The Associations between Leaf Morphology, Phenylalanine Ammonia Lyase Activity, Reactive Oxygen Species, and Fusarium Resistance in Selected Species of Wheat with Different Ploidy Levels

 and
and
Abstract
1. Introduction
2. Results
2.1. Susceptibility of Wheat Lines to Pathogenic Infections
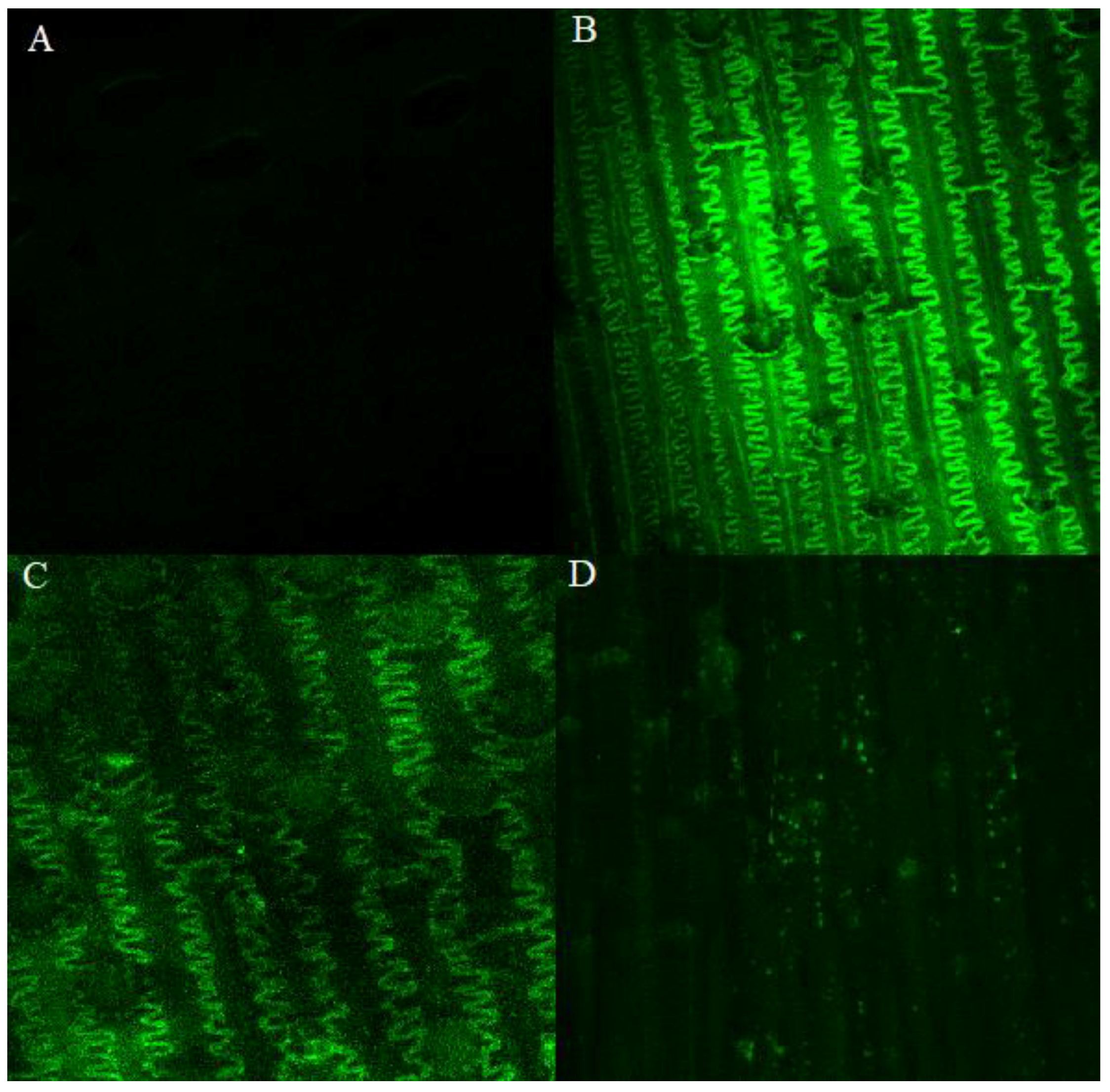
2.2. Leaf Morphology
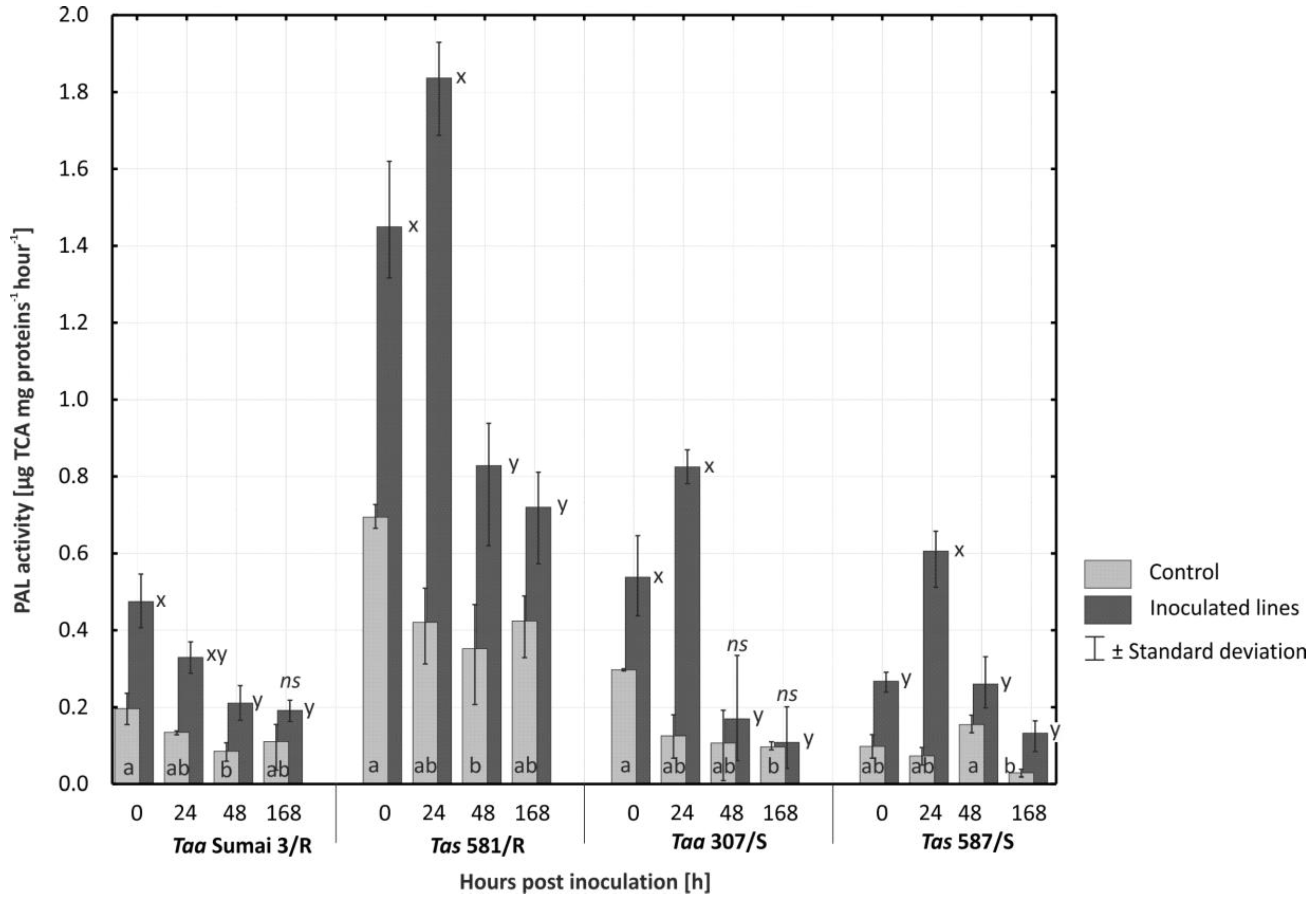
2.3. Activity of PAL
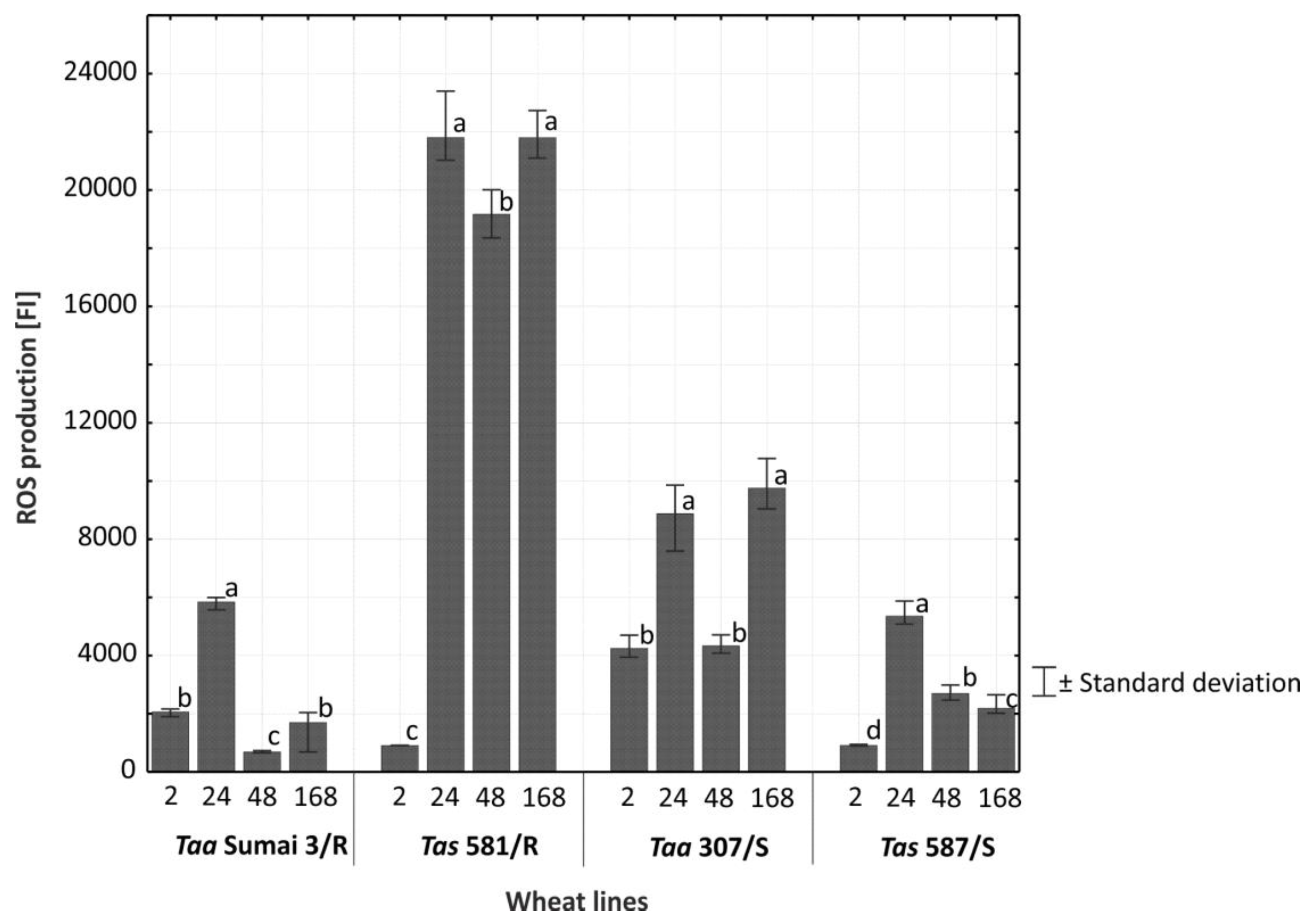
2.4. Reactive Oxygen Species (ROS)
3. Discussion
4. Materials and Methods
4.1. Plant Materials Used in This Study
4.2. Susceptibility to Inoculation with Fusarium culmorum
4.3. Scanning Electron Microscopy (SEM)
4.4. Extraction and Analysis of Phenylalanine Ammonia Lyase (PAL)
4.5. Determination of Reactive Oxygen Species (ROS)
4.6. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Somai-Jemmali, L.; Randoux, B.; Siah, A.; Magnin-Robert, M.; Halama, P.; Reignault, P.; Hamada, W. Similar infection process and induced defense patterns during compatible interactions between Zymoseptoria tritici and both bread and durum wheat species. Eur. J. Plant Pathol. 2017, 147, 787–801. [Google Scholar] [CrossRef]
- Fedak, G. Alien Introgressions from wild Triticum species, T. monococcum, T. urartu, T. turgidum, T. dicoccum, T. dicoccoides, T. carthlicum, T. araraticum, T. timopheevii, and T. miguschovae. In Alien Introgression in Wheat; Molnar, M., Ceoloni, C., Doležel, J., Eds.; Springer International Publishing: Basel, Switzerland, 2015; pp. 191–219. ISBN 978-3-319-23493-9. [Google Scholar]
- Wiwart, M.; Suchowilska, E.; Kandler, W.; Sulyok, M.; Wachowska, U.; Krska, R. The Response of Selected Triticum spp. Genotypes with Different Ploidy Levels to Head Blight Caused by Fusarium culmorum (WG Smith) Sacc. Toxins 2016, 8, 112. [Google Scholar] [CrossRef] [PubMed]
- Wu, H.; Shabala, L.; Barry, K.; Zhou, M.; Shabala, S. Ability of leaf mesophyll to retain potassium correlates with salinity tolerance in wheat and barley. Physiol. Plant. 2013, 149, 515–527. [Google Scholar] [CrossRef] [PubMed]
