
Evaluating Douglas Fir’s Provenances in Romania Through Multi-Trait Selection

 , , and
, , and 
Abstract
1. Introduction
2. Materials and Methods
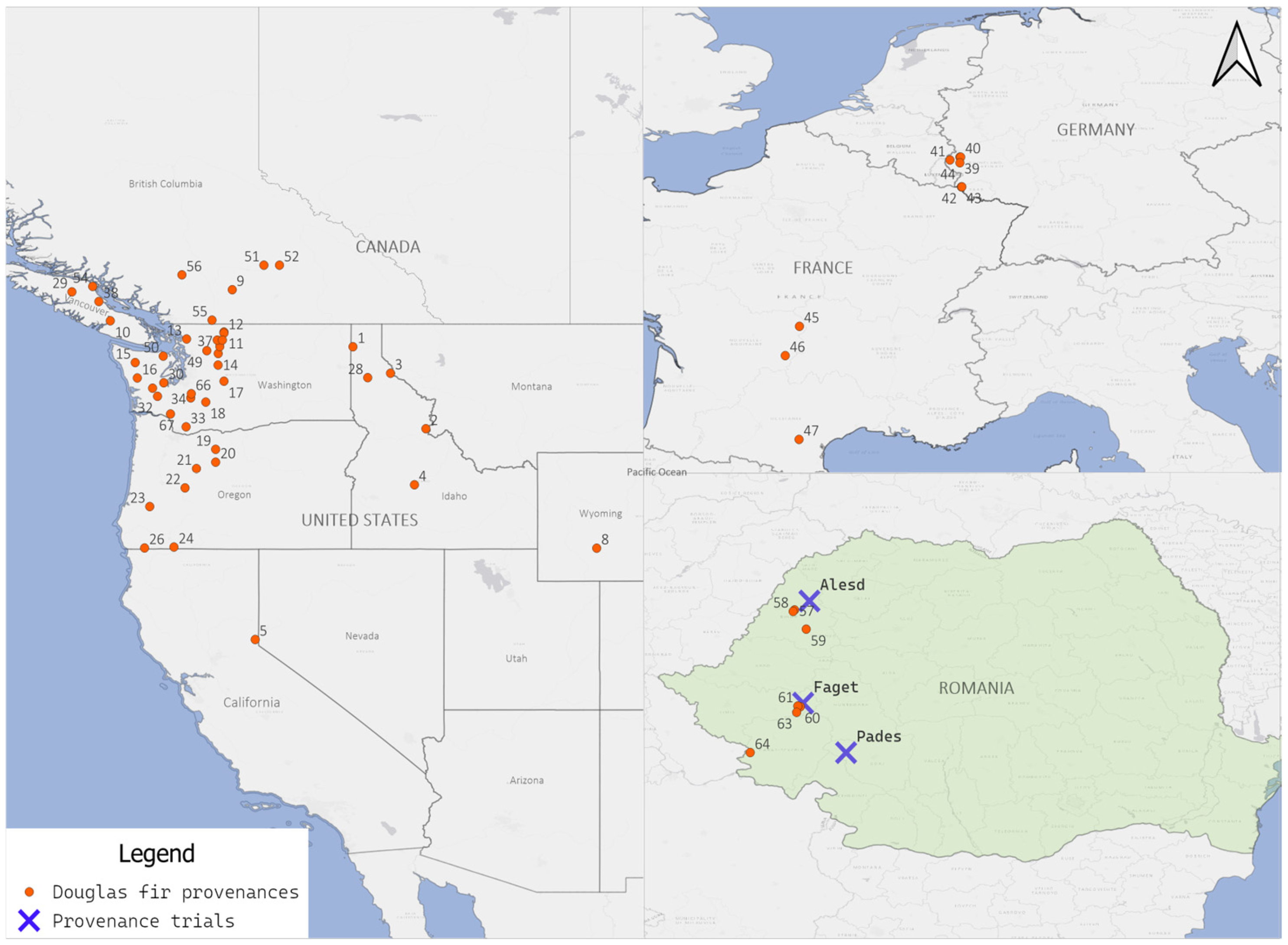
2.1. Experimental Design
2.2. Statistical Analysis
- (1)
- Rescaling traits: values were scaled to 0–100, with higher scores indicating desired increases or decreases depending on the trait.
