
Architectural Analysis for Novel Olive Crop Management

 ,
,  , , and
, , and
Abstract

1. Introduction
2. Results
2.1. Analysis of the Topographic Architectural Variability
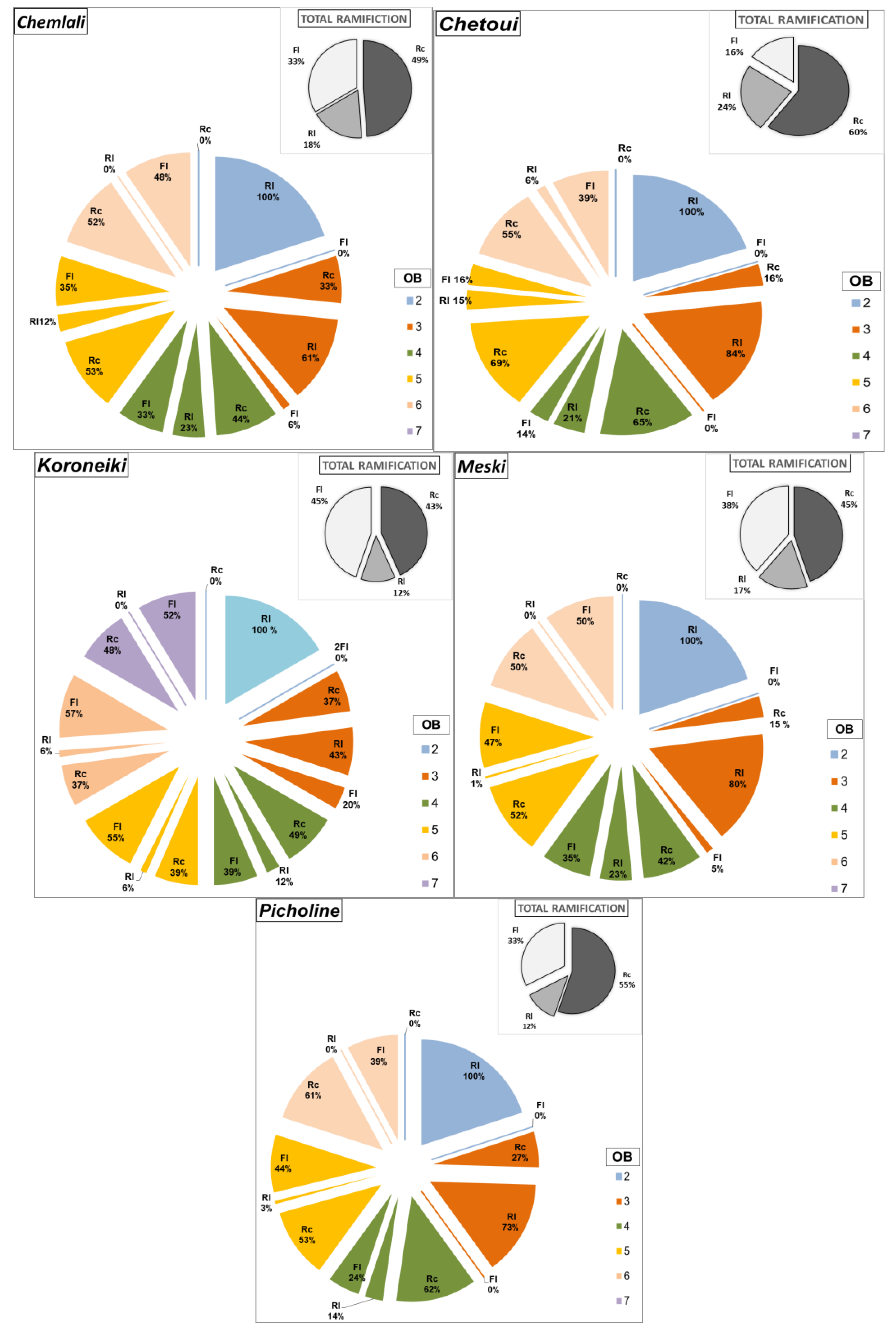
2.1.1. Evaluation of the Importance of Branching
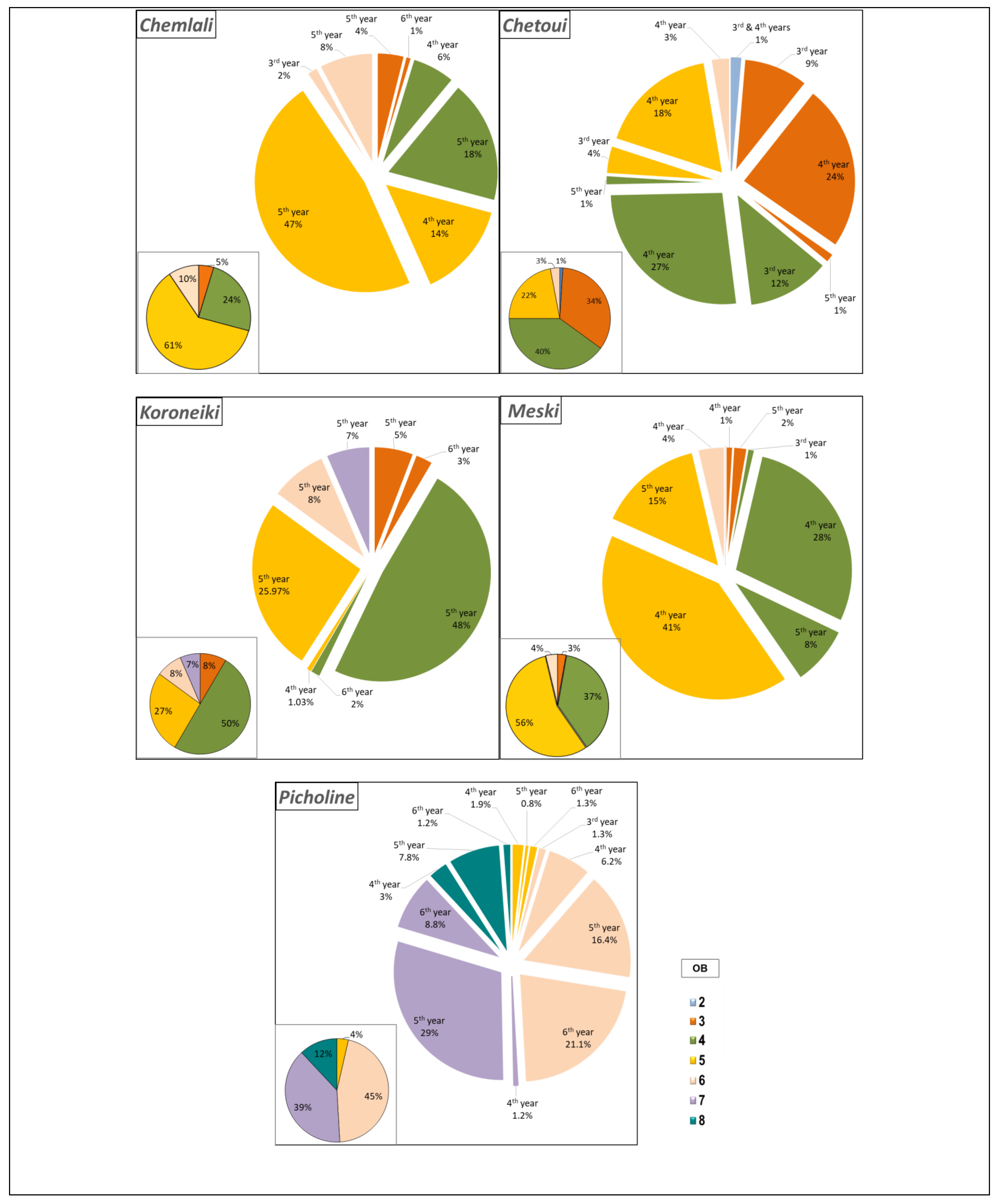
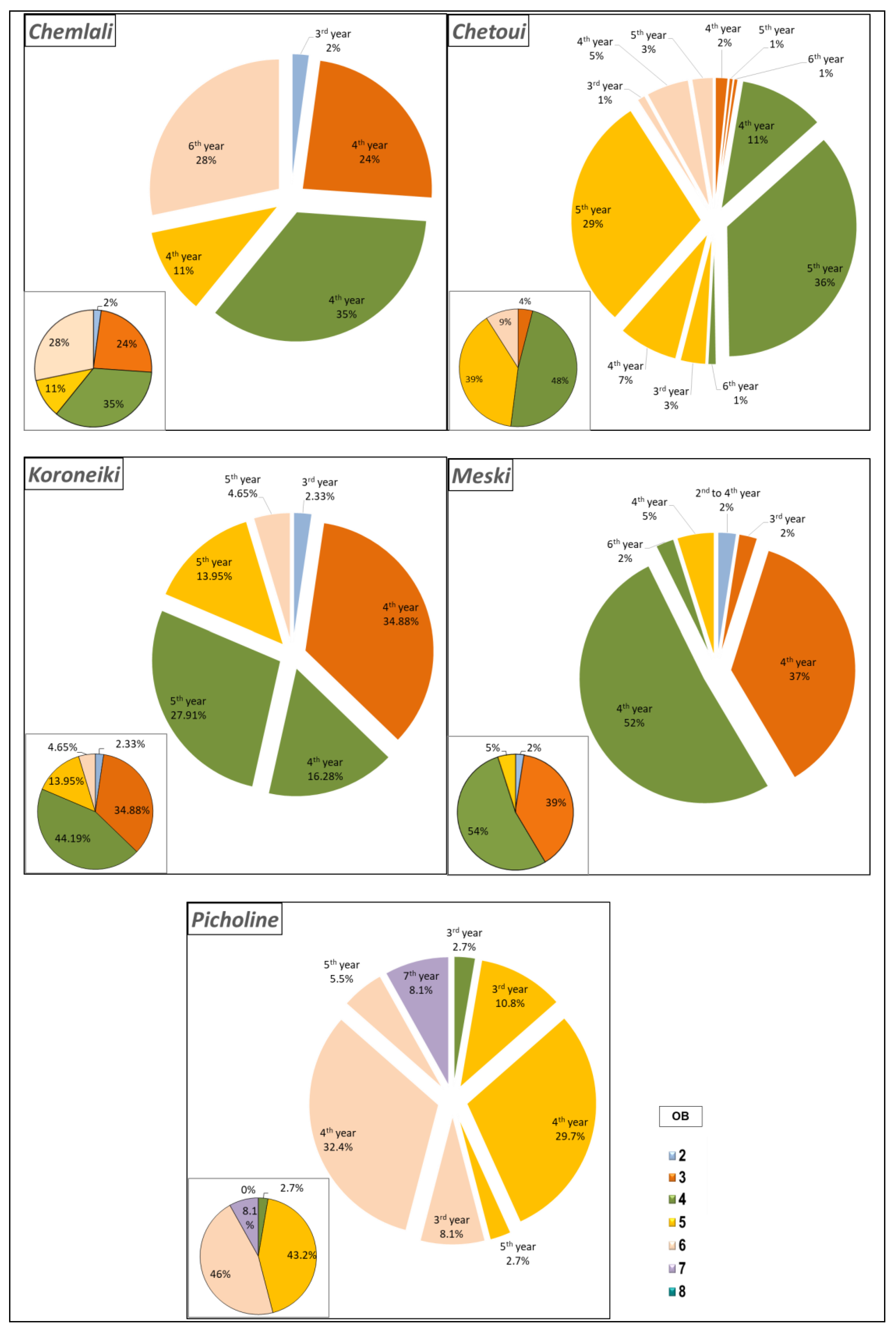
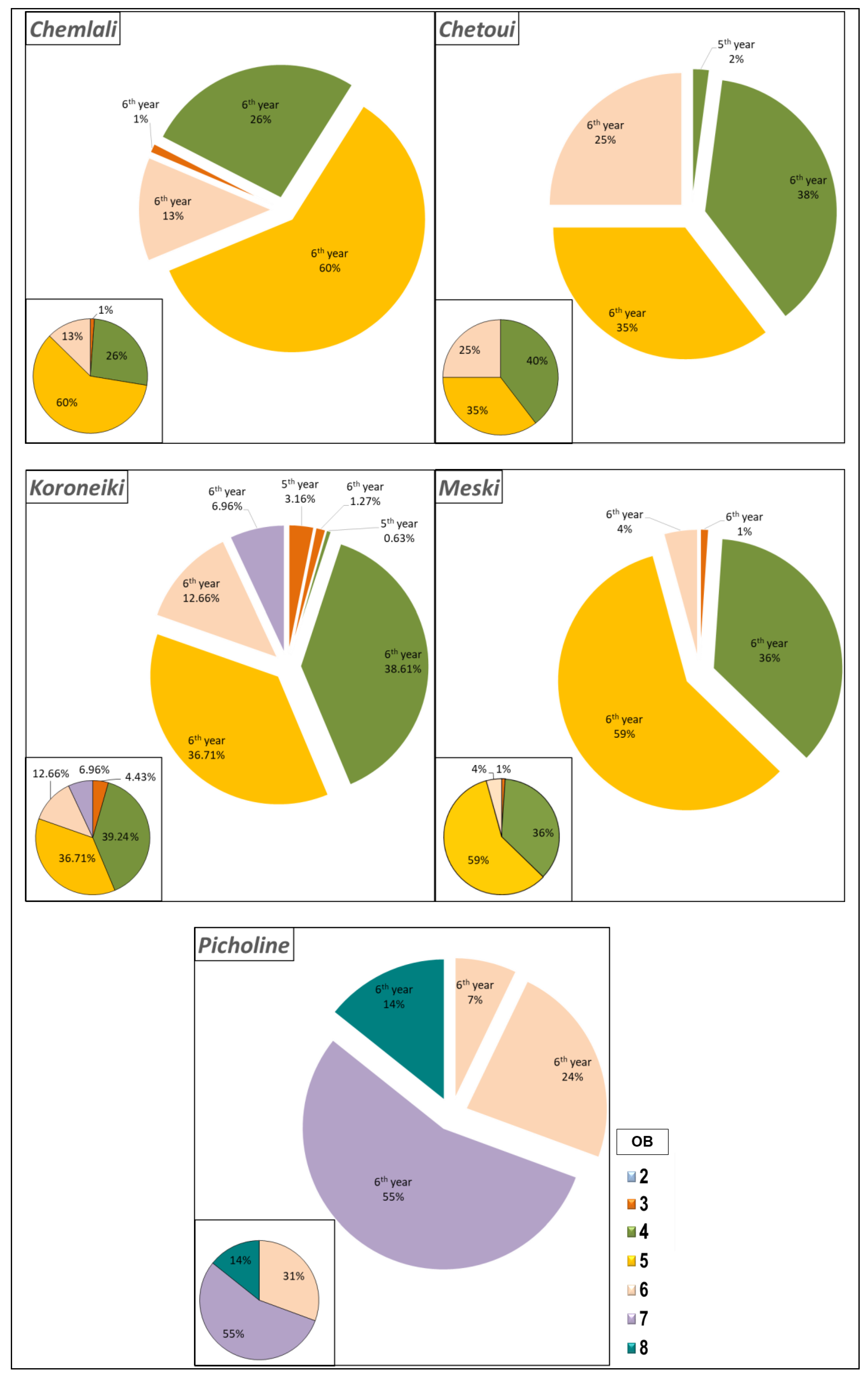
2.1.2. Estimation of Branching Density
2.1.3. Characterisation of the Shoots Carrying Architectural Units
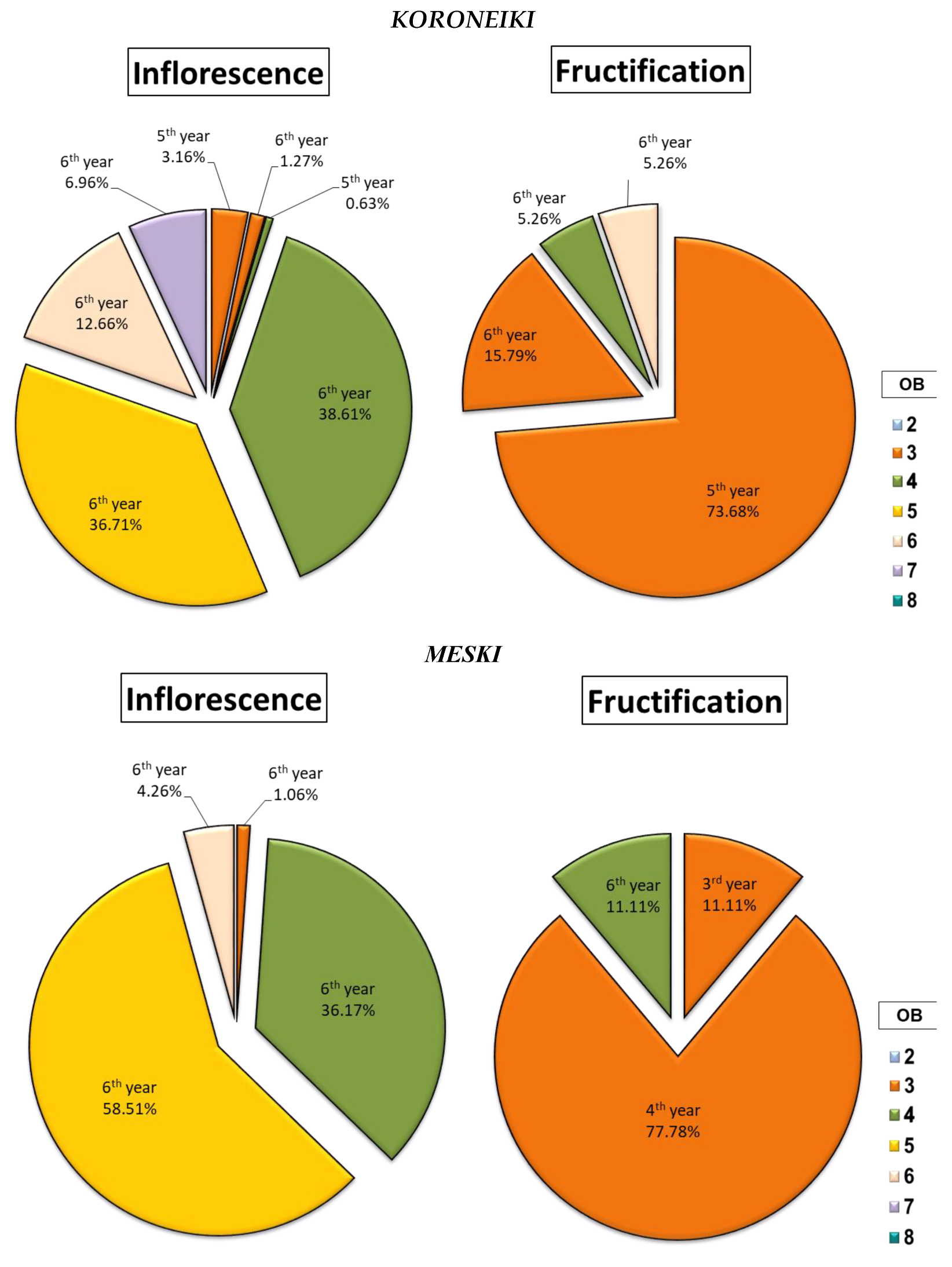
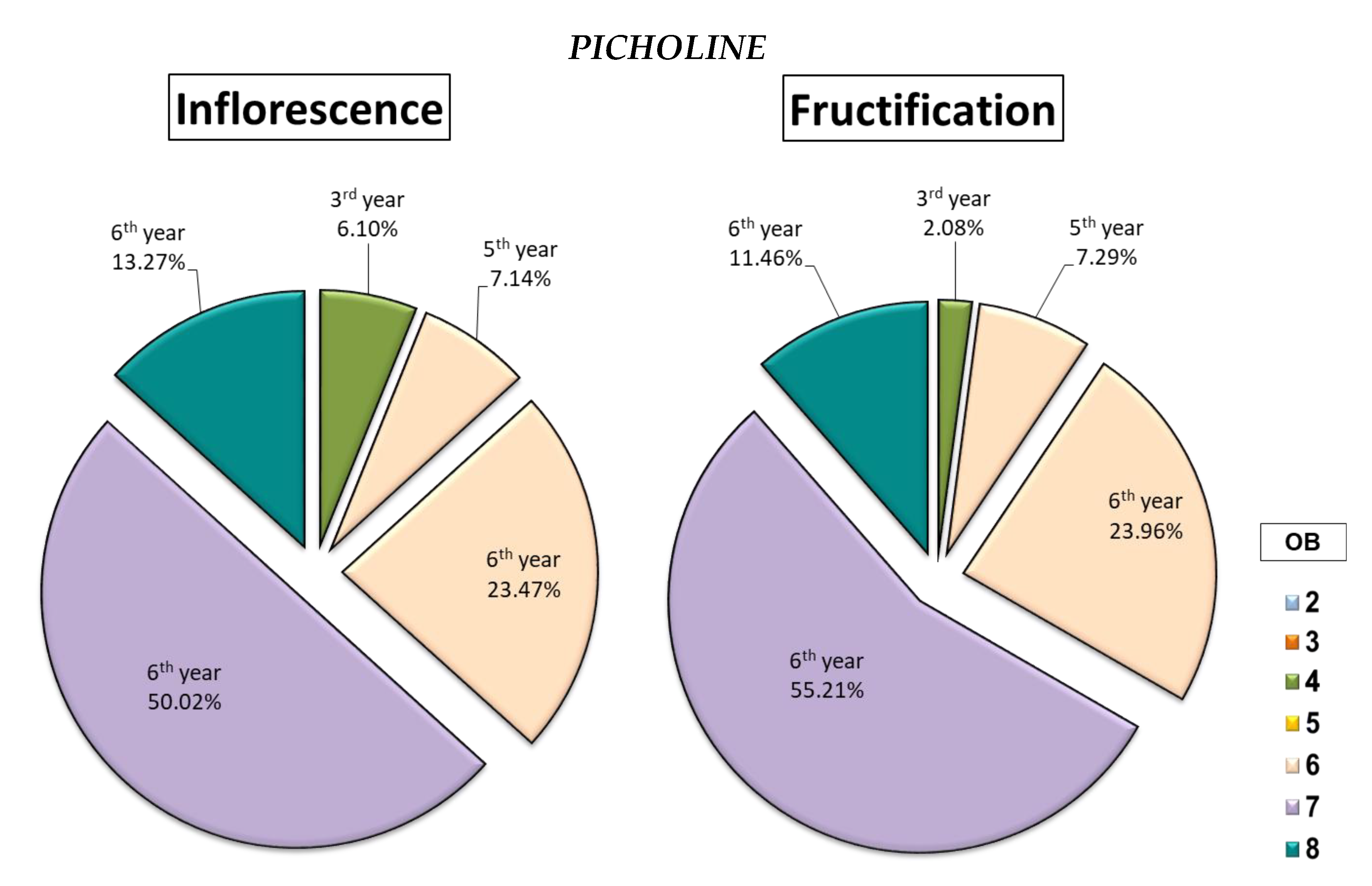
2.1.4. Characterisation of Flowering and Fruiting Sites
2.2. Biometric Characterisation of Long Shoots
2.2.1. Variation of Shoot Length
2.2.2. Variation in the Number of Internodes
