Growth and Nutritional Responses of Bean and Soybean Genotypes to Elevated CO2 in a Controlled Environment



Abstract
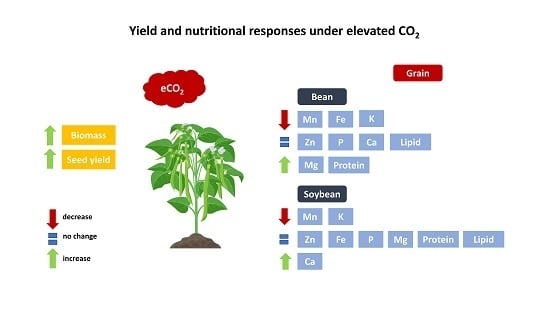
1. Introduction
2. Results
2.1. Genotypic Variation of Yield Responses to eCO2
2.2. Variation of Grain Nutritional Composition Due to eCO2
3. Discussion
4. Materials and Methods
4.1. Plant Material
4.2. Growth Conditions
4.3. Growth and Yield Measurements
4.4. Nutritional Analysis
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Preece, K.E.; Hooshyar, N.; Zuidam, N.J. Whole soybean protein extraction processes: A review. Innov. Food Sci. Emerg. 2017, 43, 163–172. [Google Scholar] [CrossRef]
- Myers, S.S.; Zanobetti, A.; Kloog, I.; Huybers, P.; Leakey, A.D.B.; Bloom, A.J.; Carlisle, E.; Dietterich, L.H.; Fitzgerald, G.; Hasegawa, T.; et al. Increasing CO2 threatens human nutrition. Nature 2014, 510, 139–142. [Google Scholar] [CrossRef] [PubMed]
- Ebert, A.W.; Chang, C.-H.; Yan, M.-R.; Yang, R.-Y. Nutritional composition of mungbean and soybean sprouts compared to their adult growth stage. Food Chem. 2017, 237, 15–22. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.; Singh, J.P.; Kaur, A.; Singh, N. Phenolic composition and antioxidant potential of grain legume seeds: A review. Food Res. Int. 2017, 101, 1–16. [Google Scholar] [CrossRef]
- Cernay, C.; Ben-Ari, T.; Pelzer, E.; Meynard, J.-M.; Makowski, D. Estimating variability in grain legume yields across Europe and the Americas. Sci. Rep. 2015, 5, 11171. [Google Scholar] [CrossRef]
- FAOSTAT. Food and Agriculture Organization of the United Nations. Available online: http://www.fao.org/faostat/en/#data/QC (accessed on 1 August 2019).
- Ma, Z.; Flynn, J.; Libra, G.; Shi, Z. Elevated CO2 Accelerates Depletion of Phosphorus by Common Bean (Phaseolus vulgaris) in association with Altered Leaf Biochemical Properties. Pedosphere 2017, 28, 422–429. [Google Scholar] [CrossRef]
- Earth System Research Laboratory, G.M.S. Available online: https://www.esrl.noaa.gov/gmd/ccgg/trends/ (accessed on 30 July 2019).
- Resco de Dios, V.; Mereed, T.E.; Ferrio, J.P.; Tissue, D.T.; Voltas, J. Intraspecific variation in juvenile tree growth under elevated CO2 alone and with O3: A meta-analysis. Tree Physiol. 2016, 36, 682–693. [Google Scholar] [CrossRef]
- Kimball, B.; Kobayashi, K.; Bindi, M. Responses of agricultural crops to free-air CO2 enrichment. In ADV AGRON; Elsevier: Amsterdam, The Netherlands, 2002; Volume 77, pp. 293–368. [Google Scholar]
- Moya, T.B.; Ziska, L.H.; Namuco, O.S.; Olsyk, D. Growth dynamics and genotypic variation in tropical, field-grown paddy rice (Oryza sativa L.) in response to increasing carbon dioxide and temperature. Glob. Chang. Biol. 1998, 4, 645–656. [Google Scholar] [CrossRef]
- Baker, J.T. Yield responses of southern US rice cultivars to CO2 and temperature. Agric. For. Meteorol. 2004, 122, 129–137. [Google Scholar] [CrossRef]
- Ziska, L.H.; Manalo, P.A.; Ordonez, R.A. Intraspecific variation in the response of rice (Oryza sativa L.) to increased CO2 and temperature: Growth and yield response of 17 cultivars1. J. Exp. Bot. 1996, 47, 1353–1359. [Google Scholar] [CrossRef]
- Ahmed, F.E.; Hall, A.E.; Madore, M.A. Interactive effects of high temperature and elevated carbon dioxide concentration on cowpea [Vigna unguiculata (L.) Walp.]. Plant Cell Environ. 1993, 16, 835–842. [Google Scholar] [CrossRef]