- Duncan, K.E.; Howard, R.J. Cytological analysis of wheat infection by the leaf blotch pathogen Mycosphaerella graminicola. Mycol. Res. 2013, 104, 1074–1082. [Google Scholar] [CrossRef]
- Scherm, B.; Balmas, V.; Spanu, F.; Pani, G.; Delogu, G.; Pasquali, M.; Migheli, Q. The wheat pathogen Fusarium culmorum. Mol. Plant. Pathol. 2013, 14, 323–341. [Google Scholar] [CrossRef] [PubMed]
- Covarelli, L.; Beccari, G.; Steed, A.; Nicholson, P. Colonization of soft wheat following infection of the stem base by Fusarium culmorum and translocation of deoxynivalenol to the head. Plant Pathol. 2012, 61, 1121–1129. [Google Scholar] [CrossRef]
- Brown, N.A.; Urban, M.; Van de Meene, A.M.L.; Hammond-Kosack, K.E. The infection biology of Fusarium graminearum: Defining the pathways of spikelet to spikelet colonisation in wheat ears. Fungal Biol. 2010, 114, 555–571. [Google Scholar] [CrossRef]
- Xu, X.; Nicholson, P. Community ecology of fungal pathogens causing wheat head blight. Annu. Rev. Phytopathol. 2009, 47, 83–103. [Google Scholar] [CrossRef]
- Buerstmayr, H.; Ban, T.; Anderson, J.A. QTL mapping and marker-assisted selection for Fusarium head blight resistance in wheat: a review. Plant Breeding 2009, 128, 1–26. [Google Scholar] [CrossRef]
- Langevin, F.; Eudes, F.; Comeau, A. Effect of trichothecenes produced by Fusarium graminearum during Fusarium head blight development in six cereal species. Eur. J. Plant Pathol. 2004, 110, 735–746. [Google Scholar] [CrossRef]
- Góral, T.; Ochodzki, P. Fusarium head blight resistance and mycotoxin profiles of four Triticum species genotypes. Phytopathol. Mediterr. 2017, 56, 175–186. [Google Scholar]
- Chekali, S.; Gargouri, S.; Paulitz, T.; Nicol, J.M.; Rezgui, M.; Nasraoui, B. Effects of Fusarium culmorum and water stress on durum wheat in Tunisia. Crop Prot. 2011, 30, 718–725. [Google Scholar] [CrossRef]