2.2.3. Variation in Perennial Shoot Dimensions: Internode Length, Apex Diameter, and Basal Vigour
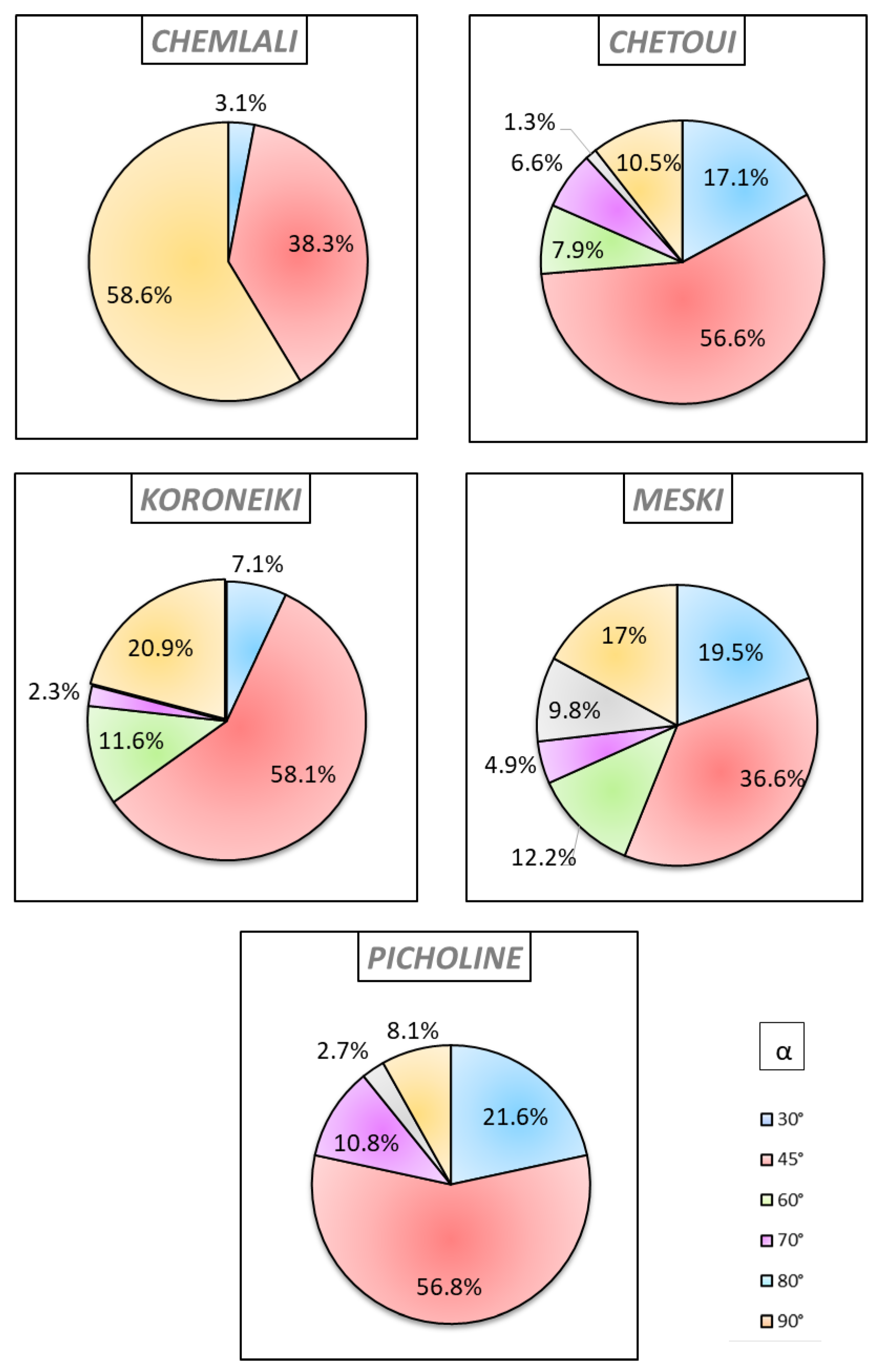
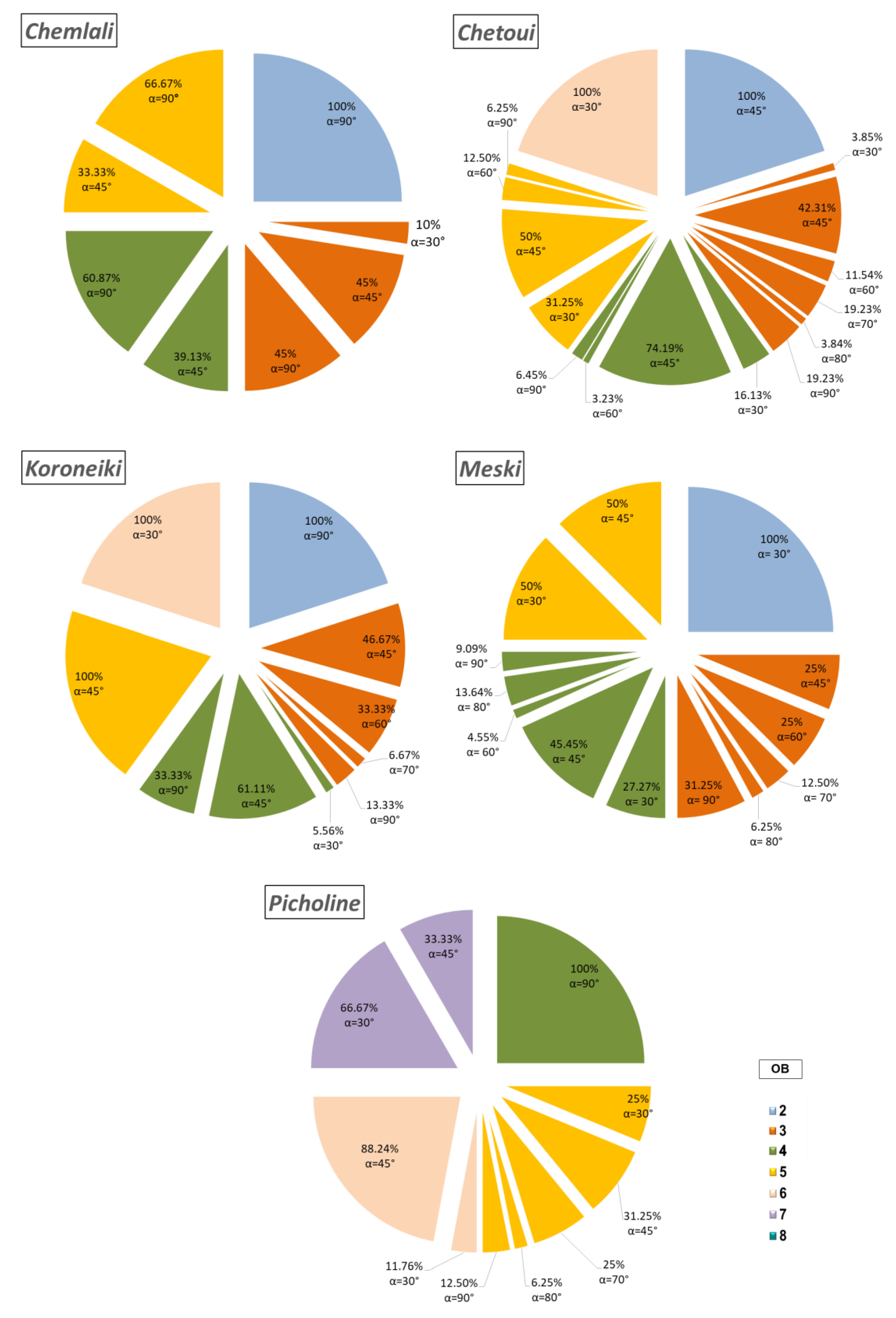
2.3. Analysis of the Geometric Architectural Variability
3. Discussion
3.1. Canopy Architecture Variability
3.2. Fruiting Characteristics
3.3. Plant Vigour and Geometric Architectural Variability
4. Materials and Methods
4.1. Plant Material and Growth Conditions
4.2. Measured Parameters
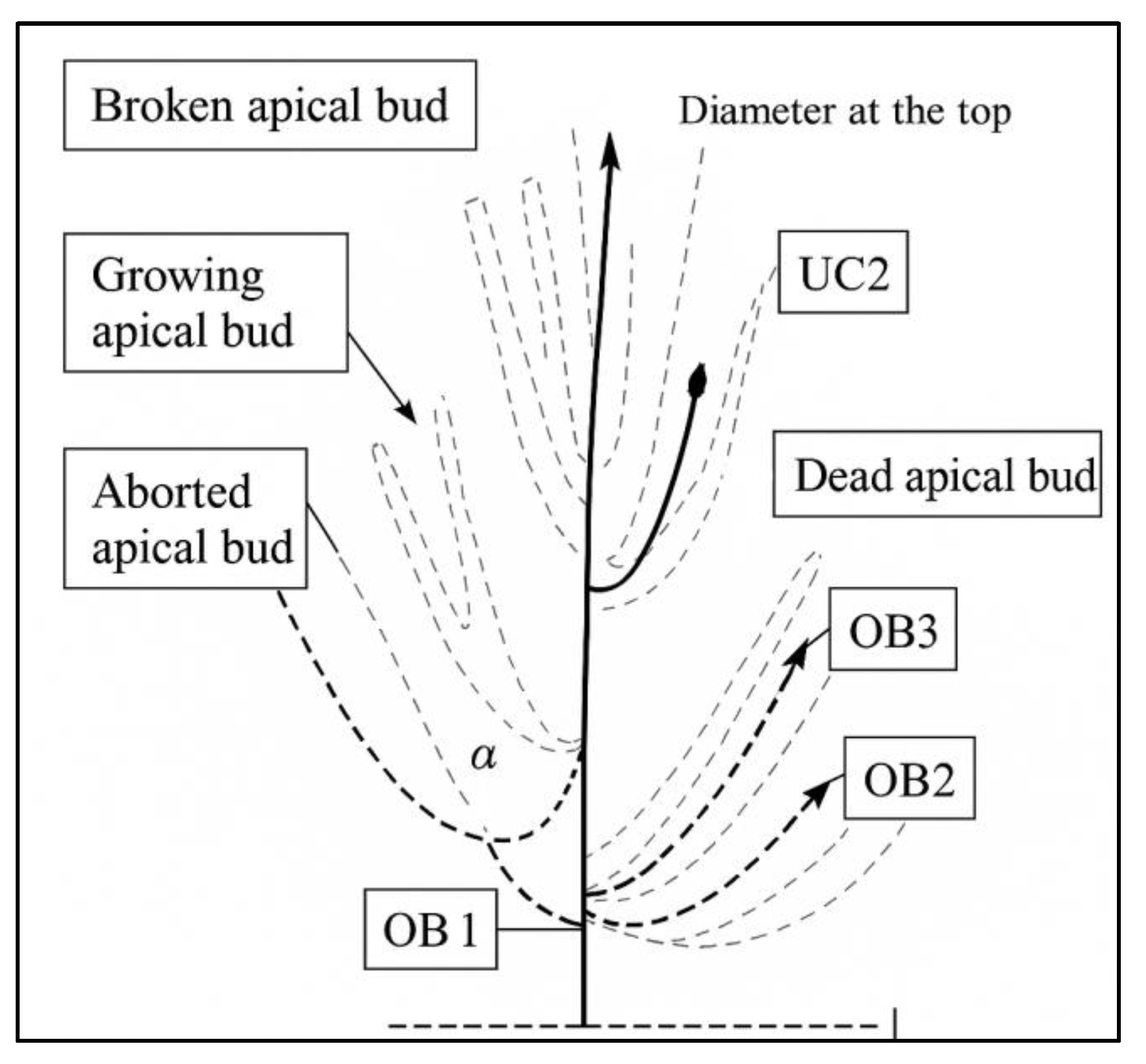
4.3. Architectural Studies
4.4. Analysis of Topographic Architectural Variability
4.5. Annual Growth Analysis
4.6. Analysis of Geometric Architectural Variability
