



Carrier-Based Application of Phyto-Benefic and Salt-Tolerant Bacillus wiedmannii and Bacillus paramobilis for Sustainable Wheat Production Under Salinity Stress


Abstract
1. Introduction
2. Results
2.1. Isolation and Purification of Bacteria
2.2. PGP Characterization of Salt-Tolerant Rhizobacteria
2.3. Molecular Characterization of Bacterial Strains
2.4. Characterization of the Carrier and Survival Percentage of PGPR in Carriers
2.5. Soil Analysis
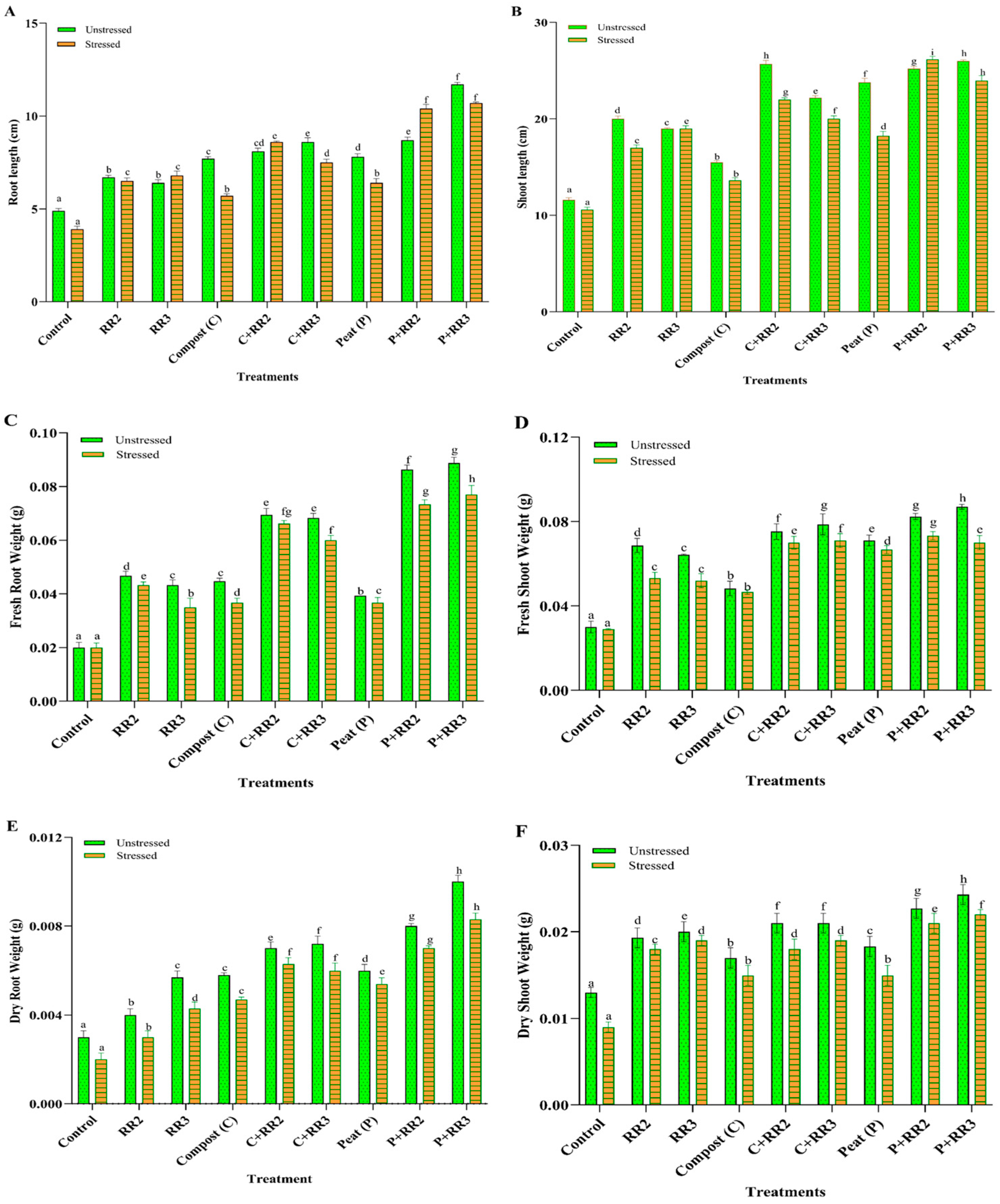
2.6. Seedling Characteristics of Wheat Under Laboratory Conditions
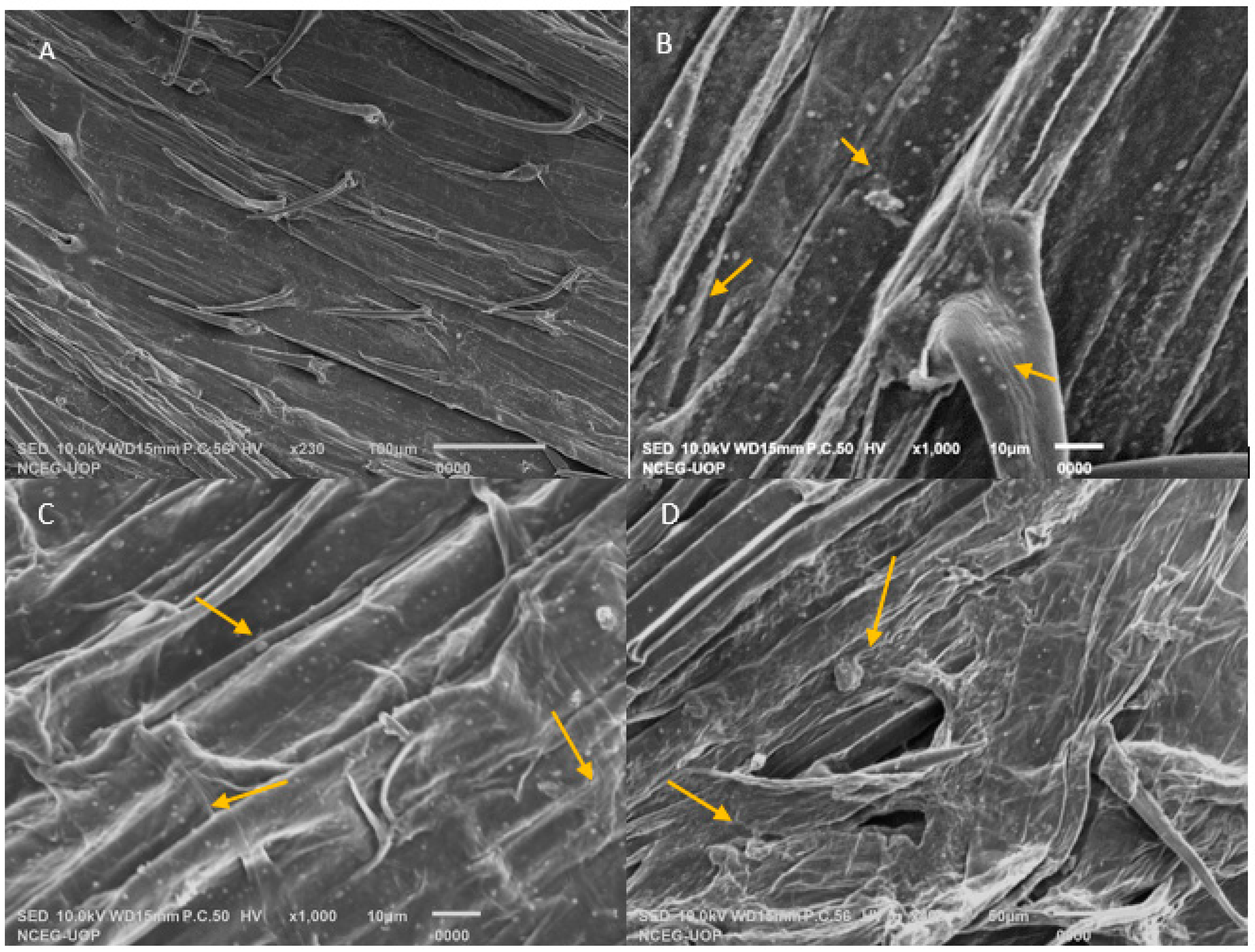
2.7. Root Colonization
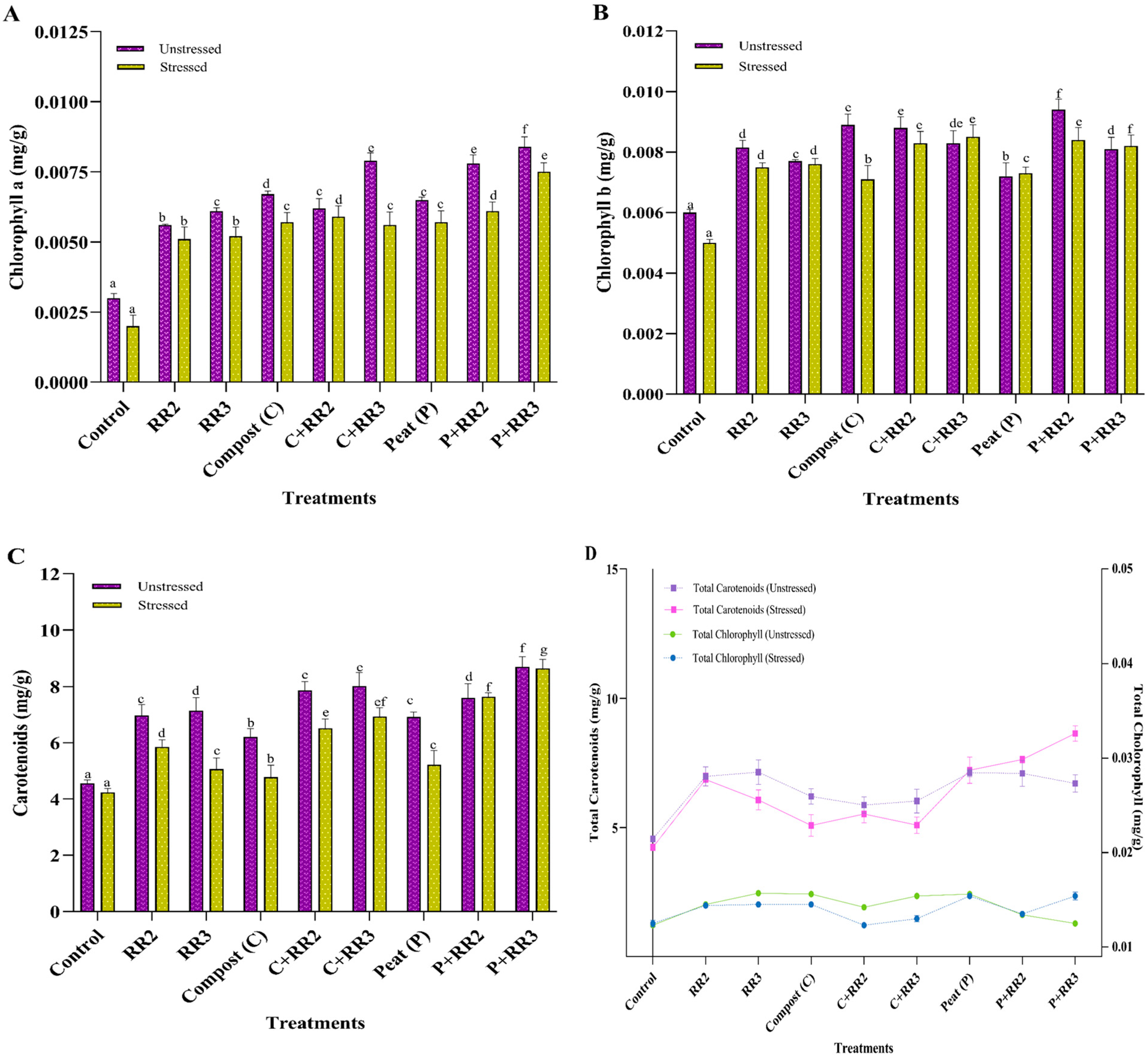
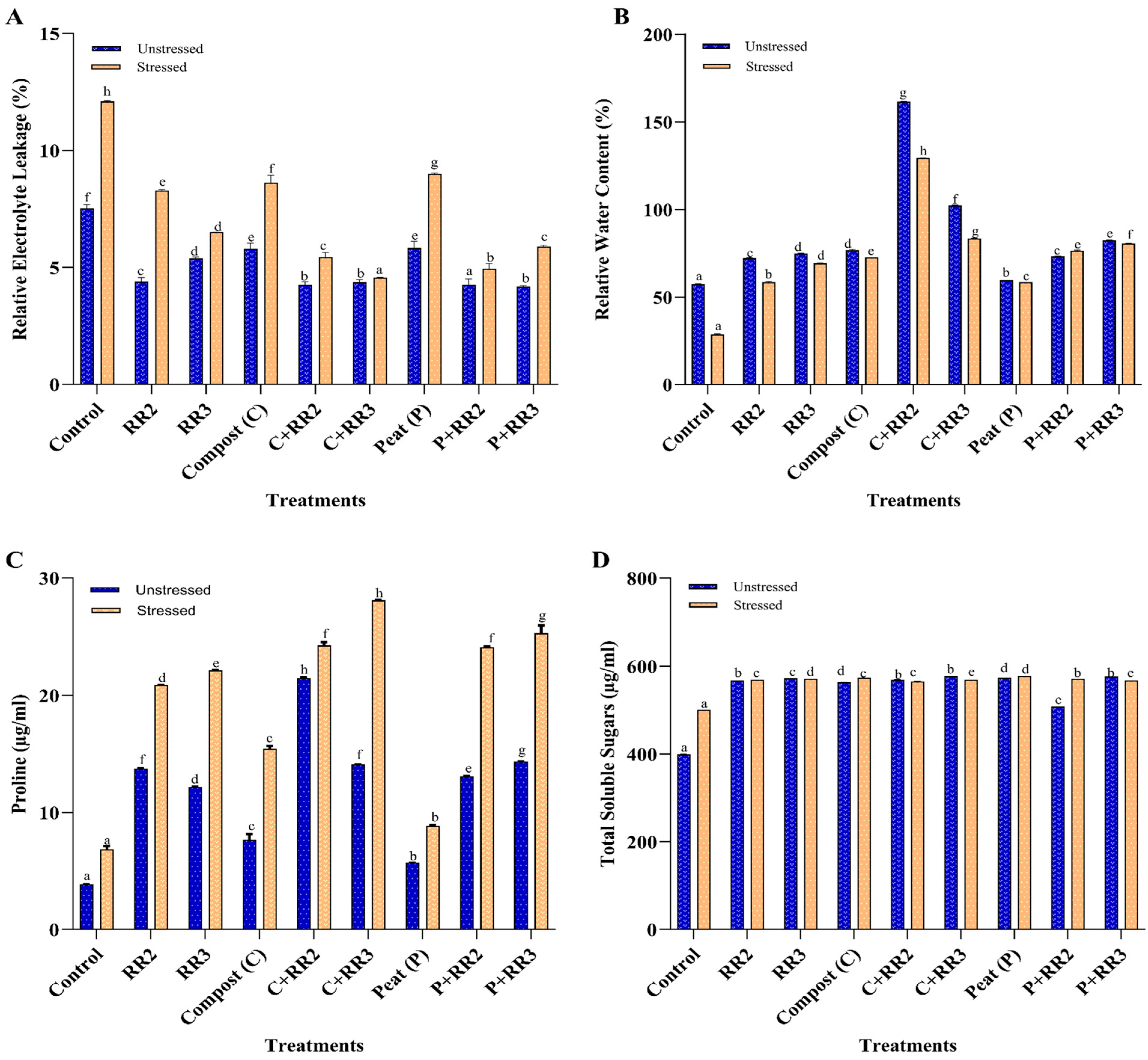
2.8. Physiological and Biochemical Parameters of Wheat Under Natural Conditions
2.9. Yield Parameters of Wheat Under Natural Conditions
3. Discussion
4. Materials and Methodology
4.1. Plant and Bacterial Strains
4.2. Sampling, Isolation, and Purification of Bacteria
4.3. Screening of Salt-Tolerant Strains
4.4. Identification of Salt-Tolerant Strains
4.5. PGPR Characterizations
4.5.1. Screening Based on Auxin Production
4.5.2. Screening Based on Phosphate Solubilization
4.5.3. Screening Based on Nitrogen Fixation
4.5.4. Screening Based on HCN, Ammonia, and Exopolysaccharide Production
4.5.5. Screening Based on the Production of Cell Wall-Degrading Enzymes
4.6. Characterization of Carrier Material
Stability of Bacteria in Carriers
4.7. Nutrient Analysis of Soil
4.8. Experimental Layout for Plant–Microbe Interaction
Plant–Microbe Interaction
4.9. Root Colonization Assay
4.10. Physiological Parameters
4.10.1. Chlorophyll Content
4.10.2. Relative Electrolyte Leakage (REL)
4.10.3. Relative Water Content (RWC)
4.11. Biochemical Parameters
4.12. Yield Parameters
4.13. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Seleiman, M.F.; Aslam, M.T.; Alhammad, B.A.; Hassan, M.U.; Maqbool, R.; Chattha, M.U.; Khan, I.; Gitari, H.I.; Uslu, O.S.; Roy, R. Salinity stress in wheat: Effects, mechanisms and management strategies. Phyton (0031-9457) 2022, 91, 667–694. [Google Scholar] [CrossRef]
- Kumar, A.; Maurya, B.R.; Raghuwanshi, R. The microbial consortium of indigenous rhizobacteria improving plant health, yield and nutrient content in wheat (Triticum aestivum). J. Plant Nutr. 2021, 44, 1942–1956. [Google Scholar] [CrossRef]
- Zahra, S.T.; Tariq, M.; Abdullah, M.; Azeem, F.; Ashraf, M.A. Dominance of Bacillus species in the wheat (Triticum aestivum L.) rhizosphere and their plant growth promoting potential under salt stress conditions. PeerJ 2023, 11, e14621. [Google Scholar] [CrossRef]
- Adil, M.; Bashir, S.; Bashir, S.; Aslam, Z.; Ahmad, N.; Younas, T.; Asghar, R.M.A.; Alkahtani, J.; Dwiningsih, Y.; Elshikh, M.S. Zinc oxide nanoparticles improved chlorophyll contents, physical parameters, and wheat yield under salt stress. Front. Plant Sci. 2022, 13, 932861. [Google Scholar] [CrossRef]
- Huang, Z.; Wang, C.; Feng, Q.; Liou, R.-M.; Lin, Y.-F.; Qiao, J.; Lu, Y.; Chang, Y. The mechanisms of sodium chloride stress mitigation by salt-tolerant plant growth promoting rhizobacteria in wheat. Agronomy 2022, 12, 543. [Google Scholar] [CrossRef]
