

Low NaCl Concentrations Increase Cotyledon Growth in Chinese White Radish (Raphanus sativus L. var. longipinnatus Bailey) Seedlings via Aquaporin-Mediated Water Transport


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
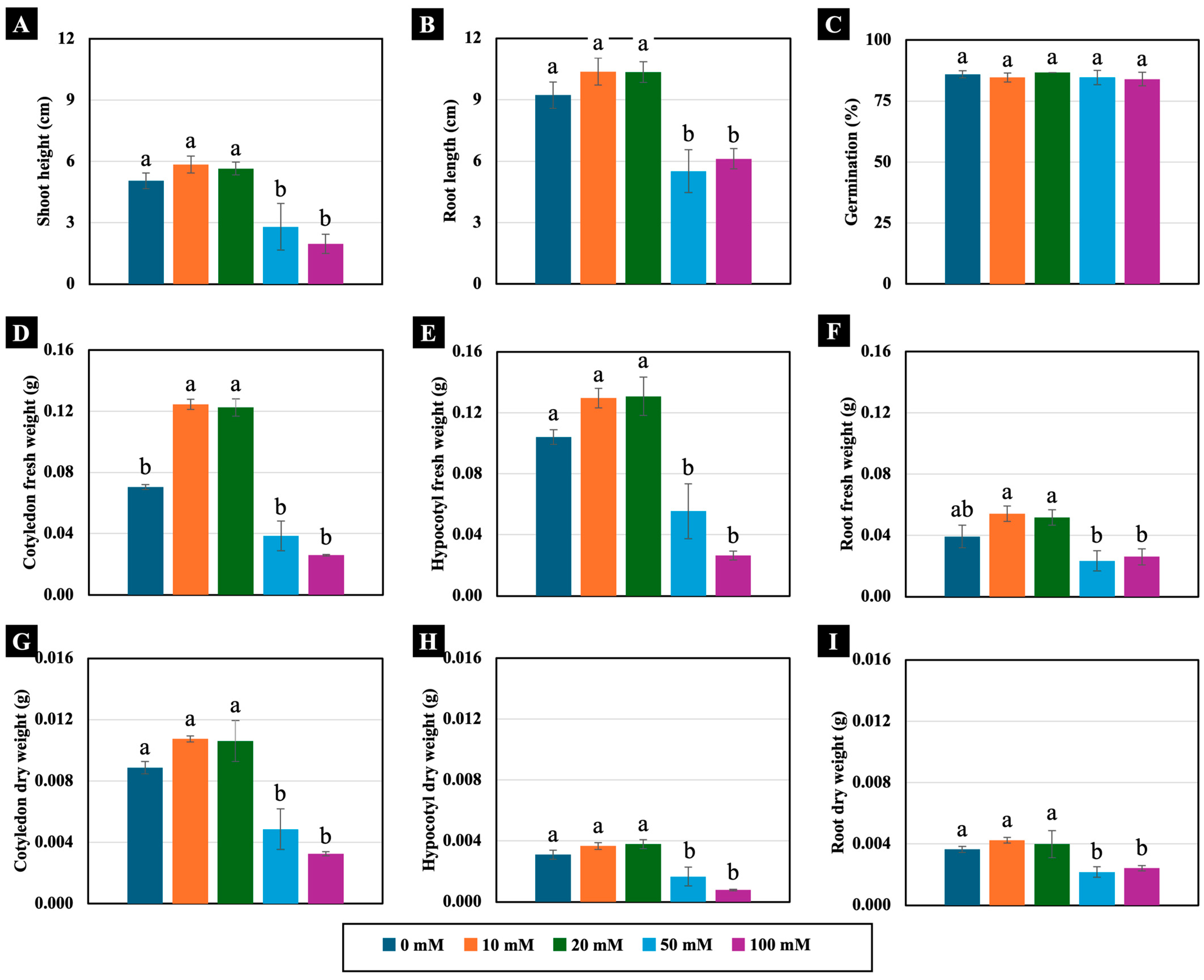
2.1. Effects of Low to High NaCl Concentrations on Growth of Chinese White Radish Seedlings
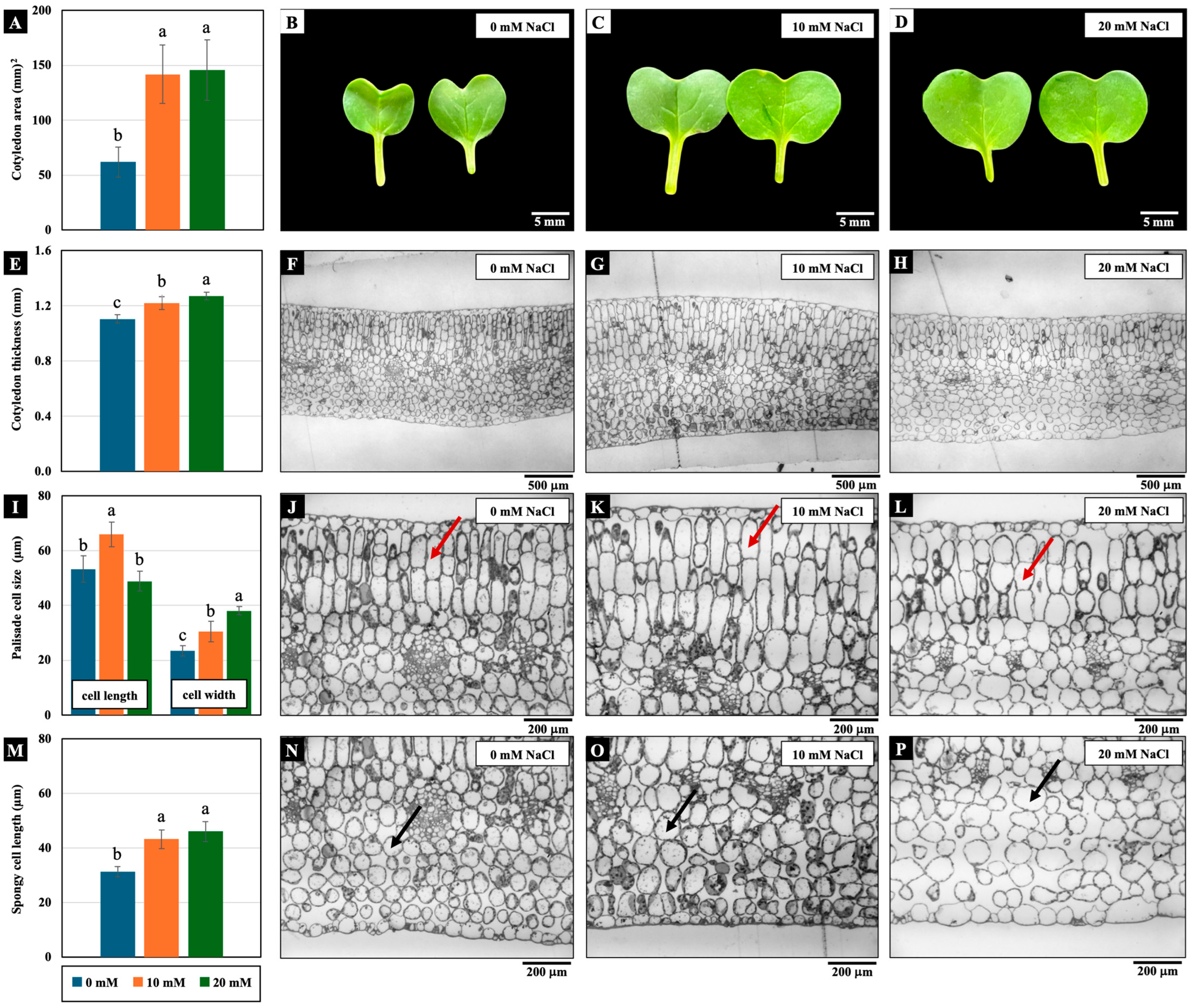
2.2. Effects of Low NaCl Concentrations on Structure and Function of Chinese White Radish Cotyledons
2.2.1. Growth Parameters
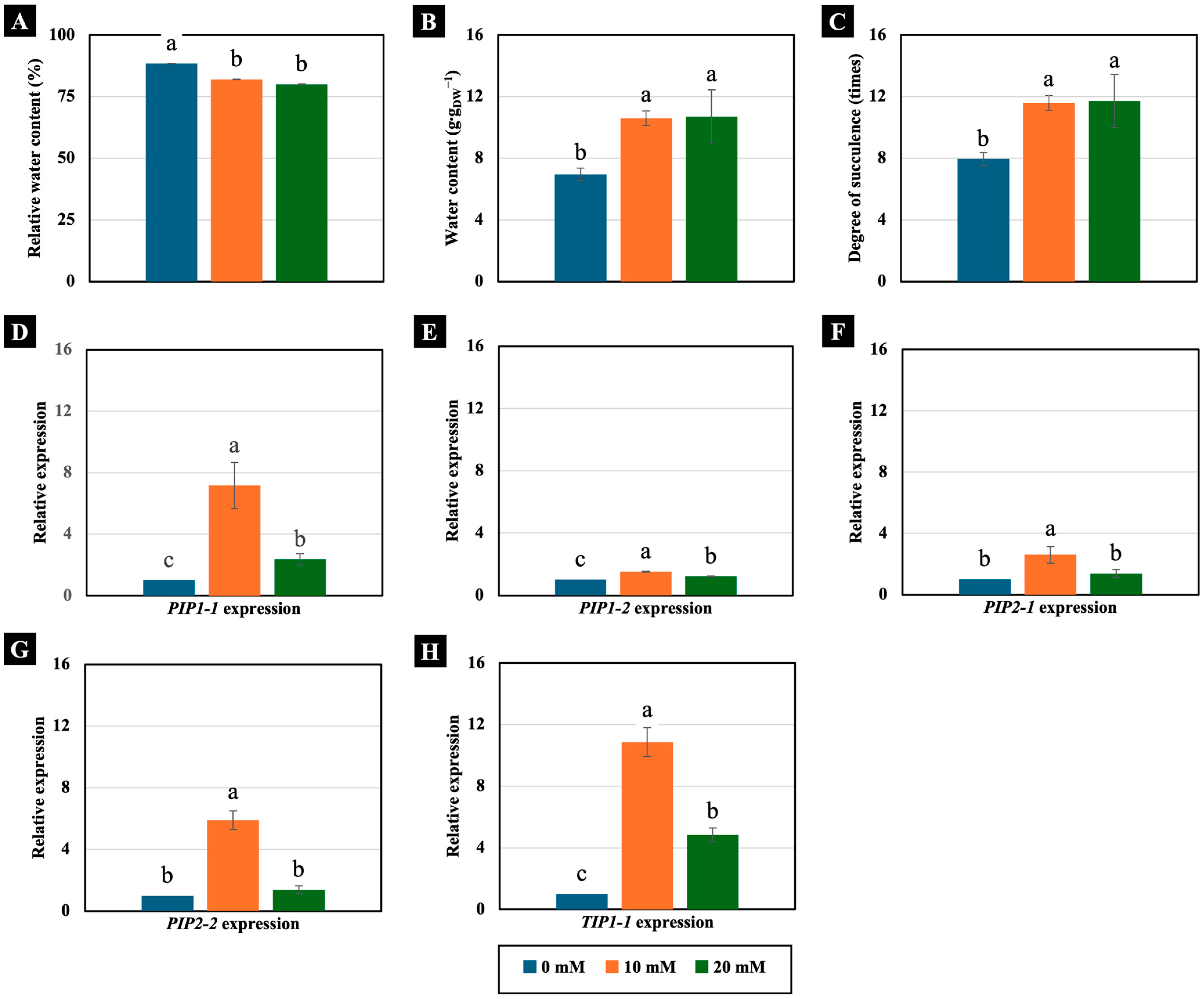
2.2.2. Water Status and Aquaporin Gene Expression
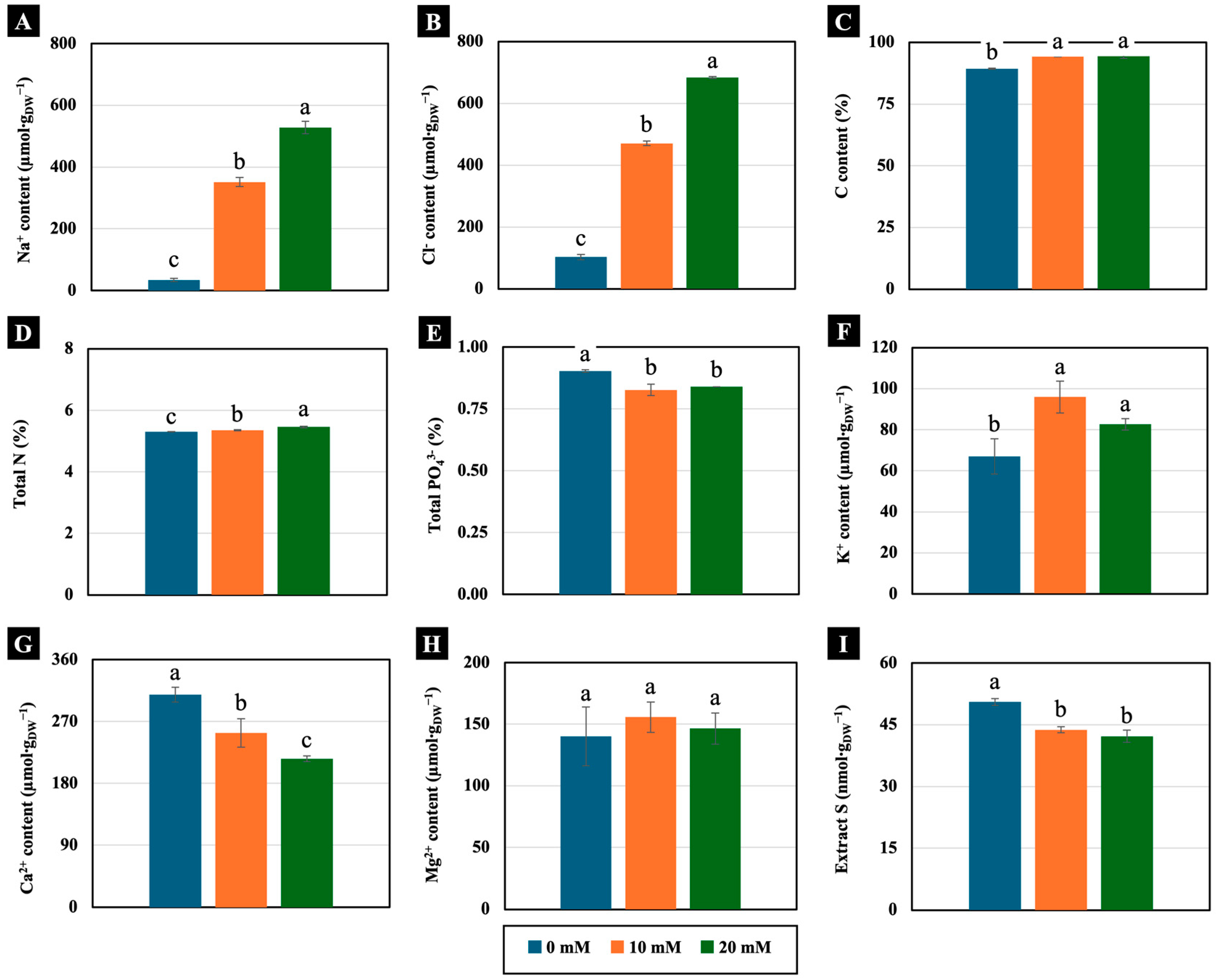
2.2.3. Ion Contents
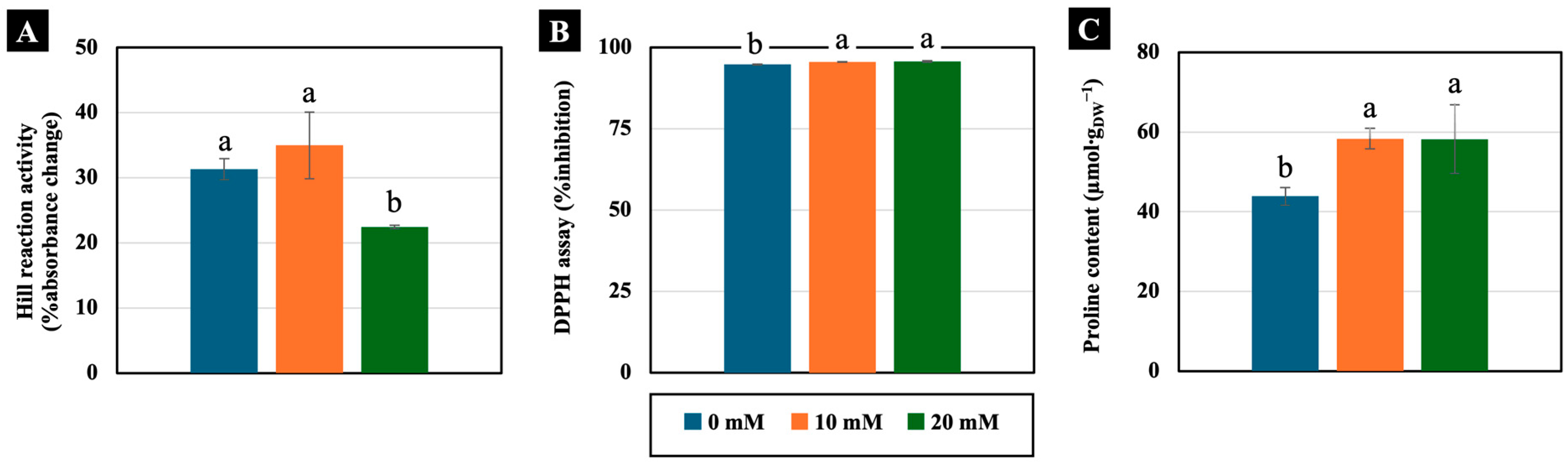
2.2.4. Physiology-Related Parameters
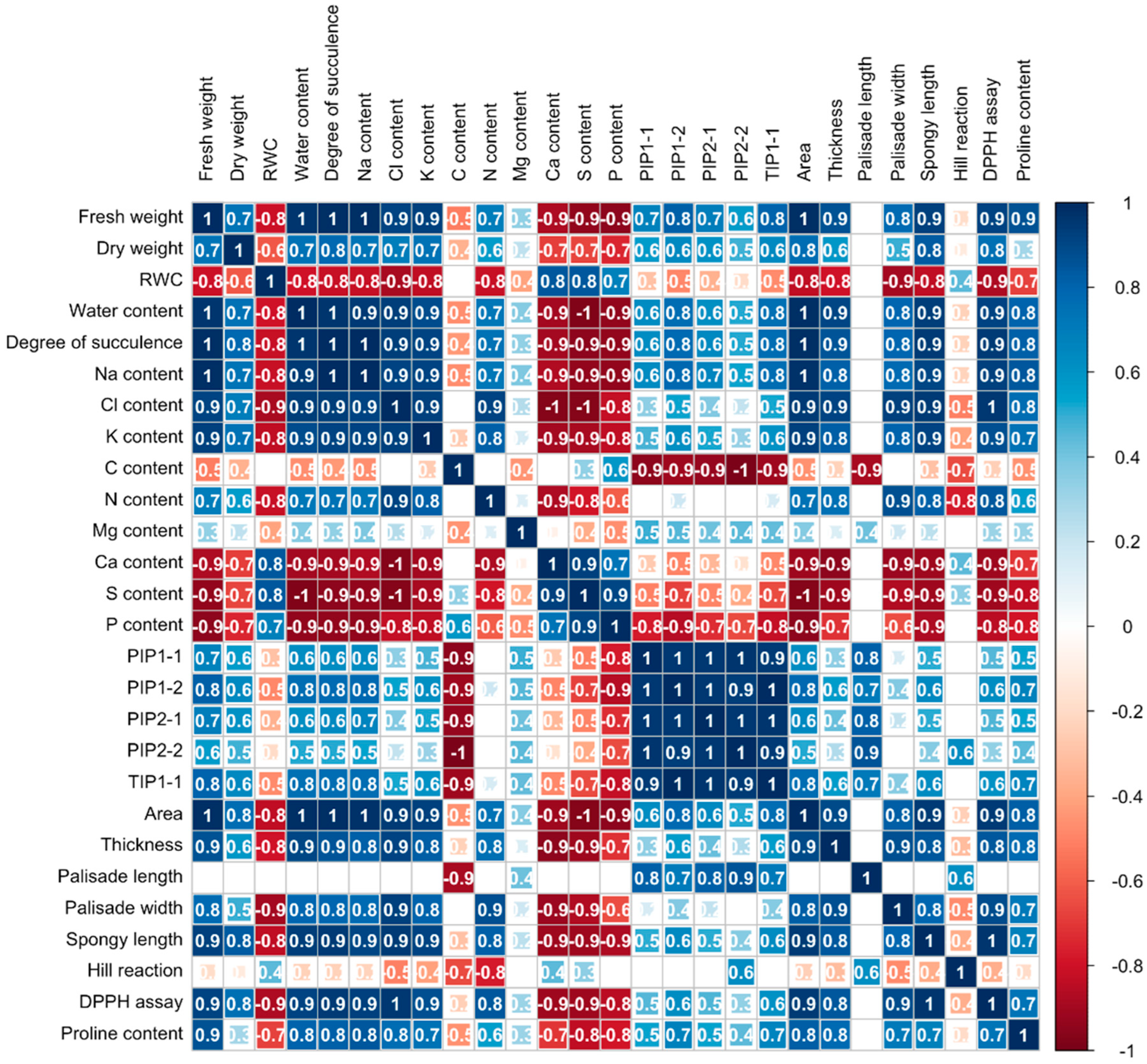
2.2.5. Correlation Coefficients Among Cotyledon Parameters Under Low NaCl Concentrations
3. Discussion
4. Materials and Methods
4.1. Plant Material and NaCl Treatments
4.2. Measurement of Growth Parameters
4.3. Measurement of Water Status Parameters
4.4. Measurement of Aquaporin Gene Expression
- ΔCt (Delta Ct) was obtained by subtracting the Ct (Cycle threshold) value of the reference gene from the Ct value of the target gene in each sample: ΔCt = CtAQPs − CtGADPH
