Micronutrient Status and Selected Physiological Parameters of Roots in Nickel-Exposed Sinapis alba L. Affected by Different Sulphur Levels



Abstract
1. Introduction
2. Results
2.1. Micronutrient Concentrations
- -
- no changes in the root Fe concentration under intensive S nutrition in plants under the lowest and the highest Ni exposure and the Fe increase under medium Ni concentration,
- -
- no changes in the root B concentration under both elevated S levels,
- -
- an increase in the shoot Mn at both the high S levels and a decrease in root Mn at the 9 mM S/0.0004 mM Ni treatment,
- -
- an increase or no changes in the root Zn concentration at 6 and 9 mM S, respectively.
2.2. Micronutrient Translocation Factor (TF)
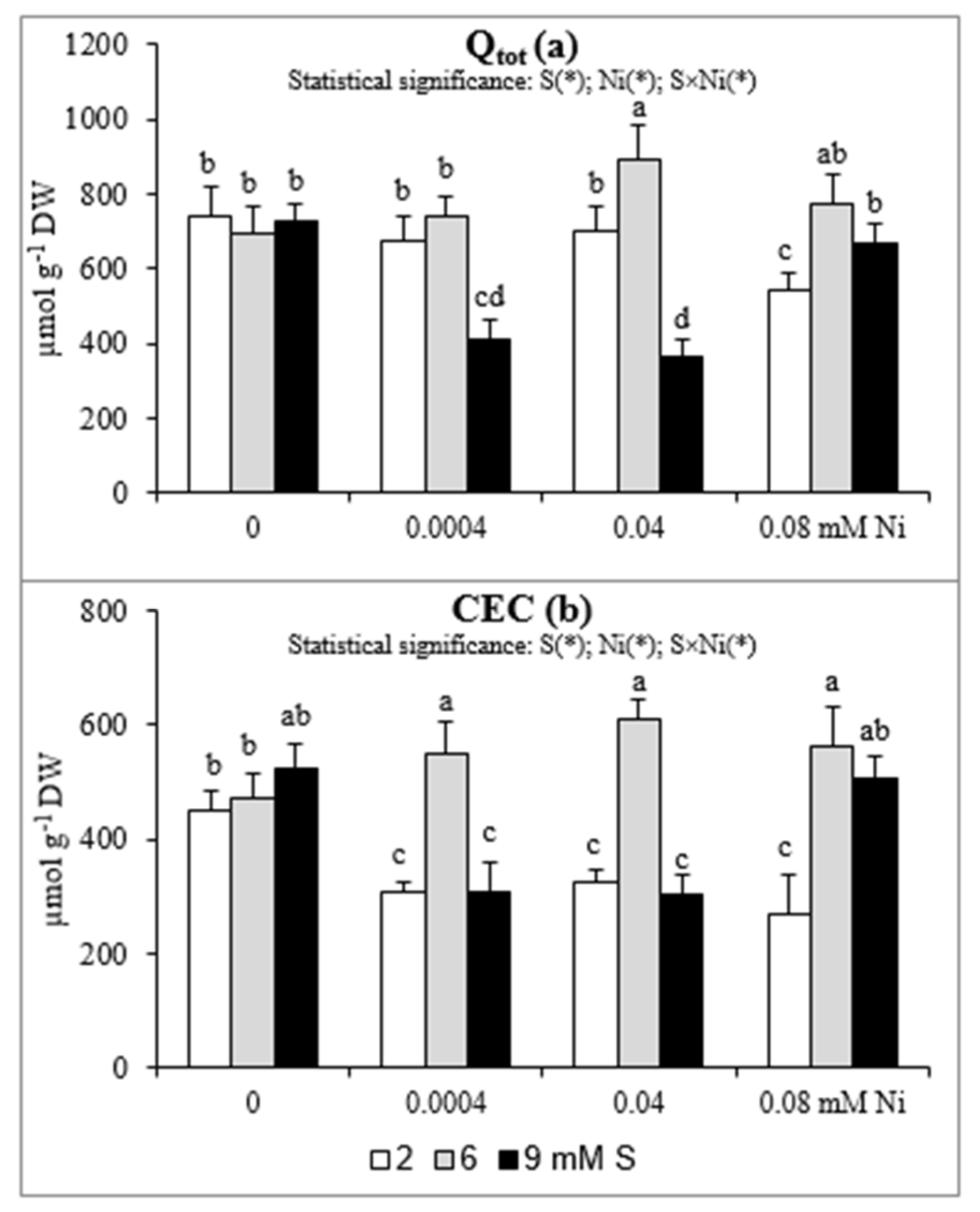
2.3. Total Surface Charge (Qtot) and Cation Exchange Capacity (CEC)
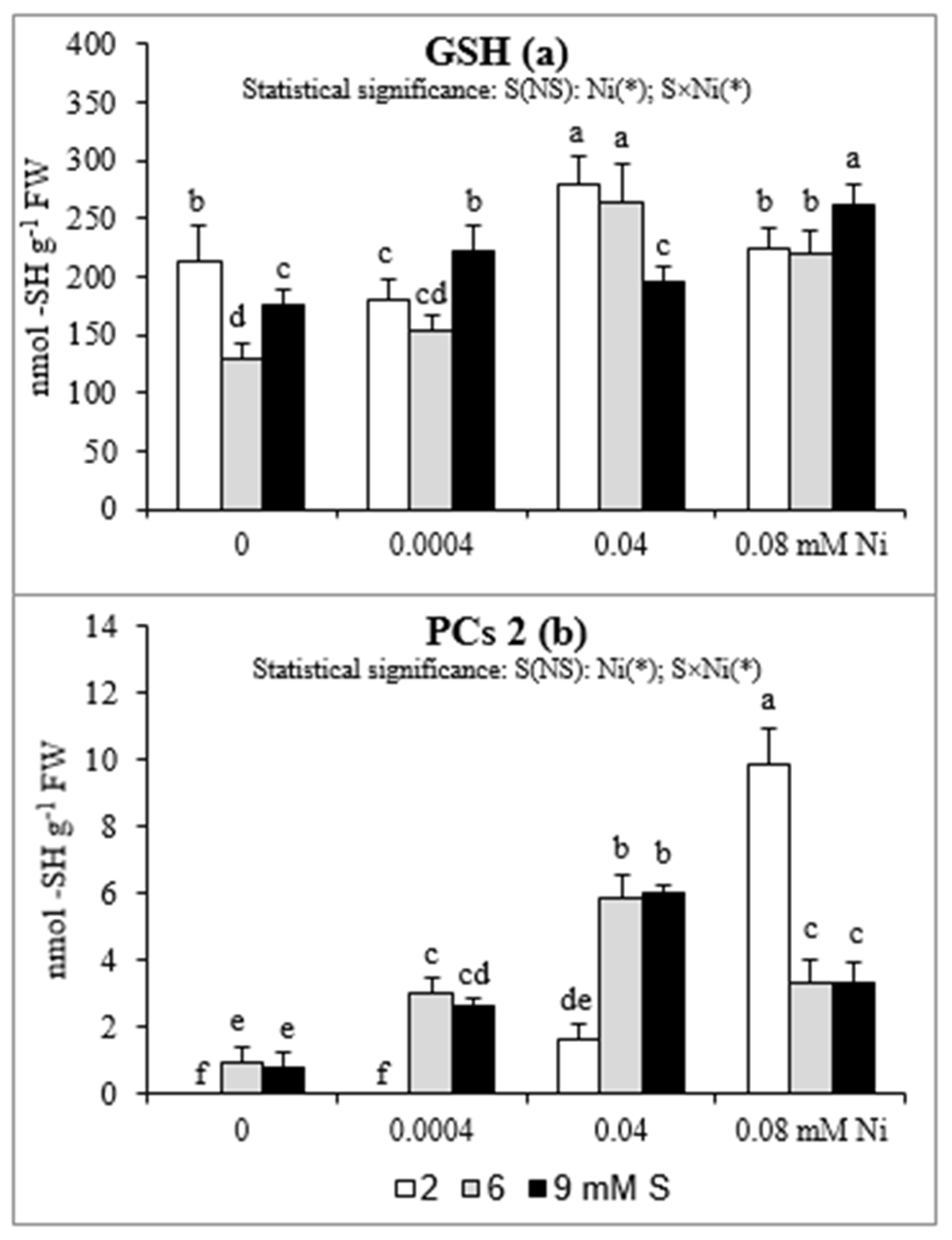
2.4. GSH and PCs Accumulation in Roots
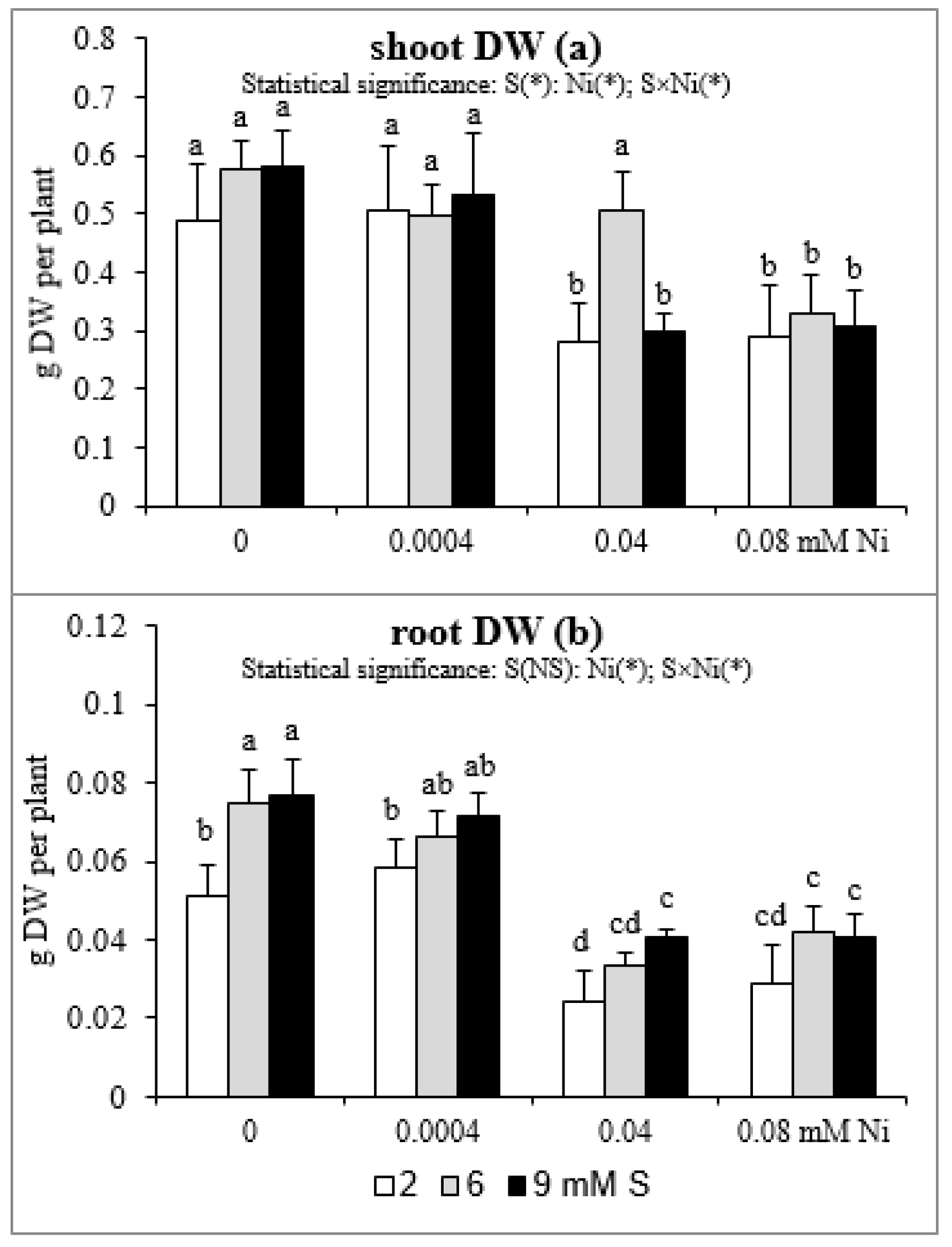
2.5. Root and Shoot Biomass
3. Discussion
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Determination of Biomass and Micronutrient Concentrations
4.3. Determination of CEC and Qtot by Potentiometric Titration
4.4. Determination of γ-Glu-Cys Peptides by HPLC Method
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ragsdale, S.W. Nickel-based enzyme systems. J. Biol. Chem. 2009, 284, 18571–18575. [Google Scholar] [CrossRef] [PubMed]
- Poonkothai, M.; Shyamala Vijayavathi, B. Nickel as an essential element and a toxicant. Int. J. Environ. Sci. 2012, 1, 285–288. [Google Scholar]