- Ziska, L.H.; Morris, C.F.; Goins, E.W. Quantitative and qualitative evaluation of selected wheat varieties released since 1903 to increasing atmospheric carbon dioxide: Can yield sensitivity to carbon dioxide be a factor in wheat performance? Glob. Chang. Biol. 2004, 10, 1810–1819. [Google Scholar] [CrossRef]
- Bunce, J.A. Contrasting responses of seed yield to elevated carbon dioxide under field conditions within Phaseolus vulgaris. Agric. Ecosyst. Environ. 2008, 128, 219–224. [Google Scholar] [CrossRef]
- Ziska, L.H.; Bunce, J.A. Sensitivity of field-grown soybean to future atmospheric CO2: Selection for improved productivity in the 21st century. Funct. Plant Biol. 2000, 27, 979–984. [Google Scholar] [CrossRef]
- Ziska, L.H.; Bunce, J.A.; Caulfield, F.A. Rising Atmospheric Carbon Dioxide and Seed Yield of Soybean Genotypes. Crop Sci. 2001, 41, 385–391. [Google Scholar] [CrossRef]
- Kumagai, E.; Aoki, N.; Masuya, Y.; Shimono, H. Phenotypic Plasticity Conditions the Response of Soybean Seed Yield to Elevated Atmospheric CO2 Concentration. Plant Physiol. 2015, 169, 2021–2029. [Google Scholar] [CrossRef]
- Li, Y.; Yu, Z.; Jin, J.; Zhang, Q.; Wang, G.; Liu, C.; Wu, J.; Wang, C.; Liu, X. Impact of Elevated CO2 on Seed Quality of Soybean at the Fresh Edible and Mature Stages. Front. Plant Sci. 2018, 9. [Google Scholar] [CrossRef]
- Loladze, I. Hidden shift of the ionome of plants exposed to elevated CO₂ depletes minerals at the base of human nutrition. eLife 2014, 3, e02245. [Google Scholar] [CrossRef]
- Högy, P.; Fangmeier, A. Effects of elevated atmospheric CO2 on grain quality of wheat. J. Cereal Sci. 2008, 48, 580–591. [Google Scholar] [CrossRef]
- Singh, S.K.; Barnaby, J.Y.; Reddy, V.R.; Sicher, R.C. Varying Response of the Concentration and Yield of Soybean Seed Mineral Elements, Carbohydrates, Organic Acids, Amino Acids, Protein, and Oil to Phosphorus Starvation and CO2 Enrichment. Front. Plant Sci. 2016, 7, 1967. [Google Scholar] [CrossRef]
- Dong, J.-L.; Li, X.; Nazim, G.; Duan, Z.-Q. Interactive effects of elevated carbon dioxide and nitrogen availability on fruit quality of cucumber (Cucumis sativus L.). J. Integr. Agric. 2018, 17, 2438–2446. [Google Scholar] [CrossRef]
- Dong, J.; Gruda, N.; Lam, S.K.; Li, X.; Duan, Z. Effects of Elevated CO2 on Nutritional Quality of Vegetables: A Review. Front. Plant Sci. 2018, 9, 924. [Google Scholar] [CrossRef] [PubMed]
- Ziska, L.H.; Palowsky, R.; Reed, D.R. A quantitative and qualitative assessment of mung bean (Vigna mungo (L.) Wilczek) seed in response to elevated atmospheric carbon dioxide: Potential changes in fatty acid composition. J. Sci. Food Agric. 2007, 87, 920–923. [Google Scholar] [CrossRef]
- Thomas, J.; Prasad, P.; Boote, K.; Allen, L., Jr. Seed composition, seedling emergence and early seedling vigour of red kidney bean seed produced at elevated temperature and carbon dioxide. J. Agron. Crop Sci. 2009, 195, 148–156. [Google Scholar] [CrossRef]
- Palacios, C.J.; Grandis, A.; Carvalho, V.J.; Salatino, A.; Buckeridge, M.S. Isolated and combined effects of elevated CO2 and high temperature on the whole-plant biomass and the chemical composition of soybean seeds. Food Chem. 2019, 275, 610–617. [Google Scholar] [CrossRef]
