
Isolation and Expression Pattern Analysis of Larix olgensis LoNAC5: LoNAC5 Acts as a Positive Regulator of Drought and Salt Tolerance

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
1. Introduction
2. Results
2.1. LoNAC5 Belongs to the AtNAC3 Subgroup
2.2. LoNAC5 Conforms to the Characteristics of NAC Transcription Factors
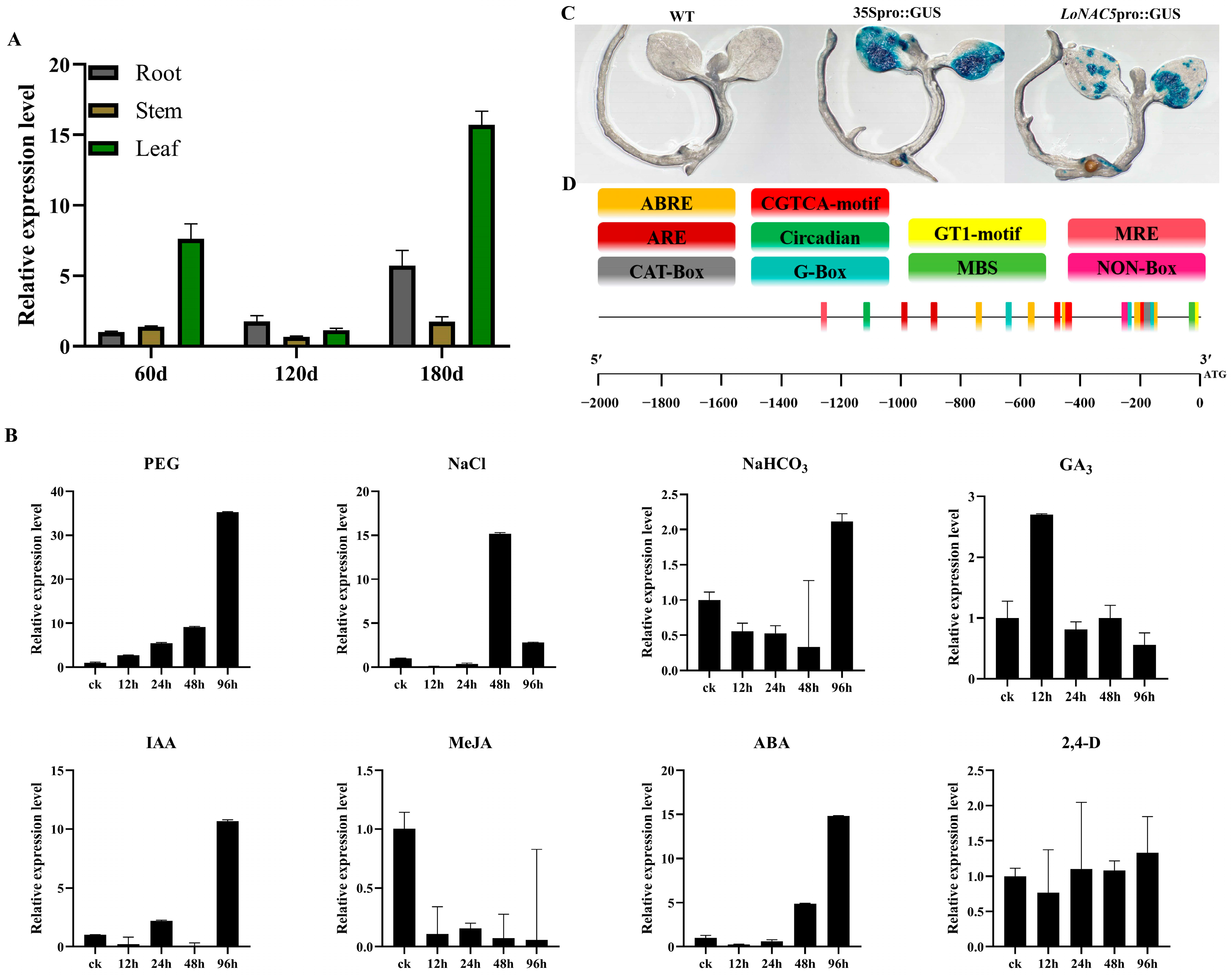
2.3. LoNAC5 Can Respond to Drought and Salt Stress
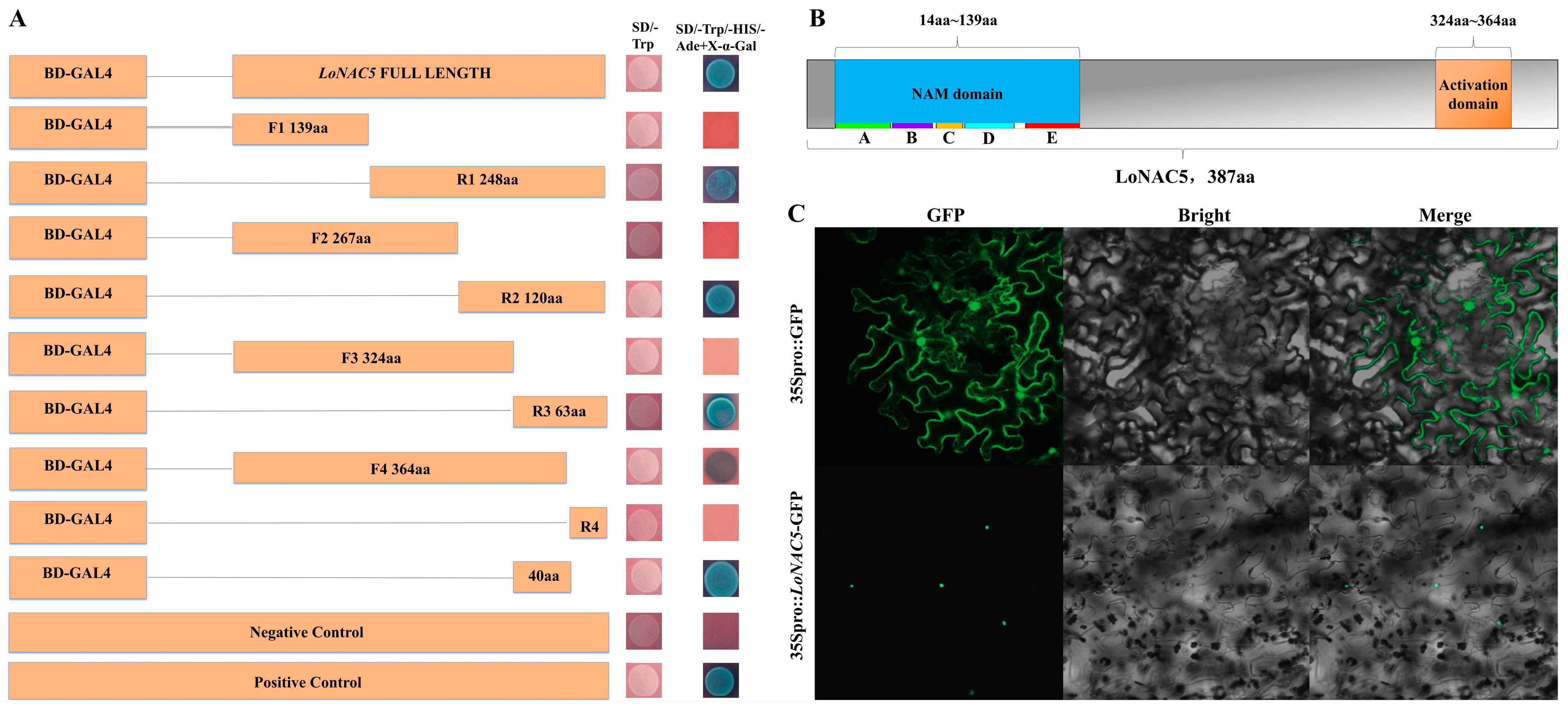
2.4. Functional Characterization of LoNAC5 in Yeast
2.5. LoNAC5 Can Form Homodimers and Bind to CATGTG Motif
3. Discussion
4. Materials and Methods
4.1. Isolation of LoNAC5 and LoNAC5 Promoter
4.2. Web Tools and Software Used for Bioinformatics Analysis
4.3. Analysis of LoNAC5 Promoter Activity
4.4. Subcellular Localization of LoNAC5
4.5. Research on Mechanism of LoNAC5
4.6. Functional Characterization of LoNAC5
4.7. Analysis of LoNAC5 Expression in L. olgensis
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Yamasaki, K.; Kigawa, T.; Seki, M.; Shinozaki, K.; Yokoyama, S. DNA-binding domains of plant-specific transcription factors: Structure, function, and evolution. Trends Plant Sci. 2013, 18, 267–276. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Li, B.; Rizwan, H.M.; Sun, K.; Zeng, J.; Shi, M.; Guo, T.; Chen, F. Genome-wide identification and comprehensive analyses of NAC transcription factor gene family and expression analysis under fusarium kyushuense and drought stress conditions in passiflora edulis. Front. Plant Sci. 2022, 13, 972734. [Google Scholar] [CrossRef]
- Zafar, Z.; Fatima, S.; Bhatti, M.F. Comprehensive analyses of NAC transcription factor family in almond (prunus dulcis) and their differential gene expression during fruit development. Plants 2021, 10, 2200. [Google Scholar] [CrossRef]
- Olsen, A.N.; Ernst, H.A.; Leggio, L.L.; Skriver, K. NAC transcription factors: Structurally distinct, functionally diverse. Trends Plant Sci. 2005, 10, 79–87. [Google Scholar] [CrossRef]
- Wang, J.Y.; Wang, J.P.; Yang, H.F. Identification and functional characterization of the NAC gene promoter from populus euphratica. Planta 2016, 244, 417–427. [Google Scholar] [CrossRef] [PubMed]
- Tran, L.S.; Nakashima, K.; Sakuma, Y.; Simpson, S.D.; Fujita, Y.; Maruyama, K.; Fujita, M.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Isolation and functional analysis of Arabidopsis stress-inducible NAC transcription factors that bind to a drought-responsive cis-element in the early responsive to dehydration stress 1 promoter. Plant Cell 2004, 16, 2481–2498. [Google Scholar] [CrossRef]