- ΔΔCt (Delta Delta Ct) was calculated by subtracting the ΔCt of the control group from the ΔCt of the treatment group for each target gene: ΔΔCt = ΔCttreatment − ΔCtcontrol
- The relative expression level of each gene was determined by the following equation: relative expression = 2−ΔΔCt
4.5. Measurement of Ion Contents
4.6. Measurement of Physiology-Related Parameters
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ouerghi, Z.; Cornic, G.; Roudani, M.; Ayadi, A.; Brulfert, J. Effect of NaCI on photosynthesis of two wheat species (Triticum durum and T. aestivum) differing in their sensitivity to salt stress. J. Plant Physiol. 2000, 156, 335–340. [Google Scholar] [CrossRef]
- Hongqiao, L.; Suyama, A.; Mitani-Ueno, N.; Hell, R.; Maruyama-Nakashita, A. A low level of NaCl stimulates plant growth by improving carbon and sulfur assimilation in Arabidopsis thaliana. Plants 2021, 10, 2138. [Google Scholar] [CrossRef] [PubMed]
- Wakeel, A.; Steffens, D.; Schubert, S. Potassium substitution by sodium in sugar beet (Beta vulgaris) nutrition on K-fixing soils. J. Plant Nutr. Soil Sci. 2010, 173, 127–134. [Google Scholar] [CrossRef]
- Franco-Navarro, J.D.; Brumós, J.; Rosales, M.A.; Cubero-Font, P.; Talón, M.; Colmenero-Flores, J.M. Chloride regulates leaf cell size and water relations in tobacco plants. J. Exp. Bot. 2016, 67, 873–891. [Google Scholar] [CrossRef]