- Bunce, J.A. Elevated Carbon Dioxide Effects on Reproductive Phenology and Seed Yield among Soybean Cultivars. Crop Sci. 2015, 55, 339–343. [Google Scholar] [CrossRef]
- Bishop, K.A.; Betzelberger, A.M.; Long, S.P.; Ainsworth, E.A. Is there potential to adapt soybean (Glycine max Merr.) to future [CO2]? An analysis of the yield response of 18 genotypes in free-air CO2) enrichment. Plant Cell Environ. 2015, 38, 1765–1774. [Google Scholar] [CrossRef]
- Prasad, P.V.; Boote, K.J.; Allen, L.H., Jr.; Thomas, J.M. Effects of elevated temperature and carbon dioxide on seed-set and yield of kidney bean (Phaseolus vulgaris L.). Glob. Chang. Biol. 2002, 8, 710–721. [Google Scholar] [CrossRef]
- Ainsworth, E.A.; Davey, P.A.; Bernacchi, C.J.; Dermody, O.C.; Heaton, E.A.; Moore, D.J.; Morgan, P.B.; Naidu, S.L.; Ra, H.S.Y.; Zhu, X.G. A meta-analysis of elevated [CO2] effects on soybean (Glycine max) physiology, growth and yield. Glob. Chang. Biol. 2002, 8, 695–709. [Google Scholar] [CrossRef]
- Palta, J.; Ludwig, C. Elevated CO2 during pod filling increased seed yield but not harvest index in indeterminate narrow-leafed lupin. Aust. J. Agric. Res. 2000, 51, 279–286. [Google Scholar] [CrossRef]
- Wu, D.-X.; Wang, G.-X.; Bai, Y.-F.; Liao, J.-X. Effects of elevated CO2 concentration on growth, water use, yield and grain quality of wheat under two soil water levels. Agric. Ecosyst. Environ. 2004, 104, 493–507. [Google Scholar] [CrossRef]
- Pang, J.; Zhu, J.-G.; Xie, Z.-B.; Liu, G.; Zhang, Y.-L.; Chen, G.-P.; Zeng, Q.; Cheng, L. A new explanation of the N concentration decrease in tissues of rice (Oryza sativa L.) exposed to elevated atmospheric CO2. Environ. Exp. Bot. 2006, 57, 98–105. [Google Scholar] [CrossRef]
- Parvin, S.; Uddin, S.; Tausz-Posch, S.; Armstrong, R.; Fitzgerald, G.; Tausz, M. Grain mineral quality of dryland legumes as affected by elevated CO2 and drought: A FACE study on lentil (Lens culinaris) and faba bean (Vicia faba). Crop Pasture Sci. 2019, 70, 244–253. [Google Scholar] [CrossRef]
- Bloom, A.J. Photorespiration and nitrate assimilation: A major intersection between plant carbon and nitrogen. Photosynth. Res. 2015, 123, 117–128. [Google Scholar] [CrossRef]
- Pleijel, H.; Gelang, J.; Sild, E.; Danielsson, H.; Younis, S.; Karlsson, P.-E.; Wallin, G.; Skärby, L.; Selldén, G. Effects of elevated carbon dioxide, ozone and water availability on spring wheat growth and yield. Physiol. Plant. 2000, 108, 61–70. [Google Scholar] [CrossRef]
- Gifford, R.M.; Barrett, D.J.; Lutze, J.L. The effects of elevated [CO2] on the C:N and C:P mass ratios of plant tissues. Plant Soil 2000, 224, 1–14. [Google Scholar] [CrossRef]
- Qiao, Y.; Miao, S.; Li, Q.; Jin, J.; Luo, X.; Tang, C. Elevated CO2 and temperature increase grain oil concentration but their impacts on grain yield differ between soybean and maize grown in a temperate region. Sci. Total Environ. 2019, 666, 405–413. [Google Scholar] [CrossRef]
- Jablonski, L.M.; Wang, X.; Curtis, P.S. Plant reproduction under elevated CO2 conditions: A meta-analysis of reports on 79 crop and wild species. New Phytol. 2002, 156, 9–26. [Google Scholar] [CrossRef]
- Taub, D.R.; Wang, X. Why are Nitrogen Concentrations in Plant Tissues Lower under Elevated CO2? A Critical Examination of the Hypotheses. J. Integr. Plant Biol. 2008, 50, 1365–1374. [Google Scholar] [CrossRef]