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- International Olive Council. 2017. Available online: http://www.internationaloliveoil.org (accessed on 1 February 2018).
- De Castro, A.I.; Rallo, P.; Suárez, M.P.; Torres-Sánchez, J.; Casanova, L.; Jiménez-Brenes, F.M.; Morales-Sillero, A.; Jiménez, M.R.; López-Granados, F. High-Throughput System for the Early Quantification of Major Architectural Traits in Olive Breeding Trials Using UAV Images and OBIA Techniques. Front. Plant Sci. 2019, 10, 1472. [Google Scholar] [CrossRef] [PubMed]
- Mairech, H.; López-Bernal, Á.; Moriondo, M.; Dibari, C.; Regni, L.; Proietti, P.; Villalobos, F.J.; Testi, L. Is new olive farming sustainable? A spatial comparison of productive and environmental performances between traditional and new olive orchards with the model OliveCan. Agric. Syst. 2020, 181, 102816. [Google Scholar] [CrossRef]
- Stilliano, T.; de Luca, A.I.; Falcone, G.; Spada, E.; Gulisano, G.; Strano, A. Economic profitability assessment of mediterranean olive growing systems. Bulg. J. Agric. Sci. 2016, 22, 517–526. [Google Scholar]
- Rugini, E.; Baldoni, L.; Muleo, R.; Sebastiani, L. The Olive Tree Genome; Springer: Berlin, Germany, 2016; pp. 13–25. [Google Scholar] [CrossRef]
- Manna, M.; Rengasamy, B.; Sinha, A.K. Revisiting the role of MAPK signalling pathway in plants and its manipulation for crop improvement. Plant Cell Environ. 2023, 46, 2277–2295. [Google Scholar] [CrossRef]
- Ye, Y.J.; Wu, J.Y.; Feng, L.; Ju, Y.Q.; Cai, M.; Cheng, T.R.; Pan, H.T.; Zhang, Q.X. Heritability and gene effects for plant architecture traits of crape myrtle using major gene plus polygene inheritance analysis. Sci. Hortic. 2017, 225, 335–342. [Google Scholar] [CrossRef]
- Lodolini, E.M.; Tarragoni, A.; Cioccolanti, T.; Pollastri, L.; Neri, D. Architectural Characteristics of Six Olive Cultivars with Respect to Their Suitability for High-Density Orchards. Acta Hortic. 2017, 1160, 127–134. [Google Scholar] [CrossRef]
- Annabi, K. Diversité Architecturale de L’olivier et Réponses de L’arbre aux Manipulations Techniques. Ph.D. Thesis, University of Sousse, Sousse, Tunisia, 5 November 2020. Available online: http://www.theses.rnu.tn/fr/dynamique/fiche.php?id=6100 (accessed on 18 February 2021).