- Fabiano, C.C.; Tezotto, T.; Favarin, J.L.; Polacco, J.C.; Mazzafera, P. Essentiality of nickel in plants: A role in plant stresses. Front. Plant Sci. 2015, 6, 754. [Google Scholar] [CrossRef] [PubMed]
- Bhatia, N.P.; Nkang, A.E.; Walsh, K.B.; Baker, A.J.; Ashwath, N.; Midmore, D.J. Successful seed germination of the nickel hyperaccumulator Stackhousia tryonii. Ann. Bot. 2005, 96, 159–163. [Google Scholar] [CrossRef][Green Version]
- Polacco, J.C.; Mazzafera, P.; Tezotto, T. Opinion: Nickel and urease in plants: Still many knowledge gaps. Plant Sci. 2013, 199, 79–90. [Google Scholar] [CrossRef]
- Ahmad, M.S.A.; Ashraf, M. Essential roles and hazardous effects of nickel in plants. Rev. Environ. Contam. Toxicol. 2011, 214, 125–167. [Google Scholar] [CrossRef]
- López, M.Á.; Magnitski, S. Nickel: The last of the essential micronutrients. Agron. Colomb. 2011, 29, 49–56. [Google Scholar]
- Bhalerao, S.A.; Sharma, A.S.; Poojari, A.C. Toxicity of nickel in plants. Int. J. Pure Appl. Biosci. 2015, 3, 345–355. [Google Scholar]
- Pavlovkin, J.; Fiala, R.; Čiamporová, M.; Martinka, M.; Repka, V. Impact of nickel on grapevine (Vitis vinifera L.) root plasma membrane, ROS generation, and cell viability. Acta Bot. Croat. 2016, 75, 25–30. [Google Scholar] [CrossRef]
- Nieminen, T.M.; Ukonmaanaho, L.; Rausch, N.; Shotyk, W. Biogeochemistry of nickel and its release into the environment. In Nickel and Its Surprising Impact in Nature, 1st ed.; Sigel, A., Sigel, H., Sigel, R.K.O., Eds.; John Wiley & Sons: Chichester, UK, 2007; pp. 1–30. [Google Scholar] [CrossRef]