- Ayaz, M.; Ali, Q.; Jiang, Q.; Wang, R.; Wang, Z.; Mu, G.; Khan, S.A.; Khan, A.R.; Manghwar, H.; Wu, H. Salt tolerant Bacillus strains improve plant growth traits and regulation of phytohormones in wheat under salinity stress. Plants 2022, 11, 2769. [Google Scholar] [CrossRef]
- Kulkova, I.; Dobrzyński, J.; Kowalczyk, P.; Bełżecki, G.; Kramkowski, K. Plant growth promotion using Bacillus cereus. Int. J. Mol. Sci. 2023, 24, 9759. [Google Scholar] [CrossRef]
- Zhou, Y.; Sang, T.; Tian, M.; Jahan, M.S.; Wang, J.; Li, X.; Guo, S.; Liu, H.; Wang, Y.; Shu, S. Effects of Bacillus cereus on photosynthesis and antioxidant metabolism of cucumber seedlings under salt stress. Horticulturae 2022, 8, 463. [Google Scholar] [CrossRef]
- Jung, H.; Choi, S.; Kim, Y.; Han, J.A.; Lee, H.-S.; Kim, E.Y. Complete genome sequence of Bacillus paramobilis sp. strain IMGN7 from soil. Microbiol. Resour. Announc. 2024, 13, e00540-24. [Google Scholar] [CrossRef]
- July, E.; Gillis, A. Antiviral defence arsenal across members of the Bacillus cereus group. Sci. Rep. 2025, 15, 4958. [Google Scholar] [CrossRef]
- Orozco-Mosqueda, M.D.C.; Santoyo, G.; Glick, B.R. Recent advances in the bacterial phytohormone modulation of plant growth. Plants 2023, 12, 606. [Google Scholar] [CrossRef]
- Yin, Z.-y.; Yuan, Y.-c.; Zhang, R.; Gan, J.-t.; Yu, L.; Qiu, X.-h.; Chen, R.-p.; Wang, Q. Understanding Bacillus response to salt stress: Growth inhibition, enhanced EPS secretion, and molecular adaptation mechanisms. Process Biochem. 2024, 146, 412–422. [Google Scholar] [CrossRef]
- Roy, B.; Maitra, D.; Biswas, A.; Chowdhury, N.; Ganguly, S.; Bera, M.; Dutta, S.; Golder, S.; Roy, S.; Ghosh, J. Efficacy of high-altitude biofilm-forming novel Bacillus subtilis species as plant growth-promoting rhizobacteria on Zea mays L. Appl. Biochem. Biotechnol. 2024, 196, 643–666. [Google Scholar] [CrossRef]
- Perveen, R.; Hussain, A.; Ditta, A.; Dar, A.; Aimen, A.; Ahmad, M.; Alataway, A.; Dewidar, A.Z.; Mattar, M.A. Growth and yield of Okra exposed to a consortium of rhizobacteria with different organic carriers under controlled and natural field conditions. Horticulturae 2022, 9, 8. [Google Scholar] [CrossRef]
- Omara, A.E.-D.; Hafez, E.M.; Osman, H.S.; Rashwan, E.; El-Said, M.A.; Alharbi, K.; Abd El-Moneim, D.; Gowayed, S.M. Collaborative impact of compost and beneficial rhizobacteria on soil properties, physiological attributes, and productivity of wheat subjected to deficit irrigation in salt affected soil. Plants 2022, 11, 877. [Google Scholar] [CrossRef]
- Kumar, S.; Li, G.; Yang, J.; Huang, X.; Ji, Q.; Liu, Z.; Ke, W.; Hou, H. Effect of Salt Stress on Growth, Physiological Parameters, and Ionic Concentration of Water Dropwort (Oenanthe javanica) Cultivars. Front. Plant Sci. 2021, 12, 660409. [Google Scholar] [CrossRef]
- Noureen, S.; Iqbal, A.; Muqeet, H.A. Potential of drought tolerant Rhizobacteria amended with biochar on growth promotion in Wheat. Plants 2024, 13, 1183. [Google Scholar] [CrossRef]