- Cui, Y.N.; Li, X.T.; Yuan, J.Z.; Wang, F.Z.; Guo, H.; Xia, Z.R.; Wang, S.M.; Ma, Q. Chloride is beneficial for growth of the xerophyte Pugionium cornutum by enhancing osmotic adjustment capacity under salt and drought stresses. J. Exp. Bot. 2020, 71, 4215–4231. [Google Scholar] [CrossRef]
- Hasegawa, P.M.; Bressan, R.A.; Zhu, J.K.; Bohnert, H.J. Plant cellular and molecular responses to high salinity. Annu. Rev. Plant Physiol. 2000, 51, 463–499. [Google Scholar] [CrossRef]
- Wang, Y.; Zhao, Z.; Liu, F.; Sun, L.; Hao, F. Versatile roles of aquaporins in plant growth and development. Int. J. Mol. Sci. 2020, 21, 9485. [Google Scholar] [CrossRef]
- Wang, J.; Yang, L.; Chai, S.; Ren, Y.; Guan, M.; Ma, F.; Liu, J. An aquaporin gene MdPIP1;2 from Malus domestica confers salt tolerance in transgenic Arabidopsis. J. Plant Physiol. 2022, 273, 153711. [Google Scholar] [CrossRef]
- Kong, W.; Huang, H.; Du, W.; Jiang, Z.; Luo, Y.; Yi, D.; Yang, G.; Pang, Y. Overexpression of MsNIP2 improves salinity tolerance in Medicago sativa. J. Plant Physiol. 2024, 295, 154207. [Google Scholar] [CrossRef]
- Xu, Y.; Hu, W.; Liu, J.; Zhang, J.; Jia, C.; Miao, H.; Xu, B.; Jin, Z. A banana aquaporin gene, MaPIP1;1, is involved in tolerance to drought and salt stresses. BMC Plant Biol. 2014, 14, 59. [Google Scholar] [CrossRef]
