
Agronomic Traits, Nutrient Accumulation, and Their Correlations in Wheat, as Affected by Nitrogen Supply in Rainfed Coastal Saline Soils


 and
and 
Abstract
1. Introduction
2. Results
2.1. Agronomic Performance of Wheat Genotypes in Response to N Supply
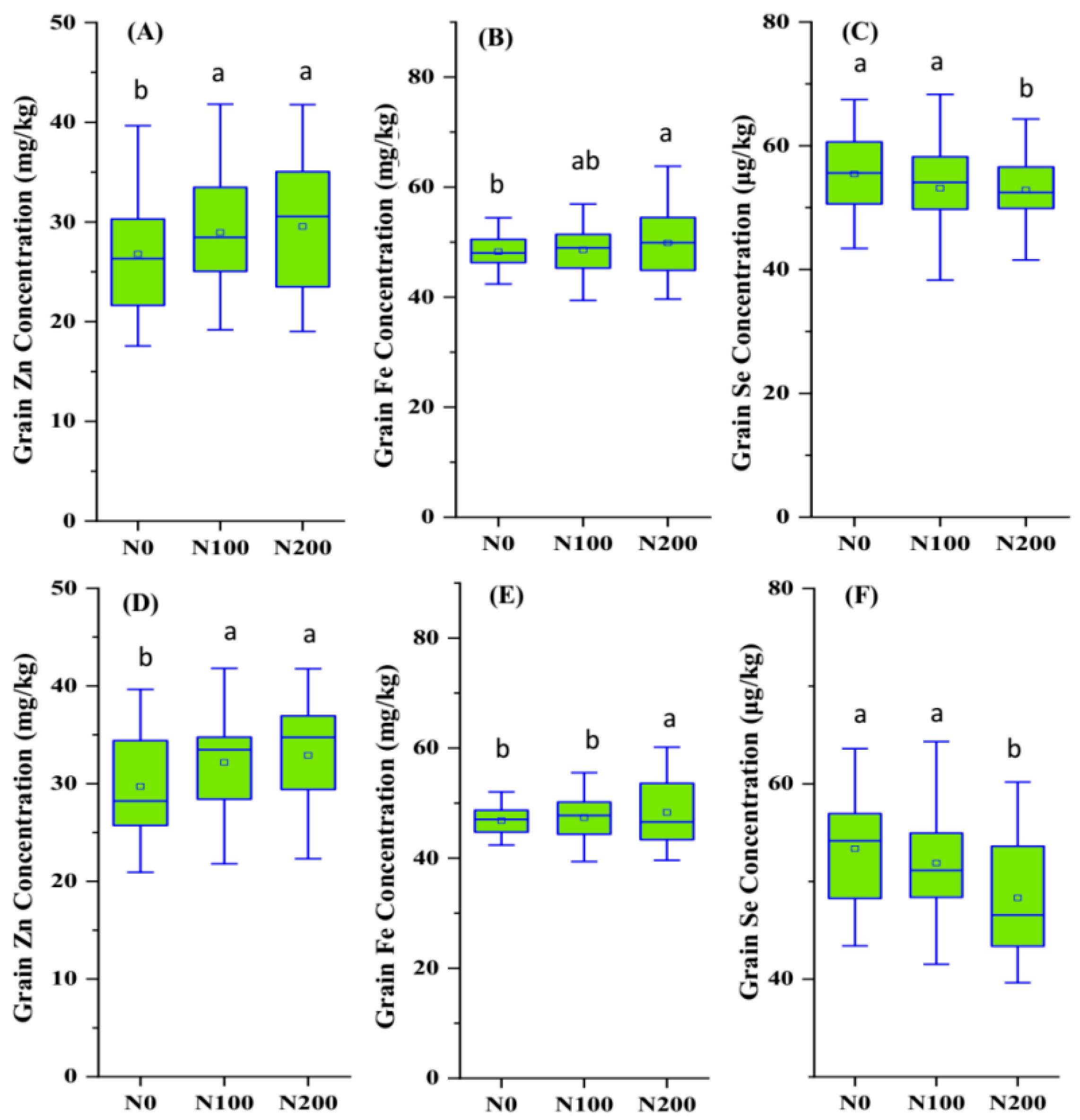
2.2. Nutrient Accumulation
2.3. Nitrogen Agronomic Efficiency of Eight Wheat Genotypes
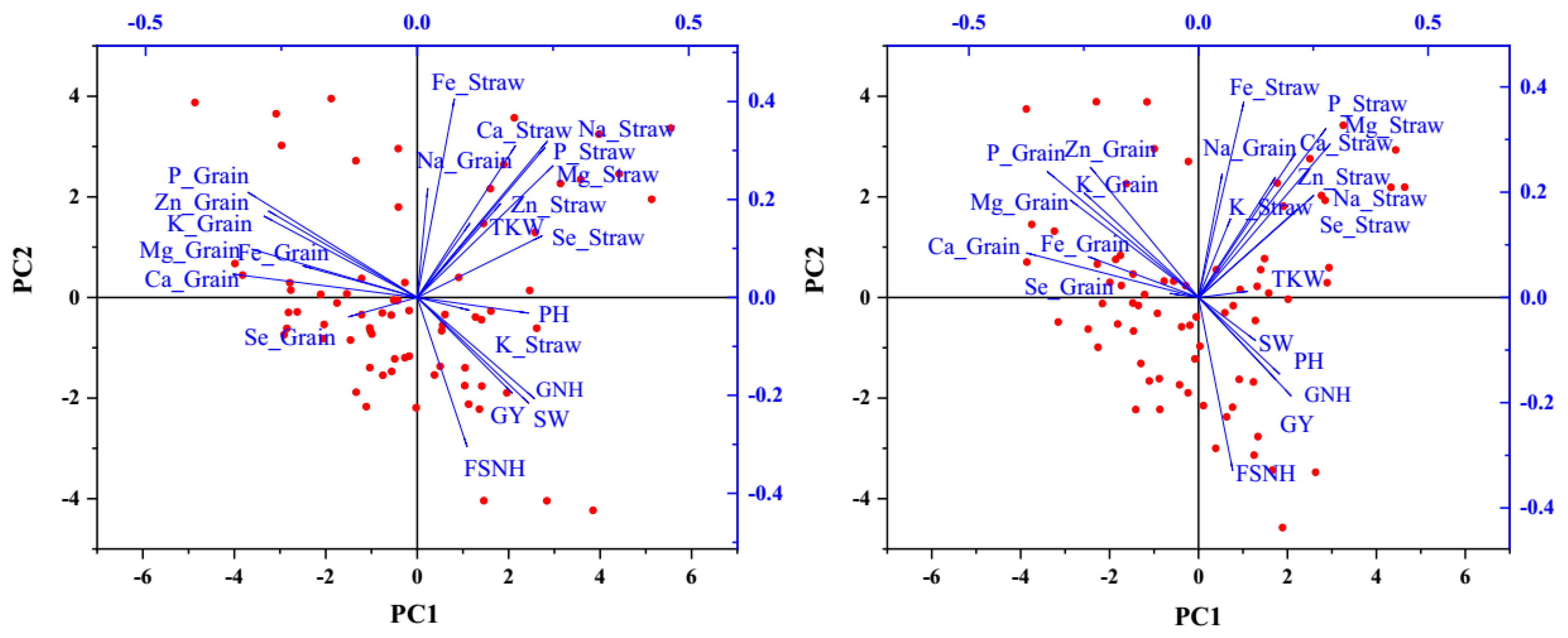
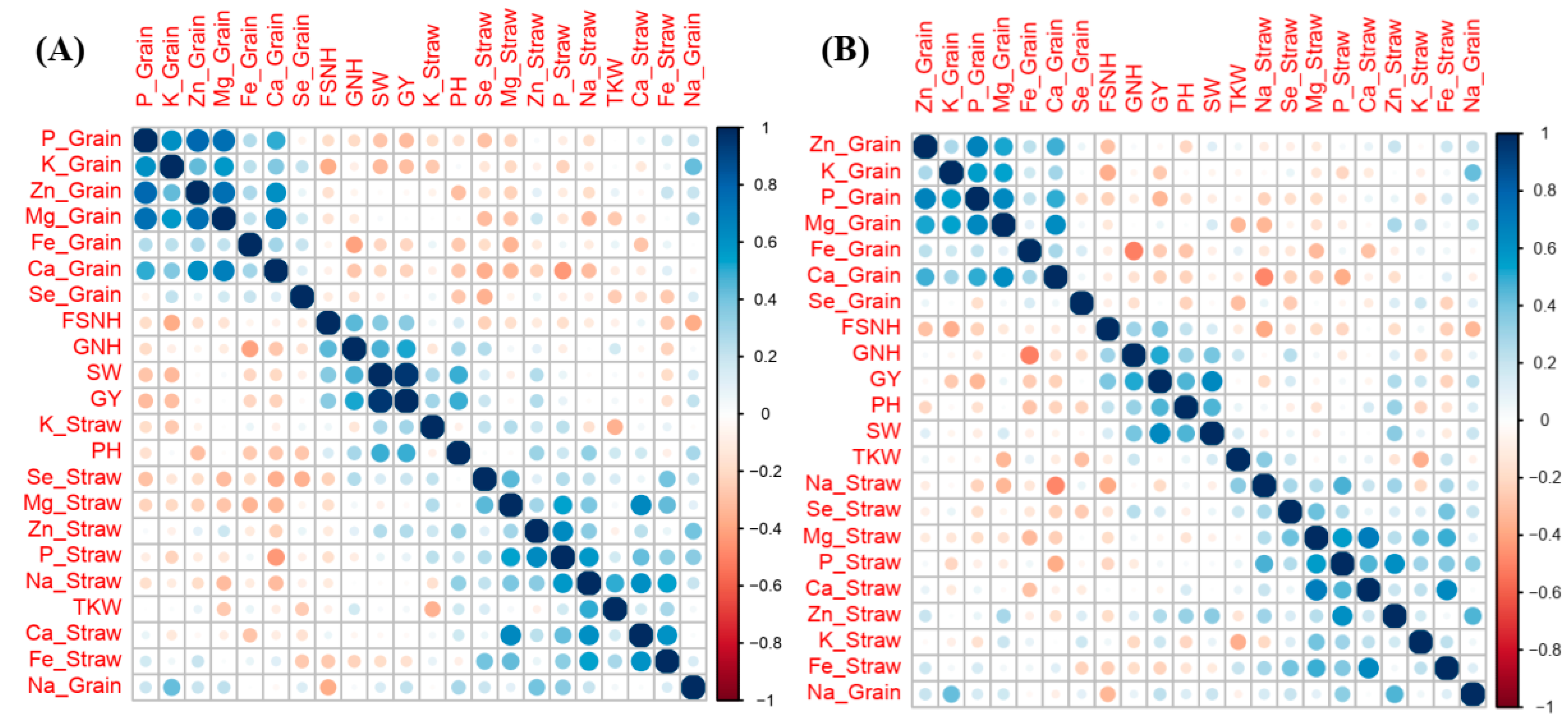
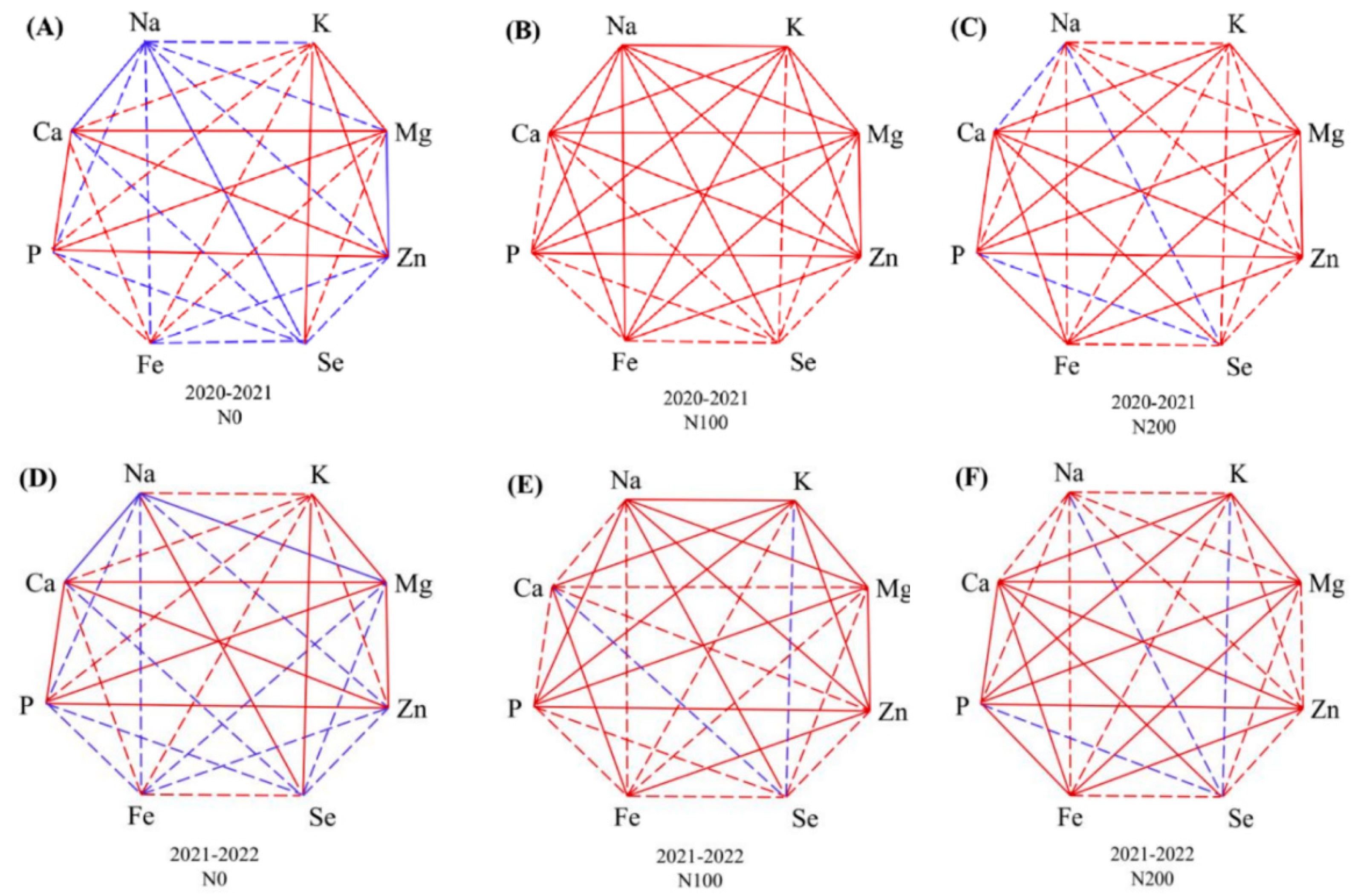
2.4. Associations Between Agronomic Performance and Nutrient Accumulation
3. Discussion
3.1. Seasonal Variation of Agronomic Performance and Nutrient Utilization Under Coastal Soil Conditions
3.2. Correlations Between Agronomic Traits and Nutrient Uptake Were Modified by N Supply
4. Materials and Methods
4.1. Experimental Setup
4.2. Agronomic Performance
4.3. Elemental Measurements
4.4. Nutrient Accumulation Parameters
4.5. Nitrogen Agronomic Efficiency
4.6. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Negacz, K.; Malek, Z.; De Vos, A.; Vellinga, P. Saline soils worldwide: Identifying the most promising areas for saline agriculture. J. Arid Environ. 2022, 203, 104775. [Google Scholar]
- Alfarrah, N.; Walraevens, K. Groundwater overexploitation and seawater intrusion in coastal areas of arid and semi-arid regions. Water 2018, 10, 143. [Google Scholar] [CrossRef]
- Prasad, A.S. Discovery of human zinc deficiency: Its impact on human health and disease. Adv. Nutr. 2013, 4, 176–190. [Google Scholar]
- Piskin, E.; Cianciosi, D.; Gulec, S.; Tomas, M.; Capanoglu, E. Iron absorption: Factors, limitations, and improvement methods. ACS Omega 2022, 7, 20441–20456. [Google Scholar] [PubMed]
- Danso, O.P.; Asante-Badu, B.; Zhang, Z.; Song, J.; Wang, Z.; Yin, X.; Zhu, R. Selenium Biofortification: Strategies, Progress and Challenges. Agriculture 2023, 13, 416. [Google Scholar] [CrossRef]
- Bouis, H.E.; Welch, R.M. Biofortification—A sustainable agricultural strategy for reducing micronutrient malnutrition in the global South. Crop Sci. 2010, 50, 20–32. [Google Scholar] [CrossRef]