- Souer, E.; van Houwelingen, A.; Kloos, D.; Mol, J.; Koes, R. The no apical meristem gene of petunia is required for pattern formation in embryos and flowers and is expressed at meristem and primordia boundaries. Cell 1996, 85, 159–170. [Google Scholar] [CrossRef] [PubMed]
- Liu, L.; White, M.J.; MacRae, T.H. Transcription factors and their genes in higher plants functional domains, evolution and regulation. Eur. J. Biochem. 1999, 262, 247–257. [Google Scholar] [CrossRef]
- Taoka, K.; Yanagimoto, Y.; Daimon, Y.; Hibara, K.; Aida, M.; Tasaka, M. The NAC domain mediates functional specificity of cup-shaped cotyledon proteins. Plant J. 2004, 40, 462–473. [Google Scholar] [CrossRef]
- Riechmann, J.L.; Heard, J.; Martin, G.; Reuber, L.; Jiang, C.; Keddie, J.; Adam, L.; Pineda, O.; Ratcliffe, O.J.; Samaha, R.R.; et al. Arabidopsis transcription factors: Genome-wide comparative analysis among eukaryotes. Science 2000, 290, 2105–2110. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Sharoni, A.M.; Satoh, K.; Moumeni, A.; Venuprasad, R.; Serraj, R.; Kumar, A.; Leung, H.; Attia, K.; Kikuchi, S. Comprehensive gene expression analysis of the NAC gene family under normal growth conditions, hormone treatment, and drought stress conditions in rice using near-isogenic lines (nils) generated from crossing aday selection (drought tolerant) and ir64. Mol. Genet. Genom. 2012, 287, 389–410. [Google Scholar] [CrossRef] [PubMed]
- Kikuchi, K.; UeguchiTanaka, M.; Yoshida, K.T.; Nagato, Y.; Matsusoka, M.; Hirano, H.Y. Molecular analysis of the NAC gene family in rice. Mol. Gen. Genet. 2000, 262, 1047–1051. [Google Scholar] [CrossRef]
- Fang, Y.; You, J.; Xie, K.; Xie, W.; Xiong, L. Systematic sequence analysis and identification of tissue-specific or stress-responsive genes of NAC transcription factor family in rice. Mol. Genet. Genom. 2008, 280, 547–563. [Google Scholar] [CrossRef]
- Ooka, H.; Satoh, K.; Doi, K.; Nagata, T.; Otomo, Y.; Murakami, K.; Matsubara, K.; Osato, N.; Kawai, J.; Carninci, P.; et al. Comprehensive analysis of NAC family genes in Oryza sativa and Arabidopsis thaliana. DNA Res. 2003, 10, 239–247. [Google Scholar] [CrossRef]