- Guo, J.; Zhang, X.; Hou, Y.; Cai, J.; Shen, X.; Zhou, T.; Xu, H.; Ohm, H.W.; Wang, H.; Li, A.; et al. High-density mapping of the major FHB resistance gene Fhb7 derived from Thinopyrum ponticum and its pyramiding with Fhb1 by marker-assisted selection. Theor. Appl. Genet. 2015, 128, 2301–2316. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Q.; Axtman, J.E.; Faris, J.D.; Chao, S.; Zhang, Z.; Friesen, T.L.; Zhong, S.; Cai, X.; Elias, E.M.; Xu, S.S. Identification and molecular mapping of quantitative trait loci for Fusarium head blight resistance in emmer and durum wheat using a single nucleotide polymorphism based linkage map. Mol. Breeding 2014, 34, 1677–1687. [Google Scholar] [CrossRef]
- Wiwart, M.; Suchowilska, E.; Kandler, W.; Sulyok, M.; Groenwald, P.; Krska, R. Can Polish wheat (Triticum polonicum L.) be an interesting gene source for breeding wheat cultivars with increased resistance to Fusarium head blight? Genet. Resour. Crop Ev. 2013, 60, 2359–2373. [Google Scholar] [CrossRef]
- Qi, P.F.; Johnston, A.; Balcerzak, M.; Rocheleau, H.; Harris, L.J.; Long, X.; Wei, Y.M.; Zheng, Y.L.; Ouellet, T. Effect of salicylic acid on Fusarium graminearum, the major causal agent of fusarium head blight in wheat. Fungal Biol. 2012, 116, 413–426. [Google Scholar] [CrossRef]
- Barna, B.; Fodor, J.; Harrach, B.D.; Pogány, M.; Király, Z. The Janus face of reactive oxygen species in resistance and susceptibility of plants to necrotrophic and biotrophic pathogens. Plant Physiol. Bioch. 2012, 59, 37–43. [Google Scholar] [CrossRef]
- Walter, S.; Nicholson, P.; Doohan, F.M. Action and reaction of host and pathogen during Fusarium head blight disease. New Phytol. 2010, 185, 54–66. [Google Scholar] [CrossRef]
- Khaledi, N.; Taheri, P.; Falahati-Rastegar, M. Reactive oxygen species and antioxidant system responses in wheat cultivars during interaction with Fusarium species. Australas. Plant Path. 2016, 45, 653–670. [Google Scholar] [CrossRef]
- Shetty, N.P.; Mehrabi, R.; Lütken, H.; Haldrup, A.; Kema, G.H.; Collinge, D.B.; Jørgensen, H.J.L. Role of hydrogen peroxide during the interaction between the hemibiotrophic fungal pathogen Septoria tritici and wheat. New Phytol. 2007, 174, 637–647. [Google Scholar] [CrossRef]