- White, P.J.; Broadley, M.R. Biofortification of crops with seven mineral elements often lacking in human diets—Fe, Zn, Cu, calcium, magnesium, selenium and iodine. New Phytol. 2008, 182, 49–84. [Google Scholar]
- Cakmak, I.; Kutman, U.B. Agronomic biofortification of cereals with zinc: A review. Eur. J. Soil Sci. 2018, 69, 172–180. [Google Scholar]
- Hawkesford, M.J.; Zhao, F.J. Strategies for increasing the selenium content of wheat. J. Cereal Sci. 2007, 46, 282–292. [Google Scholar]
- Velu, G.; Ortiz-Monasterio, I.; Cakmak, I.; Hao, Y.; Singh, R.Á. Biofortification strategies to increase grain zinc and iron concentrations in wheat. J. Cereal Sci. 2014, 59, 365–372. [Google Scholar]
- Zou, C.Q.; Zhang, Y.Q.; Rashid, A.; Ram, H.; Savasli, E.; Arisoy, R.Z.; Ortiz-Monasterio, I.; Simunji, S.; Wang, Z.H.; Sohu, V.; et al. Biofortification of wheat with zinc through zinc fertilization in seven countries. Plant Soil 2012, 361, 119–130. [Google Scholar] [CrossRef]
- Ramkissoon, C.; Degryse, F.; da Silva, R.C.; Baird, R.; Young, S.D.; Bailey, E.H.; McLaughlin, M.J. Improving the efcacy of selenium fertilizers for wheat biofortifcation. Sci. Rep. 2019, 9, 19520. [Google Scholar] [CrossRef]
- Fageria, N.K.; Baligar, V.C.; Jones, C.A. Growth and Mineral Nutrition of Field Crops, 3rd ed.; CRC Press: Boca Raton, FL, USA, 2010. [Google Scholar]
- Nie, Z.; Zhao, P.; Shi, H.; Wang, Y.; Qin, S.; Liu, H. Nitrogen supply enhances zinc uptake and root-to-shoot translocation via up-regulating the expression of TaZIP3 and TaZIP 7 in winter wheat (Triticum aestivum). Plant Soil 2019, 444, 501–517. [Google Scholar] [CrossRef]
- Recena, R.; García-López, A.M.; Delgado, A. Zinc uptake by plants as affected by fertilization with Zn sulfate, phosphorus availability, and soil properties. Agronomy 2021, 11, 390. [Google Scholar] [CrossRef]
- Kutman, U.B.; Yildiz, B.; Cakmak, I. Effect of nitrogen on uptake, remobilization and partitioning of zinc and iron throughout the development of durum wheat. Plant Soil 2011, 342, 149–164. [Google Scholar] [CrossRef]
- Kutman, U.B.; Yildiz, B.; Cakmak, I. Improved nitrogen status enhances zinc and iron concentrations both in the whole grain and the endosperm fraction of wheat. J. Cereal Sci. 2011, 53, 118–125. [Google Scholar] [CrossRef]