- Ekman, A.; Bulow, L.; Stymne, S. Elevated atmospheric CO2 concentration and diurnal cycle induce changes in lipid composition in Arabidopsis thaliana. New Phytol. 2007, 174, 591–599. [Google Scholar] [CrossRef]
- Fernando, N.; Panozzo, J.; Tausz, M.; Norton, R.M.; Fitzgerald, G.J.; Myers, S.; Nicolas, M.E.; Seneweera, S. Intra-specific variation of wheat grain quality in response to elevated [CO2] at two sowing times under rain-fed and irrigation treatments. J. Cereal Sci. 2014, 59, 137–144. [Google Scholar] [CrossRef]
- Ziska, L.H.; Bunce, J.A. Predicting the impact of changing CO2 on crop yields: Some thoughts on food. New Phytol. 2007, 175, 607–618. [Google Scholar] [CrossRef] [PubMed]
- Ainsworth, E.A.; Long, S.P. What have we learned from 15 years of free-air CO2 enrichment (FACE)? A meta-analytic review of the responses of photosynthesis, canopy properties and plant production to rising CO2. New Phytol. 2005, 165, 351–372. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.S.; Roriz, M.; Carvalho, S.M.P.; Vasconcelos, M.W. Iron partitioning at an early growth stage impacts iron deficiency responses in soybean plants (Glycine max L.). Front. Plant Sci. 2015, 6. [Google Scholar] [CrossRef] [PubMed]
- AOAC. Official Methods of Analysis of AOAC International; AOAC: Rockville, MD, USA, 2006. [Google Scholar]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Parameter | Mean CO2 Effect | CO2 | Variety | C x V | Correlation | |
|---|---|---|---|---|---|---|
| Aboveground dry weight, g plt−1 | 5.8% | * | **** | ns | 0.747 | * |
| Height, cm plt−1 | 4.8% | * | **** | ns | 0.593 | ** |
| Seed yield, g plant−1 | 5.0% | * | **** | * | ‒ | ‒ |
| Harvest index, g g−1 | −0.2% | ns | ** | ns | 0.096 | ns |
| No. of pods, plt−1 | 2.9% | ns | **** | * | 0.736 | * |
| No. of seeds, plt−1 | 3.8% | ns | **** | *** | 0.838 | ** |
| No. of seeds, pod−1 | 7.5% | ** | **** | ns | 0.314 | ns |
| 100-seed weight, g | −13.1% | **** | **** | * | −0.108 | ns |
| Parameter | Mean CO2 Effect | CO2 | Variety | C x V | Correlation | |
|---|---|---|---|---|---|---|
| Aboveground dry weight, g plt−1 | 6.9% | * | **** | ns | 0.625 | ** |
| Height, cm plt−1 | 3.6% | * | **** | **** | 0.119 | ns |
| Seed yield, g plt−1 | 7.1% | * | **** | ** | ‒ | |
| Harvest index, g g−1 | −1.0% | ns | **** | ns | 0.396 | ns |
| No. of pods, plt−1 | 7.2% | ** | **** | ns | 0.784 | ** |
| No. of seeds, plt−1 | 5.5% | * | **** | **** | 0.600 | * |
| No. of seeds, pod−1 | 5.9% | * | * | * | 0.665 | ** |
| 100-seed weight, g | −12.3% | **** | **** | ** | −0.280 | ns |
| Seed Element | CO2 | Variety | C x V |
|---|---|---|---|
| Zn | ns | **** | ns |
| Mn | **** | **** | *** |
| Fe | **** | **** | **** |
| P | ns | **** | ** |
| Mg | **** | **** | ns |
| Ca | ns | ** | **** |
| K | **** | **** | ** |
| Protein | **** | **** | **** |
| Lipid | ns | ** | ns |
| Seed Element | CO2 | Variety | C x V |
|---|---|---|---|
| Zn | ns | **** | ** |
| Mn | **** | **** | ** |
| Fe | ns | **** | **** |
| P | ns | **** | ns |
| Mg | ns | **** | ** |
| Ca | ** | *** | ** |
| K | *** | **** | ** |
| Protein | ns | **** | ** |
| Lipid | ns | *** | ns |
| Crop | Acession Number | Growth Habit | Common Name | Origin | Performance at eCO2 |
|---|---|---|---|---|---|
| Bean a | PI 203929 | D | G1274 | Mexico | Strong-responsive |
| Bean a | PI 458586 | D or I | NHB | Netherlands | Strong-responsive |