- Lodolini, E.M.; Lucchese, P.G.; de Iudicibus, A. Canopy Growth and Architecture of Five Olive Cultivars for High-Density Orchards. Acta Hortic. 2023, 1366, 275–282. [Google Scholar] [CrossRef]
- Neri, D.; Cioccolanti, T.; Zuccherelli, G.; Navacchi, O.; Giorgi, V.; Lodolini, E.M. Micropropagation effects on juvenile traits, flower differentiation, and tree architecture in young olive trees. Agronomy 2020, 10, 1742. [Google Scholar] [CrossRef]
- Carella, A.; Massenti, R.; Milazzo, G.; Caruso, T.; Lo Bianco, R. Fruiting, morphology, and architecture of ‘Arbequina’ and ‘Ca-latina’ olive branches. Horticulturae 2022, 8, 109. [Google Scholar] [CrossRef]
- Rosati, A.; Paoletti, A.; Lodolini, E.M.; Famiani, F. Cultivar ideotype for intensive olive orchards: Plant vigor, biomass partitioning, tree architecture and fruiting characteristics. Front. Plant Sci. 2024, 15, 1345182. [Google Scholar] [CrossRef]
- Hammami, S.B.M.; De la Rosa, R.; Sghaier-Hammami, B.; León, L.; Rapoport, H.F. Reliable and relevant qualitative descriptors for evaluating complex architectural traits in olive progenies. Sci. Hortic. 2012, 143, 157–166. [Google Scholar] [CrossRef]
- Catania, P.; Roma, E.; Orlando, S.; Vallone, M. Evaluation of multispectral data acquired from UAV platform in olive orchard. Horticulturae 2023, 9, 133. [Google Scholar] [CrossRef]
- Bentaher, H.; Haddar, M.; Fakhfakh, T.; Mâalej, A. Finite elements modeling of olive tree mechanical harvesting using different shakers. Trees 2013, 27, 1537–1545. [Google Scholar] [CrossRef]
- Anthony, B.M.; Minas, I.S. Optimizing peach tree canopy architecture for efficient light use, increased productivity and improved fruit quality. Agronomy 2021, 11, 1961. [Google Scholar] [CrossRef]
- Qiu, T.; Wang, T.; Han, T.; Kuehn, K.; Cheng, L.; Meng, C.; Xu, X.; Xu, K.; Yu, J. Apple QSM: Geometry-Based 3D Characterization of Apple Tree Architecture in Orchards. Plant Phenomics 2024, 6, 0179. [Google Scholar] [CrossRef]
- Sangjan, W.; Sankaran, S. Phenotyping architecture traits of tree species using remote sensing techniques. Trans. ASABE 2021, 64, 1611–1624. [Google Scholar] [CrossRef]
- Liu, J.; Zhou, Z.; Li, B. Remote sensing for field-based crop phenotyping. Front. Plant Sci. 2024, 15, 1368694. [Google Scholar] [CrossRef]
- Monasterio, R.P.; Banco, A.P.; Caderón, F.J.; Trentacoste, E.R. Effects of pre-harvest deficit irrigation during the oil accumulation period on fruit characteristics, oil yield extraction, and oil quality in olive cv. Genovesa in an arid region of Argentina. Agric. Water Manag. 2021, 252, 106901. [Google Scholar] [CrossRef]
- Catania, P.; Ferro, M.V.; Roma, E.; Orlando, S.; Vallone, M. Olive tree canopy assessment based on UAV multispectral images. In Proceedings of the 7th International Conference on Geographical Information Systems Theory, Applications and Management (GISTAM 2023), Prague, Czech Republic, 25–27 April 2023; Lecture Notes in Civil Engineering; Springer: Cham, Switzerland, 2023; p. 337. [Google Scholar] [CrossRef]
- Dhiab, A.B.; Zaier, H.; Gaaliche, B.; Fendri, M.; Ayadi, M.; Larbi, A. Vegetative growth, fruit distribution and oil quality in the canopy of olive trees grown under super high-density cropping system. Italus Hortus 2020, 27, 8295. [Google Scholar] [CrossRef]
- Belhassine, F.; Pallas, B.; Pierru-Bluy, S.; Martinez, S.; Fumey, D.; Costes, E. A genotype-specific architectural and physiological profile is involved in the flowering regularity of apple trees. Tree Physiol. 2022, 42, 2306–2318. [Google Scholar] [CrossRef]
- Ahmad, I.; Sohail, M.; Hameed, M.; Fatima, S.; Ahmad, M.S.A.; Mehmood, A.; Basharat, S.; Asghar, A.; Raza Shah, S.M.; Ahmad, K.S. Morpho-anatomical determinants of yield potential in Olea europaea L. cultivars belonging to diversified origin grown in semi-arid environments. PLoS ONE 2023, 18, e0286736. [Google Scholar] [CrossRef]
- Rosati, A.; Paoletti, A.; Caporali, S.; Perri, E. The role of tree architecture in super high density olive orchards. Sci. Hortic. 2013, 161, 24–29. [Google Scholar] [CrossRef]
- Coupel-Ledru, A.; Pallas, B.; Delalande, M.; Segura, V.; Guitton, B.; Muranty, H.; Costes, E. Tree architecture, light interception and water-use related traits are controlled by different genomic regions in an apple tree core collection. New Phytol. 2022, 234, 209–226. [Google Scholar] [CrossRef] [PubMed]