- Fan, Z.J.; Mi, D.M.; Huo, R.W.; Kong, W.L.; Noor, H.; Ren, A.X.; Ding, P.C.; Li, L.H.; Ren, Y.K.; Sun, M.; et al. Effects of Nitrogen application amount on soil selenium speciations, physiological characteristics and yield of triticales in selenium-enriched area. Russ. J. Plant Physiol. 2023, 70, 207. [Google Scholar]
- Zhang, H.; Du, B.; Jiang, S.; Zhu, J.; Wu, Q. Potential Assessment of Selenium for Improving Nitrogen Metabolism, Yield and Nitrogen Use Efficiency in Wheat. Agronomy 2023, 13, 110. [Google Scholar] [CrossRef]
- Guo, X.; Ma, X.; Zhang, J.; Zhu, J.; Lu, T.; Wang, Q.; Wang, X.; Hua, W.; Xu, S. Meta-analysis of the role of zinc in coordinating absorption of mineral elements in wheat seedlings. Plant Methods 2021, 17, 105. [Google Scholar]
- Zhang, W.; Liu, D.; Liu, Y.; Chen, X.; Zou, C. Overuse of phosphorus fertilizer reduces the grain and flour protein contents and zinc bioavailability of winter wheat (Triticum aestivum L.). J. Agric. Food Chem. 2017, 65, 1473–1482. [Google Scholar] [CrossRef]
- Huang, T.; Huang, Q.; She, X.; Ma, X.; Huang, M.; Cao, H.; He, G.; Liu, J.; Liang, D.; Malhi, S.S.; et al. Grain zinc concentration and its relation to soil nutrient availability in different wheat cropping regions of China. Soil Till. Res. 2019, 191, 57–65. [Google Scholar]
- Agarwala, S.C.; Mehrotra, S.C. Iron-magnesium antagonism in growth and metabolism of radish. Plant Soil 1984, 80, 355–361. [Google Scholar]
- Mehrabi, F.; Sepaskhah, A.R.; Ahmadi, S.H. Winter wheat root distribution with irrigation, planting methods, and nitrogen application. Nutr. Cycl. Agroecosystems 2021, 119, 231–245. [Google Scholar] [CrossRef]
- Zhao, D.; Zhang, X.; Zhao, W.; Zhao, S.; Liu, G.; Siddique, K.H.M. Root distribution, agronomic performance, and phosphorus utilization in wheat as mediated by phosphorus placement under rainfed coastal saline conditions. Agronomy 2023, 13, 2700. [Google Scholar] [CrossRef]
- Setter, T.L.; Waters, I.; Stefanova, K.; Munns, R.; Barrett-Lennard, E.G. Salt tolerance, date of flowering and rain affect the productivity of wheat and barley on rainfed saline land. Field Crop Res. 2016, 194, 31–42. [Google Scholar]