- Sreekanth, T.V.M.; Nagajyothi, P.C.; Lee, K.D.; Prasad, T.N.V.K.V. Occurrence, physiological responses and toxicity of nickel in plants. Int. J. Environ. Sci. Technol. 2013, 10, 1129–1140. [Google Scholar] [CrossRef]
- Harasim, P.; Filipek, T. Nickel in the environment. J. Elem. 2015, 20, 525–534. [Google Scholar] [CrossRef]
- Chen, C.; Huang, D.; Liu, J. Functions and toxicity of nickel in plants: Recent advances and future prospects. Clean Soil Air Water 2009, 37, 304–313. [Google Scholar] [CrossRef]
- Shafeeq, A.; Butt, Z.A.; Muhammad, S. Response of nickel pollution on physiological and biochemical attributes of wheat (Triticum aestivum L.) var. Bhakar-02. Pak. J. Bot. 2012, 44, 111–116. [Google Scholar]
- Hussain, M.B.; Ali, S.; Azam, A.; Hina, S.; Farooq, M.A.; Ali, B.; Bharwana, S.A.; Gill, M.B. Morphological, physiological and biochemical responses of plants to nickel stress: A review. Afr. J. Agric. Res. 2013, 8, 1596–1602. [Google Scholar] [CrossRef]
- Ghasemi, R.; Ghaderian, S.M.; Krämer, U. Interference of nickel with copper and iron homeostasis contributes to metal toxicity symptoms in the nickel hyperaccumulator plant Alyssum inflatum. New Phytol. 2009, 184, 566–580. [Google Scholar] [CrossRef] [PubMed]
- Yadav, S.S.; Shukla, R.; Sharma, Y.K. Nickel toxicity on seed germination and growth in radish (Raphanus sativus) and its recovery using copper and boron. J. Environ. Biol. 2009, 30, 461–466. [Google Scholar]
