De Novo Assembly Discovered Novel Structures in Genome of Plastids and Revealed Divergent Inverted Repeats in Mammillaria (Cactaceae, Caryophyllales)

 ,
,
Abstract
1. Introduction
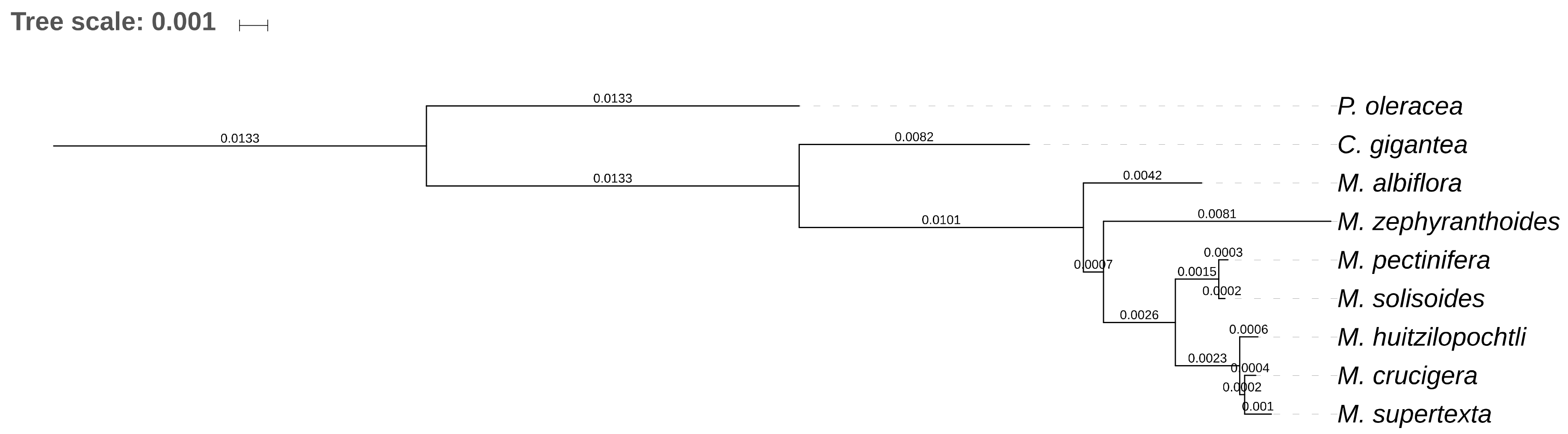