- El Sabagh, A.; Islam, M.S.; Skalicky, M.; Ali Raza, M.; Singh, K.; Anwar Hossain, M.; Hossain, A.; Mahboob, W.; Iqbal, M.A.; Ratnasekera, D. Salinity stress in wheat (Triticum aestivum L.) in the changing climate: Adaptation and management strategies. Front. Agron. 2021, 3, 661932. [Google Scholar] [CrossRef]
- Farooq, A.; Farooq, N.; Akbar, H.; Hassan, Z.U.; Gheewala, S.H. A critical review of climate change impact at a global scale on cereal crop production. Agronomy 2023, 13, 162. [Google Scholar] [CrossRef]
- Alamer, K.H. Alleviatory Role of Boron Supplementation on the Adverse Effects of Salinity Stress in Wheat. J. Plant Growth Regul. 2025, 44, 3179–3192. [Google Scholar] [CrossRef]
- Joshi, S.; Gangola, S.; Jaggi, V.; Sahgal, M. Functional characterization and molecular fingerprinting of potential phosphate solubilizing bacterial candidates from Shisham rhizosphere. Sci. Rep. 2023, 13, 7003. [Google Scholar] [CrossRef] [PubMed]
- Amri, M.; Rjeibi, M.R.; Gatrouni, M.; Mateus, D.M.; Asses, N.; Pinho, H.J.; Abbes, C. Isolation, identification, and characterization of phosphate-solubilizing bacteria from Tunisian soils. Microorganisms 2023, 11, 783. [Google Scholar] [CrossRef] [PubMed]
- Tu, Q.; Tang, S.; Huang, S. Mitigation of salinity stress via improving growth, chlorophyll contents and antioxidants defense in sunflower with Bacillus pumilis and biochar. Sci. Rep. 2025, 15, 9641. [Google Scholar] [CrossRef] [PubMed]
- Haroon, U.; Khizar, M.; Liaquat, F.; Ali, M.; Akbar, M.; Tahir, K.; Batool, S.S.; Kamal, A.; Chaudhary, H.J.; Munis, M.F.H. Halotolerant plant growth-promoting rhizobacteria induce salinity tolerance in wheat by enhancing the expression of SOS genes. J. Plant Growth Regul. 2021, 41, 2435–2448. [Google Scholar] [CrossRef]
- Kerbab, S.; Silini, A.; Chenari Bouket, A.; Cherif-Silini, H.; Eshelli, M.; El Houda Rabhi, N.; Belbahri, L. Mitigation of NaCl stress in wheat by rhizosphere engineering using salt habitat adapted PGPR halotolerant bacteria. Appl. Sci. 2021, 11, 1034. [Google Scholar] [CrossRef]
- Devi, S.; Sharma, S.; Tiwari, A.; Bhatt, A.K.; Singh, N.K.; Singh, M.; Kaushalendra; Kumar, A. Screening for multifarious plant growth promoting and biocontrol attributes in bacillus strains isolated from indo gangetic soil for enhancing growth of rice crops. Microorganisms 2023, 11, 1085. [Google Scholar] [CrossRef]
- Iqbal, M.; Naveed, M.; Sanaullah, M.; Brtnicky, M.; Hussain, M.I.; Kucerik, J.; Holatko, J.; Mustafa, A. Plant microbe mediated enhancement in growth and yield of canola (Brassica napus L.) plant through auxin production and increased nutrient acquisition. J. Soils Sediments 2023, 23, 1233–1249. [Google Scholar] [CrossRef]
- Jensen, C.N.G.; Pang, J.K.Y.; Gottardi, M.; Kračun, S.K.; Svendsen, B.A.; Nielsen, K.F.; Kovács, Á.T.; Moelbak, L.; Fimognari, L.; Husted, S. Bacillus subtilis promotes plant phosphorus (P) acquisition through P solubilization and stimulation of root and root hair growth. Physiol. Plant. 2024, 176, e14338. [Google Scholar] [CrossRef]