- Lightfoot, D.J.; Mcgrann, G.R.; Able, A.J. The role of a cytosolic superoxide dismutase in barley–pathogen interactions. Mol. Plant Pathol. 2017, 18, 323–335. [Google Scholar] [CrossRef] [PubMed]
- Trail, F. For blighted waves of grain: Fusarium graminearum in the post genomics era. Plant Physiol. 2009, 149, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Gunnaiah, R.; Kushalappa, A.C. Metabolomics deciphers the host resistance mechanisms in wheat cultivar Sumai-3, against trichothecene producing and non-producing isolates of Fusarium graminearum. Plant Physiol. Bioch. 2014, 83, 40–50. [Google Scholar] [CrossRef] [PubMed]
- Doroshkov, A.V.; Pshenichnikova, T.A.; Afonnikov, D.A. Morphological and genetic characteristics of leaf hairiness in wheat (Triticum aestivum L.) as analyzed by computer-aided phenotyping. Genetika 2011, 47, 836. [Google Scholar]
- Hameed, M.; Mansoor, U.; Ashraf, M.; Rao, A. Variation in leaf anatomy in wheat germplasm from varying drought-hit habitats. Int. J. Agric. Biol. 2002, 4, 12–16. [Google Scholar]
- Lai, A.; Cianciolo, V.; Chiavarini, S.; Sonnino, A. Effects of glandular trichomes on the development of Phytophthora infestans infection in potato (S. tuberosum). Euphytica 2000, 114, 165–174. [Google Scholar] [CrossRef]
- Zarinkamar, F. Density, Size and Distribution of Stornata in Different Monocotyledons. Pak. J. Biol. Sci. 2006, 9, 1650–1659. [Google Scholar]
- Váry, Z.; Mullins, E.; McElwain, J.C.; Doohan, F.M. The severity of wheat diseases increases when plants and pathogens are acclimatized to elevated carbon dioxide. Glob. Change Biol. 2015, 21, 2661–2669. [Google Scholar] [CrossRef]
- Devireddy, A.R.; Zandalinas, S.I.; Gómez-Cadenas, A.; Blumwald, E.; Mittler, R. Coordinating the overall stomatal response of plants: Rapid leaf-to-leaf communication during light stress. Sci. Signal 2018, 11, eaam9514. [Google Scholar] [CrossRef]
- Singh, R.; Parihar, P.; Singh, S.; Mishra, R.K.; Singh, V.P.; Prasad, S.M. Reactive oxygen species signaling and stomatal movement: Current updates and future perspectives. Redox Biol. 2017, 11, 213–218. [Google Scholar] [CrossRef]