- Hu, R.; Qi, G.; Kong, Y.; Kong, D.; Gao, Q.; Zhou, G. Comprehensive analysis of NAC domain transcription factor gene family in Populus trichocarpa. BMC Plant Biol. 2010, 10, 145. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Zi, Y.; Yang, X.; Yang, X.; Zhu, L.; Cai, H.; Yin, T.; Liu, X.; Zhang, H. Genome-wide analysis of the CsAP2/ERF gene family of sweet orange (Citrus sinensis) and joint analysis of transcriptional metabolism under salt stress. Ann. Bot. 2025, mcaf 006, 39847492. [Google Scholar] [CrossRef]
- Fang, S.; Shang, X.; Yao, Y.; Li, W.; Guo, W. NST- and SND-subgroup NAC proteins coordinately act to regulate secondary cell wall formation in cotton. Plant Sci. 2020, 301, 110657. [Google Scholar] [CrossRef] [PubMed]
- Vroemen, C.W.; Mordhorst, A.P.; Albrecht, C.; Kwaaitaal, M.A.; de Vries, S.C. The CUP-shaped cotyledon3 gene is required for boundary and shoot meristem formation in Arabidopsis. Plant Cell 2003, 15, 1563–1577. [Google Scholar] [CrossRef]
- Sun, S.; Li, X.; Nie, N.; Chen, Y.; Gao, S.; Zhang, H.; He, S.; Liu, Q.; Zhai, H. Sweet potato NAC transcription factor NAC43 negatively regulates plant growth by causing leaf curling and reducing photosynthetic efficiency. Front. Plant Sci. 2023, 14, 1095977. [Google Scholar] [CrossRef]
- Hao, Y.J.; Wei, W.; Song, Q.X.; Chen, H.W.; Zhang, Y.Q.; Wang, F.; Zou, H.F.; Lei, G.; Tian, A.G.; Zhang, W.K.; et al. Soybean NAC transcription factors promote abiotic stress tolerance and lateral root formation in transgenic plants. Plant J. 2011, 68, 302–313. [Google Scholar] [CrossRef]
- Pooam, M.; El-Ballat, E.M.; Jourdan, N.; Ali, H.M.; Hano, C.; Ahmad, M.; El-Esawi, M.A. SNAC3 transcription factor enhances arsenic stress tolerance and grain yield in rice (Oryza sativa L.) Through regulating physio-biochemical mechanisms, stress-responsive genes, and cryptochrome 1b. Plants 2023, 12, 2731. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, B.; Cheng, C.; Fu, B.; Qi, M.; Du, H.; Geng, S.; Zhang, X. Comparative transcriptomics analysis reveals the differences in transcription between resistant and susceptible pepper (Capsicum annuum L.) varieties in response to anthracnose. Plants 2024, 13, 527. [Google Scholar] [CrossRef] [PubMed]
- Cucinotta, M.; Manrique, S.; Cuesta, C.; Benkova, E.; Novak, O.; Colombo, L. CUP-shaped cotyledon1 (CUC1) and CUC2 regulate cytokinin homeostasis to determine ovule number in Arabidopsis. J. Exp. Bot. 2018, 69, 5169–5176. [Google Scholar] [CrossRef]