- Timofeeva, A.M.; Galyamova, M.R.; Sedykh, S.E. Plant growth-promoting soil bacteria: Nitrogen fixation, phosphate solubilization, siderophore production, and other biological activities. Plants 2023, 12, 4074. [Google Scholar] [CrossRef]
- Balaji, L.; Chittoor, J.T.; Jayaraman, G. Optimization of extracellular lipase production by halotolerant Bacillus sp. VITL8 using factorial design and applicability of enzyme in pretreatment of food industry effluents. Prep. Biochem. Biotechnol. 2020, 50, 708–716. [Google Scholar] [CrossRef]
- Jabborova, D.P.; Narimanov, A.A.; Enakiev, Y.I.; Davranov, K.D. Effect of Bacillus subtilis 1 strain on the growth and development of wheat (Triticum aestivum L.) under saline condition. Bulg. J. Agric. Sci. 2020, 26, 744–747. [Google Scholar]
- Afzal, A.; Bahader, S.; Ul Hassan, T.; Naz, I.; Din, A.-u.-. Rock phosphate solubilization by plant growth-promoting Bacillus velezensis and its impact on wheat growth and yield. Geomicrobiol. J. 2023, 40, 131–142. [Google Scholar] [CrossRef]
- Ibarra-Villarreal, A.L.; Gándara-Ledezma, A.; Godoy-Flores, A.D.; Herrera-Sepúlveda, A.; Díaz-Rodríguez, A.M.; Parra-Cota, F.I.; de los Santos-Villalobos, S. Salt-tolerant Bacillus species as a promising strategy to mitigate the salinity stress in wheat (Triticum turgidum subsp. durum). J. Arid. Environ. 2021, 186, 104399. [Google Scholar] [CrossRef]
- Saddiq, M.S.; Iqbal, S.; Hafeez, M.B.; Ibrahim, A.M.; Raza, A.; Fatima, E.M.; Baloch, H.; Jahanzaib; Woodrow, P.; Ciarmiello, L.F. Effect of salinity stress on physiological changes in winter and spring wheat. Agronomy 2021, 11, 1193. [Google Scholar] [CrossRef]
- Gul, S.; Javed, S.; Azeem, M.; Aftab, A.; Anwaar, N.; Mehmood, T.; Zeshan, B. Application of Bacillus subtilis for the alleviation of salinity stress in different cultivars of Wheat (Tritium aestivum L.). Agronomy 2023, 13, 437. [Google Scholar] [CrossRef]
- Zapałowska, A.; Jarecki, W.; Skwiercz, A.; Malewski, T. Optimization of Compost and Peat Mixture Ratios for Production of Pepper Seedlings. Int. J. Mol. Sci. 2025, 26, 442. [Google Scholar] [CrossRef]
- Anđelković, J.; Mihajilov Krstev, T.; Dimkić, I.; Unković, N.; Stanković, D.; Joković, N. Growth-Promoting Effects of Ten Soil Bacterial Strains on Maize, Tomato, Cucumber, and Pepper Under Greenhouse Conditions. Plants 2025, 14, 1874. [Google Scholar] [CrossRef]
- Elbagory, M. Reducing the adverse effects of salt stress by utilizing compost tea and effective microorganisms to enhance the growth and yield of wheat (Triticum aestivum L.) plants. Agronomy 2023, 13, 823. [Google Scholar] [CrossRef]
- Sohaib, M.; Zahir, Z.A.; Khan, M.Y.; Ans, M.; Asghar, H.N.; Yasin, S.; Al-Barakah, F.N. Comparative evaluation of different carrier-based multi-strain bacterial formulations to mitigate the salt stress in wheat. Saudi J. Biol. Sci. 2020, 27, 777–787. [Google Scholar] [CrossRef]
- Yang, M.; Yang, R.; Li, Y.; Pan, Y.; Sun, J.; Zhang, Z. Effects of Different Peat Application Methods on Water and Salt Migration in a Coastal Saline Soil. J. Soil. Sci. Plant Nutr. 2022, 22, 791–800. [Google Scholar] [CrossRef]