- Annabi, K.; Laaribi, I.; Gouta, H.; Laabidi, F.; Mechri, B.; Ajmi, L.; Zouaria, I.; Muscolo, A.; Panuccio, M.R.; Mezghani Aïachia, M. Protein content, antioxidant activity, carbohydrates and photosynthesis in leaves of girdled stems of four olive cultivars. Sci. Hortic. 2019, 256, 108551. [Google Scholar] [CrossRef]
- Gaaliche, B.; Saddoud, O.; Mars, M. Morphological and pomological diversity of fig (Ficus carica L.) cultivars in northwest Tunisia. Int. Sch. Res. Not. 2012, 2012, 326461. [Google Scholar] [CrossRef]
- Sobreiro, J.; Patanita, M.I.; Patanita, M.; Tomaz, A. Sustainability of high-density olive orchards: Hints for irrigation management and agroecological approaches. Water 2023, 15, 2486. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemlali | Quantitative Parameters | |||||
|---|---|---|---|---|---|---|
| OB | Year | Lg | Nbre E.N. | Diam B | Diam S | L.E.N. |
| 2 | 3 | 72 ± 0.00 a | 41 ± 0.0 a | 20.6 ± 0.0 a | 2 ± 0.0 c | 1.76 ± 0.0 a |
| 3 | 4 | 21.2 ± 9.17 c | 13.45 ± 1.4 d | 5.17 ± 2 c | 2.9 ± 1.58 b | 1.71 ± 0.7 b |
| 4 | 4 | 25.75 ± 6.1 b | 14.94 ± 8.0 b | 7.58 ± 1 b | 14.7 ± 5.2 a | 1.65 ± 0.4 c |
| 5 | 4 | 19.5 ± 3.11 d | 14.25 ± 2 c | 2.65 ± 1 d | 0.98 ± 0.05 d | 1.4 ± 0.08 d |
| Average | 24.84 ± 8.04 | 15.16 ± 1.10 | 6.54 ± 0.78 | 8.53 ± 3.64 | 1.64 ± 0.49 | |
| Meski | Quantitative Parameters | |||||
|---|---|---|---|---|---|---|
| OB | Year | Lg | Nbre E.N. | Diam B | Diam S | L.E.N. |
| 2 | 4 | 78 ± 0.0 a | 50 ± 0.0 a | 18.5 ± 0.0 a | 1.5 ± 0.0 c | 1.56 ± 0.0 a |
| 3 | 24.31 ± 3.71 c | 22.13 ± 1.23 c | 5.41 ± 1.81 c | 1.93 ± 0.13 c | 1.11 ± 0.24 c | |
| 3 | 3 | 25 ± 0.0 | 22 ± 0.0 | 5.8 ± 0.0 | 3.9 ± 0.0 | 1.14 ± 0.0 |
| 3 | 4 | 24.27 ± 4.18 | 22.13 ± 12.74 | 5.38 ± 1.87 | 1.79 ± 0.29 | 1.11 ± 0.25 |
| 4 | 21.14 ± 9.40 d | 19.14 ± 8.34 d | 5.89 ± 0.57 b | 1.52 ± 0.74 b | 1.17 ± 0.32 b | |
| 4 | 4 | 20.47 ± 9.10 | 18.71 ± 8.30 | 5.94 ± 0.84 | 1.55 ± 0.74 | 1.17 ± 0.33 |
| 4 | 6 | 35 ± 0.0 | 28 ± 0.0 | 4.8 ± 0.0 | 0.9 ± 0.0 | 1.25 ± 0.0 |
| 5 | 4 | 24.5 ± 7.78 b | 23.5 ± 4.95 b | 3.8 ± 0.71 d | 0.9 ± 0.0 d | 1.03 ± 0.11 d |
| Average | 23.93 ± 3.99 | 21.27 ± 1.08 | 5.90 ± 0.47 | 1.65 ± 1.02 | 1.15 ± 0.29 | |
| Chetoui | Quantitative Parameters | |||||
|---|---|---|---|---|---|---|
| OB | Year | Lg | Nbre E.N. | Diam B | Diam S | L.E.N. |
| 2 | 80 ± 0.0 a | 55 ± 0.0 a | 17.6 ± 0.0 a | 1.5 ± 0.0 b | 1.45 ± 0.0 a | |
| 2 | 3_4 | 80 ± 0.0 | 55 ± 0.0 | 17.6 ± 0.0 | 1.5 ± 0.0 | 1.45 ± 0.0 |
| 3 | 27.37 ± 6.96 b | 19.96 ± 1.13 b | 4.17 ± 1.65 b | 1.55 ± 0.94 a | 1.43 ± 0.54 b | |
| 3 | 3 | 26.57 ± 9.6 | 14.86 ± 1.14 | 4.11 ± 2.27 | 1.46 ± 0.12 | 1.82 ± 0.52 |
| 3 | 4 | 28.28 ± 6.65 | 22.67 ± 1.07 | 4.28 ± 1.42 | 1.61 ± 0.89 | 1.24 ± 0.44 |
| 3 | 5 | 15 ± 0.0 | 7 ± 0.0 | 2.6 ± 0.0 | 1.2 ± 0.0 | 2.14 ± 0.00 |
| 4 | 20.85 ± 7.81 c | 17.81 ± 6.28 c | 3.4 ± 0.91 b | 1.29 ± 0.7 c | 1.22 ± 0.42 d | |
| 4 | 3 | 21.61 ± 1.28 | 18.78 ± 7.03 | 3.61 ± 0.87 | 1.51 ± 0.90 | 1.15 ± 0.41 |
| 4 | 4 | 20.4 ± 6.28 | 17.6 ± 6.26 | 3.33 ± 0.97 | 1.22 ± 0.6 | 1.22 ± 0.38 |
| 4 | 5 | 22 ± 7.07 | 15.5 ± 4.95 | 3.2 ± 0.28 | 0.95 ± 0.07 | 1.57 ± 0.96 |
| 5 | 14.97 ± 8.11 d | 12.94 ± 3.87 d | 2.65 ± 0.56 b | 1.06 ± 0.49 d | 1.25 ± 0.78 c | |
| 5 | 3 | 10 ± 3.61 | 13 ± 1.73 | 2.82 ± 0.59 | 0.93 ± 0.41 | 0.80 ± 0.38 |
| 5 | 4 | 16.12 ± 8.51 | 12.92 ± 4.27 | 2.61 ± 0.57 | 1.09 ± 0.52 | 1.35 ± 0.82 |
| 6 | 4 | 6.25 ± 1.06 e | 11.5 ± 2.12 e | 1.66 ± 0.04 c | 0.86 ± 0.0 e | 0.54 ± 0.01 e |
| Average e | 22.22 ± 4.34 | 17.84 ± 9.38 | 3.65 ± 2.08 | 1.32 ± 0.75 | 1.28 ± 0.56 | |
| Picholine | Quantitative Parameters | |||||
|---|---|---|---|---|---|---|
| OB | Year | Lg | Nbre E.N. | Diam B | Diam S | L.E.N. |
| 4 | 3 | 39 ± 0.0 b | 32 ± 0.0 a | 14 ± 0.0 a | 1.4 ± 0.0 b | 1.22 ± 0.0 a |
| 5 | 43.38 ± 6.30 a | 20.25 ± 7.98 c | 4.31 ± 1.68 a | 1.56 ± 0.13 a | 1.95 ± 1.20 a | |
| 5 | 3 | 73.75 ± 5.16 | 24 ± 6.16 | 4.35 ± 1.66 | 1.1 ± 0.20 | 2.99 ± 1.09 |
| 5 | 4 | 34.64 ± 2.56 | 19.45 ± 8.59 | 4.27 ± 1.85 | 1.77 ± 1.53 | 1.63 ± 0.61 |
| 5 | 5 | 18 ± 0.0 | 14 ± 0.0 | 4.6 ± 0.0 | 1 ± 0.0 | 1.29 ± 0.0 |
| 6 | 18.5 ± 1.03 d | 15.82 ± 5.10 d | 3.61 ± 0.94 a | 1.11 ± 0.3 c | 1.23 ± 0.40 a | |
| 6 | 3 | 18.67 ± 8.50 | 19.66 ± 5.03 | 3.93 ± 0.40 | 1 ± 0.0 | 0.98 ± 0.05 |
| 6 | 4 | 20.25 ± 1.27 | 15.08 ± 5.21 | 3.65 ± 1.05 | 1.13 ± 0.36 | 1.31 ± 0.37 |
| 6 | 5 | 17 ± 1.13 | 14.5 ± 3.54 | 2.85 ± 0.35 | 1.1 ± 0.14 | 1.11 ± 0.51 |
| 7 | 4 | 29.33 ± 2.52 c | 22 ± 3.46 b | 3.53 ± 0.25 a | 1 ± 0.0 d | 1.34 ± 0.11 a |
| Average | 30.37 ± 6.97 | 18.67 ± 7.04 | 4.19 ± 2.11 | 1.3 ± 0.89 | 5.74 ± 0.22 | |