- Sengar, R.S.; Gupta, S.; Gautam, M.; Sharma, A.; Sengar, K. Occurrence, uptake, accumulation and physiological responses of nickel in plants and its effects on environment. Res. J. Phytochem. 2008, 2, 44–60. [Google Scholar] [CrossRef]
- Yusuf, M.; Fariduddin, Q.; Hayat, S.; Ahmad, A. Nickel: An overview of uptake, essentiality and toxicity in plants. Bull. Environ. Contam. Toxicol. 2011, 86, 1–17. [Google Scholar] [CrossRef]
- Stanisławska-Głubiak, E.; Korzeniowska, J. Tolerance of white mustard (Sinapis alba L.) to soil pollution with several heavy metals. Ecol. Chem. Eng. Ser. A 2011, 18, 445–450. [Google Scholar]
- Putnik-Delić, M.I.; Maksimović, I.V.; Gani-Novaković, I.; Zeremski, T.; Marjanović-Jaromela, A. The effect of Ni concentration of the most abundant essential cations in several Brassica species. J. Nat. Sci. 2014, 126, 15–23. [Google Scholar] [CrossRef]
- Manohar, P.; Ram; Pushpan, R.; Rohini, S. Mustard and its uses in Ayurveda. Indian J. Tradit. Knowl. 2009, 8, 400–404. [Google Scholar]
- Damian, C. Physical properties of mustard seeds (Sinapis alba L.). Lucr. Ştiinţifice Ser. Zooteh. 2014, 61, 39–44. [Google Scholar]
- Paunović, D.; Šolević Knudsen, T.; Krivokapić, M.; Zlatković, B.; Antić, M. Sinalbin degradation products in mild yellow mustard paste. Hem. Ind. 2012, 66, 29–32. [Google Scholar] [CrossRef]
- Young-Mathews, A. Plant guide for field mustard (Brassica rapa ssp. rapa). USDA-Natural Resources Conservation Service, Plant Materials Center, Corvallis. Available online: https://plants.usda.gov/plantguide/pdf/pg_brrar.pdf (accessed on 20 September 2019).