- Shultana, R.; Kee Zuan, A.T.; Yusop, M.R.; Saud, H.M. Characterization of salt-tolerant plant growth-promoting rhizobacteria and the effect on growth and yield of saline-affected rice. PLoS ONE 2020, 15, e0238537. [Google Scholar] [CrossRef] [PubMed]
- Saleem, S.; Iqbal, A.; Ahmed, F.; Ahmad, M. Phytobeneficial and salt stress mitigating efficacy of IAA producing salt tolerant strains in Gossypium hirsutum. Saudi J. Biol. Sci. 2021, 28, 5317–5324. [Google Scholar] [CrossRef] [PubMed]
- Holt, J.G.; Krieg, N.R.; Sneath, P.H.; Staley, J.T.; Williams, S.T. Bergey’s Manual of Determinate Bacteriology; Williams & Wilkins: Baltimore, MD, USA, 1994. [Google Scholar]
- Acuña, J.J.; Rilling, J.I.; Inostroza, N.G.; Zhang, Q.; Wick, L.Y.; Sessitsch, A.; Jorquera, M.A. Variovorax sp. strain P1R9 applied individually or as part of bacterial consortia enhances wheat germination under salt stress conditions. Sci. Rep. 2024, 14, 2070. [Google Scholar] [CrossRef]
- Rico-Jiménez, M.; Muñoz-Mira, S.; Lomas-Martínez, C.; Krell, T.; Matilla, M.A. Regulation of indole-3-acetic acid biosynthesis and consequences of auxin production deficiency in Serratia plymuthica. Microb. Biotechnol. 2023, 16, 1671–1689. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, X.-l.; Zhou, J.; Zhou, W.; Zhou, S.-Q. Construction of phosphate-solubilizing microbial consortium and its effect on the remediation of saline-alkali soil. Microb. Ecol. 2025, 88, 11. [Google Scholar] [CrossRef]
- Leal, M.; Tovar, D.; Infante, A.; Barriga, O.; Ruíz, E.; Sánchez, J.; Melgarejo, L. Phosphate solubilization by microorganisms in pyroclastic material from Half Moon Island in Antarctica: Perspectives for astrobiology. Polar Biol. 2025, 48, 29. [Google Scholar] [CrossRef]
- Shomi, F.Y.; Uddin, M.B.; Zerin, T. Isolation and characterization of nitrogen-fixing bacteria from soil sample in Dhaka, Bangladesh. Stamford J. Microbiol. 2021, 11, 11–13. [Google Scholar] [CrossRef]
- Batool, S.; Iqbal, A. Phosphate solubilizing rhizobacteria as alternative of chemical fertilizer for growth and yield of Triticum aestivum (Var. Galaxy 2013). Saudi J. Biol. Sci. 2019, 26, 1400–1410. [Google Scholar] [CrossRef]
- Chaiharn, M.; Chunhaleuchanon, S.; Kozo, A.; Lumyong, S. Screening of rhizobacteria for their plant growth promoting activities. Curr. Appl. Sci. Technol. 2008, 8, 18–23. [Google Scholar]
- Thangamanil, P.; Thiruvengadam, R.; Thillaigovindan, K. Morphological characterization and reaction of partial purified toxin of sugarcane red rot pathogen Colletotrichum falcatum collected from Southern India. Int. J. Agric. Sci. 2013, 3, 60–78. [Google Scholar]
- Patil, P.S.; Kotia, P.; Azad, A.; Gedam, S.; Bansal, D.; Tyson, K. In vivo study to evaluate of human breast milk, infant milk formula on cariogenicity in children: A comparative study. Cuest. Fisioter. 2025, 54, 1246–1251. [Google Scholar]
