
Geobotanical Study and Preservation of Rare and Endangered Rosaceae Species
 , , , , and
, , , , and 
Abstract
1. Introduction
2. Results and Discussion
2.1. Description of the Rosaceae Species Cultivated in Their Natural Habitat
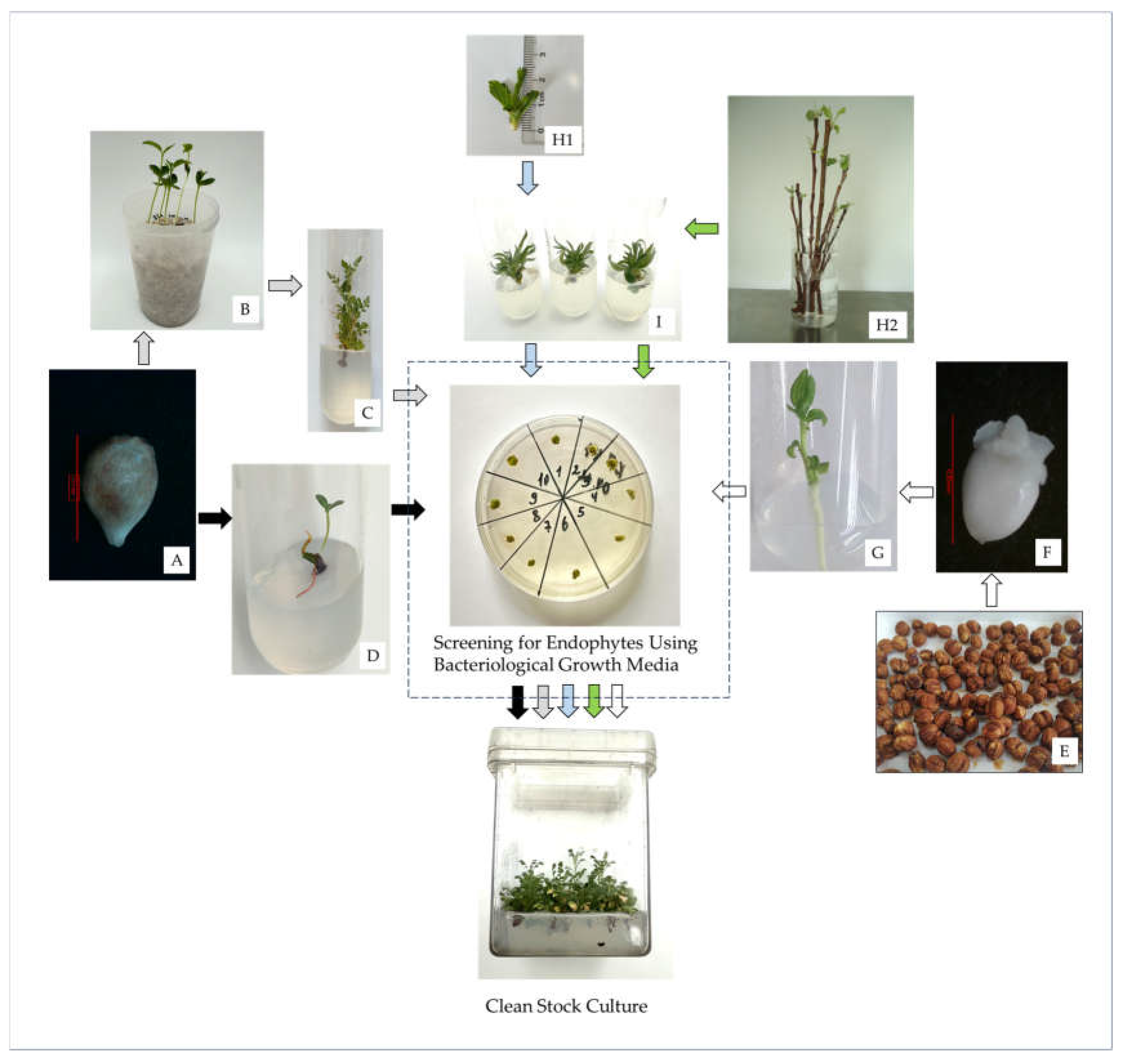
2.2. In Vitro Establishment of Rosaceae Accessions
2.2.1. Sexual Initiation Using Seeds or Embryonic Axes
2.2.2. Asexual Initiation Using Shoots
2.3. In Vitro Multiplication
2.4. Seed Storage at −20 °C or Storage of Seeds and Embryonic Axes at Cryogenic Temperature
2.5. Creation of a Cryogenic DNA Bank
3. Materials and Methods
3.1. Plant Material Acquisition
3.2. In Vitro Establishment of Rosaceae Accessions
3.2.1. Sexual Initiation Using Seeds or Embryonic Axes
3.2.2. Asexual Initiation Using Shoots
3.2.3. Screening for Endophytes Using Bacteriological Growth Media
3.3. In Vitro Multiplication
3.4. Seed Storage at −20 °C or at Cryogenic Temperatures
3.5. Creation of a Cryogenic Embryonic Axes Bank
3.6. Creation of a Cryogenic DNA Bank
3.7. Statistical Analysis
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Fernández-Palacios, J.M.; Kreft, H.; Irl, S.D.H.; Norder, S.; Ah-Peng, C.; Borges, P.A.V.; Burns, K.C.; de Nascimento, L.; Meyer, J.-Y.; Montes, E.; et al. Scientists’ warning—The outstanding biodiversity of islands is in peril. Glob. Ecol. Conserv. 2021, 31, E01847. [Google Scholar] [CrossRef] [PubMed]
- Heywood, V.V. Conserving plants within and beyond protected areas—still problematic and future uncertain. Plant Divers. 2019, 41, 36–39. [Google Scholar] [CrossRef] [PubMed]
- Marchese, C. Biodiversity hotspots: A shortcut for a more complicated concept. Glob. Ecol. Conserv. 2015, 3, 297–309. [Google Scholar] [CrossRef]
- Molkanova, O.I.; Vasilyeva, O.G.; Konovalova, L.N. The scientific basis for conservation and sustainable reproduction of plant genofond in in vitro culture. Bull. UdSU-Ser. Biol. 2015, 25, 95–100. (In Russian) [Google Scholar]
- Myers, N.; Mittermeier, R.A.; Mittermeier, C.G.; Fonseca, G.A.B.; Kent, J. Biodiversity hotspots for conservation priorities. Nature 2000, 403, 853–858. [Google Scholar] [CrossRef]
- Romadanova, N.V.; Kushnarenko, S.V. Conservation of plant biodiversity by biotechnology methods. Proc. Appl. Bot. Genet. Breed. 2023, 184, 239–248. [Google Scholar] [CrossRef]
- Fazio, G.; Bettoni, J.C.; Costa, L.C.; Hurtado-Gonzales, O.P.; Al Rwahnih, M.; Steinberger, A.; Nedrow, A.; Volk, G.M.; Adams, S.; Adams, R.; et al. Virus studies in the Geneva® apple rootstock breeding program. In New York Fruit Quarterly; Robinson, T., Ed.; New York State Horticultural Society: New York, NY, USA, 2022; pp. 20–24. [Google Scholar]
- Forsline, P.L.; Aldwinckle, H.S. Evaluation of Malus sieversii seedling populations for disease resistance and horticultural traits. Acta Hort. 2004, 663, 529–534. [Google Scholar] [CrossRef]
- Volk, G.M.; Bramel, P. A strategy to conserve worldwide apple genetic resources: Survey results. Acta Hortic. 2017, 1172, 99–106. [Google Scholar] [CrossRef]
- Robinson, T.; Aldwinckle, H.S.; Fazio, G.; Holleran, T. The Geneva series of apple rootstocks from Cornell: Performance, disease resistance, and commercialization. Acta Hortic. 2003, 622, 513–520. [Google Scholar] [CrossRef]
- Liu, Z.; Li, X.; Wen, X.; Zhang, Y.; Ding, Y.; Zhang, Y.; Gao, B.; Zhang, D. PacBio full-length transcriptome of wild apple (Malus sieversii) provides insights into canker disease dynamic response. BMC Genom. 2021, 22, 52. [Google Scholar] [CrossRef]
- Petruzzello, M. List of Economically Important Members of The Family Rosaceae. Encyclopedia Britannica. 2024. Available online: https://www.britannica.com/plant/Rosaceae (accessed on 28 February 2025).
