Mechanisms of Plant Tolerance to RNA Viruses Induced by Plant-Growth-Promoting Microorganisms



 and
and
Abstract
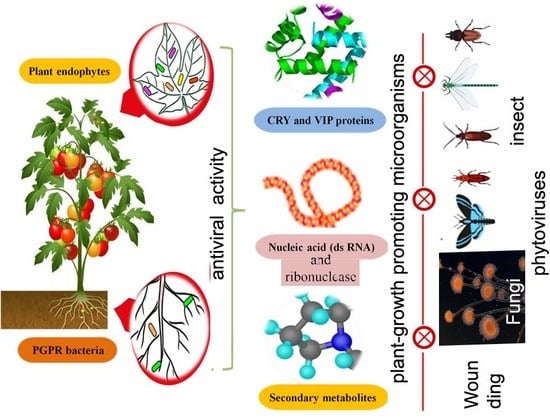
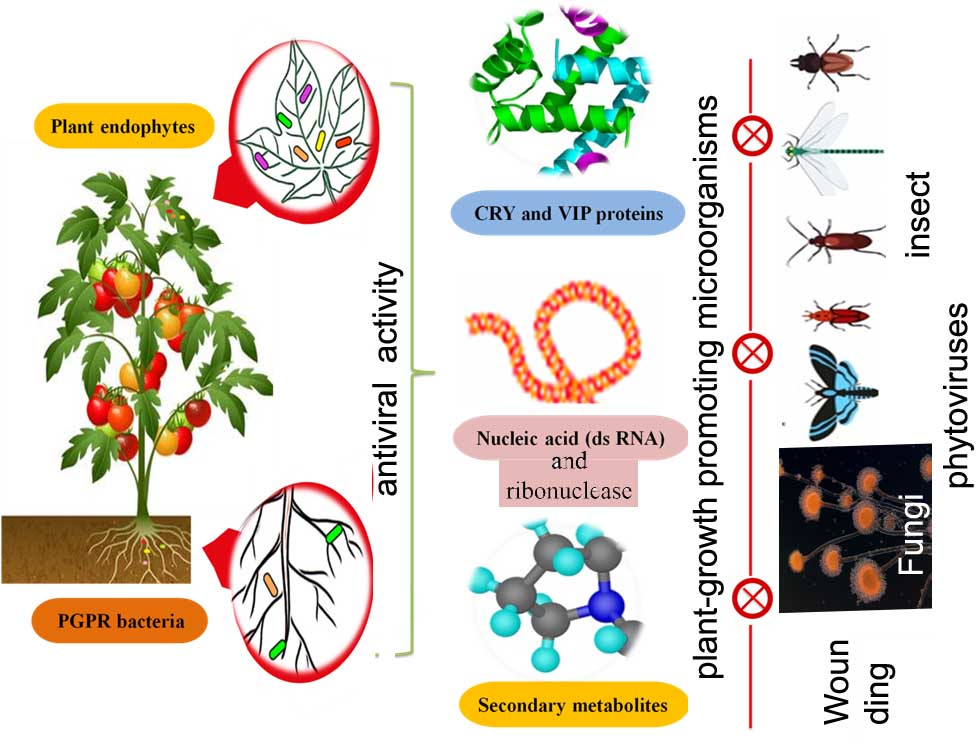
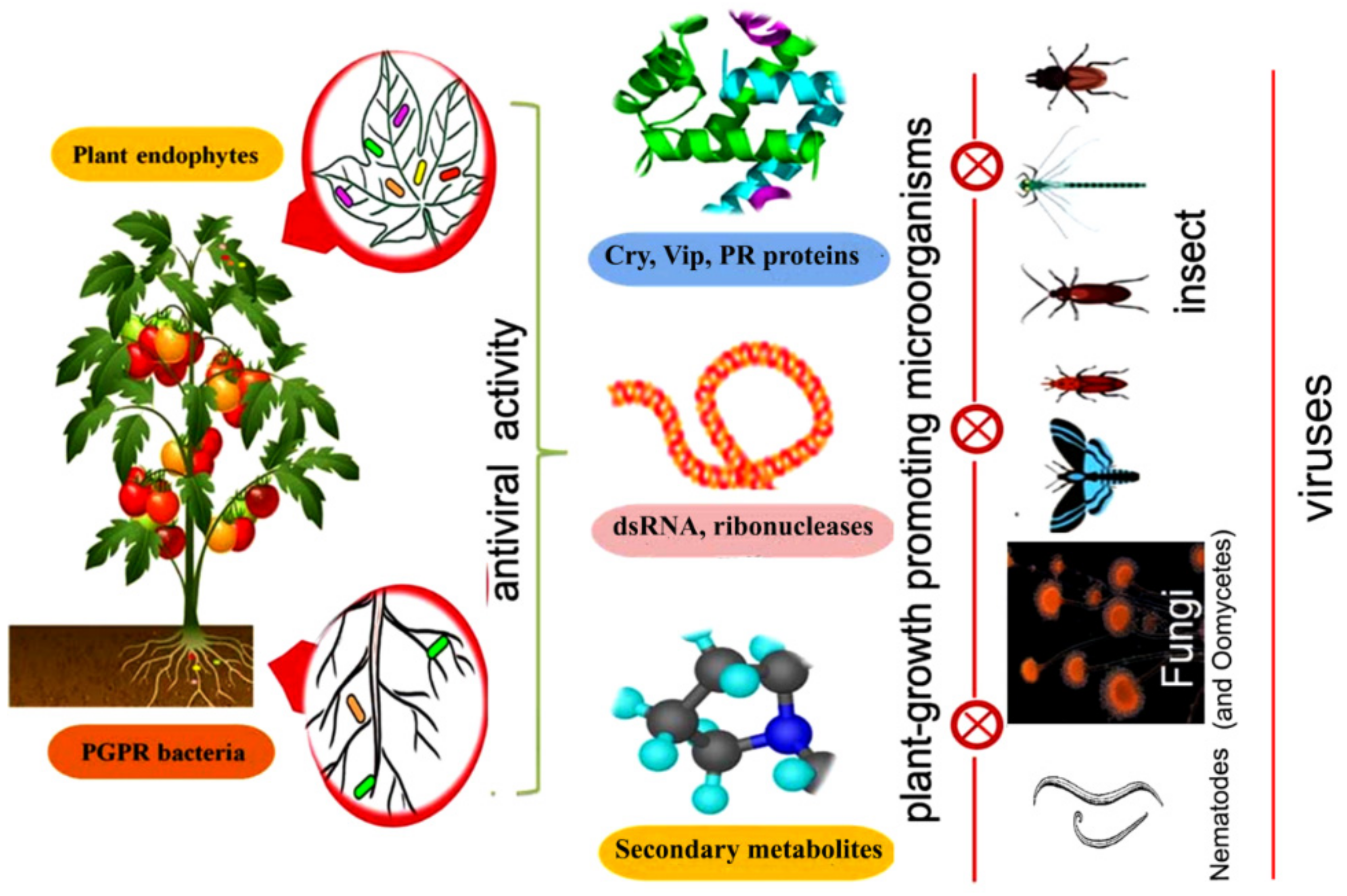
1. Introduction
2. Microorganisms as the Means of Biocontrol of Plant Viral Infections
3. Bacterial Nucleases and Their Antiviral Activity
4. Signal Pathways and Mechanisms of Plant Resistance to Viruses Induced by Microorganisms
4.1. Virus Recognition and Systemic Resistance in Plants
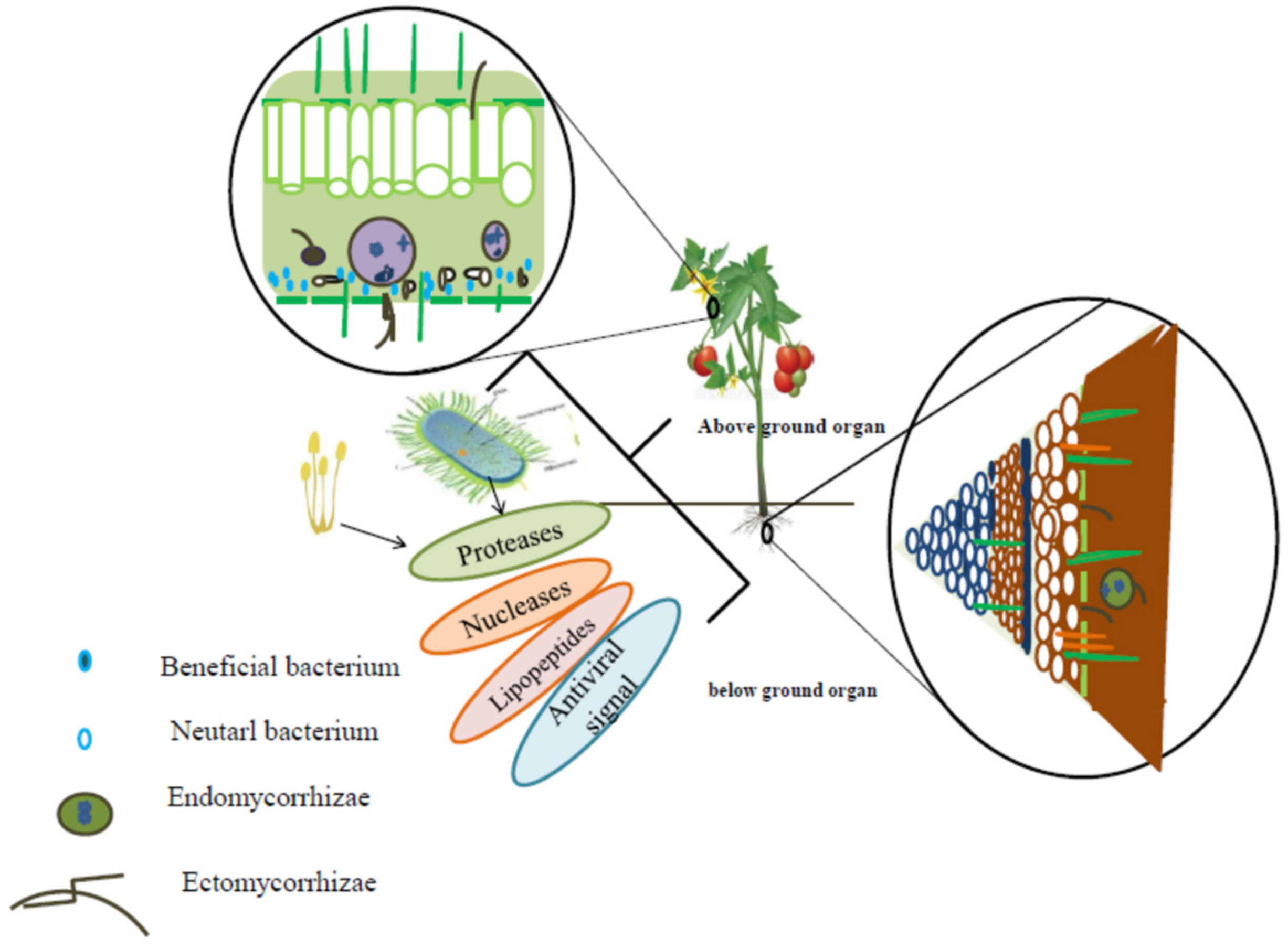
4.2. Plant-Growth Promoting Microorganism (PGPM) and Regulation of Plant Defense Mechanisms Against Viruses
5. Endophytic PGPM as Vectors of RNA Insecticides: Future Approaches
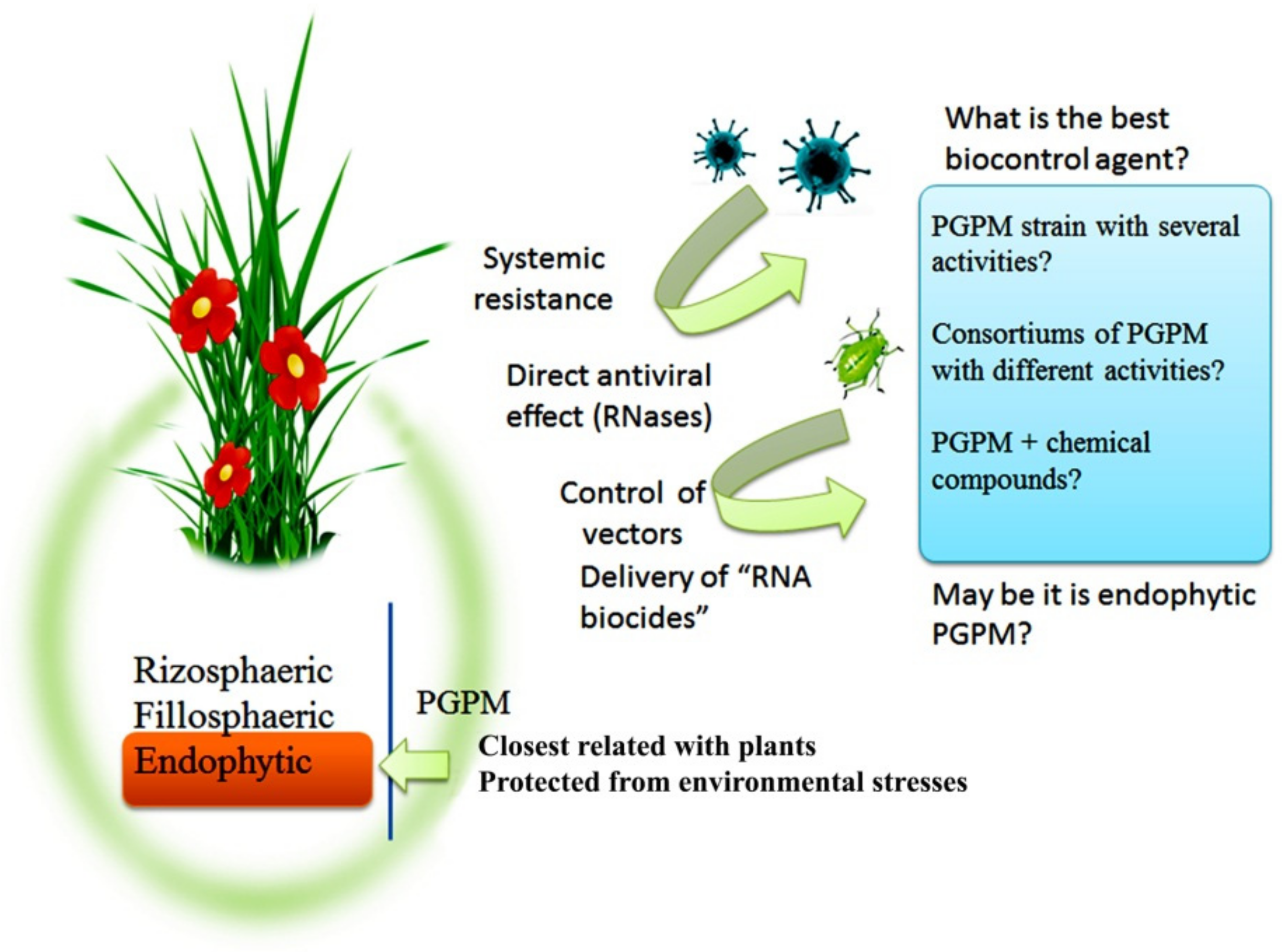
6. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Kunh, J.H. Classify viruses—the gaint is worth the pain. Nature 2019, 566, 318–320. [Google Scholar]
- Soosaar, J.L.; Burch-Smith, T.M.; Dinesh-Kumar, S.P. Mechanisms of plant resistance to viruses. Nat. Rev. Microbiol. 2005, 3, 789–798. [Google Scholar] [CrossRef] [PubMed]
- Makarova, S.S.; Makarov, V.V.; Taliansky, M.E.; Kalinina, N.O. Resistance to viruses of potato: Current status and prospects. Vavilov J. Genet. Breed. 2017, 21, 62–73. [Google Scholar] [CrossRef][Green Version]