- Siedt, M.; Schäffer, A.; Smith, K.E.; Nabel, M.; Roß-Nickoll, M.; Van Dongen, J.T. Comparing straw, compost, and biochar regarding their suitability as agricultural soil amendments to affect soil structure, nutrient leaching, microbial communities, and the fate of pesticides. Sci. Total Environ. 2021, 751, 141607. [Google Scholar] [CrossRef]
- Pour-Aboughadareh, A.; Bocianowski, J.; Jamshidi, B.; Turkoglu, A. Physiological and molecular responses of wild relatives of wheat possessing the D genome to salinity stress. Genet. Resour. Crop Evol. 2025, 72, 1819–1834. [Google Scholar] [CrossRef]
- Nadeem, M.; Shahbaz, M.; Ahmad, F.; Waraich, E.A. Enhancing wheat resistance to salinity: The role of gibberellic acid and β-Carotene in morphological, yielding and ionic adaptations. J. Ecol. Eng. 2025, 26, 76–94. [Google Scholar] [CrossRef]
- Ansari, F.; Jabeen, M.; Ahmad, I. Pseudomonas azotoformans FAP5, a novel biofilm-forming PGPR strain, alleviates drought stress in wheat plant. Int. J. Environ. Sci. Technol. 2021, 18, 3855–3870. [Google Scholar] [CrossRef]
- Morgvan, C.M.; Petrehele, A.I.G.; Fodor, A.; Cipleu, M. Influence of Manganese Ions of Chlorophyl Concentration from Wheat Grass. Fasc. Chim. 2020, XXVII, 32. [Google Scholar]
- Saeed, R.; Mirza, S.; Ahmad, R. Electrolyte leakage and relative water content as affected by organic mulch in okra plant (Abelmoschus esculentus (L.) Moench) grown under salinity. FUUAST J. Biol. 2014, 4, 221. [Google Scholar]
- Bates, L.S.; Waldren, R.; Teare, I. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Afzal, I.; Akram, M.; Rehman, H.; Rashid, S.; Basra, S. Moringa leaf and sorghum water extracts and salicylic acid to alleviate impacts of heat stress in wheat. S. Afr. J. Bot. 2020, 129, 169–174. [Google Scholar] [CrossRef]
- Mustafa, N.; Raja, N.I.; Ilyas, N.; Abasi, F.; Ahmad, M.S.; Ehsan, M.; Mehak, A.; Badshah, I.; Proćków, J. Exogenous application of green titanium dioxide nanoparticles (TiO2 NPs) to improve the germination, physiochemical, and yield parameters of wheat plants under salinity stress. Molecules 2022, 27, 4884. [Google Scholar] [CrossRef]
- İbrahimova, U.; Kumari, P.; Yadav, S.; Rastogi, A.; Antala, M.; Suleymanova, Z.; Zivcak, M.; Tahjib-Ul-Arif, M.; Hussain, S.; Abdelhamid, M. Progress in understanding salt stress response in plants using biotechnological tools. J. Biotechnol. 2021, 329, 180–191. [Google Scholar] [CrossRef] [PubMed]
- Awan, S.A.; Ilyas, N.; Khan, I.; Raza, M.A.; Rehman, A.U.; Rizwan, M.; Rastogi, A.; Tariq, R.; Brestic, M. Bacillus siamensis reduces cadmium accumulation and improves growth and antioxidant defense system in two wheat (Triticum aestivum L.) varieties. Plants 2020, 9, 878. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sr. No. | Sample Name | No. of Isolates | Plant Source | Soil Texture | Location |
|---|---|---|---|---|---|
| 1. | MUL1 | 4 | Eriobotrya japonica | Loamy soil | Multan |
| 2. | MZG1 | 4 | Bauhinia variegate | Sandy soil | Muzaffargarh |
| 3. | MUL2 | 5 | Citrus limon | Saline soil | Multan |
| 4. | MZG2 | 6 | Bauhinia variegate | Sandy soil | Muzaffargarh |
| 5. | MUL3 | 4 | Citrus limon | Saline soil | Multan |
| 6. | MUL4 | 8 | Eriobotrya japonica | Loamy soil | Multan |