- Qi, C.H.; Chen, M.; Song, J.; Wang, B.S. Increase in aquaporin activity is involved in leaf succulence of the euhalophyte Suaeda salsa, under salinity. Plant Sci. 2009, 176, 200–205. [Google Scholar] [CrossRef]
- Banihani, S.A. Radish (Raphanus sativus) and diabetes. Nutrients 2017, 9, 1014. [Google Scholar] [CrossRef] [PubMed]
- Manivannan, A.; Kim, J.; Kim, D.; Lee, E.; Lee, H. Deciphering the nutraceutical potential of Raphanus sativus—A comprehensive overview. Nutrients 2019, 11, 402. [Google Scholar] [CrossRef] [PubMed]
- Arunin, S. Saline Soil in Thailand; Land Development Department: Bangkok, Thailand, 1996. (In Thai) [Google Scholar]
- Hoffman, G.J.; Shannon, M.C. Salinity. In Microirrigation for Crop Production; Lamm, F.R., Ayars, J.E., Nakayama, F.S., Eds.; Elsevier: Amsterdam, The Netherlands, 2007; pp. 131–160. [Google Scholar] [CrossRef]
- Jungklang, J. Effects of sodium chloride on germination, growth, relative water content, and chlorophyll, proline, malondialdehyde and vitamin C contents in Chinese white radish seedlings (Raphanus sativus L. var. longipinnatus Bailey). Maejo Int. J. Sci. Technol. 2018, 12, 89–100. [Google Scholar]
- Yuan, G.; Wang, X.; Guo, R.; Wang, Q. Effect of salt stress on phenolic compounds, glucosinolates, myrosinase and antioxidant activity in radish sprouts. Food Chem. 2010, 121, 1014–1019. [Google Scholar] [CrossRef]