- Pagliarani, G.; Dapena, E.; Miñarro, M.; Denancé, C.; Lespinasse, Y.; RatMorris, E.; Troggio, M.; Durel, C.E.; Tartarini, S. Fine mapping of the rosy apple aphid resistance locus Dp-fl on linkage group 8 of the apple cultivar ‘Florina’. Tree Genet. Genom. 2016, 12, 56. [Google Scholar] [CrossRef]
- Afifi, L.; Lapin, K.; Tremetsberger, K.; Konrad, H. A systematic review of threats, conservation, and management measures for tree species of the family Rosaceae in Europe. Flora 2023, 301, 152244. [Google Scholar] [CrossRef]
- Mammadov, T.; Abbaszade, Z. Rare endangered species of the Rosaceae family of the Rosa L. genus in the flora of Azerbaijan. Bull. Sci. Pract. 2022, 8, 51–57. (In Russian) [Google Scholar] [CrossRef]
- Nurtaza, A.; Dyussembekova, D.; Islamova, S.; Samatova, I.; Zhanybekova, Z.; Umirzakova, A.; Magzumova, G.; Muranets, A.; Kakimzhanova, A. In vitro conservation and genetic diversity analysis of rare species Ribes janczewskii. Sci. Rep. 2024, 14, 31117. [Google Scholar] [CrossRef]
- Dzhangaliev, A.D. The wild apple tree of Kazakhstan. Hortic. Rev. 2003, 29, 63–303. [Google Scholar] [CrossRef]
- Volk, G.M.; Henk, A.D.; Richards, C.M.; Forsline, P.L.; Chao, C.T. Malus sieversii: A diverse central Asian apple species in the USDA-ARS National Plant Germplasm System. HortScience 2013, 48, 1440–1444. [Google Scholar] [CrossRef]
- Baitulin, I.O. (Ed.) Red Book of Kazakhstan, 2nd ed.; ArtPrintXX1 LLP: Astana, Kazakhstan, 2014; Volume 2, 452p. (In Russian) [Google Scholar]
- Bramel, P.; Volk, G.M. A Global Strategy for the Conservation and Use of Apple Genetic Resources; Crop Trust: Bonn, Germany, 2019; p. 50. [Google Scholar] [CrossRef]
- Lala, S.; Amri, A.; Maxted, N. Towards the conservation of crop wild relative diversity in North Africa: Checklist, prioritisation and inventory. Genet. Resour. Crop Evol. 2018, 65, 113–124. [Google Scholar] [CrossRef]
- Vincent, H.; Wiersema, J.; Kell, S.; Fielder, H.; Dobbie, S.; Castañeda-Álvarez, N.P.; Guarino, L.; Eastwood, R.; Leόn, B.; Maxted, N. A prioritized crop wild relative inventory to help underpin global food security. Biol. Conserv. 2013, 167, 265–275. [Google Scholar] [CrossRef]
- Tian, Z.; Song, H.; Wang, Y.; Li, J.; Maimaiti, M.; Liu, Z.; Zhang, H.; Zhang, J. Wild apples are not that wild: Conservation status and potential threats of Malus sieversii in the mountains of central Asia biodiversity hotspot. Diversity 2022, 14, 489. [Google Scholar] [CrossRef]
- Volk, G.M.; Chao, C.T.; Norelli, J.; Brown, S.K.; Fazio, G.; Peace, C.; McFerson, J.; Zhong, G.-H.; Bretting, P. The vulnerability of US apple (Malus) genetic resources. Genet. Resour. Crop Evol. 2015, 62, 765–794. [Google Scholar] [CrossRef]
- Reim, S.; Schiffler, J.; Braun-Lüllemann, A.; Schuster, M.; Flachowsky, H.; Höfer, M. Genetic and pomological determination of the trueness-to-type of sweet cherry cultivars in the German National Fruit Genebank. Plants 2023, 12, 205. [Google Scholar] [CrossRef] [PubMed]
- Buiteveld, J.; Koehorst-van Putten, H.J.J.; Kodde, L.; Laros, I.; Tumino, G.; Howard, N.; van de Weg, E.; Smulders, M.J.M. Advanced genebank management of genetic resources of European wild apple, Malus sylvestris, using genome-wide SNP array data. Tree Genet. Genomes 2021, 17, 32. [Google Scholar] [CrossRef]
- Romadanova, N.V.; Mishustina, S.A.; Matakova, G.N.; Kuhsnarenko, S.V.; Rakhimbaev, I.R.; Reed, B.M. In vitro collection of Malus shoot cultures for cryogenic bank development in Kazakhstan. Acta Hortic. 2016, 1113, 271–277. [Google Scholar] [CrossRef]
- Romadanova, N.V.; Aralbayeva, M.M.; Zemtsova, A.S.; Alexandrova, A.M.; Kazybayeva, S.Z.; Mikhailenko, N.V.; Kushnarenko, S.V.; Bettoni, J.C. In vitro collection for the safe storage of grapevine hybrids and identification of the presence of Plasmopora viticola resistance genes. Plants 2024, 13, 1089. [Google Scholar] [CrossRef] [PubMed]
- Panis, B.; Nagel, M.; Van den Houwe, I. Challenges and prospects for the conservation of crop genetic resources in field genebanks, in in vitro collections and/or in liquid nitrogen. Plants 2020, 9, 1634. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Bonnart, R.; Volk, G.M. Challenges in implementing plant shoot tip cryopreservation technologies. Plant Cell Tissue Organ Cult. 2021, 144, 21–34. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Markovic, Z.; Bi, W.; Volk, G.M.; Matsumoto, T.; Wang, Q.-C. Grapevine shoot tip cryopreservation and cryotherapy: Secure storage of disease-free plants. Plants 2021, 10, 2190. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Wang, M.-R.; Li, J.-W.; Fan, X.; Fazio, G.; Hurtado-Gonzales, O.P.; Volk, G.M.; Wang, Q.-C. Application of biotechniques for in vitro virus and viroid elimination in pome fruit crops. Phytopathology 2024, 114, 930–954. [Google Scholar] [CrossRef]
- Volk, G.M.; Bonnart, R.M.; Henk, A.D.; Chen, K.Y.; Bettoni, J.C.; Wang, Q.-C.; Kreckel, H.D.; Levinger, N.E. Fundamentals of plant cryopreservation: Dormant bud two-step cooling and shoot tip vitrification. Acta Hortic. 2025, 1421, 117–124. [Google Scholar] [CrossRef]