2. Results
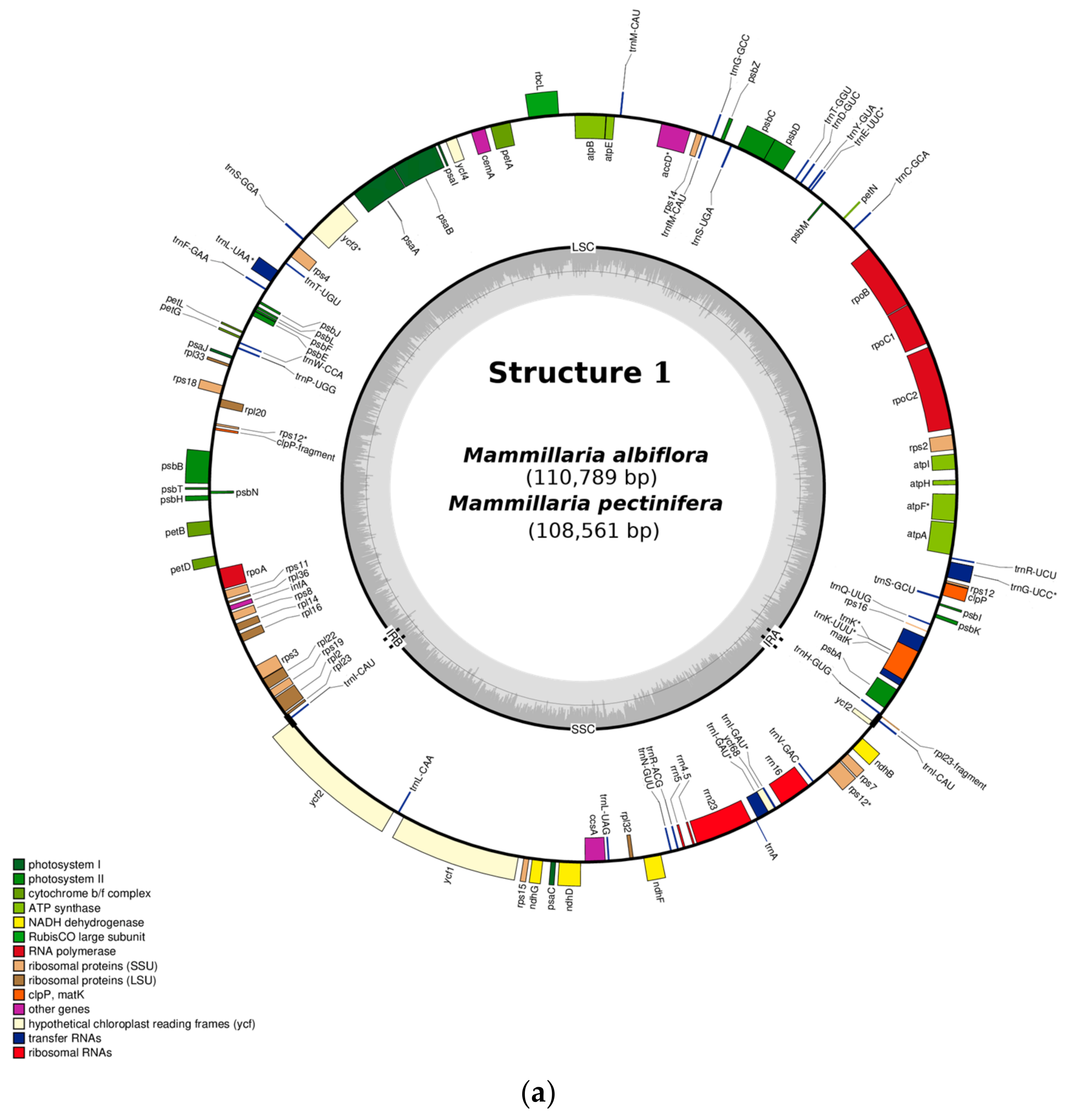
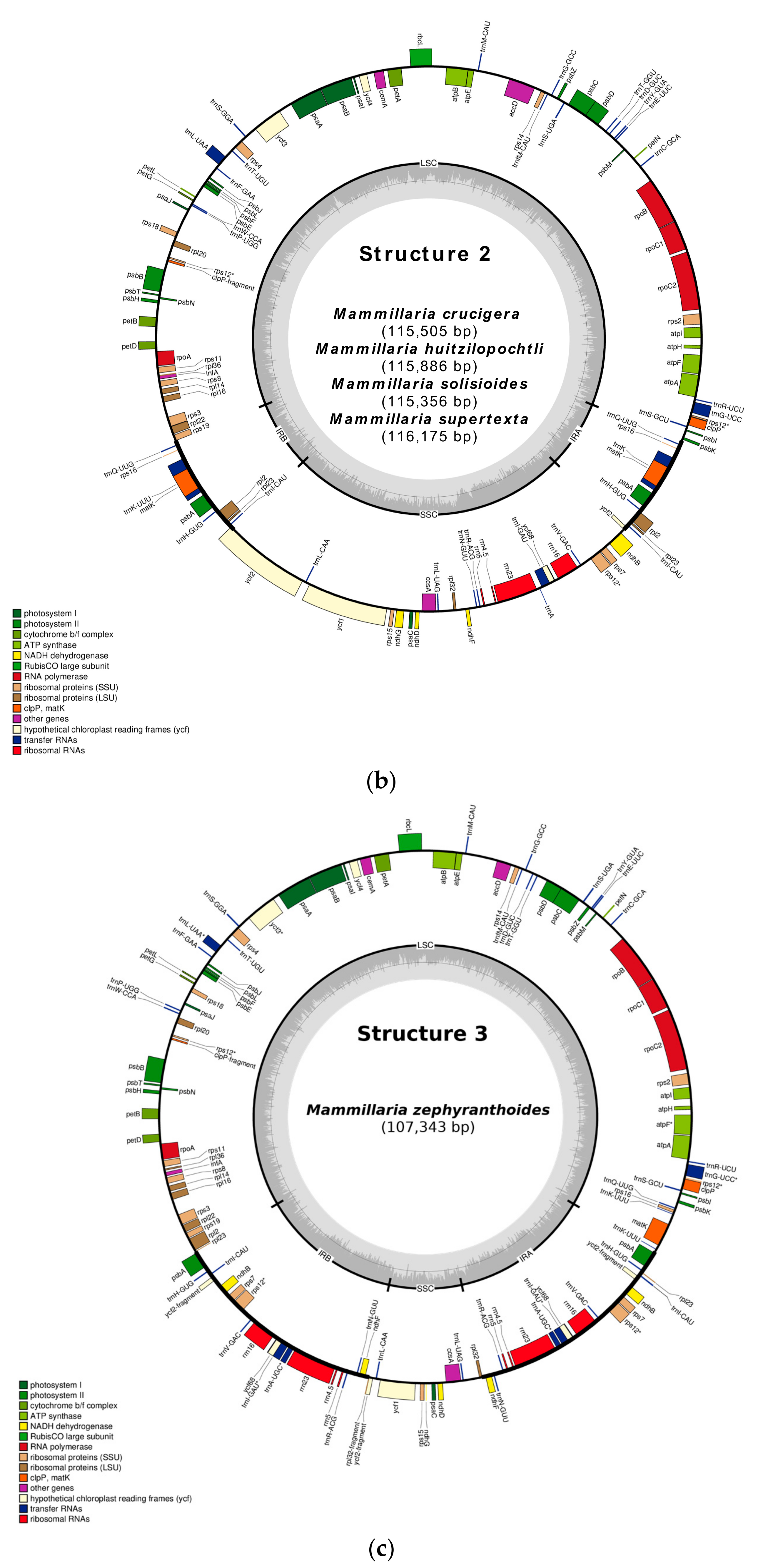
2.1. Gene Composition and Length Variation in Three Novel cpDNA Structures Identified in Mammillaria
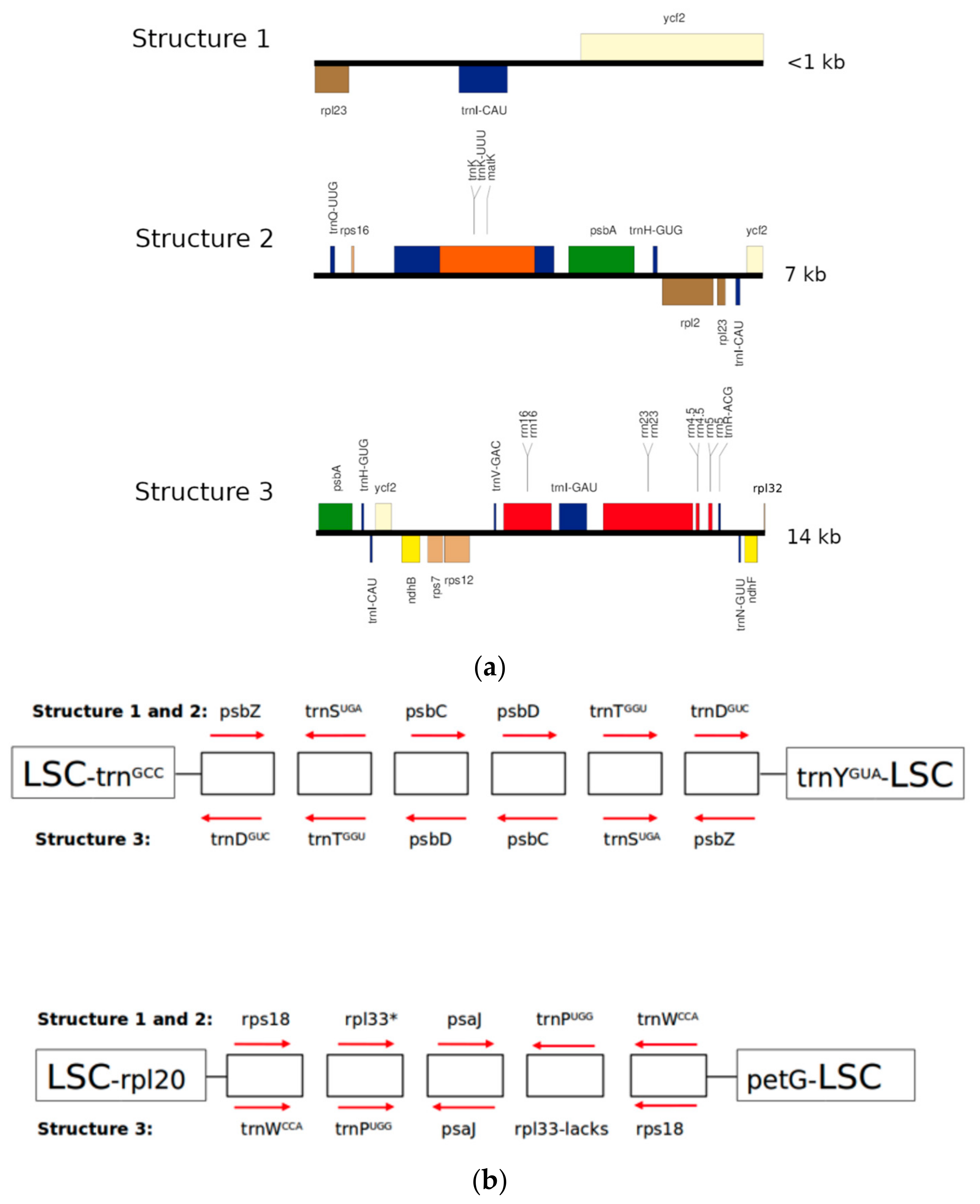
2.2. Structure 1: Shortest IRs, Composed of Three Genes, rpl23-trnI-CAU-ycf2
2.3. Structure 2: IRs Composed by an Unusual Complete Battery of 11 Concatenated Genes and One Identical Intergenic Spacer.