- Borah, B.K.; Sharma, S.; Kant, R.; Johnson, A.M.A.; Venkata, D.; Saigopal, R.; Dasgupta, I. Bacilliform DNA-containing plant viruses in the tropics: Commonalities within a genetically diverse group. Mol. Plant Pathol. 2013, 14, 759–771. [Google Scholar] [CrossRef]
- Sanford, J.C.; Johnston, S.A. The concept of parasite-derived resistance–deriving reistance genes from the parasite’s own genome. J. Theor. Biol. 1985, 113, 395–405. [Google Scholar] [CrossRef]
- Prins, M.; Laimer, M.; Noris, E.; Schubert, J.; Wassenegger, M.; Tepfer, M. Strategies for anti-viral resistance in transgenic plants. Mol. Plant Pathol. 2008, 9, 73–83. [Google Scholar]
- Romay, G.; Bragard, C. Antiviral Defenses in Plants through Genome Editing. Front. Microbiol. 2017, 8, 47. [Google Scholar] [CrossRef]
- Arif, M.; Azhar, U.; Arshad, M.; Zafar, Y.; Mansoor, S.; Asad, S. Engineering broad-spectrum resistance against RNA viruses in potato. Transgenic Res. 2012, 21, 303–311. [Google Scholar] [CrossRef]
- Chung, B.N.; Yoon, J.Y.; Palukaitis, P. Engineered resistance in potato against potato leafroll virus, potato virus A and potato virus Y. Virus Genes. 2013, 47, 86–92. [Google Scholar] [CrossRef]