- Wang, M.-R.; Chen, L.; da Silva, J.A.; Volk, G.M.; Wang, Q.-C. Cryobiotechnology of apple (Malus spp.): Development, progress and future prospects. Plant Cell Rep. 2018, 37, 689–709. [Google Scholar] [CrossRef]
- Bettoni, J.; Kretzschmar, A.A.; Bonnart, R.; Shepherd, A.; Volk, G.M. Cryopreservation of 12 Vitis species using apical shoot tips derived from plants grown in vitro. HortScience 2019, 54, 976–981. [Google Scholar] [CrossRef]
- Benelli, C.; Tarraf, W.; Izgu, T.; De Carlo, A. In vitro conservation through slow growth storage technique of fruit species: An overview of the last 10 years. Plants 2022, 11, 3188. [Google Scholar] [CrossRef]
- Kushnarenko, S.V.; Romadanova, N.V.; Aralbayeva, M.M. Current state and in vitro conservation of the only endangered population of Corylus avellana in Kazakhstan. Res. Crops 2020, 21, 681–686. [Google Scholar] [CrossRef]
- Dobránszki, J.; Teixeira da Silva, J.A. Micropropagation of apple—A review. Biotechnol. Adv. 2010, 28, 462–488. [Google Scholar] [CrossRef]
- Kovalchuk, I.Y.; Zhumagulova, Z.; Turdiyev, T.T.; Reed, B.M. Growth medium alterations improve in vitro cold storage of pear germplasm. Cryo Lett. 2014, 35, 197–203. [Google Scholar]
- Sedlak, J.; Zidova, P.; Paprstein, F. Slow growth in vitro conservation of fruit crops. Acta Hortic. 2019, 1234, 119–124. [Google Scholar] [CrossRef]
- Kim, H.H.; Popova, E.; Shin, D.J.; Yi, J.Y.; Kim, C.H.; Yoon, M.K. Cryobanking of Korean Allium Germplasm Collections: Results from a 10 Year Experience. Cryo Lett. 2012, 33, 45–57. [Google Scholar]
- Kushnarenko, S.V.; Rymkhanova, N.K.; Aralbayeva, M.M.; Romadanova, N.V. PVS2 vitrification technique for preservation of Juglans regia L. genetic resources in Kazakhstan. Cryo Lett. 2023, 44, 240–248. [Google Scholar] [CrossRef]
- Pence, V.C.; Ballesteros, D.; Walters, C.; Reed, B.M.; Philpott, M.; Dixon, K.W.; Pritchard, H.W.; Culley, T.M.; Vanhove, A.C. Cryobiotechnologies: Tools for expanding long-term ex situ conservation to all plant species. Biol. Conserv. 2020, 250, 108736. [Google Scholar] [CrossRef]
- Reed, B.M. Plant Cryopreservation: A Practical Guide; Reed, B.M., Ed.; Springer: New York, NY, USA, 2008; p. 513. [Google Scholar]
- Popov, A.S.; Popova, E.V.; Nikishina, T.V.; Vysotskaya, O.N. Cryobank of plant genetic resources in Russian Academy of Sciences. Int. J. Refrig. 2006, 29, 403–410. [Google Scholar] [CrossRef]
- Migicovsky, Z.; Warschefsky, E.; Klein, L.L.; Miller, A.J. Using living germplasm collection to characterize, improve, and conserve wood perennials. Crop Sci. 2019, 59, 2365–2380. [Google Scholar] [CrossRef]
- Reed, B.M.; Gupta, S.; Uchendu, E.E. In vitro genebanks for preserving tropical biodiversity. In Conservation of Tropical Plant Species; Normah, M.N., Chin, H.F., Reed, B.M., Eds.; Springer: New York, NY, USA, 2013; pp. 77–106. [Google Scholar]
- Jenderek, M.M.; Reed, B.M. Cryopreserved storage of clonal germplasm in the USDA National plant germplasm system. Vitr. Cell. Dev. Biol.-Plant 2017, 53, 299–308. [Google Scholar] [CrossRef]
- Zhang, A.-L.; Wang, M.-R.; Li, Z.; Panis, B.; Bettoni, J.C.; Vollmer, R.; Xu, L.; Wang, Q.-C. Overcoming challenges for shoot tip cryopreservation of root and tuber crops. Agronomy 2023, 13, 219. [Google Scholar] [CrossRef]
- Vollmer, R.; Villagaray, R.; Castro, M.; Cárdenas, J.; Pineda, S.; Espirilla, J.; Anglin, N.; Ellis, D.; Azevedo, V.C.R. The world’s largest potato cryobank at the International Potato Center (CIP)—Status quo, protocol improvement through large-scale experiments and long-term viability monitoring. Front. Plant Sci. 2022, 13, 1059817. [Google Scholar] [CrossRef]
- Bettoni, J.C.; Fazio, G.; Carvalho Costa, L.; Hurtado-Gonzales, O.P.; Rwahnih, M.A.; Nedrow, A.; Volk, G.M. Thermotherapy followed by shoot tip cryotherapy eradicates latent viruses and Apple Hammerhead viroid from in vitro apple rootstocks. Plants 2022, 11, 582. [Google Scholar] [CrossRef]
- Walters, C.; Richards, C.M.; Volk, G.M. Genebank conservation of germplasm collected from wild species. In North American Crop Wild Relatives; Greene, S., Williams, K., Khoury, C., Kantar, M., Marek, L., Eds.; Springer: New York, NY, USA, 2018; pp. 245–280. [Google Scholar]
- FAO (Food and Agriculture Organization of the United Nations). Country Report on the State of Plant Genetic Resources for Food and Agriculture in the Kazakhstan Republic. Scientific and Production Center of Farming and Plant Growing Ministry of Agriculture of Kazakhstan Republic. Production in 2007. Available online: https://www.fao.org/4/i1500e/Kazakhstan.pdf (accessed on 27 March 2025).
- Ha, Y.-H.; Oh, S.-H.; Lee, S.-R. Genetic admixture in the population of wild apple (Malus sieversii) from the Tien Shan Mountains, Kazakhstan. Genes 2021, 12, 104. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.-H.; Popova, E. Improvement and innovation of cryopreservation and in vitro methods in plant resource protection. Biology 2025, 13, 741. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). The Third Report on The State of the World’s Plant Genetic Resources for Food and Agriculture. Production in 2025. Available online: https://openknowledge.fao.org/handle/20.500.14283/cd4711en (accessed on 27 March 2025).