- Ryant, P. Effect of sulphur fertilisation on yield and quality of white mustard seeds. Acta Univ. Agric. Silvic. Mendel. Brun. 2009, 57, 95–104. [Google Scholar] [CrossRef]
- Bloem, E.; Haneklaus, S.; Daniels, R.; Schnug, E. Influence of sulfur fertilization on floral scent patterns of crops in full bloom. Landbauforsch Volk. Appl. Agric. For. Res. 2010, 60, 45–50. [Google Scholar]
- Filipek-Mazur, B.; Gondek, K. Yielding and sulphur content in white mustard as the effect of application of multi-component fertilizers containing sulphur. Acta Agrophys. 2005, 6, 343–351. [Google Scholar]
- Barczak, B.; Kozera, W.; Knapowski, T.; Ralcewicz, M. Selected field components in white mustard (Sinapis alba) versus sulfur fertilization. J. Cent. Eur. Agric. 2011, 12, 380–389. [Google Scholar] [CrossRef]
- Shekhawat, K.; Kandpal, B.K.; Chand, G.; Singh, D. Sulphur management for increased productivity of Indian mustard: A review. Ann. Plant. Soil Res. 2015, 17, 1–12. [Google Scholar]
- Ray, K.; Sengupta, K.; Pal, A.K.; Banerjee, H. Effects of sulphur fertilization on yield, S uptake and quality of Indian mustard under varied irrigation regimes. Plant Soil Environ. 2015, 61, 6–10. [Google Scholar] [CrossRef]
- Hawkesford, M.J. Plant responses to sulphur deficiency and the genetic manipulation of sulphate transporters to improve S-utilization efficiency. J. Exp. Bot. 2000, 51, 131–138. [Google Scholar] [CrossRef]
- Mašauskiene, A.; Mašauskas, V. Soil sulphur problems and management. In Sustainable Agriculture. Series Ecosystem Health and Sustainable Agriculture; Jakobsson, C., Ed.; Baltic University Press: Uppsala, Sweden, 2012; pp. 113–116. [Google Scholar]
- Eriksen, J. Sulphur Cycling in Agrosystems. Ph.D. Thesis, Aarhus University, Tjele, Denmark, 2010. [Google Scholar]
- Procházková, D.; Pavlíková, D.; Pavlík, M. Sulphur: Role in alleviation of environmental stress in crop plants. In Plant-Environment Interaction: Responses and Approaches to Mitigate Stress; Azooz, M.M., Ahmad, P., Eds.; John Wiley & Sons: Hoboken, NJ, USA, 2016; pp. 84–96. [Google Scholar] [CrossRef]
- Prasad, M.N.V. Heavy Metal. Stress in Plants: From Biomolecules to Ecosystems, 2nd ed.; Springer: Berlin/Heidelberg, Germany, 2004; p. 462. [Google Scholar] [CrossRef]
- Maheshwari, R.; Dubey, R.S. Nickel-induced oxidative stress and the role of antioxidant defense in rice seedlings. Plant Growth Regul. 2009, 59, 37–49. [Google Scholar] [CrossRef]
- Yadav, N.; Sharma, S. An account of nickel requirement, toxicity and oxidative stress in plants. Biol. Forum Int. J. 2016, 8, 414–419. [Google Scholar]
- Barker, A.V.; Pilbeam, D.J. Handbook of Plant Nutrition, 2nd ed.; CRC Press: Boca Raton, FL, USA, 2015; p. 773. [Google Scholar]
- Rahman, H.; Sabreen, S.; Alam, S.; Kawai, S. Effects of nickel on growth and composition of metal micronutrients in barley plants grown in nutrient solution. J. Plant Nutr. 2005, 28, 393–404. [Google Scholar] [CrossRef]
- Yadav, S. Toxicity of Nickel in Plants and Its Interaction with Certain Micronutrient. Ph.D. Thesis, University of Lucknow, Lucknow, India, 2013. [Google Scholar]
- Fageira, N.K. Mineral. Nutrition of Rice, 1st ed.; CRC Press: Boca Raton, FL, USA, 2013; p. 586. [Google Scholar] [CrossRef]