2.4. Structure 3: Largest and Divergent IRs in Which Four Ribosomal Units are Included.
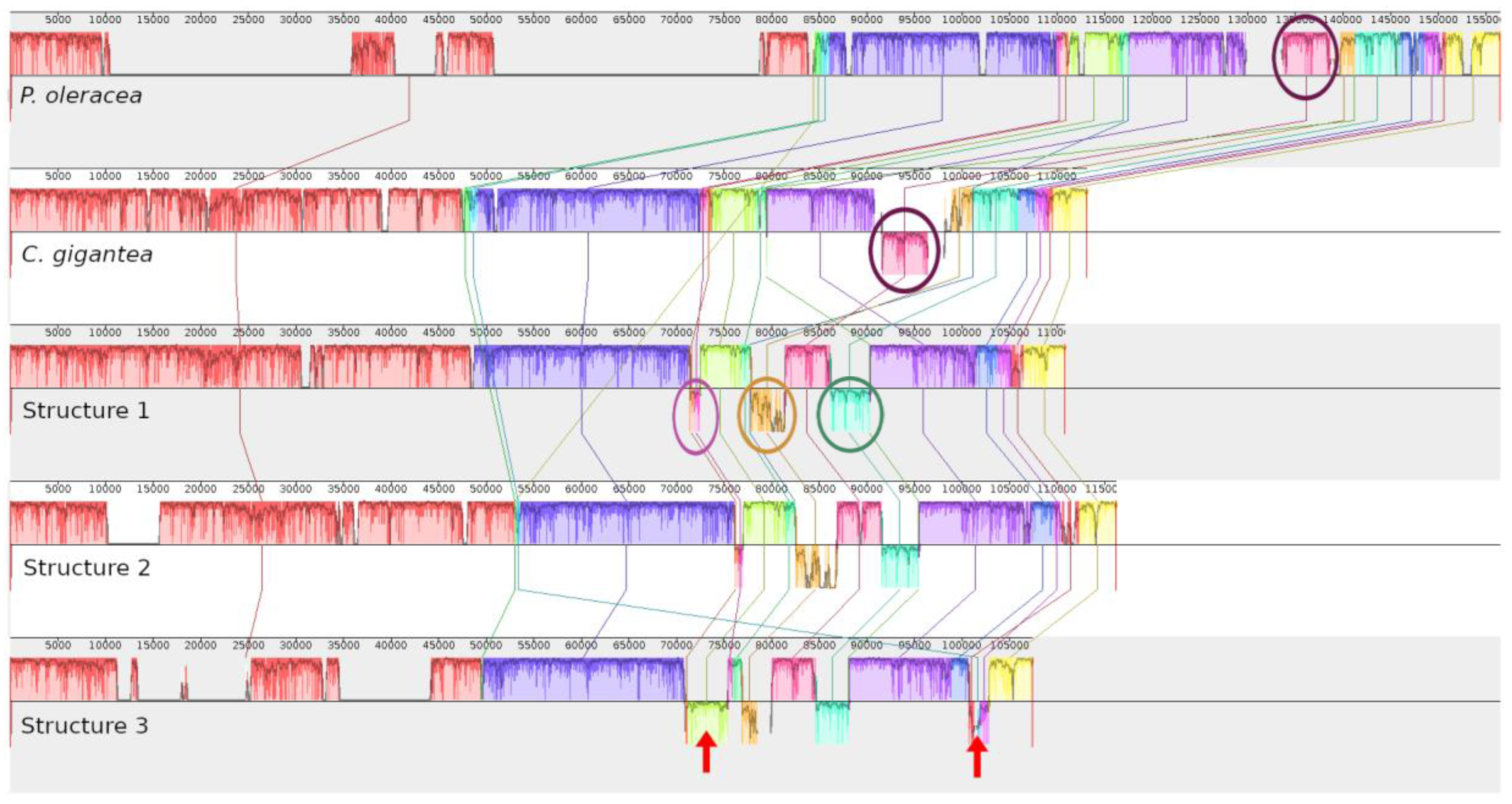
3. Discussion
4. Materials and Methods
4.1. Plant Sampling and DNA Extraction
4.2. High-Throughput Sequencing and Sanger Verification
4.3. Genome Assembly, Annotation, and Structural Alignment
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Daniell, H.; Lin, C.S.; Yu, M.; Chang, W.J. Chloroplast genomes: Diversity, evolution, and applications in genetic engineering. Genome Biol. 2016, 17, 134. [Google Scholar] [CrossRef] [PubMed]
- Morden, C.; Delwiche, C.; Kuhsel, M.; Palmer, D. Gene phylogenies and the endosymbiotic origin of plastids. Biosystems 1992, 28, 75–90. [Google Scholar] [CrossRef]
- Cavalier-Smith, T. The origins of plastids. Biol. J. Linn. Soc. 1982, 17, 289–306. [Google Scholar] [CrossRef]
- Lemieux, C.; Otis, C.; Turnel, M. Ancestral chloroplast genome in Mesostigma viride reveals an early branch of green plant evolution. Nature 2000, 403, 649–652. [Google Scholar] [CrossRef] [PubMed]
- Xiao-Ming, Z.; Junrui, W.; Li, F.; Sha, L.; Hongbo, P.; Lan, Q.; Jing, L.; Yan, S.; Weihua, Q.; Lifang, Z.; et al. Inferring the evolutionary mechanism of the chloroplast genome size by comparing whole-chloroplast genome sequences in seed plants. Sci. Rep. 2017, 7, 1555. [Google Scholar] [CrossRef] [PubMed]
- Thorsness, P.E.; Weber, E.R. Escape and migration of nucleic acids between chloroplasts, mitochondria, and the nucleus. Int. Rev. Cytol. 1996, 165, 207–234. [Google Scholar] [CrossRef]
- Yagi, Y.; Shiina, T. Recent advances in the study of chloroplast gene expression and its evolution. Front. Plant Sci. 2014, 5, 61. [Google Scholar] [CrossRef]
- Kung, S.D.; Lin, C.M. Chloroplast promoters from higher plants. Nucleic Acids Res. 1985, 13, 7543–7549. [Google Scholar] [CrossRef][Green Version]
- Mower, J.P.; Vickrey, T.L. Advances in Botanical Research Plastid Genome Evolution; Chaw, S.M., Jansen, R.K., Eds.; Academic Press of Elsevier: London, UK, 2018; Volume 85, Chapter 9; pp. 263–292. [Google Scholar]