- Kushnarenko, S.; Salnikov, E.; Nurtazin, M.; Mukhitdinova, Z.; Rakhimbaev, I.; Reed, B.M. Characterization and cryopreservation of Malus sieversii seeds. Asian Australas. J. Plant Sci. Biotechnol. 2010, 4, 5–9. [Google Scholar] [CrossRef]
- Damasco, O.P.; Huelgas, V.C.; Uy, J.N.R.; Descalsota, M.L.V.; Descalsota, J.C. Diversity assessment in the in vitro conservation response of philippine yam (Dioscorea species) accessions. Philipp. J. Sci. 2021, 151, 55–67. [Google Scholar] [CrossRef]
- Kulak, V.; Longboat, S.; Brunet, N.D.; Shukla, M.; Saxena, P. In vitro technology in plant conservation: Relevance to biocultural diversity. Plants 2022, 11, 503. [Google Scholar] [CrossRef]
- Radomir, A.-M.; Stan, R.; Florea, A.; Ciobotea, C.-M.; Bănuță, F.M.; Negru, M.; Neblea, M.A.; Sumedrea, D.I. Overview of the success of in vitro culture for ex situ conservation and sustainable utilization of endemic and subendemic native plants of Romania. Sustainability 2023, 15, 2581. [Google Scholar] [CrossRef]
- Dvirna, T.; Futorna, O.; Minarchenko, V.; Tymchencko, I. Morphological features of fruits and seeds of some species of the genus Crataegus L. of the flora of Ukraine. Acta Agrobot. 2021, 78, 7418. [Google Scholar] [CrossRef]
- Tuyakova, A.T.; Imanbayeva, A.A.; Duysenova, N.I.; Ishmuratova, M.Y. Anatomical structure study of aerial organs of Crataegus Ambigua C. A. Mey. Biotech. Res. Asia 2016, 13, 3. [Google Scholar] [CrossRef]
- Kheloufi, A.; Mansouri, L.; Vanbellinghen, C. Seed germination of Crataegus monogyna—A species with a stony endocarp. Reforesta 2019, 7, 73–80. [Google Scholar] [CrossRef]
- Murashige, T.; Skoog, F. A revised medium for rapid growth and bioassays with tobacco tissue cultures. Physiol. Plant 1962, 15, 473–497. [Google Scholar] [CrossRef]
- Viss, P.R.; Brooks, E.M.; Driver, J.A. A simplified method for the control of bacterial contamination in woody plant tissue culture. Vitr. Cell. Dev. Biol. Plant 1991, 27, 42. [Google Scholar] [CrossRef]
- Pasternak, T.P.; Steinmacher, D. Plant growth regulation in cell and tissue culture in vitro. Plants 2024, 13, 327. [Google Scholar] [CrossRef]
- Hesami, M.; Pepe, M.; Monthony, A.S.; Baiton, A.; Jones, A.M.P. Modeling and optimizing in vitro seed germination of industrial hemp (Cannabis sativa L.). Ind. Crop. Prod. 2021, 170, 113753. [Google Scholar] [CrossRef]
- Huaran, H.; Hao, l.; Feihu, L. Seed germination of hemp (Cannabis sativa L.) cultivars responds differently to the stress of salt type and concentration. Ind. Crop. Prod. 2018, 123, 254–261. [Google Scholar] [CrossRef]
- Hwang, J.E.; Park, H.B.; Jeon, D.Y.; Park, H.J.; Kim, S.; Lee, C.W.; Kim, Y.-J.; Yoon, Y.-J. Effect of different basal media and organic supplements on in vitro seedling development of the endangered orchid species Dendrobium moniliforme (L.) Swartz. Plants 2024, 13, 2721. [Google Scholar] [CrossRef]
- Romadanova, N.V.; Tolegen, A.B.; Kushnarenko, S.V.; Zholdybayeva, E.V.; Bettoni, J.C. Effect of Plant Preservative MixtureTM on endophytic bacteria eradication from in vitro-grown apple shoots. Plants 2022, 11, 2624. [Google Scholar] [CrossRef] [PubMed]
- Volk, G.M.; Bonnart, R.; Araújo de Oliveira, A.C.; Henk, A.D. Minimizing the deleterious effects of endophytes in plant shoot tip cryopreservation. Appl. Plant Sci. 2022, 10, e11489. [Google Scholar] [CrossRef]
- Gunson, H.E.; Spencer-Phillips, P.T.N. Latent bacterial infections: Epiphytes and endophytes as contaminants of micropropagated plants. In Physiology, Growth and Development of Plants in Culture; Lumsden, P.J., Nicholas, J.R., Davies, W.J., Eds.; Springer Nature: Cham, Switzerland, 1994; pp. 379–396. [Google Scholar]
- Abdalla, N.; El-Ramady, H.; Seliem, M.K.; El-Mahrouk, M.E.; Taha, N.; Bayoumi, Y.; Shalaby, T.A.; Dobránszki, J. An academic and technical overview on plant micropropagation challenges. Horticulturae 2022, 8, 677. [Google Scholar] [CrossRef]
- Pence, V.C.; Sandoval, J.A. Controlling contamination during in vitro collecting. In IPGRI Technical Bulletin No.7; Pence, V.C., Sandoval, J.A., Villalobos, V.M., Engelmann, F., Eds.; International Plant Genetic Resources Institute (IPGRI): Rome, Italy, 2002; pp. 30–40. [Google Scholar]
- Antonopoulou, C.; Dimassi, K.; Therios, I.; Chatzissavvidis, C.; Papadakis, I. The effect of Fe-EDDHA and of ascorbic acid on in vitro rooting of the peach rootstock GF-677 explants. Acta Physio. Plant. 2007, 29, 559–561. [Google Scholar] [CrossRef]
- Padmanabhan, P.; Shukla, M.R.; Sullivan, J.A.; Saxena, P.K. Iron supplementation promotes in vitro shoot induction and multiplication of Baptisia australis. Plant Cell Tissue Organ Cult. 2017, 129, 145–152. [Google Scholar] [CrossRef]
- Trejgell, A.; Libront, I.; Tretyn, A. The effect of Fe-EDDHA on shoot multiplication and in vitro rooting of Carlina onopordifolia Besser. Acta Physio. Plant. 2012, 34, 2051–2055. [Google Scholar] [CrossRef]
- Clapa, D.; Fira, A.; Rusu, T. The use of Isubgol and Sequestrene 138 for the in vitro propagation of the highbush blueberry (Vaccinium corymbosum L.). J. Food Agric. Environ. 2007, 6, 45–147. [Google Scholar]
- Chapa, D.; Bunea, C.; Borsai, O.; Pintea, A.; Hârţa, M.; Stefan, R. The role of Sequestrene 138 in highbush blueberry (Vaccinium corymbosum L.) micropropagation. HortScience 2018, 53, 1487–1493. [Google Scholar] [CrossRef]
- Fatima, N.; Ahmad, N.; Anis, M. Enhanced in vitro regeneration and change in photosynthetic pigments, biomass and proline content in Withania somnifera L. (Dunal) induced by copper and zinc ions. Plant Physiol. Biochem. 2011, 49, 1465–1471. [Google Scholar] [CrossRef]
- Aremu, A.O.; Plačková, L.; Pěnčík, A.; Novák, O.; Doležal, K.; Van Staden, J. Auxin-cytokinin interaction and variations in their metabolic products in the regulation of organogenesis in two Eucomis species. New Biotechnol. 2016, 6, 45–147. [Google Scholar] [CrossRef]
- Jakhar, M.L.; Dixit, D.; Ahmad, S.; Ola, M.P.; Ram, M.; Jat, H.R. Influence of plant growth regulators on micropropagation of Gwarpatha [Aloe vera (L.) Burm]. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 1782–1803. [Google Scholar] [CrossRef]