- Matthews, R.E.F. Plant Virology, 2nd ed.; Academic Press: Cambridge, MA, USA, 1981. [Google Scholar]
- Ali, X.X.; Keldish, M.A.; Pomazkov, U.I. A new carrier of potato virus X is the mushroom Phytophthora infestans (Mont.) De Bary. Agron. Livest. 2010, 3, 18–23. [Google Scholar]
- Desoignies, N.; Schramme, F.; Ongena, M.; Legrève, A. Systemic resistance induced by Bacillus lipopeptides in Beta vulgaris reduces infection by the rhizomania disease vector Polymyxa betae. Mol. Plant Pathol. 2013, 14, 416–421. [Google Scholar] [CrossRef] [PubMed]
- Andika, I.B.; Wei, S.; Cao, C.; Salaipeth, L.; Kondo, H.; Sun, L. Phytopathogenic fungus hosts a plant virus: A naturally occurring cross-kingdom viral infection. Proc. Natl. Acad. Sci. USA 2017, 114, 12267–12272. [Google Scholar] [CrossRef] [PubMed]
- Mascia, T.; Labarile, R.; Doohan, F.; Gallitelli, D. Tobacco mosaic virus infection triggers an RNAi-based response in Phytophthora infestans. Sci. Rep. 2019, 9, 2657. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://www.redoxagro.com/viricide-3639616.html (accessed on 4 December 2019).
- Available online: https://onzelivre.nl/disease-management.php (accessed on 4 December 2019).