- Zhu, A.; Guo, W.; Sakski, G.; Weishu, F.; Mover, J.P. Evolutionary dynamics of the plastid inverted repeat: The effects of expansion, contraction, and loss of substitution rates. New Phytol. 2016, 209, 1747–1756. [Google Scholar] [CrossRef]
- Lavin, M.; Doyle, J.J.; Palmer, J.D. Evolutionary significance of the loss of the chloroplast-DNA Inverted Repeat in the Leguminosae Subfamily Papilionoideae. Evolution 1990, 44, 390–402. [Google Scholar] [CrossRef]
- Sanderson, M.J.; Copetti, D.; Búrquez, A.; Bustamante, E.; Charboneau, J.L.M.; Eguiarte, L.; Kumar, S.; Lee, H.O.; McMahon, M.; Steele, K.; et al. Exceptional reduction of the plastid genome of saguaro cactus (Carnegiea gigantea). Am. J. Bot. 2015, 102, 1115–1127. [Google Scholar] [CrossRef] [PubMed]
- Yao, G.; Jin, J.J.; Li, H.T.; Yang, J.B.; Mandala, V.S.; Croley, M.; Mostow, R.; Douglas, N.A.; Chase, M.W.; Christenhusz, M.J.M.; et al. Plastid phylogenomic insights into the evolution of Caryophyllales. Mol. Phylogenet. Evol. 2019, 134, 74–86. [Google Scholar] [CrossRef] [PubMed]
- Huang, H.; Shi, C.; Liu, Y.; Mao, S.Y.; Gao, L.Z. Thirteen Camellia chloroplast genome sequences determined by high-throughput sequencing: Genome structure and phylogenetic relationships. BMC Evol. Biol. 2014, 14, 151. [Google Scholar] [CrossRef] [PubMed]
- Wu, Z.; Tembrock, L.R. Two complete chloroplast genomes of white campion (Silene latifolia) from male and female individuals. Mitochondrial DNA Part A 2015, 28, 375–376. [Google Scholar] [CrossRef]
- Kang, J.S.; Lee, B.Y.; Kwak, M. The complete chloroplast genome sequences of Lychnis wilfordii and Silene capitata and comparative analyses with other Caryophyllaceae genomes. PLoS ONE 2017, 27, e0172924. [Google Scholar] [CrossRef] [PubMed]
- Blazier, J.C.; Jansen, R.K.; Mower, J.P.; Govindu, M.; Zhang, J.; Weng, M.L.; Ruhlman, T.A. Variable presence of the inverted repeat and plastome stability in Erodium. Ann. Bot. 2016, 117, 1209–1220. [Google Scholar] [CrossRef]
- Hunt, D.; Taylor, N.; Charles, G. The New Cactus Lexicon; DH Books: Milborne Port, UK, 2006. [Google Scholar]
- IUCN International Union for Conservation of Nature. Consulted for Mammillaria genus. Available online: https://www.iucnredlist.org/search/list?query=Mammillaria&searchType=species (accessed on 30 June 2019).