- Sadeghi, F.; Yadollahi, A.; Kermani, M.J.; Eftekhari, M. Optimizing culture media for in vitro proliferation and rooting of Tetra (Prunus empyrean 3) rootstock. J. Genet. Eng. Biotechnol. 2015, 13, 19–23. [Google Scholar] [CrossRef] [PubMed]
- Zou, Y.-N. Micropropagation of Chinese plum (Prunus salicina Lindl.) using mature stem segments. Not. Bot. Horti Agrobot. Cluj. 2010, 38, 214–218. [Google Scholar]
- Orazov, A.E.; Mukhitdinov, N.M.; Myrzagalieva, A.B.; Turuspekov, E.K.; Shramko, G.; Kubentaev, S.A. Distribution and characteristics of cenopopulations of Amygdalus ledebouriana Schlecht. in the territory of the Narym ridge. Bull. KazNU. Biol. Ser. 2019, 1, 37–45. (In Russian) [Google Scholar] [CrossRef]
- Sumbembaev, A.A. New locations of Amygdalus ledebouriana Sclecht. (Fabaceae)—Rare endemic species of Kazakhstan flora—On the Kalbinsky ridge. Bull. Bryansk Dpt. RBS 2018, 13, 22–28. [Google Scholar] [CrossRef]
- Serafimovich, M.; Kirillov, V.; Stikhareva, T. The influence of the culture medium composition on the establishment of explants of the almond ledeburovsky (Amygdalus ledebouriana Schlecht.) in vitro. Bull. Shakarim Uni.-Tech. Sci. 2020, 3, 289–293. (In Russian) [Google Scholar]
- Isikalan, Ç.; Akbas, F.A.; Namli, S.; Tilkat, E.; Basaran, D. In vitro micropropagation of almond (Amygdalus communis L. cv. Nonpareil). Afr. J. Biotechnol. 2008, 7, 1875–1880. [Google Scholar] [CrossRef]
- Sharifmoghadam, N.; Safarnejad, A.; Tabatabaei, S.M. The effect of plant growth regulators on callus induction and regeneration of Amygdalus communis. Not. Sci. Biol. 2011, 3, 97–100. [Google Scholar] [CrossRef]
- Ahmad, A.; Ahmad, N.; Anis, M.; Alatar, A.A.; Abdel-Salam, E.M.; Qahtan, A.A.; Faisal, M. Gibberellic acid and thidiazuron promote micropropagation of an endangered woody tree (Pterocarpus marsupium Roxb.) using in vitro seedlings. Plant Cell Tissue Organ Cult. 2021, 144, 449–462. [Google Scholar] [CrossRef]
- Rad, F.A.; Jafari, M.; Khezrinejad, N.; Miandoab, M.P. An efficient plant regeneration system via direct organogenesis with in vitro flavonoid accumulation and analysis of genetic fidelity among regenerants of Teucrium polium L. Hortic. Environ. Biotechnol. 2014, 55, 568–577. [Google Scholar] [CrossRef]
- Radhakrishnan, R.; Ramachandran, A.; Ranjitha Kumari, B.D. Rooting and shooting: Dual function of thidiazuron in in vitro regeneration of soybean (Glycine max L.). Acta Physiol. Plant. 2009, 31, 1213. [Google Scholar] [CrossRef]
- Gonbad, R.A.; Sinniah, U.R.; Aziz, M.A.; Mohamad, R. Influence of cytokinins in combination with GA3 on shoot multiplication and elongation of Tea clone Iran 100 (Camellia sinensis (L.) O. Kuntze). Sci. World J. 2014, 28, 943054. [Google Scholar] [CrossRef]
- Geng, F.; Moran, R.; Day, M.; Halteman, W.; Zhang, D. Increasing in vitro shoot elongation and proliferation of ‘G.30′ and ‘G.41′ apple by chilling explants and plant growth regulators. HortScience 2016, 51, 899–904. [Google Scholar] [CrossRef]
- Zemtsova, A.; Kushnarenko, S.; Romadanova, N. Creation of in vitro collection and cryogenic bank of Berberis heteropoda seeds. Bull. KazNU. Biol. Ser. 2022, 93, 4–13. [Google Scholar] [CrossRef]
- Kushnarenko, S.V.; Mukhitdinova, Z.R.; Aralbaeva, M.M. Cryopreservation of Seeds—Methodological Recommendations; TST-Company: Almaty, Kazakhstan, 2011; p. 33. (In Russian) [Google Scholar]
- Romadanova, N.V.; Karasholakova, L.N.; Makhmutova, I.A.; Ishmuratova, M.Y.; Kopytkova, L.A.; Kabulova, F.D.; Kushnarenko, S.V. Preservation of barberry (some species) genetic material in a cryobank. Bull. KarSU-Ser. Biol. Med. Geogr. 2019, 3, 20–26. (In Russian) [Google Scholar] [CrossRef]
- Romadanova, N.V.; Aralbaeva, M.M.; Rymkhanova, N.K.; Baigaraev, D.S.; Ramazanov, A.K.; Ishmuratova, M.I.; Kushnarenko, S.V. Cryopreservation as a way to improve laboratory germination and germination energy of seeds. Bull. KarSU-Ser. Biol. Med. Geogr. 2022, 1, 86–95. [Google Scholar] [CrossRef]
- Romadanova, N.V. (Institute of Plant Biology and Biotechnology, Almaty Region, Almaty, Kazakhstan). Personal communication, 2025.
- Hay, F.R.; Probert, R.J. Advances in seed conservation of wild plant species: A review of recent research. Conserv. Physiol. 2013, 1, cot030. [Google Scholar] [CrossRef] [PubMed]
- Ishikawa, K.; Harata, K.; Mii, M.; Sakai, A.; Yoshimatsu, K.; Shimomura, K. Cryopreservation of zygotic embryos of a Japanese terrestrial orchid (Bletilla striata) by vitrification. Plant Cell Rep. 1997, 16, 754–757. [Google Scholar] [CrossRef]
- Thammasiri, K. Cryopreservation of embryonic axes of jackfruit. Cryo-Lett. 1999, 20, 21–28. [Google Scholar]
- Tacán, M.; Andrango, R.; Tapia, C.; Sørensen, M.; Vollmer, R.; Pérez, C. Developing a cryopreservation protocol for embryonic axes of six south american peanut genotypes (Arachis hypogaea L.) using desiccation–vitrification. Crops 2024, 4, 701–716. [Google Scholar] [CrossRef]
- van der Walt, K.; Nadarajan, J.; Mathew, L.; Bettoni, J.C.; Souza, J.A. Advances in cryopreservation of Syzygium maire (swamp maire, maire tawake) zygotic embryos, a critically endangered tree species endemic to New Zealand. Front. Conserv. Sci. 2023, 4, 1269881. [Google Scholar] [CrossRef]
- FAO (Food and Agriculture Organization of the United Nations). Crops and Livestock Products. Production in 2022. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 27 March 2025).
- Lateur, M.; Dapena, E.; Szalatnay, D.; Gantar, M.E.; Guyader, A.; Hjalmarsson, I.; Höfer, M.; Ikase, L.; Kelerhals, M.; Lacis, G.; et al. ECPGR Characterization and Evaluation. Descriptors for Apple Genetic Resources; European Cooperative Pro-gramme for Plant Genetic Resources: Rome, Italy, 2022; p. 57. [Google Scholar]
- Knop, W. Quantitative investigations into the nutritional process of plants. Landwirtsch. Vers. 1865, 7, 93–107. [Google Scholar]
- Doyle, J.J.; Doyle, J.L. Isolation of plant DNA from fresh tissue. Focus 1990, 12, 13–15. [Google Scholar]
- SYSTAT. Statistics Software, SYSTAT 13.0; SYSTAT Software, Inc.: San Jose, CA, USA, 2009. Available online: https://systatsoftware.com/ (accessed on 26 February 2025).