- Yang, F.; Zhao, Y.; Sjöblom, J.; Li, C.; Paso, K.G. Polymeric wax inhibitors and pour point depressants for waxy crude oils: a critical review. J. Disper. Sci. Technol. 2015, 36, 213–225. [Google Scholar] [CrossRef]
- Wang, Y.; Wang, J.; Chai, G.; Li, C.; Hu, Y.; Xinhong, C.X.; Wang, Z. Developmental changes in composition and morphology of cuticular waxes on leaves and spikes of glossy and glaucous wheat (Triticum aestivum L.). PLoS ONE 2015, 10, e0141239. [Google Scholar] [CrossRef] [PubMed]
- Wicki, W.; Winzeler, M.; Schmid, J.E.; Stamp, P.; Messmer, M. Inheritance of resistance to leaf and glume blotch caused by Septoria nodorum Berk. in winter wheat. Theor. Appl. Genet. 1999, 99, 1265–1272. [Google Scholar] [CrossRef]
- Mandal, S.; Mallick, N.; Mitra, A. Salicylic acid-induced resistance to Fusarium oxysporum f. sp. lycopersici in tomato. Plant Physiol. Biochem. 2009, 47, 642–649. [Google Scholar] [CrossRef] [PubMed]
- Makandar, R.; Nalam, V.J.; Lee, H.; Trick, H.N.; Dong, Y.; Shah, J. Salicylic acid regulates basal resistance to Fusarium head blight in wheat. Mol. Plant Microbe In. 2012, 25, 431–439. [Google Scholar] [CrossRef] [PubMed]
- Beccari, G.; Covarelli, L.; Nicholson, P. Infection processes and soft wheat response to root rot and crown rot caused by Fusarium culmorum. Plant Pathol. 2011, 60, 671–684. [Google Scholar] [CrossRef]
- Pritsch, C.; Vance, C.P.; Bushnell, W.R.; Somers, D.A.; Hohn, T.M.; Muehlbauer, G.J. Systemic expression of defense response genes in wheat spikes as a response to Fusarium graminearum infection. Physiol. Mol. Plant Pathol. 2001, 58, 1–12. [Google Scholar] [CrossRef]
- Motallebi, P.; Niknam, V.; Ebrahimzadeh, H.; Hashemi, M. Sattar Tahmasebi Enferadi Exogenous Methyl Jasmonate Treatment Induces Defense Response Against Fusarium culmorum in Wheat Seedlings. J. Plant Growth Regul. 2017, 36, 71–82. [Google Scholar] [CrossRef]
- Golkari, S.; Gilbert, J.; Ban, T.; Procunier, J.D. QTL-specific microarray gene expression analysis of wheat resistance to Fusarium head blight in Sumai-3 and two susceptible NILs. Genome 2009, 52, 409–418. [Google Scholar] [CrossRef]