- (2)
- Factor analysis: traits were grouped into factors based on correlations between them, and scores were assigned for each provenance.
- (3)
- Defining the ideotype: an ideal genotype was set with a value of 100 for all traits
- (4)
- Calculating distances: the distance between each genotype and the ideotype was computed to derive the MGIDI [32].
- (1)
- Rescaling values: both the WAASB (Weighted Average of Absolute Scores, measuring genotype stability across environments using linear mixed-effect models) and trait performance values were scaled to a uniform range of 0 to 100 (0 represents the minimum and 100 represents the maximum).
- (2)
- Calculating the WAASBY index: the WAASBY index was derived to balance stability (WAASB) and mean performance (Y), with the flexibility to adjust weights to emphasize either stability or performance [33].
- (3)
- Defining the ideotype: similar to the MGIDI, but in this case, the ideotype was assigned a value of 100 for the WAASBY index to represent the ideal combination of stability and performance.
- (4)
- MTSI estimation: the MTSI value was then calculated using the following formula:
3. Results
3.1. Genetic Variability Analysis for Trial Sites
3.2. Genotype by Environment Analysis
3.3. Multi-Trait Selection in Each Provenance Trial
3.4. The Multi-Trait Selection Across Common Garden Experiments
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Pâques, L.E. Forest Tree Breeding in Europe: Current State-of-the-Art and Perspectives; Springer Science & Business Media: Dordrecht, The Netherlands, 2013; ISBN 978-94-007-6146-9. [Google Scholar]
- Ronch, F.D.; Caudullo, G.; de Rigo, D. Pseudotsuga menziesii in Europe: Distribution, Habitat, Usage and Threats. In European Atlas of Forest Tree Species; Publication Office of the European Union: Luxembourg, 2016. [Google Scholar]
- Lavender, D.P.; Hermann, R.K. Douglas-Fir: The Genus Pseudotsuga; Oregon State University: Corvallis, OR, USA, 2014; ISBN 978-0-615-97995-3. [Google Scholar]
- European Forest Institute. Douglas fir—An Option for Europe; Leskinen, P., Lindner, M., European Forest Institute, Verkerk, P.J., Nabuurs, G.-J., Van Brusselen, J., Kulikova, E., Hassegawa, M., Lerink, B., Eds.; What Science Can Tell Us; European Forest Institute: Joensuu, Finland, 2020; Volume 11, ISBN 978-952-7426-00-5. [Google Scholar]
- Popa-Costea, V. Research on the Behaviour of Some Commercial Provenances of Green Douglas-fir Under the Conditions of Our Country; Studii și Cercetări, Seria I; ICAS: București, Romania, 1973; Volume XXIX, pp. 249–305. [Google Scholar]
- Enescu, V. Ameliorarea Principalelor Specii Forestiere. Breeding of the Main Forest Tree Species; CERES: București, Romania, 1975; pp. 11–72. [Google Scholar]
- Şofletea, N.; Curtu, L. Dendrologie; Editura Universităţii: Transilvania, Romania, 2007; ISBN 973-635-885-2. [Google Scholar]
- Konnert, M. Pseudotsuga menziesii. Stud. Slov. 2016, 151, 30–35. [Google Scholar]
- Hermann, R.K.; Lavender, D.P. Planted Forests: Contributions to the Quest for Sustainable Societies; Kluwer Academic Publishers: Dordrecht, The Netherlands, 1999. [Google Scholar]
- Aitken, S.N. Genetics and Genetic Resources|Genecology and Adaptation of Forest Trees. In Encyclopedia of Forest Sciences; Burley, J., Ed.; Elsevier: Oxford, UK, 2004; pp. 197–204. ISBN 978-0-12-145160-8. [Google Scholar]