- Aibara, I.; Miwa, K. Strategies for optimization of mineral nutrient transport in plants: Multilevel regulation of nutrient-dependent dynamics of root architecture and transporter activity. Plant Cell Physiol. 2014, 55, 2027–2036. [Google Scholar] [CrossRef]
- Mitra, G.N. Regulation of Nutrient Uptake by Plants, 1st ed.; Springer: New Delhi, India, 2015; p. 195. [Google Scholar] [CrossRef]
- Moosavi, A.A.; Mansouri, S.; Zahedifar, M. Effect of soil water stress and nickel application on micronutrient status of canola grown on two calcareous soils. Plant. Prod. Sci. 2015, 18, 377–387. [Google Scholar] [CrossRef]
- Ernst, W.H.O.; Krauss, G.J.; Verkleij, J.A.C.; Wesenberg, D. Interaction of heavy metals with the sulphur metabolism in angiosperms from an ecological point of view. Plant Cell Environ. 2008, 31, 123–143. [Google Scholar] [CrossRef]
- Ranade-Malvi, U. Interaction of micronutrients with major nutrients with special reference to potassium. Karnataka J. Agric. Sci. 2011, 24, 106–109. [Google Scholar]
- Jankowski, K.J.; Budzyński, W.S.; Kijewski, Ł.; Klasa, A. Concentrations of cooper, zinc and manganese in the roots, straw and oil cake of white mustard (Sinapis alba L.) and Indian mustard (Brassica juncea (L.) Czern. et Coss) depending on sulphur fertilization. Plant Soil Environ. 2014, 60, 364–371. [Google Scholar] [CrossRef]
- Matraszek, R.; Hawrylak-Nowak, B.; Chwil, M.; Chwil, S.; Rudaś, M. Effect of the interaction of nickel stress and sulphur supplementation on the content and accumulation of macronutrients in white mustard (Sinapis alba L.). Electron. J. Pol. Agric. Univ. 2017, 20, 1. [Google Scholar] [CrossRef]
- Szatanik-Kloc, A.; Szerement, J.; Józefaciuk, G. The role of cell walls and pectins in cation exchange and surface area of plant roots. J. Plant Physiol. 2017, 215, 85–90. [Google Scholar] [CrossRef]
- Freeman, J.L.; Persans, M.W.; Nieman, K.; Albrecht, C.; Peer, W.; Pickering, I.J.; Salt, D.E. Increased glutathione biosynthesis plays a role in nickel tolerance in Thlaspi nickel hyperaccumulators. Plant Cell 2004, 16, 2176–2191. [Google Scholar] [CrossRef] [PubMed]
- Hossain, M.A.; Piyatida, P.; Texteira da Silva, J.A.; Fujita, M. Molecular mechanism of heavy metal toxicity and tolerance in plants: Central role of glutathione in detoxification of Reactive Oxygen Species and methylglyoxal and in heavy metal chelation. J. Bot. 2012, 2012, 872875. [Google Scholar] [CrossRef]
- Viehweger, K. How plants cope with heavy metals? Bot. Stud. 2014, 55, 35. [Google Scholar] [CrossRef] [PubMed]
- Hasanuzzaman, M.; Alam, M.M.; Nahar, K.; Mohsin, S.M.; Bhuyan, M.H.M.B.; Parvin, K.; Hawrylak-Nowak, B.; Fujita, M. Silicon-induced antioxidant defense and methylglyoxal detoxification works coordinately in alleviating nickel toxicity in Oryza sativa L. Ecotoxicology 2019, 28, 261. [Google Scholar] [CrossRef] [PubMed]
- Jones, J.B., Jr. Laboratory Guide for Conducting Soil Tests and Plant Analysis, 1st ed.; CRC Press: Boca Raton, FL, USA, 2001; p. 363. [Google Scholar]
- Aref, F. Concentration and uptake of zinc and boron in corn leaf as affected by zinc sulfate and boric acid fertilizers in a deficient soil. Life Sci. J. 2011, 8, 26–31. [Google Scholar]
- Wolf, B. Improvements in the azomethine-H method for determination of boron. Commun. Soil Sci. Plant Anal. 1974, 5, 39–44. [Google Scholar] [CrossRef]