- Butterworth, C.A.; Wallace, R.S. Phylogenetic studies of Mammillaria (Cactaceae)—Insights from chloroplast sequence variation and hypothesis testing using the parametric bootstrap. Am. J. Bot. 2004, 91, 1086–1098. [Google Scholar] [CrossRef] [PubMed]
- Crozier, B.S. Systematics of Cactaceae Juss: Phylogeny, cpDNA Evolution, and Classification, with Emphasis on the Genus Mammillaria Haw. Ph.D. Thesis, University of Texas, Austin, TX, USA, 2005. [Google Scholar]
- Hernández-Hernández, T.; Hernández, H.M.; De Nova, J.A.; Puente, R.; Eguiarte, L.; Magallón, S. Phylogenetic relationships and evolution of growth form in Cactaceae (Caryophyllales, Eudicotyledoneae). Am. J. Bot. 2011, 98, 44–61. [Google Scholar] [CrossRef]
- Arakaki, M.; Christin, P.A.; Nyffeler, R.; Lendel, A.; Eggli, U.; Ogburn, M.; Spriggs, E.; Moore, M.J.; Edwards, E.J. Contemporaneous and recent radiations of the world’s major succulent plant lineages. Proc. Natl. Acad. Sci. USA 2011, 108, 8379–8384. [Google Scholar] [CrossRef]
- Turmel, M.; Otis, C.; Lemieux, C. Divergent copies of the large inverted repeat in the chloroplast genomes of ulvophycean green algae. Sci. Rep. 2017, 7, 994. [Google Scholar] [CrossRef]
- Ocampo, G.; Columbus, J.T. Molecular phylogenetics of suborder Cactineae (Caryophyllales), including insight into photosynthetic diversification and historical biogeography. Am. J. Bot. 2010, 97, 1827–1847. [Google Scholar] [CrossRef] [PubMed]
- Alkatib, S.; Fleischmann, T.T.; Scharff, L.B.; Bock, R. Evolutionary constraints on the plastid tRNA set decoding methionine and isoleucine. Nucleic Acid Res. 2012, 40, 6713–6724. [Google Scholar] [CrossRef] [PubMed]
- Ni, Z.; Ye, Y.; Bal, T.; Xu, M.; Xu, L.A. Complete chloroplast genome of Pinus massoniana (Pinaceae): Gene rearrangements, loss of ndh genes, and short inverted repeats contraction, expansion. Molecules 2017, 22, 1528. [Google Scholar] [CrossRef]
- Hao, D.C.; Chen, S.L.; Huang, B.L. Evolution of the chloroplast trnL-trnF region in the Gymnosperm lineages Taxaceae and Cephalotaxaceae. Biochem. Genet. 2009, 47, 351–369. [Google Scholar] [CrossRef] [PubMed]
- Drescher, A.; Ruf, S.; Calsa, T., Jr.; Carrer, H.; Bock, R. The two largest chloroplast genome-encoded open reading frames of higher plants are essential genes. Plant J. 2000, 22, 97–104. [Google Scholar] [CrossRef] [PubMed]
- Park, S.; An, B.; Park, S. Reconfiguration of the plastid genome in Lamprocapnos spectabilis: IR boundary shifting, inversion, and intraspecific variation. Sci. Rep. 2018, 8, 13568. [Google Scholar] [CrossRef] [PubMed]
- Barthet, M.M.; Hilu, K.W. Expression of matK: Functional and evolutionary implications. Am. J. Bot. 2007, 94, 1402–1412. [Google Scholar] [CrossRef]
- Arias, S.; Gama-López, S.; Guzmán-Cruz, L.U.; Vázquez-Benítez, B. Cactaceae. In Flora del Valle de Tehuacán-Cuicatlán; Medina, L.R., Ed.; Instituto de Biología, Universidad Nacional Autónoma de México: Mexico City, Mexico, 2012; Volume 95. [Google Scholar]
- Rozen, S.; Skaletsky, H. Primer3 on the WWW for general users and for biologist programmers. Methods Mol. Biol. 2000, 132, 365–386. [Google Scholar] [CrossRef]
- Dierckxsens, N.; Mardulyn, P.; Smits, G. NOVOPlasty: De novo assembly of organelle genomes from whole genome data. Nucleic Acids Res. 2017, 45, e18. [Google Scholar] [CrossRef]
- Love, R.R.; Weisenfeld, N.I.; Jaffe, D.B.; Besansky, N.J.; Neafsey, D.E. Evaluation of DISCOVAR de novo using a mosquito simple for cost-effective short-read genome assembly. BMC Genom. 2016, 17, 18. [Google Scholar] [CrossRef]
- Kolmogorov, M.; Raney, B.; Paten, B.; Pham, S. Ragout-a reference-assisted assembly tool for bacterial genomes. Bioinformatics 2014, 30, i302–i309. [Google Scholar] [CrossRef] [PubMed]