- Zhao, D.; Gao, S.; Zhang, X.; Zhang, Z.; Zheng, H.; Rong, K.; Zhao, W.; Khan, S.A. Impact of saline stress on the uptake of various macro and micronutrients and their associations with plant biomass and root traits in wheat. Plant Soil Environ. 2021, 67, 61–70. [Google Scholar]
- Joshi, A.K.; Crossa, J.; Arun, B.; Chand, R.; Trethowan, R.; Vargas, M.; Ortiz-Monasterio, I. Genotype× environment interaction for zinc and iron concentration of wheat grain in eastern Gangetic plains of India. Field Crop Res. 2010, 116, 268–277. [Google Scholar]
- Li, C.; Guo, Z.; Wang, X.; Ma, Y.; Liu, J.; Shi, M.; Zhang, D.; Malhi, S.S.; Siddique, K.H.M.; Wang, Z.H. Field-scale studies quantify limitations for wheat grain zinc biofortification in dryland areas. Eur. J. Agron. 2023, 142, 126687. [Google Scholar]
- Feng, D.; Wang, X.; Gao, J.; Zhang, C.; Liu, H.; Liu, P.; Sun, X. Exogenous calcium: Its mechanisms and research advances involved in plant stress tolerance. Front. Plant Sci. 2023, 14, 1143963. [Google Scholar] [CrossRef]
- Li, L.; Tang, C.; Rengel, Z.; Zhang, F.S. Calcium, magnesium and microelement uptake as affected by phosphorus sources and interspecific root interactions between wheat and chickpea. Plant Soil 2004, 261, 29–37. [Google Scholar]
- Zhao, D.; Zhang, X.; Zhao, S.; Liu, G.; Zhang, Z.; Zhao, W.; Li, X.; Khan, S.A.; Siddique, K.H.M. Biomass allocation and nutrients utilization in wheat as affected by phosphorus placement and salt stress. Agronomy 2023, 13, 1570. [Google Scholar] [CrossRef]
- Hui, X.; Wang, X.; Luo, L.; Wang, S.; Guo, Z.; Shi, M.; Wang, R.; Lyons, G.; Chen, Y.; Cakmak, I.; et al. Wheat grain zinc concentration as affected by soil nitrogen and phosphorus availability and root mycorrhizal colonization. Eur. J. Agron. 2022, 134, 126469. [Google Scholar]
- Zhao, F.J.; Su, Y.H.; Dunham, S.J.; Rakszegi, M.; Bedo, Z.; McGrath, S.P.; Shewry, P.R. Variation in mineral micronutrient concentrations in grain of wheat lines of diverse origin. J. Cereal Sci. 2009, 49, 290–295. [Google Scholar]
- Shaukat, M.; Sun, M.; Ali, M.; Mahmood, T.; Naseer, S.; Maqbool, S.; Rehman, S.; Mahmood, Z.; Hao, Y.; Xia, X.; et al. Genetic Gain for Grain Micronutrients and Their Association with Phenology in Historical Wheat Cultivars Released between 1911 and 2016 in Pakistan. Agronomy 2021, 11, 1247. [Google Scholar] [CrossRef]