- Schober, R. Douglasien Provenienzversuche [Douglas-fir Provenance Trials]. Allg. Forst-Und Jagdzeitung 1954, 125, 60–179. [Google Scholar]
- Stimm, B.; Dong, P.H. The Kaiserslautern Douglas Fir Provenance Trial After Nine Decades of Observation. Eur. J. For. Res. 2001, 120, 173–186. [Google Scholar] [CrossRef]
- Enescu, V. Cercetări de Provenineță la Duglas si Larice; Institutul de Cercetări și Amenajări Silvice: Voluntari, Romania, 1984. [Google Scholar]
- Maria, M. Studiul Variabilităţii Genetice Interpopulaţionale la Larice (Larix decidua Mill.) Şi Duglas (Psudotsuga menziesii Mirb. Franco) în Culturi Comparative Multistaţionale din România; Academia de Științe Agricole și Silvice: București, Romania, 1998. [Google Scholar]
- Callaham, R.Z. Provenance Research: Investigation of Genetic Diversity Associated with Geography. Unasylua 1963, 18, 73–74. [Google Scholar]
- White, T.L.; Adams, W.T.; Neale, D.B.; Adams, W.T. Forest Genetics; Nachdr. CABI Publishing: Wallingford, UK, 2009; ISBN 978-0-85199-083-5. [Google Scholar]
- Apostol, E.N.; Stuparu, E.; Scarlatescu, V.; Budeanu, M. Testing Hungarian oak (Quercus frainetto Ten.) Provenances in Romania. iForest—Biogeosciences For. 2020, 13, 9. [Google Scholar] [CrossRef]
- Blada, I.; Popescu, F. Swiss Stone Pine Provenance Experiment in Romania: II Variation in Growth and Branching Traits to Age 14. Silvae Genet. 2017, 56, 148–158. [Google Scholar] [CrossRef]
- Krakowski, J.; Stoehr, M.U. Coastal Douglas-Fir Provenance Variation: Patterns and Predictions for British Columbia Seed Transfer. Ann. For. Sci. 2009, 66, 811. [Google Scholar] [CrossRef]
- Li, Y.; Suontama, M.; Burdon, R.D.; Dungey, H.S. Genotype by Environment Interactions in Forest Tree Breeding: Review of Methodology and Perspectives on Research and Application. Tree Genet. Genomes 2017, 13, 60. [Google Scholar] [CrossRef]
- Klisz, M.; Chakraborty, D.; Cvjetković, B.; Grabner, M.; Lintunen, A.; Mayer, K.; George, J.-P.; Rossi, S. Functional Traits of Boreal Species and Adaptation to Local Conditions. In Boreal Forests in the Face of Climate Change: Sustainable Management; Girona, M.M., Morin, H., Gauthier, S., Bergeron, Y., Eds.; Springer International Publishing: Cham, Switzerland, 2023; pp. 323–355. ISBN 978-3-031-15988-6. [Google Scholar]
- Alberto, F.J.; Aitken, S.N.; Alía, R.; González-Martínez, S.C.; Hänninen, H.; Kremer, A.; Lefèvre, F.; Lenormand, T.; Yeaman, S.; Whetten, R.; et al. Potential for Evolutionary Responses to Climate Change—Evidence from Tree Populations. Glob. Chang. Biol. 2013, 19, 1645–1661. [Google Scholar] [CrossRef]
- Liepe, K.J.; van der Maaten, E.; van der Maaten-Theunissen, M.; Kormann, J.M.; Wolf, H.; Liesebach, M. Ecotypic Variation in Multiple Traits of European Beech: Selection of Suitable Provenances Based on Performance and Stability. Eur. J. For. Res. 2024, 143, 831–845. [Google Scholar] [CrossRef]
- Pégard, M.; Segura, V.; Muñoz, F.; Bastien, C.; Jorge, V.; Sanchez, L. Favorable Conditions for Genomic Evaluation to Outperform Classical Pedigree Evaluation Highlighted by a Proof-of-Concept Study in Poplar. Front. Plant Sci. 2020, 11, 581954. [Google Scholar] [CrossRef] [PubMed]
- Tahina, R.; Ramamonjisoa, L.; Verhaegen, D.; Tsy, J.-M.; Jacquin, L.; Cao, T.-V.; Garel, M.; Bouvet, J.-M. Performance of Multi-Trait Genomic Selection for Eucalyptus Robusta Breeding Program. Tree Genet. Genomes 2018, 14, 71. [Google Scholar] [CrossRef]