- Omuto, C.T.; Vargas, R.R.; El Mobarak, A.M.; Mohamed, N.; Viatkin, K.; Yigini, Y. Mapping of Salt-Affected Soils: Technical Manual; FAO: Rome, Italy, 2020. [Google Scholar] [CrossRef]
- Blaylock, A.D. Soil Salinity, Salt Tolerance and Growth Potential of Horticultural and Landscape Plants; Co-Operative Extension Service, University of Wyoming, Department of Plant, Soil and Insect Sciences, College of Agriculture: Laramie, WY, USA, 1994. [Google Scholar]
- Slama, I.; Ghnaya, T.; Messedi, D.; Hessini, K.; Labidi, N.; Savoure, A.; Abdelly, C. Effect of sodium chloride on the response of the halophyte species Sesuvium portulacastrum grown in mannitol-induced water stress. J. Plant Res. 2007, 120, 291–299. [Google Scholar] [CrossRef]
- Nieves-Cordones, M.; Al Shiblawi, F.R.; Sentenac, H. Role and transport of sodium and potassium in plants. Met. Ions Life Sci. 2016, 16, 291–324. [Google Scholar] [CrossRef]
- Taiz, L.; Zeiger, E. Plant Physiology, 5th ed.; Sinauer Associates Inc. Publishers: Massachusetts, UK, 2010. [Google Scholar]
- Colmenero-Flores, J.M.; Franco-Navarro, J.D.; Cubero-Font, P.; Peinado-Torrubia, P.; Rosales, M. Chloride as a beneficial macronutrient in higher plants: New roles and regulation. Int. J. Mol. Sci. 2019, 20, 4686. [Google Scholar] [CrossRef]