- Nowosielski, O. Methods for the Determination of Fertilisation Requirements, 1st ed.; PWRiL: Warszawa, Poland, 1974. (In Polish) [Google Scholar]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Concentration in the Nutrient Solution (mM) | Concentration of the Micronutrients in the Roots (mg kg−1 DM) | |||||||
|---|---|---|---|---|---|---|---|---|
| S | Ni | Fe | B | Cl | Cu | Mn | Mo | Zn |
| 2 | 1378 ± 42.6a | 17.24 ± 1.58c | 8485 ± 53.87 | 5.48 ± 0.22ab | 47.18 ± 1.02e | 0.642 ± 0.093 | 34.89 ± 0.95d–f | |
| 6 | 0.00 | 1382 ± 43.0a | 22.13 ± 2.73a | 8478 ± 70.42 | 5.72 ± 0.35a | 53.37 ± 0.97bc | 0.585 ± 0.060 | 45.57 ± 1.14a |
| 9 | 1377 ± 52.2a | 20.77 ± 1.65b | 8498 ± 55.39 | 5.13 ± 0.56a–c | 55.74 ± 0.76a | 0.663 ± 0.074 | 42.61 ± 0.99ab | |
| 2 | 1372 ± 32.5a | 18.14 ± 2.51c | 8492 ± 97.01 | 5.01 ± 0.46a–d | 54.81 ± 0.84a–c | 0.608 ± 0.087 | 36.28 ± 1.22de | |
| 6 | 0.0004 | 1380 ± 61.7a | 17.29 ± 1.67c | 8484 ± 41.66 | 5.36 ± 0.70ab | 53.78 ± 0.66a–c | 0.597 ± 0.058 | 40.67 ± 1.48bc |
| 9 | 1370 ± 62.7a | 17.71 ± 2.49c | 8503 ± 84.72 | 5.29 ± 0.39a–c | 51.13 ± 0.80d | 0.591 ± 0.051 | 38.52 ± 1.09b–d | |
| 2 | 1263 ± 21.9c | 17.05 ± 2.63c | 8473 ± 72.38 | 4.42 ± 0.31cd | 53.19 ± 0.75c | 0.617 ± 0.052 | 31.12 ± 1.14h | |
| 6 | 0.04 | 1279 ± 42.0c | 18.09 ± 2.57c | 8488 ± 89.84 | 4.73 ± 0.66b–d | 55.31 ± 0.88ab | 0.580 ± 0.041 | 35.43 ± 0.99de |
| 9 | 1310 ± 30.4b | 17.42 ± 1.66c | 8482 ± 75.05 | 4.16 ± 0.54d | 54.02 ± 0.76ac | 0.635 ± 0.049 | 33.74 ± 1.16e–h | |
| 2 | 1307 ± 36.1b | 18.59 ± 1.67c | 8500 ± 59.32 | 4.87 ± 0.57a–d | 51.04 ± 0.64d | 0.659 ± 0.102 | 28.13 ± 0.94hi | |
| 6 | 0.08 | 1299 ± 33.2b | 17.35 ± 1.48c | 8477 ± 93.41 | 5.07 ± 0.61a–c | 54.28 ± 0.93a–c | 0.668 ± 0.044 | 31.37 ± 1.21f–h |
| 9 | 1313 ± 42.2b | 18.28 ± 2.53c | 8492 ± 84.85 | 5.34 ± 0.43ab | 52.95 ± 0.75cd | 0.584 ± 0.057 | 26.48 ± 0.87i | |
| Main effects | ||||||||
| S | ||||||||
| 2 | 1330 ± 52.4 | 17.76 ± 1.48b | 8488 ± 70.21 | 4.95 ± 0.37 | 51.56 ± 0.43b | 0.632 ± 0.027 | 32.61 ± 0.73c | |
| 6 | 1335 ± 41.4 | 18.72 ± 1.52a | 8482 ± 82.08 | 5.22 ± 0.23 | 54.19 ± 0.41a | 0.608 ± 0.024 | 38.26 ± 1.64a | |
| 9 | 1342 ± 59.0 | 18.55 ± 1.41ab | 8494 ± 70.83 | 4.98 ± 0.69 | 53.46 ± 0.48a | 0.618 ± 0.019 | 35.34 ± 0.68b | |
| Ni | ||||||||
| 0 | 1379 ± 44.8a | 20.05 ± 1.64a | 8487 ± 91.37 | 5.44 ± 0.45a | 52.10 ± 0.67c | 0.630 ± 0.037 | 41.02 ± 0.70a | |
| 0.0004 | 1374 ± 51.3a | 17.71 ± 1.58b | 8493 ± 72.44 | 5.22 ± 0.68a | 53.24 ± 0.48ab | 0.599 ± 0.018 | 38.49 ± 0.75b | |
| 0.04 | 1283 ± 32.4b | 17.52 ± 1.46b | 8481 ± 63.36 | 4.44 ± 0.47b | 54.17 ± 0.41a | 0.611 ± 0.022 | 33.43 ± 0.63c | |
| 0.08 | 1306 ± 42.3ab | 18.07 ± 1.53b | 8490 ± 70.09 | 5.09 ± 0.64a | 52.76 ± 0.45bc | 0.637 ± 0.028 | 28.66 ± 0.59d | |
| Statistical significance | ||||||||
| S | NS | * | NS | NS | * | NS | * | |
| Ni | * | * | NS | * | * | NS | * | |
| S × Ni | * | * | NS | * | * | NS | * | |
| Concentration in the Nutrient Solution (mM) | Concentration of the Micronutrients in the Shoots (mg kg−1 DM) | |||||||
|---|---|---|---|---|---|---|---|---|
| S | Ni | Fe | B | Cl | Cu | Mn | Mo | Zn |
| 2 | 114.27 ± 3.29ab | 42.73 ± 2.89cd | 12,359 ± 85.2a | 5.96 ± 0.19b | 26.72 ± 1.55a | 0.983 ± 0.057a | 42.15 ±0.87c–e | |
| 6 | 0.00 | 111.86 ± 2.91ab | 49.36 ± 1.14b | 12,311 ± 98.5bc | 6.88 ± 0.15a | 22.45 ± 1.63e | 1.052 ± 0.071a | 40.73 ± 1.11e |
| 9 | 115.09 ± 3.84ab | 54.22 ± 2.93a | 12,348 ± 60.3ab | 7.25 ± 0.26a | 23.96 ± 1.72de | 0.928 ± 0.049a–d | 41.06 ± 1.03e | |