- Luo, Y.; Li, Y.; Li, W.; Zhang, S.; Mu, W.; Huang, N.; Sun, R.; Ding, Y.; She, W.; Song, W.; et al. Iron concentrations in grain and its different parts of newly developed wheat Varieties (lines) in China and influencing factors. Sci. Agric. Sin. 2025, 58, 416–430. [Google Scholar]
- Marcos-Barbero, E.L.; Pérez, P.; Martínez-Carrasco, R.; Arellano, J.B.; Morcuende, R. Genotypic Variability on Grain Yield and Grain Nutritional Quality Characteristics of Wheat Grown under Elevated CO2 and High Temperature. Plants 2021, 10, 1043. [Google Scholar] [CrossRef]
- Klikocka, H.; Cybulska, M.; Barczak, B.; Narolski, B.; Wójcik, E. The effect of sulphur and nitrogen fertilization on grain yield and technological quality of spring wheat. Plant Soil Environ. 2016, 62, 230–236. [Google Scholar]
- Bouzid, N. Selenium as a powerful trace element for mitigation of plant salt stress: A review. J. Trace Elem. Miner. 2024, 8, 100123. [Google Scholar]
- Hussain, S.; Ahmed, S.; Akram, W.; Ahmad, A.; Yasin, N.A.; Fu, M.; Li, G.; Sardar, R. The potential of selenium to induce salt stress tolerance in Brassica rapa: Evaluation of biochemical, physiological and molecular phenomenon. Plant Stress 2024, 11, 100331. [Google Scholar]
- Dresler, S.; Hawrylak-Nowak, B.; Kováik, J.; Woniak, M.; Sowa, I. Organic nitrogen modulates not only cadmium toxicity but also microbial activity in plants. J. Hazard Mater. 2021, 402, 123887. [Google Scholar]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Season | N Supply | PH (cm) | FSNH | GNH | TKW (g) | GY (kg/ha) | SW (kg/ha) |
|---|---|---|---|---|---|---|---|
| 2020–2021 | N0 | 53.0 ± 4.4 c | 15.4 ± 0.7 a | 41.6 ± 2.7 b | 40.1 ± 2.2 c | 4013.7 ± 611.8 c | 4404.5 ± 602.1 c |
| N100 | 54.3 ± 4.3 b | 15.6 ± 0.6 a | 41.4 ± 3.4 b | 40.5 ± 2.2 b | 4706.6 ± 710.2 b | 5136.8 ± 775.8 b | |
| N200 | 57.1 ± 5.2 a | 15.8 ± 0.9 a | 44.6 ± 2.0 a | 40.8 ± 2.3 a | 5782.3 ± 890.0 a | 6110.8 ± 1009.9 a | |
| 2021–2022 | N0 | 55.0 ± 4.4 b | 16.0 ± 0.8 b | 42.6 ± 3.3 b | 41.2 ± 2.3 c | 4158.2 ± 619.7 c | 4120.3 ± 658.1 c |