- Shaddad, M.A. The effect of proline application on the physiology of Raphanus sativus plant grown under salinity stress. Biol. Plant. 1990, 32, 104–112. [Google Scholar] [CrossRef]
- Nuanjan, N.; Nghia, P.T.; Theerakulpisut, P. Exogenous proline and trehalose promote recovery of rice seedlings from salt-stress and differentially modulate antioxidant enzymes and expression of related genes. J. Plant Physiol. 2012, 169, 596–604. [Google Scholar] [CrossRef]
- Wu, G.Q.; Jiao, Q.; Shui, Q.Z. Effect of salinity on seed germination, seedling growth and inorganic and organic solutes accumulation in sunflower (Helianthus annuus L.). Plant Soil Environ. 2015, 61, 220–226. [Google Scholar] [CrossRef]
- Inayat, H.; Mehmood, H.; Danish, S.; Alharbi, S.A.; Ansari, M.J.; Datta, R. Impact of cobalt and proline foliar application for alleviation of salinity stress in radish. BMC Plant Biol. 2024, 24, 287. [Google Scholar] [CrossRef] [PubMed]
- Blatt, M.R.; Chaumont, F.; Farquhar, G. Focus on Water. Plant Physiol. 2014, 164, 1553–1555. [Google Scholar] [CrossRef]
- Ievinsh, G. Water content of plant tissues: So simple that almost forgotten? Plants 2023, 12, 1238. [Google Scholar] [CrossRef]
- Kramer, P.J. Water Relation of Plants; New York Academic Press: New York, NY, USA, 1983; Available online: https://www.sciencedirect.com/book/9780124250406/water-relations-of-plants (accessed on 17 July 2024).