| Bean b | PI 169920 | D | Kazak | Turkey | Weak-responsive |
| Bean a | PI 324691 | D | ZK | Hungary | Weak-responsive |
| Bean a | W6 9628 | I | Dama | Czechoslovakia | Weak-responsive |
| Bean a | W6 12428 | NS | PP 63 | Bulgaria | Strong-responsive |
| Bean a | PI 550128 | I | Trend | Netherlands | Weak-responsive |
| Bean a | PI 550038 | NS | Garnet | United States | Weak-responsive |
| Bean b | PI 212027 | D | G1378 | Iran | Weak-responsive |
| Bean a | PI 598287 | I | PV1-4 | Japan | Weak-responsive |
| Bean a | PI 368715 | D or I | Rosomanska | Macedonia | Strong-responsive |
| Bean a | PI 550035 | D | Agate | United States | Weak-responsive |
| Bean b | PI 149484 | D | Logan | United States | Weak-responsive |
| Bean a | PI 136687 | D | Yamal | Canada | Weak-responsive |
| Bean a | PI 165933 | D | Shimi | India | Weak-responsive |
| Bean a | PI 550037 | D | Dandy | United States | Strong-responsive |
| Bean b | G 8853 | D | Medra | Germany | Strong-responsive |
| Beana | PI 477023 | D or I | CBB | Netherlands | Strong-responsive |
| Soybean a | PI 361085 A | I | L.117 | Romania | Strong-responsive |
| Soybean a | PI 437413 | I | Ussurijscaja | Russia | Weak-responsive |
| Soybean a | PI 424194 | D | ISZ-II | Hungary | Weak-responsive |
| Soybean a | PI 445823 | I | Tubinger | Germany | Weak-responsive |
| Soybean a | PI 378676 A | I | Primorskaja | Russia | Strong-responsive |
| Soybean a | PI 561302 A | I | BMS | China | Weak-responsive |
| Soybean a | PI 437101 | I | DV-0197 | Russia | Weak-responsive |
| Soybean a | PI 319537 A | I | Tono | China | Strong-responsive |
| Soybean a | PI 437224 | I | CSchi 675 | Moldova | Strong-responsive |
| Soybean a | PI 319534 A | I | Honshu | China | Strong-responsive |
| Soybean a | PI 437676 A | I | MTTPDH | China | Weak-responsive |
| Soybean a | PI 445829 A | I | Dunayka | Romania | Strong-responsive |
| Soybean a | PI 361097 A | I | Novosadska | Serbia | Strong-responsive |
| Soybean a | PI 360952 | I | Amurskaja | Russia | Weak-responsive |
| Soybean a | PI 417554 | I | EM | Poland | Strong-responsive |
| Soybean a | PI 538409 | D | Shironomai | Japan | Strong-responsive |
| Soybean a | PI 153271 | I | WB | Belgium | Strong-responsive |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Soares, J.; Deuchande, T.; Valente, L.M.P.; Pintado, M.; Vasconcelos, M.W. Growth and Nutritional Responses of Bean and Soybean Genotypes to Elevated CO2 in a Controlled Environment. Plants 2019, 8, 465. https://doi.org/10.3390/plants8110465
Soares J, Deuchande T, Valente LMP, Pintado M, Vasconcelos MW. Growth and Nutritional Responses of Bean and Soybean Genotypes to Elevated CO2 in a Controlled Environment. Plants. 2019; 8(11):465. https://doi.org/10.3390/plants8110465
Chicago/Turabian StyleSoares, José, Teresa Deuchande, Luísa M.P. Valente, Manuela Pintado, and Marta W. Vasconcelos. 2019. "Growth and Nutritional Responses of Bean and Soybean Genotypes to Elevated CO2 in a Controlled Environment" Plants 8, no. 11: 465. https://doi.org/10.3390/plants8110465
APA StyleSoares, J., Deuchande, T., Valente, L. M. P., Pintado, M., & Vasconcelos, M. W. (2019). Growth and Nutritional Responses of Bean and Soybean Genotypes to Elevated CO2 in a Controlled Environment. Plants, 8(11), 465. https://doi.org/10.3390/plants8110465