| N100 | 57.1 ± 5.7 a | 16.2 ± 0.7 a | 43.3 ± 3.8 b | 41.5 ± 2.4 b | 4921.2 ± 784.8 b | 5095.3 ± 1068.2 b | |
| N200 | 58.9 ± 4.6 a | 16.4 ± 1.0 a | 46.6 ± 2.4 a | 41.9 ± 2.4 a | 6073.9 ± 916.7 a | 6096.9 ± 1326.6 a |
| Growing Season | Genotype | PH (cm) | FSNH | GNH | TKW (g) | GY (kg/ha) | SW (kg/ha) |
|---|---|---|---|---|---|---|---|
| 2020–2021 | Hongdi95 | 49.6 ± 0.9 d | 15.1 ± 0.5 cd | 41.0 ± 3.3 bc | 40.3 ± 0.6 bc | 3667.8 ± 558.4 d | 3964.1 ± 723.4 d |
| Jimai106 | 50.7 ± 1.3 d | 15.9 ± 0.7 b | 44.2 ± 1.2 a | 43.3 ± 0.9 a | 4759.1 ± 1176.8 c | 5219.7 ± 1245.3 bc | |
| Jimai60 | 62.9 ± 2.2 a | 15.5 ± 0.7 bc | 44.2 ± 2.1 a | 43.5 ± 0.5 a | 5457.7 ± 781.3 ab | 5730.2 ± 650.4 ab | |
| Jimai775 | 54.6 ± 2.7 c | 16.4 ± 0.7 a | 44.4 ± 3.9 a | 36.7 ± 0.8 e | 5782.7 ± 1363.7 a | 6135.1 ± 1472.2 a | |
| LJJ803 | 55.4 ± 5.0 bc | 15.5 ± 0.6 bc | 38.6 ± 4.4 c | 38.8 ± 0.9 d | 4572.5 ± 906.3 c | 5166.9 1118.3 bc | |
| LS018R | 54.4 ± 3.9 c | 14.8 ± 0.4 d | 40.4 ± 3.4 c | 40.6 ± 0.6 b | 4836.0 ± 372.7 c | 5148.5 ± 357.9 bc | |
| LS3666 | 56.5 ± 3.9 b | 15.8 ± 0.7 b | 43.2 ± 1.0 ab | 39.7 ± 1.3 c | 4591.5 ± 549.2 c | 4904.5 ± 514.7 c | |
| Shannong25 | 54.3 ± 3.6 c | 15.7 ± 0.5 b | 42.9 ± 3.2 ab | 40.8 ± 0.8 b | 5006.3 ± 988.3 bc | 5469.8 ± 769.9 abc | |
| 2021–2022 | Hongdi95 | 51.6 ± 1.7 d | 15.6 ± 0.6 c | 43.1 ± 4.0 bcd | 41.2 ± 1.0 bc | 3865.3 ± 585.8 c | 4033.9 ± 860.0 b |
| Jimai106 | 53.1 ± 1.7 d | 16.4 ± 0.7 b | 46.0 ± 1.3 ab | 44.9 ± 1.0 a | 4817.0 ± 1229.7 b | 5128.9 ± 1028.4 ab | |
| Jimai60 | 64.9 ± 3.4 a | 16.1 ± 0.6 bc | 46.4 ± 2.2 a | 44.2 ± 0.5 a | 5696.0 ± 807.7 a | 5820.1 ± 979.5 a | |
| Jimai775 | 57.7 ± 3.6 bc | 17.3 ± 0.7 a | 45.8 ± 4.4 ab | 37.3 ± 0.9 e | 6081.7 ± 1444.8 a | 5078.9 ± 1484.1 ab | |
| LJJ803 | 57.6 ± 5.8 bc | 16.0 ± 0.8 bc | 41.2 ± 3.3 d | 39.8 ± 1.2 d | 4770.6 ± 949.7 b | 5655.9 ± 1817.6 a | |
| LS018R | 56.0 ± 3.7 c | 15.6 ± 0.5 c | 41.7 ± 3.7 cd | 42.0 ± 0.7 b | 5120.7 ± 413.2 b | 5135.2 ± 1060.2 ab | |
| LS3666 | 58.4 ± 4.2 b | 16.4 ± 0.8 b | 44.6 ± 1.8 abc | 40.9 ± 1.3 c | 4867.4 ± 527.3 b | 4297.9 ± 707.8 b | |
| Shannong25 | 56.4 ± 3.5 bc | 16.2 ± 0.4 bc | 44.5 ± 3.8 abc | 41.7 ± 1.1 bc | 5190.3 ± 1210.6 b | 5682.6 ± 1496.8 a |