- Tsuda, Y.; Semerikov, V.; Sebastiani, F.; Giovanni Giuseppe, V.; Lascoux, M. Multispecies Genetic Structure and Hybridization in the Betula Genus across Eurasia. Mol. Ecol. 2016, 26, 589–605. [Google Scholar] [CrossRef]
- Packham, J.R.; Thomas, P.A.; Atkinson, M.D.; Degen, T. Biological Flora of the British Isles: Fagus sylvatica. J. Ecol. 2012, 100, 1557–1608. [Google Scholar] [CrossRef]
- Matsushita, M.; Nishikawa, H.; Tamura, A. Effects of Girdling Intensity, Pruning Season and Thinning on Tree Growth, Crown Vigor and Wound Recovery in Japanese Larch. Forests 2022, 13, 449. [Google Scholar] [CrossRef]
- Alexandru, A.-M.; Mihai, G.; Stoica, E.; Curtu, A.L. Multi-Trait Selection and Stability in Norway Spruce (Picea abies) Provenance Trials in Romania. Forests 2023, 14, 456. [Google Scholar] [CrossRef]
- Wang, T.; Ren, J.; Huang, Q.; Li, J. Genetic Parameters of Growth and Leaf Traits and Genetic Gains with MGIDI in Three Populus Simonii × P. Nigra Families at Two Spacings. Front. Plant Sci. 2024, 15, 1483580. [Google Scholar] [CrossRef]
- Ding, X.; Zhang, Y.; Sun, J.; Tan, Z.; Huang, Q.; Diao, S.; Wu, Y.; Luan, Q.; Jiang, J. Genetic Selection for Growth, Wood Quality and Resin Traits of Potential Slash Pine for Multiple Industrial Uses. For. Res. 2024, 4, e023. [Google Scholar] [CrossRef]
- Olivoto, T.; Nardino, M. MGIDI: Toward an Effective Multivariate Selection in Biological Experiments. Bioinformatics 2021, 37, 1383–1389. [Google Scholar] [CrossRef]
- Olivoto, T.; Lúcio, A.D.C.; da Silva, J.A.G.; Sari, B.G.; Diel, M.I. Mean Performance and Stability in Multi-Environment Trials II: Selection Based on Multiple Traits. Agron. J. 2019, 111, 2961–2969. [Google Scholar] [CrossRef]
- Bucci, G. ClimateDT—Climate Downscaling Tool. Available online: https://www.ibbr.cnr.it/climate-dt/ (accessed on 10 February 2025).
- Yin, M.; Wang, C.; Wang, H.; Han, Q.; Zhao, Z.; Tang, C.; Guo, J.; Zeng, J. Modeling and Evaluating Site and Provenance Variation in Height–Diameter Relationships for Betula Alnoides Buch.–Ham. Ex D. Don in Southern China. Front. Plant Sci. 2023, 14, 1248278. [Google Scholar] [CrossRef] [PubMed]
- Víquez, E.; Pérez, D. Effect of Pruning on Tree Growth, Yield, and Wood Properties of Tectona Grandis Plantations in Costa Rica. Silva Fenn. 2005, 39, 375. [Google Scholar] [CrossRef]
- Mihai, G.; Alexandru, A.; Nita, I.-A.; Birsan, M.-V. Climate Change in the Provenance Regions of Romania over the Last 70 Years: Implications for Forest Management. Forests 2022, 13, 1203. [Google Scholar] [CrossRef]
- Yang, Z.; Liu, Q.; Luo, P.; Ye, Q.; Duan, G.; Sharma, R.; Zhang, H.; Wang, G.; Fu, L. Prediction of Individual Tree Diameter and Height to Crown Base Using Nonlinear Simultaneous Regression and Airborne LiDAR Data. Remote Sens. 2020, 12, 2238. [Google Scholar] [CrossRef]
- Nanson, A. Génétique et Amélioration des Arbres Forestiers; Presses Agronomiques de Gembloux: Gembloux, Belgium, 2004; ISBN 978-2-87016-070-1. [Google Scholar]
- Kuznetsova, A.; Brockhoff, P.B.; Christensen, R.H.B.; Jensen, S.P. lmerTest: Tests in Linear Mixed Effects Models. 23 October 2020. Available online: https://CRAN.R-project.org/package=lmerTest (accessed on 10 September 2024).