- Soto-Jiménez, L.M.; Estrada, K.; Sanchez-Flores, A. GARM: Genome assembly, reconciliation and merging pipeline. Curr. Top. Med. Chem. 2014, 14, 418–424. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.; De Sila, N.; Otto, T.D.; Parkhill, J.; Keane, J.A.; Harris, S.R. Circlator: Automated circularization of genome assemblies using long sequencing reads. Genome Biol. 2015, 16, 294. [Google Scholar] [CrossRef] [PubMed]
- Hunt, M.; Kikuchi, T.; Sanders, M.; Newbold, C.; Berriman, M.; Otto, T.D. REAPR: A universal tool for genome assembly evaluation. Genome Biol. 2013, 14, R47. [Google Scholar] [CrossRef] [PubMed]
- Tillich, M.; Lehwark, P.; Pellizzer, T.; Ulbricht-Jones, E.S.; Fischer, A.; Bock, R.; Greiner, S. GeSeq–versatile and accurate annotation of organelle genomes. Nucleic Acids Res. 2017, 45, W6–W11. [Google Scholar] [CrossRef] [PubMed]
- Lohse, M.; Drechsel, O.; Bock, R. OrganellarGenomeDRAW (OGDRAW): A tool for the easy generation of high-quality custom graphical maps of plastid and mitochondrial genomes. Curr. Genet. 2007, 52, 267–274. [Google Scholar] [CrossRef]
- Liu, X.; Yang, H.; Zhao, J.; Zhou, B.; Li, T.; Xiang, B. The complete chloroplast genome sequence of the folk medicinal and vegetable plant purslane (Portulaca oleracea L.). J. Hortic. Sci. Biotech. 2018, 93, 356–365. [Google Scholar] [CrossRef]
- Darling, A.C.; Mau, B.; Blattner, F.R.; Perna, N.T. Mauve: Multiple alignment of conserved genomic sequence with rearrangements. Genome Res. 2004, 14, 1394–1403. [Google Scholar] [CrossRef]
- Darriba, D.; Taboada, G.L.; Doallo, R.; Posada, D. ProtTest 3: Fast selection of best-fit models of protein evolution. Bioinformatics 2011, 27, 1164–1165. [Google Scholar] [CrossRef]
- Katoh, K.; Daron, M.S. MAFFT multiple sequence alignment software version 7: Improvements in performance and usability. Mol. Biol. Evol. 2013, 30, 772–780. [Google Scholar] [CrossRef]
- Posada, D. JModelTest: Phylogenetic model averaging. Mol. Biol. Evol. 2008, 25, 1253–1256. [Google Scholar] [CrossRef] [PubMed]
- Stamatakis, A. RAxML version 8: A tool for phylogenetic analysis and post-analysis of large phylogenies. Bioinformatics 2014, 30, 1312–1313. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Type of Structure | Total Length | IRs | LSC | SCC | Total Number of Genes | Access Number 1 |
|---|---|---|---|---|---|---|
| I. Structure 1 | ||||||
| 1.1 M. albiflora | 110789 | 1348 | 78380 | 31061 | 113 | MN517610 |
| 1.2. M. pectinifera | 108561 | 1544 | 72273 | 29744 | 113 | MN519716 |
| II. Structure 2 | ||||||
| 1. M. crucigera | 115505 | 14522 | 71565 | 29418 | 120 | MN517613 |
| 2. M. huitzilopochtli | 115886 | 14488 | 71997 | 29401 | 120 | MN517612 |
| 3. M. solisioides | 115356 | 14428 | 71690 | 29238 | 120 | MN518341 |
| 4. M. supertexta | 116175 | 14490 | 72240 | 29445 | 119 | MN508963 |
| III. Structure 3 | ||||||
| 1. M. zephyranthoides | 107343 | 28252 | 71811 | 7281 | 131 | MN517611 |
| Gene Type/Structure | Region | Structure 1 | Structure 2 | Structure 3 |
|---|---|---|---|---|
| 1. Ribosomal RNA (rrn) | SSC | rrn4.5, 5,16, 23 | rrn4.5, 5,16, 23 | |
| IRs | rrn4.5, 5,16, 23 (2X) | |||
| 2. Transfer RNA (trn) | LSC | trnCGCA, trnDGUC, trnEUUC, trnFGAA, trnGGCC, trnGUCC, trnHGUG, trnKUUU, trnLUAA, trnMCAU, trnMCAU, trnPUGG, trnQUUG, trnRUCU, trnSGGA, trnSGGU, trnSUGA, trnTGGU, trnTUGU, trnYGUA | trnCGCA, trnDGUC, trnEUUC, trnFGAA, trnGGCC, trnGUCC, trnLUAA, trnMCAU, trnfMCAU, trnPUGG, trnRUCU, trnSGGA, trnSGCU, trnSUGA, trnTGGU, trnTUGU, trnWCCA, trnYGUA | trnCGCA, trnDGUC, trnEUUC, trnFGAA, trnGGCC, trnGUCC, trnKUUU, trnLUAA, trnMCAU, trnfMCAU, trnPUGG, trnQUUG, trnRUCU, trnSGGA, trnSGCU, trnSUGA, trnTGGU, trnTUGU, trnWCCA, trnYGUA |
| SSC | trnA-f, trnIGAU, trnIGAU, trnLCAA, trnLUAG, trnNGUU, trnRACG, trnVGAG | trnA-f, trnIGAU, trnIUAG, trnNGUU, trnLCAA, trnRACG, trnVGAG | trnLUAG, trnLCAA | |