- De Fazio, G.; Caner, J.; Vicente, M. Effect of virazole (ribavirin) on tomato spotted wilt virus in two systemic hosts, tomato and tobacco. Arch. Virol. 1980, 63, 305–309. [Google Scholar] [CrossRef]
- Kondakova, V.G.; Schuster, G. Elimination of Strawberry Mottle Virus and Strawberry Crinkle Virus from Isolated Apices of Three Strawberry Varieties by the Addition of 2,4-Dioxohexahydro-1,3,5-triazine (5-Azadihydrouracil) to the Nutrient Medium. J. Phytopathol. 1991, 132, 84–86. [Google Scholar] [CrossRef]
- Palukaitis, P.; Yoon, J.-Y.; Choi, S.-K.; Carr, J.P. Manipulation of induced resistance to viruses. Curr. Opin. Virol. 2017, 26, 141–148. [Google Scholar] [CrossRef]
- Kavino, M.; Harish, S.; Kumar, N.; Saravanakumar, D.; Samiyappan, R. Induction of systemic resistance in banana (Musa spp.) against Banana bunchy top virus (BBTV) by combining chitin with root-colonizing Pseudomonas fluorescens strain CHA0. Eur. J. Plant Pathol. 2008, 120, 353–362. [Google Scholar] [CrossRef]
- Mishra, S.; Jagadeesh, K.S.; Krishnaraj, P.U.; Prem, S. Biocontrol of tomato leaf curl virus (ToLCV) in tomato with chitosan supplemented formulations of Pseudomonas sp. Under field conditions. Aust. J. Crop Sci. 2014, 8, 347–355. [Google Scholar]
- Firmansyah, D.; Widodo Hidayat, S.H. Chitosan and Plant Growth Promoting Rhizobacteria Application to Control Squash mosaic virus on Cucumber Plants. Asian J. Plant Pathol. 2017, 11, 148–155. [Google Scholar]
- Yi, H.-S.; Yang, J.W.; Ryu, C.-M. ISR meets SA Routside: Additive action of the endophyte Bacillus pumilus INR7 and the chemical inducer, benzothiadiazole, on induced resistance against bacterial spotin field-grown pepper. Front. Plant Sci. 2013, 4, 122. [Google Scholar] [CrossRef]
- Beris, D.; Theologidis, I.; Skandalis, N.; Vassilakos, N. Bacillus amyloliquefaciens strain MBI600 induces salicylic acid-dependent resistance in tomato plants against Tomato spotted wilt virus and Potato virus Y. Sci. Rep. 2018, 8, 10320. [Google Scholar] [CrossRef] [PubMed]
- Clay, K. Fungal endophytes of grasses: A defensive mutualism between plants and fungi. Ecology 1988, 69, 10–16. [Google Scholar] [CrossRef]
- Siegel, M.R.; Latch, G.C.M. Expression of antifungal activity in agar culture by isolates of grass endophytes. Mycologia 1991, 83, 529–537. [Google Scholar] [CrossRef]
- Le Cocq, K.; Gurr, S.J.; Hirsch, P.R.; Mauchline, T.H. Exploitation of endophytes for sustainable agricultural intensification. Mol. Plant Pathol. 2017, 18, 469–473. [Google Scholar] [CrossRef]
- Bouizgarne, B. Bacteria for Plant Growth Promotion and Disease Management. In Bacteria in Agrobiology: Disease Management; Maheshwari, D.K., Ed.; Chapter 2; Springer: Berlin/Heidelberg, Germany, 2012; pp. 15–46. [Google Scholar]
- Yue, Q.; Miller, C.J.; White, J.F., Jr.; Richardson, M.D. Isolation and characterization of fungal inhibitors from Epichloë festucae. J. Agric. Food Chem. 2000, 48, 4687–4692. [Google Scholar] [CrossRef]
- Saritha, M.; Prasad, N.V.; Tollamadugu, K.V. The Status of Research and Application of Biofertilizers and Biopesticides: Global Scenario. In Recent Developments in Applied Microbiology and Biochemistry; Academic Press: Cambridge, MA, USA, 2019; pp. 195–207. [Google Scholar]
- Available online: https://agrow.agribusinessintelligence.informa.com/-/media/agri/agrow/ag-market-reviews-pdfs/supplements/agrow-biopesticides_2013.pdf (accessed on 05 December 2019).
- Available online: http://en.sibbio.ru/catalog/crop/bitoksibatsilin/ (accessed on 4 December 2019).
- Sudhakar, N.; Thajuddin, N.; Murugesana, K. Plant growth-promoting rhizobacterial mediated protection of tomato in the field against cucumber mosaic virus and its vector Aphis gossypii. Biocontrol Sci. Technol. 2011, 21, 367–386. [Google Scholar] [CrossRef]
- Muvea, A.M.; Subramanian, S.; Maniania, N.K.; Poehling, H.-M.; Ekesi, S.; Meyhöfer, R. Endophytic Colonization of Onions Induces Resistance Against Viruliferous Thrips and Virus Replication. Front. Plant Sci. 2018, 9, 1785. [Google Scholar] [CrossRef]
- Loebenstein, G.; Lovrekovich, L. Interference with tobacco mosaic virus local lesion formation in tobacco by injection heat-killed cells of Pseudomonas syringae. Virology 1966, 30, 587–591. [Google Scholar] [CrossRef]
- Mann, E.W. Inhibition of tobacco mosaic virus by a bacterial extract. Phytopathology 1969, 59, 658–662. [Google Scholar]
- Bergstrom, G.C.; Johnson, M.C.; Kuc, J. Effects of local infection of cucumber by Colletotrichum lagenarium, Pseudomonas lachrymans, or tobacco necrosis virus on systemic resistance to cucumber mosaic virus. Phytopathology 1982, 72, 922–926. [Google Scholar] [CrossRef]
- Vinodkumar, S.; Nakkeeran, S.; Renukadevi, P.; Mohankumar, S. Diversity and antiviral potential of rhizospheric and endophytic Bacillus species and phyto-antiviral principles against tobacco streak virus in cotton. Agric. Ecosyst. Environ. 2018, 267, 42–51. [Google Scholar]
- Wang, S.; Wu, H.; Qiao, J.; Ma, L.; Liu, J.; Xia, Y.; Gao, X. Molecular Mechanism of Plant Growth Promotion and Induced Systemic Resistance to Tobacco Mosaic Virus by Bacillus spp. J. Microbiol. Biotechnol. 2009, 19, 1250–1258. [Google Scholar] [CrossRef] [PubMed]
- Lian, L.; Xie, L.; Zheng, L.; Lin, Q. Induction of systemic resistance in tobacco against Tobacco mosaic virus by Bacillus spp. Biocontrol Sci. Technol. 2011, 21, 281–292. [Google Scholar] [CrossRef]
- Zeyruk, V.N.; Barkalov, A.G.; Tikhonova, L.V.; Bessonov, A.S.; Nazarov, N.M.; Paremsky, I.Y.; Korshunov, A.V.; Chernikov, V.I.; Masyuk, Y.; Marianovskaya, M.V.; et al. Method of Reproduction of Healthy Potato Plants. Patent of the Russian Federation No. 2001123454/13, 23 August 2001. [Google Scholar]
- Shankar, A.C.; Udaya, N.S.; Chandra, N.-R.S.; Kumar, H.B.; Reddy, M.S.; Niranjana, S.R.; Prakash, H.S. Rhizobacteria mediated resistance against the blackeye cowpea mosaic strain of bean common mosaic virus in cowpea (Vigna unguiculata). Pest Manag. Sci. 2009, 65, 1059–1064. [Google Scholar] [CrossRef] [PubMed]
- Murphy, J.F.; Zehnder, G.W.; Schuster, D.J.; Sikora, E.J.; Polston, J.E.; Kloepper, J.W. Plant Growth-Promoting Rhizobacterial Mediated Protection in Tomato Against Tomato mottle virus. Plant Dis. 2000, 84, 779–784. [Google Scholar] [CrossRef] [PubMed]
- Maurhofer, M.; Hase, C.; Meuwly, P.; Métraux, J.P.; Défago, G. Induction of systemic resistance of tobacco to tobacco necrosis virus by the root-colonizing Pseudomonas fluorescens strain CHA0: Influence of the gacA gene and of pyoverdine production. Phytopathology 1994, 84, 139–146. [Google Scholar] [CrossRef]
- Karthikeyan, G.; Doraisamy, S.; Rabindran, R. Pseudomonas fluorescens mediated systemic resistance against urdbean leaf crinkle virus in blackgram (Vigna mungo). Arch. Phytopathol. Plant Protect. 2009, 42, 201–212. [Google Scholar] [CrossRef]
- Maurhofer, M.; Reimmann, C.; Sacherer, S.P.; Heeb, S.; Haas, D.; Defago, G. Salicylic acid biosynthetic genes expressed in Pseudomonas fluorescens strain P3 improve the induction of systemic resistance in tobacco against Tobacco necrosis virus. Phytopathology 1998, 88, 678–684. [Google Scholar] [CrossRef]
- Damayanti, T.A.; Katerina, T. Protection of hot pepper against multiple infections of viruses by utilizing root colonizing bacteria. J. ISSAAS 2008, 14, 92–100. [Google Scholar]
- Lee, G.; Lee, S.H.; Kim, K.M.; Ryu, C.M. Foliar application of the leaf-colonizing yeast Pseudozyma churashimaensis elicits systemic defense of pepper against bacterial and viral pathogens. Sci. Rep. 2017, 10, 39432. [Google Scholar] [CrossRef]
- Kumar, S.; Chauhan, P.S.; Agrawal, L.; Raj, R.; Srivastava, A.; Gupta, S.; Mishra, S.K.; Yadav, S.; Singh, P.C.; Raj, S.K.; et al. Paenibacillus lentimorbus Inoculation Enhances Tobacco Growth and Extenuates the Virulence of Cucumber mosaic virus. PLoS ONE 2016, 11, e0149980. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://minsemlab.ru/agrobakter/biogran (accessed on 4 December 2019).