- Li, Y.; Xia, T.; Gao, F.; Li, Y. Control of plant branching by the CUC2/CUC3-DA1-UBP15 regulatory module. Plant Cell 2020, 32, 1919–1932. [Google Scholar] [CrossRef] [PubMed]
- Zimmermann, R.; Werr, W. Pattern formation in the monocot embryo as revealed by NAM and CUC3 orthologues from Zea mays L. Plant Mol. Biol. 2005, 58, 669–685. [Google Scholar] [CrossRef]
- Kubo, M.; Udagawa, M.; Nishikubo, N.; Horiguchi, G.; Yamaguchi, M.; Ito, J.; Mimura, T.; Fukuda, H.; Demura, T. Transcription switches for protoxylem and metaxylem vessel formation. Genes Dev. 2005, 19, 1855–1860. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, M.; Kubo, M.; Fukuda, H.; Demura, T. Vascular-related NAC-domain7 is involved in the differentiation of all types of xylem vessels in Arabidopsis roots and shoots. Plant J. 2008, 55, 652–664. [Google Scholar] [CrossRef]
- Yamaguchi, M.; Ohtani, M.; Mitsuda, N.; Kubo, M.; Ohme-Takagi, M.; Fukuda, H.; Demura, T. VND-interacting2, a NAC domain transcription factor, negatively regulates xylem vessel formation in Arabidopsis. Plant Cell 2010, 22, 1249–1263. [Google Scholar] [CrossRef]
- Tan, T.T.; Endo, H.; Sano, R.; Kurata, T.; Yamaguchi, M.; Ohtani, M.; Demura, T. Transcription factors VND1-VND3 contribute to cotyledon xylem vessel formation. Plant Physiol. 2018, 176, 773–789. [Google Scholar] [CrossRef]
- Xie, Q.; Frugis, G.; Colgan, D.; Chua, N.H. Arabidopsis NAC1 transduces auxin signal downstream of TIR1 to promote lateral root development. Genes Dev. 2000, 14, 3024–3036. [Google Scholar] [CrossRef]
- Lu, M.; Ying, S.; Zhang, D.F.; Shi, Y.S.; Song, Y.C.; Wang, T.Y.; Li, Y. A maize stress-responsive NAC transcription factor, ZmsNAC1, confers enhanced tolerance to dehydration in transgenic Arabidopsis. Plant Cell Rep. 2012, 31, 1701–1711. [Google Scholar] [CrossRef]
- Jin, X.; Chai, Q.; Liu, C.; Niu, X.; Li, W.; Shang, X.; Gu, A.; Zhang, D.; Guo, W. Cotton GhNAC4 promotes drought tolerance by regulating secondary cell wall biosynthesis and ribosomal protein homeostasis. Plant J. 2024, 117, 1052–1068. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Wang, L.Y.; Dai, J.X.; Wang, Y.; Lin, D. The NAC-type transcription factor CaNAC46 regulates the salt and drought tolerance of transgenic Arabidopsis thaliana. BMC Plant Biol. 2021, 21, 11. [Google Scholar] [CrossRef] [PubMed]