| IRs | trnICAU (2X) | trnHGUG,tmICAU, trnKUUU, trnQUUG (2X) | trnAUGC, trnHGUG, trnICAU, trnIGAU, trnNGUU, trnRACG, trnVGAC (2X) | |
| 3. Proteins of small subunits of the ribosome (rps) | LSC | rps2, 3, 4, 8, 11, 12 (2), 14, 16Ψ, 18Ψ, 19 | rps2, 3, 4, 8, 11, 12 (2), 14, 18Ψ, 19 | rps2, 3, 4, 8 11, 12, 12Ψ, 14, 16Ψ, 18Ψ, 19 |
| SSC | rps7, 12, 15 | rps7, 12, 15 | rps15 | |
| IRs | rps16Ψ (2X) | rps7, 12, (2X) | ||
| 4. Proteins of large subunits of the ribosome (rpl) | LSC | rpl2, 14, 16, 20, 22, 33Ψ, 36Ψ | rpl14, 16Ψ, 20, 22,33Ψ*, 36Ψ | rpl2, 14, 16Ψ, 20, 22, 23Ψ, 36 |
| SSC | rpl32 | rpl32 | ||
| IRs | rpl23Ψ (2X) | rpl2, 23Ψ (2X) | rpl32 (2X), 23Ψ (IRA) | |
| 5. DNA dependent RNA polymerase (rpo) | LSC | rpoA, B, C1, C2 | rpoA, B, C1, C2 | rpoA, B, C1, C2, |
| 6. NADH dehydrogenase (ndh) | SSC | ndhBΨ, DΨ, FΨ, GΨ*** | ndhBΨ, DΨ, FΨ, G*** | |
| IRs | ndhBΨ, DΨ, FΨ, GΨ (2X) | |||
| 7. Photosystem I (psa) | LSC | psaA, B, I, J | psaA, B, I, J | psaA, B, I, J |
| SSC | psaC | psaC | psaC | |
| 8. Photosystem II (psb) | LSC | psbA, B, C, D, E, F, H, I, J, K, L, M, N, T, Z | psbB, C, D, E, F, H, I, J, K, L, M, N, T, Z | psbB, C, D, E, F, H, I, J, L, K, M, N T, Z |
| IRs | psbA (2X) | psbA (2X) | ||
| 9. Cytochrome b/f complex (pet) | LSC | petA, B, D, G, L, N | petA, B, D, G, L, N | petA, B, D, G, L, N |
| 10. ATP synthase (atp) | LSC | atpA, B, E, F, H, I | atpA, B, E, F, H, I | atpA, B, E, F, H, I |
| 11. Rubisco (rbc) | LSC | rbcL | rbcL | rbcL |
| 12. Maturase K | LSC | matK | matK | |
| IRs | matK (2X) | |||
| 13. Protease (clp) | LSC | clpPΨ, clpP | clpPΨ, clpP | clpPΨ, clpP |
| 14. Envelope membrane protein (cem) | LSC | cemA | cemA | cemA |
| 15. Subunit of acetil-CoA-carboxylase (acc) | LSC | accDΨ | accDΨ | accDΨ |
| 16. c-type cytochrome synthesis (ccs) | SSC | ccsA | SSC: ccsA | SSC: ccsA |
| 17. Translational initiation factor (inf) | LSC | infA | infA | infA |
| 18. Hypothetical chloroplast reading frames (ycf) | LSC | ycf3, ycf4Ψ | ycf3, ycf4Ψ** | ycf3, ycf4 |
| SSC | ycf1, ycf2, ycf68Ψ | ycf1, ycf2, ycf68Ψ | ycf1Ψ, ycf2Ψ | |
| IRs | ycf2-p (2X) | ycf2-p (2X) | ycf2Ψ, ycf68Ψ (2X) |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Solórzano, S.; Chincoya, D.A.; Sanchez-Flores, A.; Estrada, K.; Díaz-Velásquez, C.E.; González-Rodríguez, A.; Vaca-Paniagua, F.; Dávila, P.; Arias, S. De Novo Assembly Discovered Novel Structures in Genome of Plastids and Revealed Divergent Inverted Repeats in Mammillaria (Cactaceae, Caryophyllales). Plants 2019, 8, 392. https://doi.org/10.3390/plants8100392
Solórzano S, Chincoya DA, Sanchez-Flores A, Estrada K, Díaz-Velásquez CE, González-Rodríguez A, Vaca-Paniagua F, Dávila P, Arias S. De Novo Assembly Discovered Novel Structures in Genome of Plastids and Revealed Divergent Inverted Repeats in Mammillaria (Cactaceae, Caryophyllales). Plants. 2019; 8(10):392. https://doi.org/10.3390/plants8100392
Chicago/Turabian StyleSolórzano, Sofía, Delil A. Chincoya, Alejandro Sanchez-Flores, Karel Estrada, Clara E. Díaz-Velásquez, Antonio González-Rodríguez, Felipe Vaca-Paniagua, Patricia Dávila, and Salvador Arias. 2019. "De Novo Assembly Discovered Novel Structures in Genome of Plastids and Revealed Divergent Inverted Repeats in Mammillaria (Cactaceae, Caryophyllales)" Plants 8, no. 10: 392. https://doi.org/10.3390/plants8100392
APA StyleSolórzano, S., Chincoya, D. A., Sanchez-Flores, A., Estrada, K., Díaz-Velásquez, C. E., González-Rodríguez, A., Vaca-Paniagua, F., Dávila, P., & Arias, S. (2019). De Novo Assembly Discovered Novel Structures in Genome of Plastids and Revealed Divergent Inverted Repeats in Mammillaria (Cactaceae, Caryophyllales). Plants, 8(10), 392. https://doi.org/10.3390/plants8100392