- Yang, J.; Guo, C.; Zhai, X.; Shen, L.; Qian, Y.; Wang, F. Inactivation of Tobacco mosaic virus in soil by Pseudomonas putida A3-m strain to prevent virus mosaic disease. Afr. J. Microbiol. Res. 2012, 6, 6300–6307. [Google Scholar] [CrossRef]
- Sharipova, M.; Rockstroh, A.; Balaban, N.; Mardanova, A.; Toymentseva, A.; Tikhonova, A.; Vologin, S.; Stashevsky, Z. Antiviral Effect of Ribonuclease from Bacillus pumilus against phytopathogenic RNA-Viruses. Agric. Sci. 2015, 6, 1357–1366. [Google Scholar]
- Zehnder, G.W.; Yao, C.; Murphy, J.F.; Sikora, E.J.; Kloepper, J.W. Induction of resistance in tomato against Cucumber mosaic cucumovirus by plant growth-promoting rhizobacteria. Biolcontrol 2000, 45, 127–137. [Google Scholar] [CrossRef]
- Su, P.; Tan, X.; Li, C.; Zhang, D.; Cheng, J.; Zhang, S.; Zhou, X.; Yan, Q.; Peng, J.; Zhang, Z.; et al. Photosynthetic bacterium Rhodopseudomonas palustris GJ-22 induces systemic resistance against viruses. Microb. Biotechnol. 2017, 10, 612–624. [Google Scholar] [CrossRef]
- Ryu, C.M.; Murphy, J.F.; Mysore, K.S.; Kloepper, J.W. Plant growth-promoting rhizobacteria systemically protect Arabidopsis thaliana against Cucumber mosaic virus by a salicylic acid and NPR1-independent and jasmonic acid-dependent signaling pathway. Plant J. 2004, 39, 381–392. [Google Scholar] [CrossRef]
- Lee, G.H.; Ryu, C.M. Spraying of Leaf-Colonizing Bacillus amyloliquefaciens Protects Pepper from Cucumber mosaic virus. Plant Dis. 2016, 100, 2099–2105. [Google Scholar] [CrossRef]
- Jetiyanon, K.; Fowler, W.; Kloepper, J.W. Broad-spectrum protection against several pathogens by PGPR mixtures under field conditions in Thailand. Plant Dis. 2003, 87, 1390–1394. [Google Scholar] [CrossRef]
- Raupach, G.S.; Liu, L.; Murphy, J.F.; Tuzun, S.; Kloepper, J.W. Induced systemic resistance in cucumber and tomato against cucumber mosaic cucumovirus using plant growth-promoting rhizobacteria (PGPR). Plant Dis. 1996, 80, 891–894. [Google Scholar] [CrossRef]
- Murphy, J.F.; Reddy, M.S.; Ryu, C.M.; Kloepper, J.W.; Li, R. Rhizobacteria mediated growth promotion of tomato leads to protection against Cucumber mosaic virus. Phytopathology 2003, 93, 1301–1307. [Google Scholar] [CrossRef]
- Srinivasan, K.; Mathivanan, N. Biological control of sunflower necrosis virus disease with powder and liquid formulations of plant growth promoting microbial consortia under field conditions. Biol. Control 2009, 51, 395–402. [Google Scholar] [CrossRef]
- Abdalla, O.A.; Bibi, S.; Zhang, S. Application of plant growth-promoting rhizobacteria to control Papaya ringspot virus and Tomato chlorotic spot virus. Arch. Phytopathol. Plant Prot. 2017, 50, 584–597. [Google Scholar] [CrossRef]
- Harish, S.; Kavino, M.; Kumar, N.; Saravanakumar, D.; Soorianathasundaram, K.; Samiyappan, R. Biohardening with plant growth promoting rhizosphere and endophytic bacteria induces systemic resistance against Banana bunchy top virus. Appl. Soil Ecol. 2008, 39, 187–200. [Google Scholar] [CrossRef]
- Ulyanova, V.; Vershinina, V.; Ilinskaya, O. Barnase and binase: Twins with distinct fates. FEBS J. 2011, 278, 3633–3643. [Google Scholar] [CrossRef]
- Ilinskaya, O.; Ulyanova, V.; Lisevich, I.; Dudkina, E.; Zakharchenko, N.; Kusova, A.; Faizullin, D.; Zuev, Y. The Native Monomer of Bacillus pumilus Ribonuclease Does Not Exist Extracellularly. BioMed Res. Int. 2018, 2018, 4837623. [Google Scholar] [CrossRef]
- Khalaf, E.M.; Raizada, M.N. Bacterial Seed Endophytes of Domesticated Cucurbits Antagonize Fungal and Oomycete Pathogens Including Powdery Mildew. Front Microbiol. 2018, 9, 42. [Google Scholar] [CrossRef]
- Nakamura, A.; Koide, Y.; Miyazaki, H.; Kitamura, A.; Masaki, H.; Beppu, T.; Uozumi, T. Gene cloning and characterization of a novel extracellular ribonuclease of Bacillus subtilis. Eur. J. Biochem. 1992, 209, 121–127. [Google Scholar] [CrossRef]
- Hahnen, E.; Znamenskaya, L.; Koczan, D.; Leshchinskaya, I.; Hobom, G. A novel secreted ribonuclease from Bacillus intermedius: Gene structure and regulatory control. Mol. Gen. Genet. 2000, 263, 571–580. [Google Scholar] [CrossRef]
- Trifonova, E.A.; Ibragimova, S.M.; Volkova, O.A.; Shumny, V.K.; Kochetov, A.V. Ribonuclease activity as a new prospective disease resistance marker in potato. Vavilov J. Genet. Breed. 2018, 22, 987–991. [Google Scholar] [CrossRef]
- Fedorova, A.A.; Azzami, K.; Ryabchikova, E.I.; Spitsyna, Y.E.; Silnikov, V.N.; Ritter, W.; Gross, H.J.; Tautz, J.; Vlassov, V.V.; Beier, H.; et al. Inactivation of a non-enveloped RNA virus by artificial ribonucleases: Honey bees and acute bee paralysis virus as a new experimental model for in vivo antiviral activity assessment. Antivir. Res. 2011, 91, 267–277. [Google Scholar] [CrossRef]