- Gonzalez, L.; Gonzalez-Vilar, M. Chapter 14 Determination of relative water content. In Handbook of Plant Ecophysiology Techniques; Reigosa Roger, M.J., Ed.; Kluwer Academic Publishers: Dordrecht, The Netherlands; Springer: Berlin/Heidelberg, Germany, 2007; pp. 207–212. [Google Scholar] [CrossRef]
- Suga, S.; Komatsu, S.; Maeshima, M. Aquaporin isoform responsive to salt and water stresses and phytohormones in radish seedlings. Plant Cell Physiol. 2002, 43, 1229–1237. [Google Scholar] [CrossRef]
- Fricke, W.; Knipfer, T. Plant aquaporins and cell elongation. In Plant Aquaporins: From Transport to Signaling; Chaumont, F., Tyerman, S., Eds.; Springer: Berlin/Heidelberg, Germany, 2017; pp. 107–131. [Google Scholar]
- Leng, H.; Jiang, C.; Song, X.; Lu, M.; Wan, X. Poplar aquaporin PIP1;1 promotes Arabidopsis growth and development. BMC Plant Biol. 2021, 21, 253. [Google Scholar] [CrossRef]
- Lopaz-Zaplana, A.; Martinez-Garcia, N.; Carvajal, M.; Bárzana, G. Relationships between aquaporins gene expression and nutrient concentrations in melon plants (Cucumis melo L.) during typical abiotic stresses. Environ. Exp. Bot. 2022, 195, 104759. [Google Scholar] [CrossRef]
- Yaneff, A.; Sigaut, L.; Marquez, M.; Alleva, K.; Pietrasanta, L.I.; Amodeo, G. Heteromerization of PIP aquaporins affects their intrinsic permeability. Proc. Natl. Acad. Sci. USA 2013, 111, 231–236. [Google Scholar] [CrossRef]
- Tripathi, D.K.; Singh, V.P.; Chauhan, D.K.; Prasad, S.M.; Dubey, N.K. Chapter 8 Role of macronutrients in plant growth and acclimation: Recent advances and future prospective. In Improvement of Crops in the Era of Climatic Changes, Volume 2; Ahmad, P., Wani, M.R., Azooz, M.M., Tran, L.P., Eds.; Springer: Berlin/Heidelberg, Germany, 2014; pp. 197–216. [Google Scholar] [CrossRef]
- Mndi, O.; Sogoni, A.; Jimoh, M.O.; Wilmot, C.M.; Rautenbach, F.; Laubscher, C.P. Interactive Effects of salinity stress and irrigation intervals on plant growth, nutritional value, and phytochemical content in Mesembryanthemum crystallinum L. Agriculture 2023, 13, 1026. [Google Scholar] [CrossRef]
- Wu, H.; Zhang, X.; Giraldo, J.P.; Shabala, S. It is not all about sodium: Revealing tissue specificity and signaling roles of potassium in plant responses to salt stress. Plant Soil 2018, 431, 1–17. [Google Scholar] [CrossRef]
- Zarei, M.; Azizi, M. Evaluation of NaCl salinity tolerance of four fig genotypes based on vegetative growth and ion content in leaves, shoots, and roots. J. Am. Soc. Hortic. Sci. 2016, 51, 1427–1434. [Google Scholar] [CrossRef]
- Efrida, E. Effect of NaCl and KCl to the growth and nutrient level of upland rice in an Ultisol. In Proceedings of the 7th International Conference on Multidisciplinary Research, Munich, Germany, 19–21 October 2018; pp. 143–148. [Google Scholar] [CrossRef]
- Shi, Y.; Chen, J.; Hou, X. Similarities and differences of photosynthesis establishment related mRNAs and novel lncRNAs in early seedlings (coleoptile/cotyledon vs. true leaf) of rice and Arabidopsis. Front. Genet. 2020, 11, 565006. [Google Scholar] [CrossRef] [PubMed]
- Santos, C.V. Regulation of chlorophyll biosynthesis and degradation by salt stress in sunflower leaves. Sci. Hortic. 2004, 103, 93–99. [Google Scholar] [CrossRef]
- Turan, M.A.; Katkat, V.; Taban, S. Variation in proline, chlorophyll and mineral contents of wheat plant grown under salinity stress. J. Agron. 2007, 6, 137–141. [Google Scholar] [CrossRef]
- M’rah, S.; Ouerghi, Z.; Berthomieu, C.; Havaux, M.; Jungas, C.; Hajji, M.; Grignin, C.; Lachaâl, M. Effects of NaCl on the growth, ion accumulation and photosynthetic parameters of Thellungiella halophila. J. Plant Physiol. 2006, 163, 1022–1031. [Google Scholar] [CrossRef]
- Land Development Department. Handbook for Analyzing Samples of Soil, Water, Fertilizer, Plants, and Soil Amendments: Ensuring Product Standards Through Rigorous Analysis; Office of Science for Land Development, Land Development Department: Bangkok, Thailand, 2004. (In Thai) [Google Scholar]
- Department of Agriculture. A Handbook of Soil Analysis (Chemical and Physical Methods); Quick Print Offset: Bangkok, Thailand, 2010. (In Thai) [Google Scholar]
- Smart, R.E.; Bingham, G.E. Rapid estimates of relative water content. Plant Physiol. 1974, 53, 258–260. [Google Scholar] [CrossRef]
- Zeng, Y.; Li, L.; Yang, R.; Yi, X.; Zhang, B. Contribution and distribution of inorganic ions and organic compounds to the osmotic adjustment in Halostachys caspica response to salt stress. Sci. Rep. 2015, 5, 13639. [Google Scholar] [CrossRef]