- Olivoto, T.; Lúcio, A.D. Metan: An R Package for Multi-Environment Trial Analysis. Methods Ecol. Evol. 2020, 11, 783–789. [Google Scholar] [CrossRef]
- Ye, T.Z.; Jayawickrama, K.J.S. Predicted Responses of Genetically Improved Populations to Climate Changes Based on Second-Cycle Douglas-Fir Progeny Tests. Forests 2024, 15, 1610. [Google Scholar] [CrossRef]
- Gričar, J.; Arnič, D.; Krajnc, L.; Prislan, P.; Božič, G.; Westergren, M.; Mátyás, C.; Kraigher, H. Different Patterns of Inter-Annual Variability in Mean Vessel Area and Tree-Ring Widths of Beech from Provenance Trials in Slovenia and Hungary. Trees 2024, 38, 179–195. [Google Scholar] [CrossRef]
- Liao, J.; Hang, J.; Luo, Q.; Luo, H.; Ma, T.; Wei, Z.; Yuan, H. Seasonal Variability of Forest Cooling and Warming Effects and Response to Drought in Mid-to-High Latitudes of the Northern Hemisphere. For. Ecol. Manag. 2023, 546, 121324. [Google Scholar] [CrossRef]
- Chakraborty, D.; Ciceu, A.; Ballian, D.; Benito Garzón, M.; Bolte, A.; Bozic, G.; Buchacher, R.; Čepl, J.; Cremer, E.; Ducousso, A.; et al. Assisted Tree Migration Can Preserve the European Forest Carbon Sink Under Climate Change. Nat. Clim. Chang. 2024, 14, 845–852. [Google Scholar] [CrossRef]
- Benito Garzón, M.; Robson, T.M.; Hampe, A. ΔTraitSDMs: Species Distribution Models That Account for Local Adaptation and Phenotypic Plasticity. New Phytol. 2019, 222, 1757–1765. [Google Scholar] [CrossRef]
- Isaac-Renton, M.; Moore, B.; Degner, J.; Bealle Statland, C.; Bogdanski, B.; Sun, L.; Stoehr, M. Economic Gain of Genetically-Selected Coastal Douglas-Fir: Timber, Log and Carbon Value at Varying Planting Densities. For. Policy Econ. 2025, 171, 103397. [Google Scholar] [CrossRef]
- Peláez, P.; Lorenzana, G.P.; Baesen, K.; Montes, J.R.; De La Torre, A.R. Spatially Heterogeneous Selection and Inter-Varietal Differentiation Maintain Population Structure and Local Adaptation in a Widespread Conifer. BMC Ecol. Evol. 2024, 24, 117. [Google Scholar] [CrossRef] [PubMed]
- Nicolescu, V.-N.; Mason, W.L.; Bastien, J.-C.; Vor, T.; Petkova, K.; Podrázský, V.; Đodan, M.; Perić, S.; La Porta, N.; Brus, R.; et al. Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) in Europe: An Overview of Management Practices. J. For. Res. 2023, 34, 871–888. [Google Scholar] [CrossRef]
- Gartner, B.; Robbins, J.; Newton, M. Effects of Pruning on Wood Density and Tracheid Length in Young Douglas-Fir. Wood Fiber Sci. 2003, 37, 304–313. [Google Scholar]
- Smolnikar, P.; Brus, R.; Jarni, K. Differences in Growth and Log Quality of Douglas-Fir (Pseudotsuga menziesii (Mirb.) Franco) Provenances. Forests 2021, 12, 287. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Aleșd | Făget | Padeș | |
|---|---|---|---|
| Coordinates | 47.15° N, 22.33° E | 45.76° N, 22.29° E | 45.09° N, 22.89° E |
| Temperature (°C) (1997–2023) | 7.9 (15.3) | 10.4 (17.9) | 7.82 (15.4) |
| Precipitation (mm) (1997–2023) | 71.6 (90.0) | 70.9 (90.9) | 81.25 (100.5) |
| Elevation (m a.s.l.) | 670 | 330 | 720 |