- Zhou, W.W.; Niu, T.G. Purification and some properties of an extracellular ribonuclease withantiviral activity against tobacco mosaic virus from Bacillus cereus. Biotechnol. Lett. 2009, 31, 101–105. [Google Scholar] [CrossRef] [PubMed]
- Trifonova, E.A.; Komarova, M.L.; Leonova, N.S.; Shcherban’, A.B.; Kochetov, A.V.; Malinovskii, V.I.; Shumnyi, V.K. Transgenic Potato (Solanum tuberosum L.) Plants Expressing the Gene of Secretory Nuclease from Serratia marcescens. Dokl. Biochem. Biophys. 2004, 394, 39–41. [Google Scholar] [PubMed]
- Zhirnov, I.V.; Trifonova, E.A.; Romanova, A.V.; Filipenko, E.A.; Kochetov, A.V.; Shumny, V.K.; Sapotsky, M.V.; Malinovsky, V.I. Induced expression of Serratia marcescens ribonuclease III gene in transgenic Nicotiana tabacum L. cv. SR1 tobacco plants. Russ. J. Genet. 2016, 52, 1137–1141. [Google Scholar] [CrossRef]
- Cao, X.; Lu, Y.; Di, D.; Zhang, Z.; Liu, H.; Tian, L.; Zhang, A.; Zhang, Y.; Shi, L.; Guo, B.; et al. Enhanced virus resistance in transgenic maize expressing a dsRNA-specific endoribonuclease gene from E. coli. PLoS ONE 2013, 8, e60829. [Google Scholar] [CrossRef]
- Zhang, L.; French, R.; Langenberg, W.G.; Mitra, A. Accumulation of barley stripe mosaic virus is significantly reduced in transgenic wheat plants expressing a bacterial ribonuclease. Transgenic Res. 2001, 10, 13–19. [Google Scholar] [CrossRef]
- Yang, X.; Niu, L.; Zhang, W.; He, H.; Yang, J.; Xing, G.; Guo, D.; Zhao, Q.; Zhong, X.; Li, H.; et al. Increased multiple virus resistance in transgenic soybean overexpressing the double-strand RNA-specific ribonuclease gene PAC1. Transgenic Res. 2019, 28, 129–140. [Google Scholar] [CrossRef] [PubMed]
- Aman, R.; Ali, Z.; Butt, H.; Mahas, A.; Aljedaani, F.; Khan, M.Z.; Ding, S.; Mahfouz, M. RNA virus interference via CRISPR/Cas13a system in plants. Genome Biol. 2018, 19, 1. [Google Scholar] [CrossRef]
- Pakniat-Jahromy, A.; Behjatnia, S.A.; Dry, I.B.; Izadpanah, K.; Rezaian, M.A. A new strategy for generating geminivirus resistant plants using a DNA betasatellite/split barnase construct. J. Virol. Methods. 2010, 170, 57–66. [Google Scholar] [CrossRef]
- Chikh-Ali, M.; Rowley, J.S.; Kuhl, J.; Gray, S.M.; Karasev, A.V. Evidence of a Monogenic Nature of the Nz Gene Conferring Resistance Against Potato virus Y Strain Z (PVYZ) in Potato. Am. J. Potato Res. 2014, 91, 649. [Google Scholar] [CrossRef]
- Glais, L.; Bellstedt, D.U.; Lacomme, C. Diversity, Characterization and Classification of PVY. In Potato Virus Y: Biodiversity, Pathogenicity, Epidemiology and Management; Lacomme, C., Glais, L., Bellstedt, D., Dupuis, B., Karasev, A., Jacquot, E., Eds.; Springer: Cham, Switzerland, 2017; pp. 43–76. [Google Scholar]
- Niehl, A.; Wyrsch, I.; Boller, T.; Heinlein, M. Double-stranded RNAs induce a pattern-triggered immune signaling pathway in plants. New Phytol. 2016, 211, 1008–1019. [Google Scholar] [CrossRef]
- Zvereva, A.S.; Golyaev, V.; Turco, S.; Gubaeva, E.G.; Rajeswaran, R.; Schepetilnikov, M.V.; Srour, O.; Ryabova, L.A.; Boller, T.; Pooggin, M.M. Viral protein suppresses oxidative burst and salicylic acid dependent autophagy and facilitates bacterial growth on virus-infected plants. New Phytol. 2016, 211, 1020–1034. [Google Scholar] [CrossRef] [PubMed]
- Duan, C.G.; Wang, C.H.; Guo, H.S. Application of RNA silencing to plant disease resistance. Silence 2012, 3, 5. [Google Scholar] [CrossRef] [PubMed]
- Choudhary, D.K.; Johri, B.N. Interactions of Bacillus spp. and plants—with special reference to induced systemic resistance (ISR). Microbiol. Res. 2009, 68, 1754–1759. [Google Scholar] [CrossRef] [PubMed]
- Maksimov, I.V.; Veselova, S.V.; Nuzhnaya, T.V.; Sarvarova, E.R.; Khairullin, R.M. Plant growth-promoting bacteria in the regulation of plant resistance to stress factors. Russ. J. Plant Physiol. 2015, 62, 715–726. [Google Scholar] [CrossRef]
- Mhlongo, M.I.; Piater, L.A.; Madala, N.E.; Labuschagne, N.; Dubery, I.A. The Chemistry of Plant–Microbe Interactions in the Rhizosphere and the Potential for Metabolomics to Reveal Signaling Related to Defense Priming and Induced Systemic Resistance. Front. Plant Sci. 2018, 9, 112. [Google Scholar] [CrossRef]
- Zvereva, A.S.; Pooggin, M.M. Silencing and Innate Immunity in Plant Defense against Viral and Non-Viral Pathogens. Viruses 2012, 4, 2578–2597. [Google Scholar] [CrossRef]
- Boris, K.V.; Kochieva, E.Z. NBS-LRR Resistance Genes to Potato Virus X. Biol. Bull. Rev. 2013, 133, 124–132. [Google Scholar] [CrossRef]