| Growing Season | Genotype | Na (mg/kg) | K (g/kg) | Ca (mg/kg) | Mg (g/kg) | P (g/kg) | Zn (mg/kg) | Fe (mg/kg) | Se (µg/kg) |
|---|---|---|---|---|---|---|---|---|---|
| 2020–2021 | Hongdi95 | 51.7 ± 18.5 b | 4.6 ± 0.5 a | 524.3 ± 41.2 a | 1.6 ± 0.1 ab | 4.4 ± 0.5 a | 28.4 ± 2.8 ab | 51.9 ± 3.1 bc | 55.8 ± 6.7 ab |
| Jimai106 | 43.3 ± 3.5 c | 4.2 ± 0.4 abc | 501.8 ± 52.8 ab | 1.5 ± 0.1 bc | 3.9 ± 0.4 b | 30.2 ± 4.6 a | 55.1 ± 4.5 b | 50.6 ± 7.9 c | |
| Jimai60 | 64.3 ± 14.1 a | 4.4 ± 0.3 a | 441.8 ± 29.8 d | 1.4 ± 0.1 c | 3.4 ± 0.5 c | 23.1 ± 2.9 de | 50.1 ± 3.8 c | 54.2 ± 7.1 bc | |
| Jimai775 | 45.2 ± 5.2 c | 4.0 ± 0.4 bc | 487.9 ± 59.8 abc | 1.6 ± 0.1 ab | 3.4 ± 0.5 c | 25.1 ± 4.5 cd | 48.4 ± 4.8 cd | 57.3 ± 5.4 ab | |
| LJJ803 | 49.9 ± 14.1 b | 4.4 ± 0.4 a | 511.9 ± 40.3 ab | 1.6 ± 0.2 a | 4.0 ± 0.5 b | 26.5 ± 4.6 bc | 63.6 ± 15.3 a | 57.2 ± 3.9 ab | |
| LS018R | 35.3 ± 4.4 d | 4.3 ± 0.4 ab | 486.4 ± 43.1 abcd | 1.5 ± 0.2 bc | 3.3 ± 0.5 c | 23.8 ± 2.6 d | 49.8 ± 2.1 c | 56.8 ± 3.9 ab | |
| LS3666 | 52.2 ± 13.6 b | 4.2 ± 0.3 abc | 449.5 ± 82.6 cd | 1.4 ± 0.1 c | 3.2 ± 0.2 cd | 21.3 ± 1.2 e | 44.9 ± 2.4 d | 58.6 ± 9.1 ab | |
| Shannong25 | 44.8 ± 9.4 c | 3.9 ± 0.3 c | 478.4 ± 36.6 bcd | 1.4 ± 0.1 c | 2.9 ± 0.2 d | 23.5 ± 3.8 de | 49.6 ± 2.7 c | 60.0 ± 10.4 a | |
| 2021–2022 | Hongdi95 | 47.6 ± 17.2 b | 4.1 ± 0.4 a | 489.7 ± 31.9 a | 1.4 ± 0.1 ab | 4.1 ± 0.4 a | 35.1 ± 3.5 ab | 48.5 ± 3.2 bc | 51.8 ± 6.2 bc |
| Jimai106 | 39.7 ± 3.6 c | 3.9 ± 0.4 b | 470.1 ± 54.8 a | 1.3 ± 0.1 bc | 3.6 ± 0.2 b | 35.9 ± 3.4 a | 51.9 ± 3.0 b | 47.0 ± 5.8 d | |
| Jimai60 | 58.5 ± 11.1 a | 3.9 ± 0.2 ab | 406.8 ± 16.8 c | 1.3 ± 0.1 c | 3.2 ± 0.5 c | 30.4 ± 6.2 cd | 46.4 ± 2.9 cd | 49.5 ± 5.5 cd | |
| Jimai775 | 41.5 ± 4.8 c | 3.6 ± 0.4 c | 453.9 ± 57.2 ab | 1.4 ± 0.1 ab | 3.2 ± 0.5 c | 31.2 ± 6.9 bc | 45.2 ± 4.2 cd | 53.1 ± 4.6 abc | |
| LJJ803 | 48.1 ± 13.6 b | 4.1 ± 0.3 ab | 481.6 ± 44.5 a | 1.5 ± 0.2 a | 3.7 ± 0.5 b | 32.8 ± 5.3 abc | 60.4 ± 1.3 a | 53.8 ± 2.5 abc | |
| LS018R | 33.2 ± 3.7 d | 3.9 ± 0.4 ab | 454.8 ± 47.3 ab | 1.3 ± 0.2 bc | 3.0 ± 0.4 c | 30.8 ± 5.6 bc | 47.1 ± 3.1 cd | 52.6 ± 3.3 bc | |