- Foroud, N.A.; Eudes, F. Trichothecenes in Cereal Grains. Int. J. Mol. Sci. 2009, 10, 147–173. [Google Scholar] [CrossRef]
- Sorahinobar, M.; Niknam, V.; Ebrahimzadeh, H.; Soltanloo, H. Differential antioxidative responses of susceptible and resistant wheat cultivars against Fusarium head blight. Int. J. Farm. Alli. Sci. 2015, 4, 239–243. [Google Scholar]
- Peltonen, S. Responses of barley and wheat to pathogens, non-pathogens and wounding as indicated by induced phenylalanine ammonia-lyase activity. Acta Agricul. Scandinavica B—Plant Soil Sci. 1998, 48, 184–191. [Google Scholar] [CrossRef]
- Ma, L.J.; Geiser, D.M.; Proctor, R.H.; Rooney, A.P.; O’Donnell, K.; Trail, F.; Gardiner, D.M.; Manners, J.M.; Kazan, K. Fusarium Pathogenomics. Annu. Rev. Microbiol. 2013, 67, 399–416. [Google Scholar] [CrossRef]
- Van Ginkel, M.; Gilchrist, L. How to make intelligent crosses to accumulate Fusarium head blight resistance genes based on knowledge of the underlying resistance mechanisms. In Proceedings of the National Fusarium Head Blight Forum, Erlanger, KY, USA, 7–9 December 2002; pp. 268–272. [Google Scholar]
- Zohary, D.; Hopf, M.; Weiss, E. Domestication of Plants in the Old World: The Origin and Spread of Domesticated Plants in Southwest Asia, Europe, and the Mediterranean Basin, 4th ed.; Oxford University Press: Oxford, UK, 2012. [Google Scholar]
- Gunupuru, L.R.; Perochon, A.; Doohan, F.M. Deoxynivalenol resistance as a component of FHB resistance. Trop Plant Pathol. 2017, 42, 175–183. [Google Scholar] [CrossRef]
- Miedaner, T.; Schilling, A.G.; Geiger, H.H. Molecular genetic diversity and variation for aggressiveness in populations of Fusarium graminearum and Fusarium culmorum sampled from wheat fields in different countries. J. Phytopathol. 2001, 149, 641–648. [Google Scholar] [CrossRef]
- Polish National List of Agricultural Plant Varieties. Research Center for Cultivar Testing. Słupia Wielka, Poland. 2019. Available online: http://www.coboru.pl/Publikacje_COBORU/Listy_odmian/lo_rolnicze_2019.pdf (accessed on 22 September 2019).