- Sorokina, E.V. Toll-like receptors and primary pathogen recognition in infectious and non-infectious cutaneous pathology. Int. J. Immunopathol. Allergol. Infectol. 2012, 2, 6–15. [Google Scholar]
- Guevara-Morato, M.A.; de Lacoba, M.G.; García-Luque, I.; Serra, M.T. Characterization of a pathogenesis-related protein 4 (PR-4) induced in Capsicum chinense L3 plants with dual RNA-ase and DNA-ase activities. J. Exp. Bot. 2010, 61, 3259–3271. [Google Scholar] [CrossRef]
- Bai, S.; Dong, C.; Li, B.; Dai, H. A PR-4 gene identified from Malus domestica is involved in the defense responses against Botryosphaeria dothidea. Plant Physiol. Biochem. 2013, 62, 23–32. [Google Scholar] [CrossRef]
- Kandan, A.; Radja Commare, R.; Nandakumar, R.; Ramiah, M.; Raguchander, T.; Samiyappan, R. Induction of phenylpropanoid metabolism by Pseudomonas fluorescens against Tomato spotted wilt virus in tomato. Folia Microbiol. 2002, 47, 121–129. [Google Scholar] [CrossRef]
- Zarate, S.; Kempema, L.; Walling, L.L. Silverleaf Whitefly Induces Salicylic Acid Defenses and Suppresses Effectual Jasmonic Acid Defenses. Plant Physiol. 2007, 143, 866–875. [Google Scholar] [CrossRef] [PubMed]
- Yun, D.C.; Yang, S.Y.; Kim, Y.C.; Kim, I.S.; Kim, Y.H. Identification of surfactin as an aphicidal metabolite produced by Bacillus amyloliquefaciens G1. J. Korean Soc. Appl. Biol. Chem. 2013, 56, 751–753. [Google Scholar] [CrossRef]
- Hussein, W.; Awad, H.; Fahim, S. Systemic Resistance Induction of Tomato Plants against ToMV Virus by Surfactin Produced from Bacillus subtilis BMG02. Am. J. Microbiol. Res. 2016, 4, 153–158. [Google Scholar]
- Djavaheri, M.; Mercado-Blanco, J.; Versluis, C.; Meyer, J.M.; Loon, L.C.; Bakker, P.A. Iron-regulated metabolites produced by Pseudomonas fluorescens WCS374r are not required for eliciting induced systemic resistance against Pseudomonas syringae pv. tomato in Arabidopsis. Microbiol. Open. 2012, 1, 311–325. [Google Scholar] [CrossRef]
- Xie, S.; Jiang, H.; Ding, T.; Xu, Q.; Chai, W.; Cheng, B. Bacillus amyloliquefaciens FZB42 represses plant miR846 to induce systemic resistance via a jasmonic acid-dependent signalling pathway. Mol. Plant Pathol. 2018, 19, 1612–1623. [Google Scholar] [CrossRef]
- Nazari, F.; Safaie, N.; Soltani, B.M.; Shams-Bakhsh, M.; Sharifi, M. Bacillus subtilis affects miRNAs and flavanoids production in Agrobacterium-Tobacco interaction. Plant Physiol Biochem. 2017, 118, 98–106. [Google Scholar] [CrossRef]
- Niu, D.; Xia, J.; Jiang, C.; Qi, B.; Ling, X.; Lin, S.; Zhang, W.; Guo, J.; Jin, H.; Zhao, H. Bacillus cereus AR156 primes induced systemic resistance by suppressing miR825/825* and activating defense-related genes in Arabidopsis. J. Integr. Plant Biol. 2016, 58, 426–439. [Google Scholar] [CrossRef]
- Kong, H.G.; Shin, T.S.; Kim, T.H.; Ryu, C.-M. Stereoisomers of the Bacterial Volatile Compound 2,3-Butanediol Differently Elicit Systemic Defense Responses of Pepper against Multiple Viruses in the Field. Front. Plant Sci. 2018, 9, 90. [Google Scholar] [CrossRef]
- Pumplin, N.; Voinnet, O. RNA silencing suppression by plant pathogens: Defence, counter-defence and counter-counter-defence. Nat. Rev. Microbiol. 2013, 11, 745–760. [Google Scholar] [CrossRef]
- Zhan, X.; Zhang, F.; Zhong, Z.; Chen, R.; Wang, Y.; Chang, L.; Bock, R.; Nie, B.; Zhang, J. Generation of virus-resistant potato plants by RNA genome targeting. Plant Biotechnol. J. 2019, 17, 1814–1822. [Google Scholar] [CrossRef]
- Fuentes, A.; Carlos, N.; Ruiz, Y.; Callard, D.; Sánchez, Y.; Ochagavía, M.E.; Seguin, J.; Malpica-López, N.; Hohn, T.; Lecca, M.R. Field trial and molecular characterization of RNAi-transgenic tomato plants that exhibit resistance to tomato yellow leaf curl geminivirus. Mol. Plant Microbe Interact. 2016, 29, 197–209. [Google Scholar] [CrossRef] [PubMed]
- Whyard, S.; Singh, A.D.; Wong, S. Ingested double-stranded RNAs can act as species-specific insecticides. Insect Biochem. Mol. Biol. 2009, 39, 824–832. [Google Scholar] [CrossRef] [PubMed]
- Huvenne, H.; Smagghe, G. Mechanisms of dsRNA uptake in insects and potential of RNAi for pest control: A review. J. Insect Physiol. 2010, 56, 227–235. [Google Scholar] [CrossRef] [PubMed]
- Whitten, M.M.A.; Facey, P.D.; Sol, R.D.; Fernández-Martínez, L.T.; Evans, M.C.; Mitchell, J.J.; Bodger, O.G.; Dyson, P.J. Symbiont-mediated RNA interference in insects. Proc. R. Soc. B. 2016, 283, 20160042. [Google Scholar] [CrossRef] [PubMed]
- Available online: http://www.motherjones.com/tom-philpott/2015/08/coming-farm-field-near-you-gene-silencing-pesticides-rna-rnai (accessed on 4 December 2019).