- Suga, S.; Imagawa, S.; Maeshima, M. Specificity of the accumulation of mRNA and proteins of the plasma membrane and tonoplast aquaporins in radish organs. Planta 2001, 212, 294–304. [Google Scholar] [CrossRef]
- Tsuchihira, A.; Hanba, Y.T.; Kato, N.; Doi, T.; Kawazu, T.; Maeshima, M. Effect of overexpression of radish plasma membrane aquaporins water-use efficiency, photosunthesis and growth of Eucayptus trees. Tree Physiol. 2010, 30, 417–430. [Google Scholar] [CrossRef]
- Higuchi, T.; Suga, S.; Tsuchiya, T.; Hisada, H.; Morishima, S.; Okada, Y.; Maeshima, M. Molecular Cloning, Water Channel Activity and Tissue Specific Expression of Two Isoforms of Radish Vacuolar Aquaporin. Plant Cell Physiol. 1998, 39, 905–913. [Google Scholar] [CrossRef]
- Xu, Y.; Zhu, X.; Gong, Y.; Xu, L.; Wang, Y.; Liu, L. Evaluation of reference genes for gene expression studied in radish (Raphanus sativus L.) using quantitive real-time PCR. Biochem. Biophys. Res. Commun. 2012, 424, 398–403. [Google Scholar] [CrossRef] [PubMed]
- Duan, M.; Wang, J.; Zhang, X.; Yang, H.; Wang, H.; Qui, Y.; Song, J.; Guo, Y.; Li, X. Identification of optimal reference genes for expression analysis in radish (Raphanus sativus L.) and its relatives based on expression stability. Front. Plant Sci. 2017, 8, 1605. [Google Scholar] [CrossRef] [PubMed]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2−ΔΔCT method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Hoo, K.Y.; Shim, I.S.; Kobayashi, K.; Usui, K. Relationship between Na content or K/Na ratio in shoots and salt tolerance in several Gramineous plants. J. Weed Sci. Technol. 1999, 44, 293–299. [Google Scholar] [CrossRef]
- Science Center of Land Development. Manual for the Analysis of Soil Samples, Water, Fertilizers, Plants, Soil Improvement Materials and Analysis to Inspect and Certify Product Standards; Land Development Department: Bangkok, Thailand, 2004. (In Thai) [Google Scholar]
- Walkley, A.; Black, I.A. Acritical examination of a rapid method for determining organic carbon in soil-effect of variation in digestions and of inorganic soil constituents. Soil Sci. 1947, 63, 251–263. [Google Scholar] [CrossRef]
- Dean, R.P.; Miskiewicz, E. Rates of electron transport in the thylakoid membranes of isolated, illuminated chloroplasts are enhanced in the presence of ammonium chloride. Biochem. Mol. Biol. Educ. 2003, 31, 410–417. [Google Scholar] [CrossRef]
- Latkowska, E.; Lechowski, Z.; Bialczyk, J.; Pilarski, J. Photosynthesis and water relations in tomato plants cultivated long-term in media containing (+)-Usnic acid. J. Chem. Ecol. 2006, 32, 2053–2066. [Google Scholar] [CrossRef]
- Kumari, S.; Phogat, D.; Sehrawat, K.D.; Choudhary, R.; Rajput, V.D.; Ahlawat, J.; Karunakaran, R.; Minkina, T.; Sehrawat, A.R. The effect of Ascophyllum nodosum extract on the nutraceutical antioxidant potential of Vigna radiata sprout under salt stress. Plants 2021, 10, 1216. [Google Scholar] [CrossRef]
- Bates, L.S.; Waldren, R.P.; Teare, I.D. Rapid determination of free proline for water-stress studies. Plant Soil 1973, 39, 205–207. [Google Scholar] [CrossRef]
- Ghoulam, C.; Foursy, A.; Fares, K. Effect of salt stress on growth, inorganic ions and proline accumulation in relation to osmotic adjustment in five sugar beet cultivars. Environ. Exp. Bot. 2002, 47, 39–50. [Google Scholar] [CrossRef]

Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Masepan, N.; Intarasit, S.; Panya, A.; Jungklang, J. Low NaCl Concentrations Increase Cotyledon Growth in Chinese White Radish (Raphanus sativus L. var. longipinnatus Bailey) Seedlings via Aquaporin-Mediated Water Transport. Plants 2025, 14, 1616. https://doi.org/10.3390/plants14111616
Masepan N, Intarasit S, Panya A, Jungklang J. Low NaCl Concentrations Increase Cotyledon Growth in Chinese White Radish (Raphanus sativus L. var. longipinnatus Bailey) Seedlings via Aquaporin-Mediated Water Transport. Plants. 2025; 14(11):1616. https://doi.org/10.3390/plants14111616
Chicago/Turabian StyleMasepan, Nutkamol, Sitthisak Intarasit, Aussara Panya, and Jarunee Jungklang. 2025. "Low NaCl Concentrations Increase Cotyledon Growth in Chinese White Radish (Raphanus sativus L. var. longipinnatus Bailey) Seedlings via Aquaporin-Mediated Water Transport" Plants 14, no. 11: 1616. https://doi.org/10.3390/plants14111616
APA StyleMasepan, N., Intarasit, S., Panya, A., & Jungklang, J. (2025). Low NaCl Concentrations Increase Cotyledon Growth in Chinese White Radish (Raphanus sativus L. var. longipinnatus Bailey) Seedlings via Aquaporin-Mediated Water Transport. Plants, 14(11), 1616. https://doi.org/10.3390/plants14111616