{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Experimental Variant | N | HgCl2 Exposure Duration (min) | Indexing with Medium 523 | |||||
|---|---|---|---|---|---|---|---|---|
| 4 | 5 | 6 | 7 | 9 | ||||
| Viability of In Vitro Shoots (%) | N | Clean Shoots (%) | ||||||
| Crataegus ambigua | ||||||||
| E1 | 60 | - | 0 e | - | 0 e | - | - | - |
| E2/2 | 80 | 52.6 ± 8 ab | - | 7.3 ± 4 cd | - | 0 e | 15 | 40.3 ± 7 cd |
| E2/4 | 80 | 42.2 ± 5 b | - | 31.7 ± 7 bc | - | 7.7 ± 5 cd | 9 | 39.8 ± 5 cd |
| C1 | 80 | 0 e | - | 0 e | - | 0 e | 0 | - |
| Malus sieversii | ||||||||
| E1 | 60 | 29.4 ± 5 bc | - | 53.9 ± 4 ab | - | - | 16 | 72.8 ± 5 b |
| E2/1 | 60 | 23.7 ± 12 bc | - | 37.1 ± 4 b | - | - | 14 | 75.1 ± 14 b |
| E2/3 | 60 | 15.6 ± 12 c | - | 19.1 ± 5 c | - | - | 6 | 85.0 ± 6 b |
| C1 | 60 | 17.8 ± 13 c | - | 22.7 ± 12 bc | - | - | 6 | 66.7 ± 8 c |
| C2 | 60 | 12.8 ± 14 cd | - | 17.6 ± 12 c | - | - | 11 | 72.7 ± 7 b |
| Prunus tenella | ||||||||
| E2/2 | 80 | 22.5 ± 11 bc | - | - | 44.3 ± 7 b | 0 e | 12 | 71.4 ± 9 b |
| E2/4 | 80 | 14.9 ± 13 cd | - | - | 19.5 ± 13 c | 0 e | 5 | 72.8 ± 9 b |
| C1 | 60 | 0 e | - | - | 0 e | - | - | - |
| Sibiraea laevigata | ||||||||
| E1 | 60 | 61.5 ± 12 a | - | 38.2 ± 12 b | - | - | 30 | 100.0 ± 0 a |
| E2/1 | 60 | 56.2 ± 6 ab | - | 36.7 ± 13 b | - | - | 29 | 100.0 ± 0 a |
| E2/3 | 60 | 33.4 ± 14 bc | - | 25.8 ± 12 bc | - | - | 18 | 100.0 ± 0 a |
| C1 | 60 | 54.5 ± 13 ab | - | 34.0 ± 13 bc | - | - | 27 | 100.0 ± 0 a |
| C2 | 60 | 56.9 ± 12 ab | - | 37.3 ± 6 b | - | - | 28 | 100.0 ± 0 a |
| C3 | 60 | 27.1 ± 14 bc | - | 26.9 ± 5 bc | - | - | 16 | 100.0 ± 0 a |
| Spiraeanthus schrenckianus | ||||||||
| E1 | 60 | - | 27.3 ± 12 cd | - | 26.9 ± 12 bc | - | 19 | 100.0 ± 0 a |
| E2/1 | 60 | - | 41.1 ± 12 b | - | 35.6 ± 14 bc | - | 23 | 100.0 ± 0 a |
| E2/3 | 60 | - | 20.9 ± 15 c | - | 12.9 ± 12 cd | - | 10 | 100.0 ± 0 a |
| C1 | 60 | - | 0 e | - | 0 e | - | - | - |
| C2 | 60 | - | 30.2 ± 21 bc | - | 20.5 ± 12 c | - | 15 | 100.0 ± 0 a |
| C3 | 60 | - | 13.7 ± 12 cd | - | 9.5 ± 12 cd | - | 14 | 100.0 ± 0 a |
| Mean | 30.6 ± 18 | 20.5 ± 15 | 29.3 ± 13 | 16.9 ± 13 | 1.5 ± 3.4 | 14 | 84.8 ± 33 | |
| Plant Species, Accession | In Vitro Establishment of Shoots | Indexing with Medium 523 | |||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Clean Shoots (%) | |||||||||||||||
| Sprouted Cuttings in the Laboratory (E1) | N | Taken Directly from the Field (E2) | N | Sprouted Cuttings in the Laboratory (E1) | N | Taken Directly From the Field (E2) | N | ||||||||
| M. sieversii, 1 | 62 ± 13 | b | 39 | 30 ± 10 | c | 60 | 37.5 ± 8 | b | 24 | 31.1 ± 10 | b | 18 | |||
| M. sieversii, 2 | 51 ± 6 | b | 40 | 40 ± 9 | bc | 60 | 59.2 ± 14 | ab | 18 | 30.6 ± 6 | bc | 24 | |||
| M. sieversii, 3 | 48 ± 8 | bc | 32 | 34 ± 8 | c | 60 | 55.0 ± 18 | ab | 20 | 13.0 ± 12 | c | 20 | |||
| M. sieversii, 4 | 63 ± 10 | b | 26 | 52 ± 5 | b | 60 | 56.2 ± 19 | ab | 16 | 31.7 ± 8 | b | 31 | |||
| M. sieversii, 5 | 55 ± 13 | b | 28 | 56 ± 17 | b | 60 | 54.5 ± 18 | ab | 22 | 39.2 ± 14 | b | 23 | |||
| M. sieversii, 6 | 79 ± 10 | ab | 32 | 51 ± 13 | b | 60 | 62.2 ± 17 | a | 15 | 15.0 ± 9 | c | 30 | |||
| M. sieversii, 7 | 73 ± 8 | ab | 48 | 62 ± 12 | b | 60 | 63.0 ± 12 | a | 18 | 35.1 ± 13 | b | 37 | |||
| M. sieversii, 8 | 65 ± 8 | b | 33 | 55 ± 18 | b | 60 | 59.4 ± 9 | ab | 23 | 48.2 ± 13 | ab | 33 | |||
| Mean | 62 ± 11 | 48 ± 11 | 55.9 ± 13 | 30.5 ± 11 | |||||||||||
| P. ulmifolia, 1 | 45 ± 14 | bc | 29 | - | - | - | 52.2 ± 14 | ab | 13 | - | - | - | |||
| P. ulmifolia, 2 | 22 ± 8 | c | 37 | - | - | - | 47.8 ± 10 | ab | 8 | - | - | - | |||
| P. ulmifolia, 3 | 25 ± 9 | c | 56 | - | - | - | 57.8 ± 10 | ab | 14 | - | - | - | |||
| P. ulmifolia, 4 | 75 ± 14 | ab | 36 | - | - | - | 54.5 ± 19 | ab | 20 | - | - | - | |||
| P. ulmifolia, 5 | 0 ± 0 | d | 28 | - | - | - | - | - | - | - | - | - | |||
| P. ulmifolia, 6 | 100 ± 0 | a | 20 | - | - | - | 35.6 ± 10 | b | 19 | - | - | - | |||
| P. ulmifolia, 7 | 94 ± 5 | a | 22 | - | - | - | 60.0 ± 10 | a | 18 | - | - | - | |||
| Mean | 52 ± 39 | 51.3 ± 12 | |||||||||||||
| Culture Media | Crataegus ambigua | Prunus tenella | Sibiraea laevigata | Spiraeanthus schrenckianus | ||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N of Shoots | MR | N of Shoots | MR | N of Shoots | MR | N of Shoots | MR | |||||
| Int | Reg | Int | Reg | Int | Reg | Int | Reg | |||||
| 1 | 15 | 18 | 1.2 ± 0.1 e | 15 | 42 | 2.8 ± 0.3 cd | 15 | 39 | 2.6 ± 0.5 cd | 15 | 18 | 1.2 ± 0.4 e |
| 2 | 15 | 30 | 1.8 ± 0.2 de | 15 | 30 | 2.0 ± 0.3 d | 15 | 45 | 3.0 ± 0.4 c | 15 | 15 | 1.0 ± 0.1 e |
| 3 | 15 | 15 | 1.0 ± 0.1 e | 15 | 37 | 2.5 ± 0.4 cd | 15 | 66 | 4.4 ± 0.9 b | 15 | 16 | 1.3 ± 0.3 e |
| 4 | 15 | 18 | 1.2 ± 0.3 e | 15 | 23 | 1.5 ± 0.2 de | 15 | 83 | 5.5 ± 1.6 a | 15 | 15 | 1.0 ± 0.1 e |
| 5 | 15 | 17 | 1.1 ± 0.1 e | 15 | 27 | 1.8 ± 0.3 de | 15 | 39 | 2.6 ± 0.5 cd | 15 | 18 | 1.2 ± 0.4 e |
| 6 | 15 | 18 | 1.2 ± 0.3 e | 15 | 26 | 1.7 ± 0.2 de | 15 | 75 | 5.0 ± 1.5 ab | 15 | 52 | 3.5 ± 0.7 b |