- Available online: http://gogreenpestcontrol.ca/rna-insecticide-could-target-specific-pests (accessed on 4 December 2019).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Strain of PGPM | Plants | Method of Treatment | PGPM Effectiveness Against Viruses | Source |
|---|---|---|---|---|
| Pseudomonas syringae (heat-killed cells) | Nicotiana tabacum | Foliar spraying | Tobacco mosaic virus (TMV) | [35] |
| Bacillus uniflagellatus | Nicotiana tabacum | Soil drench | TMV | [36] |
| Pseudomonas lachrymans | Cucumis sativus | Foliar spraying | Cucumber mosaic virus (CMV) | [37] |
| Bacillus spp. | Gossypium herbaceum | Soil drench/foliar spray | Tobacco streak virus (TSV) | [38] |
| B. amyloliquefaciens MBI600 | Solanum lycopersicum/Solanum tuberosum | Foliar spraying | Tomato spotted wilt virus (TSWV) | [24] |
| B. amyloliquefaciens FZB24, FZB42 | Nicotiana tabacum | Soaking the roots | TMV | [39] |
| B. pumilus EN16 | Nicotiana tabacum | Soaking the roots | TMV | [40] |
| B. subtilis SW1 | Nicotiana tabacum | Soaking the roots | TMV | [40] |
| B. subtilis Ch13 | Solanum tuberosum | Micro-tubers spraying | PVY, potato virus X (PVX) | [41] |
| Bacillus pumilus T4 | Vigna unguiculata | Seed spraying | Bean Common Mosaic Virus (BCMV) | [42] |
| Bacillus subtilis GBO3 | Vigna unguiculata | Seed spraying | BCMV | [42] |
| B. pumilus SE34 | Solanum lycopersicum | Seed spraying | CMV | [43] |
| P. fluorescens CHA0 | Nicotiana tabacum | Foliar spraying | Tobacco necrosis virus (TNV) | [44] |
| P. fluorescens CHA0 | Vigna mungo | Foliar spraying | Urdbean leaf crinkle virus (ULCV) | [45] |
| P. fluorescens P3 | Nicotiana tabacum | Foliar spraying | TNV | [46] |
| Bacillus cereus (I-35), Stenotrophomonas sp. (II-10) | Capsicum annuum | Seed treatment and soil drench | TMV virus, Chili veinal mottle virus (ChiVMV) | [47] |
| Pseudozyma churashimaensis | Capsicum annuum | Soil drench | CMV, pepper mottle virus (PMV), pepper mild mottle virus (PMMV), and Broad bean wilt virus (BBWV) | [48] |
| Paenibacillus lentimorbus B-30488 | Nicotiana tabacum | Soil drench | CMV | [49] |
| Azotobacter vinelandii, Azotobacter chroococcum | Solanum tuberosum | Tuber drench | PVY, PVX, PLRV | [50] |
| P. putida A3 | Nicotiana tabacum | Soil drench | TMV | [51] |
| B. pumilus | Nicotiana tabacum | Leaves juice | TMV (destroying viral particles) | [52] |
| Bacillus pumilus SE34, Kluyvera cryocrescens IN114, Bacillus amyloliquefaciens IN937a, Bacillus subtilus IN937b | Solanum lycopersicum | Soil drench | CMV | [53] |
| Rhodopseudomonas palustris GJ-22 | Nicotiana tabacum | Seed treatment | TMV | [54] |
| S. marcescens 90-166 | Arabidopsis thaliana | Seed treatment | CMV | [55] |
| B. amyloliquefaciens 5B6 | Capsicum annuum | Foliar treatment | CMV | [56] |
| Microbial consortiums | ||||
| Bacillus subtilis GB03 + B. pumilus SE34/B. amyloliquefaciens IN937a/B. subtilis IN937b/B. pumilus INR7/B. pumilus T4 | Solanum lycopersicum | Seed treatment | CMV | [57] |
| Serattia marcescens 90-166 + P. putida 89B-61/B. pumilus SE34 | Cucumis sativus | Seed spraying | CMV | [58,59] |
| Bacillus licheniformis MML2501 + Bacillus sp. MML2551 + Pseudomonas aeruginosa MML2212 + Streptomyces fradiae MML1042 | Helianthus annuus | Seed treatment | Sunflower necrosis virus disease (SNVD) | [60] |
| B. amyloliquefaciens IN937a + B. pumilus SE34 + B. pumilus T4 | Carica papaya /Solanum lycopersicum | Seed treatment | Papaya ringspot virus (PRSV-W) /Tomato chlorotic spot virus (TCSV) | [61] |
| P. fluorescens Pf1. + Bacillus spp. EPB22 incidence with a final efficiency of up to 80% | Musa spp | Foliar treatment | Banana bunchy top virus (BBTV) | [62] |
| Individual strain/microbial consortiums + chemical immunoregulators | ||||
| P. fluorescens CHAO+chitin | Musa spp. | Soil drench | BBTV | [20] |
| Pseudomonas sp. (206 (4) + B-15 + JK-16 + chitosan olygomers | Solanum lycopersicum | Seed treatment | Tomato leaf curl virus (ToLCV) | [21] |
| Bacillus polymixa + Pseudomonas fluorescens + chitin olygomers | Cucumis sativus | Seed treatment | Squash mosaic virus (SqMV) | [22] |
| B. pumulus INR7 + benzothiadiazole | Capsicum annuum | Foliar treatment | CMV | [23] |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Maksimov, I.V.; Sorokan, A.V.; Burkhanova, G.F.; Veselova, S.V.; Alekseev, V.Y.; Shein, M.Y.; Avalbaev, A.M.; Dhaware, P.D.; Mehetre, G.T.; Singh, B.P.; et al. Mechanisms of Plant Tolerance to RNA Viruses Induced by Plant-Growth-Promoting Microorganisms. Plants 2019, 8, 575. https://doi.org/10.3390/plants8120575
Maksimov IV, Sorokan AV, Burkhanova GF, Veselova SV, Alekseev VY, Shein MY, Avalbaev AM, Dhaware PD, Mehetre GT, Singh BP, et al. Mechanisms of Plant Tolerance to RNA Viruses Induced by Plant-Growth-Promoting Microorganisms. Plants. 2019; 8(12):575. https://doi.org/10.3390/plants8120575
Chicago/Turabian StyleMaksimov, Igor V., Antonina V. Sorokan, Guzel F. Burkhanova, Svetlana V. Veselova, Valentin Yu. Alekseev, Mikhail Yu. Shein, Azamat M. Avalbaev, Prashant D. Dhaware, Gajanan T. Mehetre, Bhim Pratap Singh, and et al. 2019. "Mechanisms of Plant Tolerance to RNA Viruses Induced by Plant-Growth-Promoting Microorganisms" Plants 8, no. 12: 575. https://doi.org/10.3390/plants8120575
APA StyleMaksimov, I. V., Sorokan, A. V., Burkhanova, G. F., Veselova, S. V., Alekseev, V. Y., Shein, M. Y., Avalbaev, A. M., Dhaware, P. D., Mehetre, G. T., Singh, B. P., & Khairullin, R. M. (2019). Mechanisms of Plant Tolerance to RNA Viruses Induced by Plant-Growth-Promoting Microorganisms. Plants, 8(12), 575. https://doi.org/10.3390/plants8120575