Sodium silicate | Alleviated salt toxicity | [122] |
| S. lycopersicon | Potassium silicate | Alleviated the deleterious salt effect | [89] |
| C. sativus | Potassium silicate | Alleviated the salinity stress | [123,124] |
| Spinacia oleracea | Sodium silicate | Increased stress tolerance | [84] |
| C. sativus | Enhanced salinity tolerance | [125] | |
| M. charantia | Potassium silicate | Alleviated salt stress and increases antioxidant enzymes activity | [85] |
| G. max | Sodium metasilicate | Alleviated the detrimental effect of salinity stress | [126] |
| C. sativus | Sodium silicate | Increased resistance against salinity | [127] |
| Trigonella foenumgraceum | Sodium silicate | Increased the tolerance to salt stress | [128] |
| S. lycopersicon | Alleviated the effect of salinity stress | [129] | |
| S. lycopersicon | Silicon and nano silicon | Improved the salt tolerance | [82,130] |
| C. sativus | Sodium silicate | Alleviated salt-oxidative stress | [86,93] |
| Cucurbita pepo | nano-SiO2 | Improved the defense mechanisms of plants against salt stress toxicity | [92] |
| C. sativus | Silicic acid | Enhanced the salt tolerance | [131] |
| Pisum sativum | Potassium silicate | Alleviated the salinity-induced deleterious effects | [90] |
| C. annumn | Nano Silicon | Improved salinity tolerance | [91] |
| S. lycopersicon | Sodium silicate | Alleviated salinity stress | [132] |
| C. sativus | Sodium silicate | Alleviated oxidative damage and improved plant growth and photosynthetic performance | [95,133] |
| C. annuum | Potassium silicate | Mitigated salinity stress | [94] |
| S. lycopersicon | Nano-silicon | Regulated the expression of salt tolerance genes under salinity stress | [134] |
| S. tuberosum | Silicon dioxide nanoparticles | Improved the salinity tolerance | [135] |
| S. lycopersicon | Potassium silicate | Manipulated ion Distribution of plants under salt stress | [136] |
| S. lycopersicon | Silicic acid | Improved nutrient levels and yields under saline conditions | [137] |
| S. lycopersicon | Silicon nanoparticles | Enhanced salinity tolerance and improved plant growth with exopolysaccharide-producing bacteria | [138] |
| Drought | |||
| Capsicum annuum | Increased the tolerance to water deficit | [96] | |
| C. annuum | Sodium metasilicate | Alleviated negative effects of water deficiency | [97] |
| G. max | Sodium metasilicate | Alleviated seedling damage under drought and ultraviolet-B radiation | [88] |
| G. max | Silicic acid | Mitigated the adverse effects of salt and drought stress | [98] |
| S. lycopersicon | Sodium metasilicate | Increased total chlorophylls under water-deficient conditions | [139] |
| S. lycopersicon | Silicic acid | Improved seed germination and alleviated oxidative stress under water deficit stress | [99] |
| S. lycopersicon | Sodium silicate | Restrained chlorophyll degradation and increased optimal photosynthetic efficiency under drought stress | [100,101,103] |
| S. lycopersicon | Potassium silicate | Enhanced the water stress tolerance | [102,140] |
| Mineral toxicity | |||
| G. max | Soluble silicon | Alleviated the symptoms of Al toxicity | [108] |
| Vigna unguiculata | Potassium silicate | Alleviated Mn toxicity | [111] |
| C. sativus | Potassium silicate | Alleviated the Mn toxicity | [112] |
| C. sativus | Silicic acid | Alleviated Mn toxicity and modulated the metabolism and utilization of phenolic compounds | [113] |
| C. sativus | Sodium silicate | Alleviated the adverse effects of excess Mn and cadmium (Cd) toxicity | [114,118] |
| C. sativus | Improved antioxidant capacity of plant under Cd toxicity | [141] | |
| C. sativus | Silicic acid | Ameliorated manganese toxicity by decreasing hydroxyl radical accumulation | [115] |
| S. lycopersicon | Calcium silicate | Mitigated the inhibitory effects of arsenic | [142] |
| S. lycopersicon and C. sativus | Sodium metasilicate nonahydrate | Alleviated Cd stress | [119] |
| C. sativus | Sodium silicate | Alleviated autotoxicity caused by 3-phenyl propionic acid during seed germination | [87] |
| C. sativus | Silicic acid | Enhanced leaf remobilization of iron under limited iron conditions | [116] |
| C. sativus | Potassium silicate | Mitigated the toxicity of ammonium | [120] |
| S. lycopersicon | Monosilicic acid | Mitigated ammonium toxicity | [121] |
| C. sativus | Sodium silicate | Alleviated autotoxicity and Cd toxicity | [87] |
| C. sativus | Silicic acid | Mitigated the Al toxicity under acidic conditions | [109] |
| P. sativum | Orthosilicic acid | Alleviated Cd toxicity | [143] |
| S. tuberosum | Sodium silicate | Improved the defense ability against Al toxicity | [110] |
| Osmotic | |||
| C. sativus | Induced alleviation of growth reduction under osmotic stress | [104] | |
| C. sativus | Silicon spray | Enhanced the capacity of scavenging active oxygen species and improved photosynthesis | [105] |
| C. sativus | Sodium metasilicate | Contributed tolerance against osmotic stress | [106] |
| S. lycopersicon | Monosilicic acid | Regulated osmotic stress tolerance by differential accumulation of relevant amino acids | [144] |
| Cold | |||
| C. sativus | Potassium silicate | Provided chilling tolerance | [107] |
| UV-B | |||
| G. max | Sodium metasilicate | Enhanced nutrient acquisition under UV-B Radiation | [22,88] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kaushik, P.; Saini, D.K. Silicon as a Vegetable Crops Modulator—A Review. Plants 2019, 8, 148. https://doi.org/10.3390/plants8060148
Kaushik P, Saini DK. Silicon as a Vegetable Crops Modulator—A Review. Plants. 2019; 8(6):148. https://doi.org/10.3390/plants8060148
Chicago/Turabian StyleKaushik, Prashant, and Dinesh Kumar Saini. 2019. "Silicon as a Vegetable Crops Modulator—A Review" Plants 8, no. 6: 148. https://doi.org/10.3390/plants8060148
APA StyleKaushik, P., & Saini, D. K. (2019). Silicon as a Vegetable Crops Modulator—A Review. Plants, 8(6), 148. https://doi.org/10.3390/plants8060148