| 7 | 15 | 15 | 1.0 ± 0.1 e | 15 | 27 | 1.8 ± 0.2 de | 15 | 69 | 4.6 ± 1.1 ab | 15 | 60 | 4.0 ± 0.9 b |
| 8 | 15 | 38 | 2.7 ± 0.4 cd | 15 | 36 | 2.4 ± 0.3 cd | 15 | 82 | 5.5 ± 1.4 a | - | - | |
| 9 | 15 | 50 | 3.3 ± 0.5 b | 15 | 39 | 2.6 ± 0.5 cd | - | - | - | - | ||
| 10 | 15 | 53 | 3.5 ± 0.7 b | 15 | 44 | 2.9 ± 0.6 cd | - | - | - | - | ||
| 11 | - | - | - | 15 | 48 | 3.2 ± 0.8 bc | - | - | - | - | ||
| 12 | - | - | - | 15 | 42 | 2.8 ± 0.3 cd | - | - | - | - | ||
| 13 | - | - | - | 15 | 33 | 2.2 ± 0.2 d | - | - | - | - | ||
| Mean | 15 | 28 | 1.8 ± 1.0 | 15 | 35 | 2.3 ± 0.8 | 15 | 62 | 4.2 ± 1.2 | 15 | 1.9 ± 1.3 | |
| Plant Species, Accession | Seed Storage at −20 °C | Seed Cryopreservation | Embryonic Axes Cryopreservation | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Viability (%) | N | Viability (%) | N | Viability (%) | N | ||||
| C. ambigua, 1 | - | - | - | - | - | - | 73.3 ± 7.3 | ab | 30 |
| C. ambigua, 2 | - | - | - | - | - | - | 60.0 ± 10.0 | bc | 30 |
| M. niedzwetzkyana, 1 | 93.3 ± 5.8 | a | 30 | 100.0 ± 0.0 | a | 30 | - | - | - |
| M. niedzwetzkyana, 2 | 96.7 ± 5.4 | a | 30 | 93.3 ± 6.2 | a | 30 | - | - | - |
| M. sieversii, 1 | 96.7 ± 6.1 | a | 30 | 100.0 ± 0.0 | a | 30 | 50.0 ± 10.0 | bc | 30 |
| M. sieversii, 2 | 96.7 ± 5.8 | a | 30 | 93.3 ± 6.8 | a | 30 | - | - | |
| M. sieversii, 3 | - | - | - | 86.7 ± 7.6 | ab | 30 | - | - | |
| M. sieversii, 4 | - | - | - | 93.3 ± 5.8 | a | 30 | 60.0 ± 17.3 | bc | 30 |
| M. sieversii, 5 | - | - | - | - | - | 66.7 ± 15.3 | bc | 30 | |
| P. tenella, 1 | - | - | - | - | - | - | 80.0 ± 9.2 | ab | 30 |
| P. tenella, 2 | - | - | - | - | - | - | 70.0 ± 10.0 | b | 30 |
| S. laevigata, 1 | 90.0 ± 7.3 | ab | 30 | 93.3 ± 5.8 | a | 30 | - | - | - |
| S. laevigata, 2 | 100.0 ± 0.0 | a | 30 | 100.0 ± 0.0 | a | 30 | - | - | - |
| S. laevigata, 3 | 83.3 ± 11.6 | ab | 30 | 100.0 ± 0.0 | a | 30 | - | - | - |
| S. laevigata, 4 | 93.3 ± 5.8 | a | 30 | 100.0 ± 0.0 | a | 30 | - | - | - |
| S. laevigata, 5 | 80.0 ± 17.3 | ab | 30 | 90.0 ± 7.3 | ab | 30 | - | - | - |
| S. schrenckianus, 1 | 66.7 ± 12.4 | bc | 30 | 93.3 ± 5.8 | a | 30 | - | - | - |
| S. schrenckianus, 2 | 50.0 ± 8.2 | c | 30 | 73.3 ± 11.6 | b | 30 | - | - | - |
| S. schrenckianus, 3 | 90.0 ± 6.6 | ab | 30 | 100.0 ± 0.0 | a | 30 | - | - | - |
| Mean | 86.4 ± 12.7 | 94.0 ± 7.4 | 65.7 ± 9.9 | ||||||
| Species | Number of Accessions | GPS Coordinates, Elevations, m | Place of Collection, year |
|---|---|---|---|
| Crataegus ambigua | 10 | N44°27′.42.2″–N44°27′45.8″ E50°35′. 22.2″–E50°35′ 25.1″, 51–126 m | P1 *—Mangistau region, Kizim tract, 2023 |
| 10 | N44°28′.26.3″–N44°28′.27.5″ E51°00′. 45.4″–E51°00′. 50.1″, 58–81 m | P2—Mangistau region, gorge. “Sultan Uly”, 2023 | |
| 10 | N43°39′.02.7″–N43°39′.55.8″ E50°35′. 22.2″–E50°35′. 25.1″, 4–12 m | P3—Mangistau region, Mangyshlak experimental botanical garden, 2023 | |
| 10 | N44°12′.53.8″–N44°12′.55.5″ E51°59′. 29.1″–E51°59′. 30.8″, 252–277 m | P4—Mangistau region, Western Karatau, Samal gorge, 2023 | |
| Cotoneaster karatavicus | 21 | N42°36′.064″–N42°40′.044″ E070°13′.444″–E070°23′.508″, 837–844 m | P1—Turkestan region, Sairam-Ugam national park, Kokbulak gorge, 2024 |
| 9 | N42°36′.123″–N42°36′.574″ E070°23′.671″–E070°24′.047″, 932–956 m | P2—Turkestan region, Tyulkubas district, Kulan reservoir, 2024 | |
| 9 | N43°05′.101″–N43°05′.146″ E069°55′.351″–E069°55′.388″, 715–716 m | 3—Turkestan region, Bayzhansai gorge, Ulken Buken river, 2024 | |
| Malus niedzwetzkyana na | 2 | N43°17′.896″–N43°17′.880″ E079°30′.902″–E079°30′.912″, 1646–1664 m | P1—Almaty region, Uyghur district, gorge. “Bol’shoy Kyrgyzsai”, 2024 |
| 1 | N43°40′.1080″, E076°59′.8702″, 600 m | P2—Almaty region, Ili district, 2024 | |
| M. sieversii | 35 | N43°17′.118″–N43°23′.036″ E77°34′.394″–E77°39′.850″, 819–1343 m | P1—Enbekshikazakh district, Almaty region, Turgen village, 2023 |
| 19 | N43°17′.938″– N43°20′.749″ E079°29′.802″–E079°37′.927″, 1585–1657 m | P2—Almaty region, Uyghur district, gorge. “Kyrgyzsai”, 2023–2024 | |
| 1 | N43°38′.240″, E080°03′.255″, 745 m | P3—Almaty region, Uyghur district, Chundzha-Kolzhat highway, 2024 | |
| Prunus tenella | 20 | N49°42′.460″–N49°42′.466″ E083°43′.463″–E083°43′.492″, 710–710 m | P1—East Kazakhstan region, western Altai, vicinity of Pribrezhny vil., 2023 |
| 1 | N50°32′.305″, E083°54′.190″, 512 m | P2—East Kazakhstan region, outskirts of Ridder, 2024 | |
| P. ulmifolia | 15 | N43°13′29.106″, E76°54′51.132″, 867–880 m | Almaty region, Main Botanical Garden, 2023–2024 |
| Sibiraea laevigata | 10 | N50°19′.254″–N50°19′.290″ E83°32′.514″–E083°33′.083″, 730–793 m | P1—East Kazakhstan region, Ridder city, Bystrukha river valley, 2023 |
| 21 | N50°17′.826″–N50°17′.855″ E083°29′.310″–E083°29′.379″, 873–880 m | P2—East Kazakhstan region, Ridder city, Gromotukha river floodplain, 2024 | |
| Sorbus persica | 21 | N42°24′.423″–N42°53′.915″ E70°34′.813″–E70°34′.874″,1752–1883 m | P1—–Turkestan region, Aksu Zhabagly State Nature Reserve, Kshi-Kaindy gorge, 2023 |