| 2 | 115.24 ± 4.09a | 35.07 ± 1.28g | 12,351 ± 97.1ab | 4.12 ± 0.34d | 24.02 ± 1.53d | 0.770 ± 0.078b–d | 43.87 ± 0.75b–d | |
| 6 | 0.0004 | 103.26 ± 3.71c–e | 42.06 ± 1.05d | 12,301 ± 63.0c | 5.68 ± 0.26b | 25.69 ± 1.47a–c | 0.981 ± 0.043a | 47.67 ± 1.17a |
| 9 | 117.74 ± 3.35a | 44.85 ± 1.35c | 12,275 ± 60.3c | 5.81 ± 0.21b | 25.53 ± 1.61a–c | 0.975 ± 0.065a | 45.38 ± 1.21ab | |
| 2 | 88.22 ± 5.29g | 34.72 ± 1.07g | 12,385 ± 51.4a | 4.01 ± 0.28d | 24.72 ± 1.55cd | 0.759 ± 0.057d | 41.75 ± 1.09de | |
| 6 | 0.04 | 97.29 ± 4.07ef | 36.94 ± 1.22fg | 12,149 ± 88.7d | 5.16 ± 0.20bc | 26.30 ± 1.59ab | 0.960 ± 0.048a–c | 44.29 ± 0.86bc |
| 9 | 101.56 ± 5.61de | 38.12 ± 1.24f | 12,298 ± 36.0c | 5.92 ± 0.24b | 26.91 ± 1.47a | 0.972 ± 0.061a | 47.17 ± 1.14a | |
| 2 | 93.81 ± 4.48fg | 35.48 ± 1.79fg | 12,377 ± 51.6a | 4.35 ± 0.21cd | 25.19 ± 1.62b–d | 0.763 ± 0.066cd | 44.31 ± 0.92bc | |
| 6 | 0.08 | 104.77 ± 3.02cd | 37.35 ± 2.14fg | 12,304 ± 68.3c | 5.37 ± 0.22b | 26.73 ± 1.53a | 0.970 ± 0.072ab | 46.97 ± 1.17a |
| 9 | 110.01 ± 5.23bc | 41.37 ± 1.33d | 12,092 ± 54.2e | 5.59 ± 0.31b | 26.70 ± 1.58a | 0.967 ± 0.054ab | 47.06 ± 1.20a | |
| Main effects | ||||||||
| S | ||||||||
| 2 | 102.89 ± 4.61b | 37.00 ± 1.98c | 12,368 ± 78.2a | 4.61 ± 0.14b | 25.16 ± 1.44 | 0.819 ± 0.037b | 43.02 ± 0.47b | |
| 6 | 104.30 ± 3.57b | 41.43 ± 1.45b | 12,266 ± 87.1ab | 5.77 ± 0.11a | 25.29 ± 1.52 | 0.991 ± 0.029a | 44.92 ± 0.44a | |
| 9 | 111.10 ± 4.79a | 44.64 ± 1.83a | 12,253 ± 56.2b | 6.14 ± 0.19a | 25.78 ± 1.47 | 0.961 ± 0.031a | 45.17 ± 0.50a | |
| Ni | ||||||||
| 0 | 113.74 ± 2.97a | 48.77 ± 2.65a | 12,399 ± 84.3a | 6.70 ± 0.11a | 24.38 ± 1.42b | 0.988 ± 0.027 | 41.31 ± 0.78c | |
| 0.0004 | 112.08 ± 3.65a | 40.66 ± 1.23b | 12,309 ± 74.4a | 5.20 ± 0.17b | 25.08 ± 1.43ab | 0.909 ± 0.022 | 45.64 ± 0.52b | |
| 0.04 | 95.69 ± 4.72b | 36.59 ± 1.12d | 12,277 ± 71.1b | 5.03 ± 0.22b | 25.98 ± 1.51a | 0.897 ± 0.032 | 44.40 ± 0.47b | |
| 0.08 | 102.86 ± 4.53a | 38.07 ± 1.76c | 12258 ± 59.2b | 5.10 ± 0.13b | 26.21 ± 1.48a | 0.900 ± 0.029 | 56.11 ± 0.51a | |
| Statistical significance | ||||||||
| S | * | * | * | * | NS | * | * | |
| Ni | * | * | * | * | * | NS | * | |
| S × Ni | * | * | * | * | * | * | * | |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Matraszek-Gawron, R.; Hawrylak-Nowak, B. Micronutrient Status and Selected Physiological Parameters of Roots in Nickel-Exposed Sinapis alba L. Affected by Different Sulphur Levels. Plants 2019, 8, 440. https://doi.org/10.3390/plants8110440
Matraszek-Gawron R, Hawrylak-Nowak B. Micronutrient Status and Selected Physiological Parameters of Roots in Nickel-Exposed Sinapis alba L. Affected by Different Sulphur Levels. Plants. 2019; 8(11):440. https://doi.org/10.3390/plants8110440
Chicago/Turabian StyleMatraszek-Gawron, Renata, and Barbara Hawrylak-Nowak. 2019. "Micronutrient Status and Selected Physiological Parameters of Roots in Nickel-Exposed Sinapis alba L. Affected by Different Sulphur Levels" Plants 8, no. 11: 440. https://doi.org/10.3390/plants8110440
APA StyleMatraszek-Gawron, R., & Hawrylak-Nowak, B. (2019). Micronutrient Status and Selected Physiological Parameters of Roots in Nickel-Exposed Sinapis alba L. Affected by Different Sulphur Levels. Plants, 8(11), 440. https://doi.org/10.3390/plants8110440