- Meier, U. Phenological Growth Stages. Mono- and Dicotyledonous Plants. In Phenology: An Integrative Environmental Science; Schwarz, M.D., Ed.; Springer: Dordrecht, The Netherlands, 2003; pp. 269–283. [Google Scholar]
- EPPO. European and Mediterranean Plant Protection Organization Global Database, 2018. Available online: https://gd.eppo.int (accessed on 6 August 2018).
- Camacho-Cristóbal, J.J.; Anzellotti, D.; González-Fontes, A. Changes in phenolic metabolism of tobacco plants during short-term boron deficiency. Plant Physiol. Bioch. 2002, 40, 997–1002. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Benabdellah, K.; Ruiz-Lozano, J.M.; Aroca, R. Hydrogen peroxide effects on root hydraulic properties and plasma membrane aquaporin regulation in Phaseolus vulgaris. Plant Mol. Biol. 2009, 70, 647. [Google Scholar] [CrossRef]
- Patterson, B.D.; MacRae, E.A.; Ferguson, I.B. Estimation of hydrogen peroxide in plant extracts using titanium (IV). Anal. Biochem. 1984, 139, 487–492. [Google Scholar] [CrossRef]
- Aroca, R.; Irigoyen, J.J.; Sánchez-Díaz, M. Drought enhances maize chilling tolerance. II. Photosynthetic traits and protective mechanisms against oxidative stress. Physiol. Plant. 2003, 117, 540–549. [Google Scholar] [CrossRef]
- StatSoft, Inc. STATISTICA (Data Analysis Software System), Version 12. 2014. Available online: www.statsoft.com (accessed on 6 August 2018).






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Triticum Species/Line | Origin § | Symptoms of Infection with † | Susceptibility to F. culmorum ‡ | Resistance/Susceptibility (R/S) | ||||
|---|---|---|---|---|---|---|---|---|
| B. graminis | Z. tritici | P. striiformis | P. recondita | |||||
| T. monococcum ssp. monococcum 311 | Tmm 311/S | PI 427462 | low | none | none | none | 61.1 | S |
| T. monococcum ssp. monococcum 405 | Tmm 405/R | TRI 579 | none | none | none | none | 20.0 | R |
| T. turgidum ssp. dicoccum 475 | Ttd 475/S | TRI 9574 | low | none | none | none | 21.0 | S |
| T. turgidum ssp. dicoccum 495 | Ttd 495/R | TRI 11296 | none | none | none | none | 0 | R |
| T. turgidum ssp. polonicum 618 | Ttp 618/S | TRI3550 | high | low | none | none | 21.3 | S |
| T. turgidum ssp. polonicum 406 | Ttp 406/R | TRI 1997 | none | none | low | none | 0 | R |
| T. aestivum ssp. spelta 157 | Tas 157/S | PI 168679 | high | none | none | none | 24.6 | S |
| T. aestivum ssp. spelta cv. Wirtas | Tas Wirtas/R | UWM | low | none | none | none | 0 | R |
| T. aestivum ssp. spelta 587 | Tas 587/S | TRI 9871 | high | none | none | none | 43.3 | S |
| T. aestivum ssp. spelta 581 | Tas 581/R | TRI 3665 | none | low | none | none | 0 | R |
| T. aestivum ssp. aestivum 307 | Taa 307/S | PI 387475 | high | low | none | low | 30.0 | S |
| T. aestivum ssp. aestivum cv. Sumai3 | Taa Sumai3/R | UWM | low | high | none | low | 0 | R |
| Triticum Species/Line† | Leaf Surface | Trichome Number | Stomata Number | Wax Structure † | |
|---|---|---|---|---|---|
| Tmm 311/S | AB | 91.34 b–e | 32.66 c–e | 1.00 | |
| AD | 68.00 c–f | 28.00 c–e | 1.00 | ||
| Tmm 405/R | AB | 62.00 c–g | 29.34 c–e | 1.00 | |
| AD | 67.34 c–f | 24.00 e | 1.00 | ||
| Ttd 475/S | AB | 2.68 i | 47.34 a–e | 1.00 | |
| AD | 2.00 i | 32.66 c–e | 1.00 | ||
| Ttd 495/R | AB | 78.00 c–f | 47.34 a–e | 1.00 | |
| AD | 38.30 c–f | 19.33 b–e | 1.00 | ||
| Ttp 618/S | AB | 0 i | 42.66 b–e | 2.50 | |
| AD | 0 i | 34.66 c–e | 2.50 | ||
| Ttp 406/R | AB | 109.34 a–c | 54.66 a–d | 1.00 | |
| AD | 76.00 c–f | 34.00 c–e | 1.00 | ||
| Tas 157/S | AB | 37.34 f–i | 28.66 c–e | 2.00 | |