| 7. | MUL5 | 5 | Citrus limon | Saline soil | Multan |
| 8. | MZG3 | 3 | Bauhinia variegate | Sandy soil | Muzaffargarh |
| 9. | MZG4 | 5 | Syzygium cumini | Loamy soil | Muzaffargarh |
| 10. | MUL5 | 6 | Eriobotrya japonica | Loamy soil | Multan |
| Bacterial Strains | Auxin (µg/mL) | Phosphate (µg/mL) | Nitrogen Fixation Zones (cm) | HCN Production | Ammonia Production | EPS Production |
|---|---|---|---|---|---|---|
| RR1 | 41.0 ± 0.7 | 217.2 ± 0.3 | 1.13 ± 0.05 | + | − | + |
| RR2 | 428.7 ± 0.5 | 1316.1 ± 0.8 | 1.33 ± 0.05 | +++ | ++ | ++ |
| RR3 | 351.1 ± 0.8 | 780.6 ± 0.5 | 2.13 ± 0.05 | ++ | ++ | + |
| RR6 | 36.5 ± 0.5 | 200.1 ± 0.7 | 0.86 ± 0.05 | − | − | − |
| RR8 | 455.7 ± 0.5 | 255.9 ± 0.3 | 2.43 ± 0.05 | ++ | − | − |
| Salt-Tolerant Strains | Chitinase | Pectinase | Catalase | Protease | Amylase |
|---|---|---|---|---|---|
| RR1 | + | − | − | − | − |
| RR2 | + | − | − | + | + |
| RR3 | + | + | − | + | − |
| RR6 | + | − | − | − | + |
| RR8 | − | − | − | + | + |
| Analytes | RR2 | RR3 |
|---|---|---|
| Indole acetic acid | − | + |
| Benzoic acid | − | + |
| n-Propyl acetate | − | + |
| Toluene | + | + |
| 2-(Pyridin-2-ylformamido)acetic | + | − |
| 4-(4-Hydroxy-2,5-dimethylbenzyl) | + | + |
| 4-tert-Butylphenol | + | − |
| 3-Trifluoroacetoxyhexadecane | − | + |
| 4-Methyl-2-trimethylsilyloxy-acetone | + | − |
| Cyclopentasiloxane, decamethyl | − | + |
| Pyrrolo[1,2-a]pyrazine-1,4-dione | − | + |
| n-hexadeconic acid | − | + |
| Bis(2-ethylhexyl)s phthalate | + | + |
| Dichloroacetic acid, heptadecyl | − | + |
| Ticosene | − | + |
| Tris(tert-butyldimethylsilyloxy) | − | + |
| Arsenous acid, tris(rimethylsil)ester | + | + |
| Methyltris(trimethylsiloxy)silane | + | + |
| Tetrasiloxane, decamethyl | − | + |
| 4-(7-Methyloctyl)phenol | + | + |
| Cyclotrisiloxane, hexamethyl | − | + |
| 1,4-Bis(trimethylsilyl)benzene | − | + |
| Ethoxy(phenyl)silanediol | + | + |
| Parameters | Compost | Peat |
|---|---|---|
| Electrical Conductivity (dS−1) | 2.15 | 1.93 |
| pH | 5.6 | 5.28 |
| Nitrogen (%) | 1.14 | 0.92 |
| Phosphorus (%) | 0.97 | 0.64 |
| Potassium (PPM) | 87.61 | 73.56 |
| Moisture content (%) | 3.89 | 3.73 |
| Water-holding capacity (%) | 28.42 | 31.12 |
| Carriers | Strains | Bacterial Population (1 × 107 CFU mg−1) at Incubation Period (Days) | ||||||
|---|---|---|---|---|---|---|---|---|
| 15 | 30 | 60 | 90 | 120 | 150 | 180 | ||
| Compost | RR2-C | 35.1 ± 0.1 c | 31.8 ± 0.6 d | 24.8 ± 0.2 c | 21.0 ± 0.7 d | 17.5 ± 0.6 d | 11.1 ± 0.9 b | 7.9 ± 1.1 b |
| RR3-C | 30.4 ± 0.2 a | 27.8 ± 0.3 c | 213 ± 0.8 b | 18.2 ± 0.6 c | 14.8 ± 0.6 c | 10.8 ± 0.5 ab | 8.3 ± 0.7 b | |
| Peat | RR2-P | 31.8 ± 0.7 b | 25.9 ± 0.7 b | 21.1 ± 0.9 b | 15.9 ± 0.1 b | 13.2 ± 0.1 b | 9.8 ± 0.3 ab | 7.4 ± 0.8 b |
| RR3-P | 29.2 ± 0.6 a | 22.9 ± 0.6 a | 19.2 ± 0.2 a | 14.3 ± 0.2 a | 11.3 ± 0.2 a | 9.5 ± 0.2 a | 5.2 ± 0.5 a | |
| Parameters | Soil |
|---|---|
| EC (dS−1) | 2.37 |
| pH | 8.6 |
| Available nitrogen (mg kg) | 91 |
| Available phosphorus (mg kg) | 4.42 |
| Available potassium (mg kg) | 98 |
| Texture | Sandy loam |