| LS3666 | 48.9 ± 13.0 b | 3.9 ± 0.2 c | 427.9 ± 79.2 bc | 1.3 ± 0.1 c | 3.0 ± 0.2 cd | 26.1 ± 2.5 d | 42.9 ± 2.6 d | 55.3 ± 7.8 ab | |
| Shannong25 | 41.5 ± 8.5 c | 3.5 ± 0.3 c | 453.6 ± 38.5 ab | 1.3 ± 0.2 bc | 2.8 ± 0.3 d | 30.4 ± 5.2 cd | 47.5 ± 3.6 c | 57.2 ± 8.9 a |
| Growing Season | Genotype | NAE (kg/kg) | |
|---|---|---|---|
| N100 | N200 | ||
| 2020–2021 | Hongdi95 | 4.6 ± 0.3 c | 6.0 ± 0.4 b |
| Jimai106 | 6.1 ± 1.9 c | 13.0 ± 0.8 ab | |
| Jimai60 | 1.5 ± 0.2 d | 7.6 ± 0.5 b | |
| Jimai775 | 18.7 ± 2.1 a | 15.4 ± 0.9 a | |
| LJJ803 | 8.0 ± 0.7 c | 10.0 ± 0.6 b | |
| LS018R | 2.1 ± 0.6 d | 3.3 ± 0.7 c | |
| LS3666 | 0.5 ± 0.1 d | 5.1 ± 0.3 c | |
| Shannong25 | 14.9 ± 1.5 b | 10.2 ± 2.0 b | |
| 2021–2022 | Hongdi95 | 6.3 ± 0.9 c | 4.2 ± 1.7 cd |
| Jimai106 | 13.4 ± 1.7 ab | 6.4 ± 0.9 c | |
| Jimai60 | 7.8 ± 1.7 bc | 3.5 ± 0.4 cd | |
| Jimai775 | 16.0 ± 0.6 a | 20.4 ± 4.7 a | |
| LJJ803 | 10.4 ± 1.8 b | 8.2 ± 2.4 c | |
| LS018R | 3.8 ± 1.6 c | 3.1 ± 1.1 cd | |
| LS3666 | 5.4 ± 0.3 c | 2.2 ± 0.6 d | |
| Shannong25 | 13.6 ± 0.7 ab | 14.2 ± 1.4 b | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Y.; Zhao, S.; Liu, G.; Li, J.; Siddique, K.H.M.; Zhao, D. Agronomic Traits, Nutrient Accumulation, and Their Correlations in Wheat, as Affected by Nitrogen Supply in Rainfed Coastal Saline Soils. Plants 2025, 14, 1022. https://doi.org/10.3390/plants14071022
Li Y, Zhao S, Liu G, Li J, Siddique KHM, Zhao D. Agronomic Traits, Nutrient Accumulation, and Their Correlations in Wheat, as Affected by Nitrogen Supply in Rainfed Coastal Saline Soils. Plants. 2025; 14(7):1022. https://doi.org/10.3390/plants14071022
Chicago/Turabian StyleLi, Yan, Shuaipeng Zhao, Guolan Liu, Jian Li, Kadambot H. M. Siddique, and Deyong Zhao. 2025. "Agronomic Traits, Nutrient Accumulation, and Their Correlations in Wheat, as Affected by Nitrogen Supply in Rainfed Coastal Saline Soils" Plants 14, no. 7: 1022. https://doi.org/10.3390/plants14071022
APA StyleLi, Y., Zhao, S., Liu, G., Li, J., Siddique, K. H. M., & Zhao, D. (2025). Agronomic Traits, Nutrient Accumulation, and Their Correlations in Wheat, as Affected by Nitrogen Supply in Rainfed Coastal Saline Soils. Plants, 14(7), 1022. https://doi.org/10.3390/plants14071022