| Koroneiki | Quantitative Parameters | |||||
|---|---|---|---|---|---|---|
| OB | Year | Lg | Nbre E.N. | Diam B | Diam S | L.E.N. |
| 2 | 3 | 100 ± 0.0 a | 46 ± 0.0 a | 24 ± 0.0 a | 1.2 ± 0.0 c | 2.17 ± 0.0 a |
| 3 | 4 | 48.47 ± 5.62 b | 23.53 ± 11.5 b | 5.77± 2.58 c | 1.79 ± 1.42 b | 2.13 ± 0.76 a |
| 4 | 23.75 ± 9.31 d | 15.44 ± 7.06 d | 0.68 ± 0.0 e | 1.09 ± 0.32 d | 1.69 ± 0.74 a | |
| 4 | 4 | 23.67 ± 8.78 | 16 ± 1.09 | 3.77 ± 0.71 | 1.32 ± 0.49 | 1.90 ± 1.08 |
| 4 | 5 | 23.79 ± 9.94 | 15.17 ± 4.69 | 3.18 ± 0.62 | 0.98 ± 0.05 | 1.58 ± 0.53 |
| 5 | 19 ± 8.66 e | 13.43 ± 7.59 e | 8.89 ± 1.46 b | 5.97 ± 1.32 a | 1.49 ± 0.54 a | |
| 5 | 4 | 2 ± 0.00 | 2 ± 0.0 | 42 ± 0.0 | 36 ± 0.0 | 1 ± 0.0 |
| 5 | 5 | 21.83 ± 4.75 | 15.33 ± 6.22 | 3.37 ± 0.59 | 0.97 ± 0.08 | 1.58 ± 0.54 |
| 6 | 5 | 27 ± 4.24 c | 16 ± 0.0 c | 3.5 ± 0.0 d | 1 ± 0.0 e | 1.69 ± 0.27 a |
| Average | 33.52 ± 2.98 | 18.67 ± 1.04 | 5.60 ± 0.67 | 2.13 ± 0.53 | 1.82 ± 0.27 | |
| MAIN AXIS | ORDRE 2 | ORDRE3 | ORDRE | QUALITATIVE PARAMETERS | QUANTITATIVE PARAMETRES | |||||||||||||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Exp | Var | n° Arbre | Treat | OB-T | UC1 | O1-ON | OB- U | nd br B2 | nd br S2 | OB-T | UC2 | O2-ON | OB- U | nd br B3 | nd br S3 | OB | UC3 | O3-ON | OB- U | UC | Year | OB-T | OB-U | NAT | State of the Bud | Nb. Fruit | lg | Nbre EN | diam B | diam S | Ang/Insert | LEN |
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 100,000 | 1 | UC13 | 3 | 4 | 1 | 2 | 2 | 97 | 39 | 32 | 14 | 1.4 | 90 | 1.22 | ||||||||||||
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 120,000 | 1 | 12 | 21 | 5 | UC24 | 20,000 | 2 | UC24 | 4 | 5 | 2 | 2 | 2 | 17 | 13 | 2.4 | 0.9 | 80 | 1.31 | |||||||
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 120,000 | 1 | 12 | 21 | 5 | UC26 | 20,000 | 2 | UC26 | 6 | 5 | 2 | 1 | 3 | |||||||||||||
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 120,000 | 1 | 13 | 20 | 5 | UC24 | 20,000 | 2 | UC24 | 4 | 5 | 2 | 2 | 1 | 38 | 16 | 7.4 | 5.7 | 70 | 2.38 | |||||||
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 123,000 | 1 | 13 | 20 | 5 | UC24 | 23,000 | 2 | 2 | 15 | 6 | UC35 | 3000 | 3 | UC35 | 5 | 6 | 3 | 1 | 2 | |||||||
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 123,000 | 1 | 13 | 20 | 5 | UC24 | 23,000 | 2 | 2 | 15 | 6 | UC36 | 3000 | 3 | UC36 | 6 | 6 | 3 | 3 | 2 | |||||||
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 123,000 | 1 | 13 | 20 | 5 | UC24 | 23,000 | 2 | 5 | 12 | 6 | UC35 | 3000 | 3 | UC35 | 5 | 6 | 3 | 1 | 2 | |||||||
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 123,000 | 1 | 13 | 20 | 5 | UC24 | 23,000 | 2 | 5 | 12 | 6 | UC36 | 3000 | 3 | UC36 | 6 | 6 | 3 | 1 | 3 | |||||||
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 123,000 | 1 | 13 | 20 | 5 | UC24 | 23,000 | 2 | 13 | 4 | 6 | UC34 | 3000 | 3 | UC34 | 6 | 6 | 3 | 1 | 2 | |||||||
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 123,000 | 1 | 13 | 20 | 5 | UC24 | 23,000 | 2 | 13 | 4 | 6 | UC36 | 3000 | 3 | UC36 | 6 | 6 | 3 | 1 | 3 | |||||||
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 123,000 | 1 | 13 | 20 | 5 | UC24 | 23,000 | 2 | 16 | 1 | 6 | UC34 | 3000 | 3 | UC34 | 4 | 6 | 3 | 2 | 2 | 13 | 12 | 3.9 | 1 | 30 | 1.09 | |
| ettaoues | 1 | 15 | 3.1 | 4 | UC13 | 123,400 | 1 | 13 | 20 | 5 | UC24 | 23,400 | 2 | 16 | 1 | 6 | UC34 | 3400 | 3 | UC44 | 4 | 7 | 4 | 2 | 2 | 27 | 20 | 3.3 | 1 | 30 | 1.35 | |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Annabi, K.; Haouala, F.; Hamrita, A.; Kouki, R.; Laabidi, F.; Rejili, M.; Akef Bziouech, S.; Mezghani Aïachi, M. Architectural Analysis for Novel Olive Crop Management. Plants 2025, 14, 1707. https://doi.org/10.3390/plants14111707
Annabi K, Haouala F, Hamrita A, Kouki R, Laabidi F, Rejili M, Akef Bziouech S, Mezghani Aïachi M. Architectural Analysis for Novel Olive Crop Management. Plants. 2025; 14(11):1707. https://doi.org/10.3390/plants14111707
Chicago/Turabian StyleAnnabi, Khouloud, Faouzi Haouala, AbdelKarim Hamrita, Rania Kouki, Foued Laabidi, Mokhtar Rejili, Samra Akef Bziouech, and Mouna Mezghani Aïachi. 2025. "Architectural Analysis for Novel Olive Crop Management" Plants 14, no. 11: 1707. https://doi.org/10.3390/plants14111707
APA StyleAnnabi, K., Haouala, F., Hamrita, A., Kouki, R., Laabidi, F., Rejili, M., Akef Bziouech, S., & Mezghani Aïachi, M. (2025). Architectural Analysis for Novel Olive Crop Management. Plants, 14(11), 1707. https://doi.org/10.3390/plants14111707