| Organic matter (%) | 0.25 |
| Saturation (%) | 11 |
| Treatments | Length (cm) | Spike Length (cm) | No. of Spikes | Weight per 100 Grains (g) | Length (cm) | Spike Length (cm) | No. of Spikes | Weight per 100 Grains (g) |
|---|---|---|---|---|---|---|---|---|
| Unstressed | Stressed | |||||||
| Control | 55.0 ± 1.0 a | 8.1 ± 1.6 a | 13.3 ± 4.0 a | 3.20 ± 0.1 a | 44.3 ± 3.7 a | 8.0 ± 1.7 a | 9.0 ± 2.6 a | 2.90 ± 0.1 a |
| T1 | 65.6 ± 1.5 c | 10.3 ± 1.5 c | 16.0 ± 3.0 b | 4.50 ± 0.01 b | 64.6 ± 4.1 c | 9.3 ± 0.5 a | 12.3 ± 2.0 b | 3.96 ± 0.1 de |
| T2 | 63.3 ± 0.5 b | 10.0 ± 1.7 c | 16.3 ± 2.0 b | 3.83 ± 0.05 a | 64.3 ± 4.0 c | 9.6 ± 0.5 a | 16.6 ± 0.5 c | 3.20 ± 0.1 b |
| T3 | 62.6 ± 5.5 b | 10.6 ± 0.5 c | 13.3 ± 2.3 a | 4.60 ± 0.01 b | 63.3 ± 1.5 c | 10.3 ± 0.5 b | 13.3 ± 3.7 b | 3.53 ± 0.05 c |
| T4 | 67.0 ± 2.6 d | 10.9 ± 1.7 c | 17.0 ± 1.0 c | 4.82 ± 0.02 b | 68.6 ± 4.5 c | 10.0 ± 1.7 b | 16.6 ± 3.7 c | 3.70 ± 0.1 c |
| T5 | 68.3 ± 1.5 e | 11.0 ± 0.00 c | 16.0 ± 1.0 b | 4.94 ± 0.06 b | 67.3 ± 4.1 c | 9.6 ± 1.1 a | 19.6 ± 4.6 d | 4.03 ± 0.05 ef |
| T6 | 62.0 ± 2.0 b | 9.0 ± 1.7 b | 14.3 ± 3.2 a | 5.13 ± 0.05 c | 57.6 ± 4.5 b | 9.6 ± 1.1 a | 17.0 ± 6.9 cd | 4.23 ± 0.05 fg |
| T7 | 68.0 ± 2.0 e | 11.0 ± 1.7 c | 21.0 ± 7.0 d | 5.23 ± 0.05 c | 65.6 ± 3.5 c | 10.3 ± 0.5 b | 19.0 ± 1.0 d | 4.46 ± 0.05 g |
| T8 | 67.3 ± 2.0 d | 12.0 ± 1.0 d | 17.6 ± 4.1 c | 5.86 ± 0.05 c | 68.6 ± 4.5 c | 10.0 ± 1.0 b | 28.0 ± 6.0 e | 4.76 ± 0.05 h |
| Name | Unstressed | Stressed (200 mM) |
|---|---|---|
| Control | No PGPR + No carrier | No PGPR + No carrier |
| T1 | PGPR (RR-2) | PGPR (RR-2) |
| T2 | PGPR (RR-3) | PGPR (RR-3) |
| T3 | Compost (C) | Compost (C) |
| T4 | C + RR-2 | C + RR-2 |
| T5 | C + RR-3 | C + RR-3 |
| T6 | Peat (P) | Peat (P) |
| T7 | P + RR-2 | P + RR-2 |
| T8 | P + RR-3 | P + RR-3 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rashid, R.; Iqbal, A.; Shahzad, M.; Noureen, S.; Muqeet, H.A. Carrier-Based Application of Phyto-Benefic and Salt-Tolerant Bacillus wiedmannii and Bacillus paramobilis for Sustainable Wheat Production Under Salinity Stress. Plants 2025, 14, 2096. https://doi.org/10.3390/plants14142096
Rashid R, Iqbal A, Shahzad M, Noureen S, Muqeet HA. Carrier-Based Application of Phyto-Benefic and Salt-Tolerant Bacillus wiedmannii and Bacillus paramobilis for Sustainable Wheat Production Under Salinity Stress. Plants. 2025; 14(14):2096. https://doi.org/10.3390/plants14142096
Chicago/Turabian StyleRashid, Raina, Atia Iqbal, Muhammad Shahzad, Sidra Noureen, and Hafiz Abdul Muqeet. 2025. "Carrier-Based Application of Phyto-Benefic and Salt-Tolerant Bacillus wiedmannii and Bacillus paramobilis for Sustainable Wheat Production Under Salinity Stress" Plants 14, no. 14: 2096. https://doi.org/10.3390/plants14142096
APA StyleRashid, R., Iqbal, A., Shahzad, M., Noureen, S., & Muqeet, H. A. (2025). Carrier-Based Application of Phyto-Benefic and Salt-Tolerant Bacillus wiedmannii and Bacillus paramobilis for Sustainable Wheat Production Under Salinity Stress. Plants, 14(14), 2096. https://doi.org/10.3390/plants14142096