| AD | 48.60 e–i | 32.66 c–e | 2.00 | ||
| Tas Wirtas/R | AB | 96.60 b–e | 56.00 a–c | 2.00 | |
| AD | 145.34 a | 29.34 c–e | 2.00 | ||
| Tas 587/S | AB | 2.00 i | 30.00 c–e | 2.50 | |
| AD | 0 i | 26.74 c–e | 2.50 | ||
| Tas 581/R | AB | 14.66 g–i | 75.34 a | 2.00 | |
| AD | 0 i | 42.00 b–e | 2.00 | ||
| Taa 307/S | AB | 1.34 i | 51.34 a–e | 2.50 | |
| AD | 8.00 hi | 41.34 b–e | 2.50 | ||
| Taa Sumai3/R | AB | 58.00 d–g | 26.00 de | 3.00 | |
| AD | 84.00 b–f | 26.60 de | 3.00 | ||
| Mean | S | AB | 22.45(±36.6) | 38.78(±9.6) A | 1.91 |
| AD | 21.10(±29.6) | 32.67(±5.2) B | 1.92 | ||
| R | AB | 69.77(±43.8) | 48.11(±18.4) A | 1.67 | |
| AD | 68.50(±48.6) | 29.21(±8.0) B | 1.67 | ||
| S | 21.78(±31.8) Y | 35.73(±8.0) Y | 1.92 | ||
| R | 69.14(±39.8) X | 38.66(±16.7) X | 1.67 | ||
| Triticum Species/Line † | Organization and Characteristics of Stomata | Trichome Characteristics |
|---|---|---|
| Tmm 311/S | linear and parallel | single, very long trichomes |
| Tmm 405/R | linear and parallel, semi-closed stomata | long trichomes |
| Ttd 475/S | linear and parallel, closely arranged rows of stomata | trichomes practically absent |
| Ttd 495/R | linear and parallel | curved, medium-long trichomes |
| Ttp 618/S | linear and parallel, closely arranged rows of stomata | no trichomes |
| Ttp 406/R | linear and parallel, large gaps between stomata in rows, semi-closed stomata | numerous, very short trichomes |
| Tas 157/S | linear and parallel, closely arranged rows of stomata | long trichomes in parallel lines and short trichomes covering the entire leaf surface |
| Tas Wirtas/R | linear and parallel, rows of paired stomata, semi-closed stomata | very short trichomes |
| Tas 587/S | linear and parallel, closely arranged rows of stomata | single, very short trichomes |
| Tas 581/R | linear and parallel, rows of densely packed stomata, closely arranged rows of stomata, semi-closed stomata | single, very short trichomes |
| Taa 307/S | linear and parallel, large number of open stomata | lines of short trichomes |
| Taa Sumai3/R | linear and parallel, semi-closed stomata | single, short trichomes |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Duba, A.; Goriewa-Duba, K.; Wachowska, U.; Głowacka, K.; Wiwart, M. The Associations between Leaf Morphology, Phenylalanine Ammonia Lyase Activity, Reactive Oxygen Species, and Fusarium Resistance in Selected Species of Wheat with Different Ploidy Levels. Plants 2019, 8, 360. https://doi.org/10.3390/plants8100360
Duba A, Goriewa-Duba K, Wachowska U, Głowacka K, Wiwart M. The Associations between Leaf Morphology, Phenylalanine Ammonia Lyase Activity, Reactive Oxygen Species, and Fusarium Resistance in Selected Species of Wheat with Different Ploidy Levels. Plants. 2019; 8(10):360. https://doi.org/10.3390/plants8100360
Chicago/Turabian StyleDuba, Adrian, Klaudia Goriewa-Duba, Urszula Wachowska, Katarzyna Głowacka, and Marian Wiwart. 2019. "The Associations between Leaf Morphology, Phenylalanine Ammonia Lyase Activity, Reactive Oxygen Species, and Fusarium Resistance in Selected Species of Wheat with Different Ploidy Levels" Plants 8, no. 10: 360. https://doi.org/10.3390/plants8100360
APA StyleDuba, A., Goriewa-Duba, K., Wachowska, U., Głowacka, K., & Wiwart, M. (2019). The Associations between Leaf Morphology, Phenylalanine Ammonia Lyase Activity, Reactive Oxygen Species, and Fusarium Resistance in Selected Species of Wheat with Different Ploidy Levels. Plants, 8(10), 360. https://doi.org/10.3390/plants8100360