| 5 | N42°53′.910″–N42°53′.915″ E70°00′.086″–E70°00′.091″, 1044–1052 m | P2—Turkestan region, gorge “Aule Karabastau”, 2023 | |
| Spiraeanthus schrenkianus us | 12 | N42°44′35.33″–N42°44′39.81″ E71°00′01.53′’–E71°00′20.97′’, 1055–1079 m | P1—Zhambyl region, Zhualynsky district, Kuyuk pass, 2023 |
| 8 | N42°58′.608″–N42°58′.623″ E69°40′.705″–E69°40′.707″, 527–538 m | P2—Turkestan region, Karatau foothills, Ak-Mechet cave, 2023 |
| Accession | Variants of Media * |
| Crataegus ambigua | (1) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA (2) MS with 30 g L−1 sucrose, 0.2 mg L−1 BAP, 0.01 mg L−1 IBA (3) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.1 mg L−1 IBA (4) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.02 mg L−1 IBA (5) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.03 mg L−1 IBA (6) MS with 30 g L−1 sucrose, 0.1 mg L−1 BAP, 0.01 mg L−1 IBA (7) MS with 30 g L−1 sucrose, 0.1 mg L−1 BAP, 0.1 mg L−1 IBA (8) ½ MS with 30 g L−1 sucrose, 0.2 mg L−1 BAP, 0.01 mg L−1 IBA (9) ½ MS with 30 g L−1 sucrose, 0.2 mg L−1 BAP, 0.01 mg L−1 IBA, 0.1 g L−1 Sequestrene 138 Fe (10) ½ MS with 30 g L−1 sucrose, 0.2 mg L−1 BAP, 0.01 mg L−1 IBA, 0.1 g L−1 Sequestrene 138 Fe, 1.0 mg L−1 Vitamin C |
| Prunus tenella | (1) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA, 7.0 mg L−1 FeSO4·7H2O, 9.3 mg L−1 Na2EDTA 2H2O, 166 mg L−1 CaCl2 (2) ½ MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA, 7.0 mg L−1 FeSO4·7H2O, 9.3 mg L−1 Na2EDTA 2H2O, 166 mg L−1 CaCl (3) ½ MS with 20 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA, 7.0 mg L−1 FeSO4·7H2O, 9.3 mg L−1 Na2EDTA 2H2O, 166 mg L−1 CaCl2; (4) ½ MS with 20 g L−1 sucrose, 1.0 mg L−1 BAP, 0.01 mg L−1 IBA, 7.0 mg L−1 FeSO4·7H2O, 9.3 mg L−1 Na2EDTA 2H2O, 166 mg L−1 CaCl2 (5) ½ MS with 20 g L−1 sucrose, 1.0 mg L−1 BAP, 0.05 mg L−1 IBA, 7.0 mg L−1 FeSO4·7H2O, 9.3 mg L−1 Na2EDTA 2H2O, 166 mg L−1 CaCl2 (6) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA (7) MS with 20 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA (8) ½ MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA (9) ½ MS with 20 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA (10) ½ MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA, 7.0 mg L−1 FeSO4·7H2O, 9.3 mg L−1 Na2EDTA 2H2O, 166 mg L−1 CaCl2 (11) ½ MS with 30 g L−1 sucrose, 0.25 mg L−1 BAP, 0.01 mg L−1 IBA, 7.0 mg L−1 FeSO4·7H2O, 9.3 mg L−1 Na2EDTA 2H2O, 166 mg L−1 CaCl2 (12) ½ MS with 30 g L−1 sucrose, 0.25 mg L−1 BAP, 7.0 mg L−1 FeSO4·7H2O, 9.3 mg L−1 Na2EDTA 2H2O, 166 mg L−1 CaCl2 (13) ½ MS with 30 g L−1 sucrose, 7.0 mg L−1 FeSO4·7H2O, 9.3 mg L−1 Na2EDTA 2H2O, 166 mg L−1 CaCl2 |
| Sibiraea laevigata | (1) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA (2) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA, 0.1 mg L−1 GA3 (3) MS with 30 g L−1 sucrose, 1.0 mg L−1 BAP, 0.01 mg L−1 IBA, 0.1 mg L−1 GA3 (4) MS with 30 g L−1 sucrose, 1.0 mg L−1 BAP, 0.01 mg L−1 IBA, 0.2 mg L−1 GA3 (5) MS with 30 g L−1 sucrose, 1.0 mg L−1 BAP, 0.01 mg L−1 IBA, 0.3 mg L−1 GA3 (6) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.1 mg L−1 IBA, 0.1 mg L−1 GA3 (7) MS with 30 g L−1 sucrose, 1.0 mg L−1 BAP, 0.1 mg L−1 IBA, 0.1 mg L−1 GA3 (8) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA, 0.2 mg L−1 GA3 |
| Spiraeanthus schrenkianus | (1) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA (2) MS with 30 g L−1 sucrose, 1.0 mg L−1 BAP, 0.01 mg L−1 IBA (3) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.1 mg L−1 IBA (4) MS with 30 g L−1 sucrose, 1.0 mg L−1 BAP, 0.1 mg L−1 IBA (5) MS with 30 g L−1 sucrose, 0.5 mg L−1 BAP, 0.01 mg L−1 IBA, 0.2 mg L−1 GA3 (6) MS with 30 g L−1 sucrose, 0.2 mg L−1 BAP, 0.02 mg L−1 IBA (7) MS with 30 g L−1 sucrose, 0.1 mg L−1 BAP, 0.01 mg L−1 IBA |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Romadanova, N.V.; Zemtsova, A.S.; Altayeva, N.A.; Artimovich, N.A.; Alexandrova, A.M.; Kushnarenko, S.V.; Bettoni, J.C. Geobotanical Study and Preservation of Rare and Endangered Rosaceae Species. Plants 2025, 14, 1526. https://doi.org/10.3390/plants14101526
Romadanova NV, Zemtsova AS, Altayeva NA, Artimovich NA, Alexandrova AM, Kushnarenko SV, Bettoni JC. Geobotanical Study and Preservation of Rare and Endangered Rosaceae Species. Plants. 2025; 14(10):1526. https://doi.org/10.3390/plants14101526
Chicago/Turabian StyleRomadanova, Natalya V., Alina S. Zemtsova, Nazira A. Altayeva, Natalya A. Artimovich, Alyona M. Alexandrova, Svetlana V. Kushnarenko, and Jean Carlos Bettoni. 2025. "Geobotanical Study and Preservation of Rare and Endangered Rosaceae Species" Plants 14, no. 10: 1526. https://doi.org/10.3390/plants14101526
APA StyleRomadanova, N. V., Zemtsova, A. S., Altayeva, N. A., Artimovich, N. A., Alexandrova, A. M., Kushnarenko, S. V., & Bettoni, J. C. (2025). Geobotanical Study and Preservation of Rare and Endangered Rosaceae Species. Plants, 14(10), 1526. https://doi.org/10.3390/plants14101526