| Soil | Eutricambosoil | Luvosol | Eutricambosoil |
| Trial | Trait | LRTp | Vp | Vr | MS b | Mean ± SD |
|---|---|---|---|---|---|---|
| Aleșd | DBH (cm) | 23.16 *** | 7.23 | 60.77 | 80.66 ns | 36.05 ± 8.18 |
| TH (m) | 29.67 *** | 3.36 | 19.68 | 13.32 ns | 30.36 ± 4.71 | |
| PH (m) | 24.55 *** | 0.98 | 7.34 | 228.08 *** | 17.48 ± 2.91 | |
| Făget | DBH (cm) | 8.27 ** | 2.87 | 56.16 | 565.59 *** | 28.82 ± 7.75 |
| TH (m) | 12.80 *** | 2.009 | 24.73 | 923.91 *** | 29.61 ± 5.31 | |
| PH (m) | 34.01 *** | 1.73 | 12.05 | 735.91 *** | 17.32 ± 3.86 | |
| Padeș | DBH (cm) | 2.89 ns | 1.93 | 50.89 | 0.31 ns | 34.71 ± 7.25 |
| TH (m) | 18.97 *** | 1.59 | 15.29 | 20.03 ns | 28.77 ± 4.1 | |
| PH (m) | 7.78 ** | 0.38 | 6.74 | 20.02 ns | 17.68 ± 2.67 |
| Traits | LRTg | LRTge | Vp | Vge | Vr | MS Env | MS B (Env) | Mean ± SD |
|---|---|---|---|---|---|---|---|---|
| DBH (cm) | 0 ns | 20.74 *** | 3.53 (27%) | 3.91 (30%) | 5.69 (43%) | 1196 *** | 70.96 ns | 33.03 ± 8.43 |
| TH (m) | 0 ns | 28.03 *** | 0 (0%) | 1.75 (8%) | 20.36 (92%) | 221.8 *** | 162.9 *** | 29.50 ± 4.79 |
| PH (m) | 0.03 ns | 38.20 *** | 0.03 (1%) | 0.92 (9%) | 8.97 (90%) | 57.88 ** | 161.7 *** | 17.47 ± 3.24 |
| Traits | Site Mean | Selected Provenances Mean | Selection Differential % | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Aleșd | Făget | Padeș | Aleșd | Făget | Padeș | Aleșd | Făget | Padeș | |
| DBH (cm) | 35.9 | 28.8 | 34.7 | 39.8 | 30.3 | 35.8 | 10.9 | 5.2 | 3.2 |
| TH (m) | 30.2 | 29.6 | 28.8 | 31.9 | 30.9 | 30 | 5.6 | 4.4 | 4.2 |
| PH (m) | 17.3 | 17.3 | 17.7 | 17.8 | 18.5 | 18.2 | 2.9 | 6.9 | 2.8 |
| Traits | Site Mean | Selected Provenances Mean | Selection Differential % | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Aleșd | Făget | Padeș | Aleșd | Făget | Padeș | Aleșd | Făget | Padeș | |
| DBH (cm) | 35.9 | 28.8 | 34.7 | 37.1 | 29.62 | 36.75 | 3.3 | 2.8 | 5.9 |
| TH (m) | 30.2 | 29.6 | 28.8 | 30.92 | 30.22 | 29.5 | 2.4 | 2.1 | 2.4 |
| PH (m) | 17.3 | 17.3 | 17.7 | 17.82 | 17.62 | 17.8 | 3.0 | 1.8 | 0.6 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stoica, E.; Alexandru, A.M.; Mihai, G.; Scarlatescu, V.; Curtu, A.L. Evaluating Douglas Fir’s Provenances in Romania Through Multi-Trait Selection. Plants 2025, 14, 1347. https://doi.org/10.3390/plants14091347
Stoica E, Alexandru AM, Mihai G, Scarlatescu V, Curtu AL. Evaluating Douglas Fir’s Provenances in Romania Through Multi-Trait Selection. Plants. 2025; 14(9):1347. https://doi.org/10.3390/plants14091347
Chicago/Turabian StyleStoica, Emanuel, Alin Madalin Alexandru, Georgeta Mihai, Virgil Scarlatescu, and Alexandru Lucian Curtu. 2025. "Evaluating Douglas Fir’s Provenances in Romania Through Multi-Trait Selection" Plants 14, no. 9: 1347. https://doi.org/10.3390/plants14091347
APA StyleStoica, E., Alexandru, A. M., Mihai, G., Scarlatescu, V., & Curtu, A. L. (2025). Evaluating Douglas Fir’s Provenances in Romania Through Multi-Trait Selection. Plants, 14(9), 1347. https://doi.org/10.3390/plants14091347