- Shinde, H.; Dudhate, A.; Tsugama, D.; Gupta, S.K.; Liu, S.; Takano, T. Pearl millet stress-responsive NAC transcription factor PgNAC21 enhances salinity stress tolerance in Arabidopsis. Plant Physiol. Biochem. 2019, 135, 546–553. [Google Scholar] [CrossRef]
- Wang, R.; Zhang, Y.; Wang, C.; Wang, Y.C.; Wang, L.Q. ThNAC12 from Tamarix hispida directly regulates ThPIP2.5 to enhance salt tolerance by modulating reactive oxygen species. Plant Physiol. Biochem. 2021, 163, 27–35. [Google Scholar] [CrossRef]
- Zhang, H.; Pei, Y.; Zhu, F.; He, Q.; Zhou, Y.; Ma, B.; Chen, X.; Guo, J.; Khan, A.; Jahangir, M.; et al. CaSNRK2.4-mediated phosphorylation of CaNAC035 regulates abscisic acid synthesis in pepper (Capsicum annuum L.) Responding to cold stress. Plant J. 2024, 117, 1377–1391. [Google Scholar] [CrossRef] [PubMed]
- Shan, W.; Kuang, J.F.; Lu, W.J.; Chen, J.Y. Banana fruit NAC transcription factor MaNAC1 is a direct target of MaICE1 and involved in cold stress through interacting with MaCBF1. Plant Cell Environ. 2014, 37, 2116–2127. [Google Scholar] [CrossRef]
- Yan, T.; Sun, M.; Su, R.; Wang, X.; Lu, X.; Xiao, Y.; Deng, H.; Liu, X.; Tang, W.; Zhang, G. Transcriptomic profiling of cold stress-induced differentially expressed genes in seedling stage of indica rice. Plants 2023, 12, 2675. [Google Scholar] [CrossRef]
- Fang, Y.; Liao, K.; Du, H.; Xu, Y.; Song, H.; Li, X.; Xiong, L. A stress-responsive NAC transcription factor SNAC3 confers heat and drought tolerance through modulation of reactive oxygen species in rice. J. Exp. Bot. 2015, 66, 6803–6817. [Google Scholar] [CrossRef]
- Ren, T.; Qu, F.; Morris, T.J. Hrt gene function requires interaction between a NAC protein and viral capsid protein to confer resistance to turnip crinkle virus. Plant Cell 2000, 12, 1917–1926. [Google Scholar] [CrossRef]
- Xie, Q.; Sanz-Burgos, A.P.; Guo, H.; Garcia, J.A.; Gutierrez, C. Grab proteins, novel members of the NAC domain family, isolated by their interaction with a geminivirus protein. Plant Mol. Biol. 1999, 39, 647–656. [Google Scholar] [CrossRef] [PubMed]
- Selth, L.A.; Dogra, S.C.; Rasheed, M.S.; Healy, H.; Randles, J.W.; Rezaian, M.A. A NAC domain protein interacts with tomato leaf curl virus replication accessory protein and enhances viral replication. Plant Cell 2005, 17, 311–325. [Google Scholar] [CrossRef]
- Wang, S.; Han, H.; Zhang, B.; Wang, L.; Wu, J.; Chen, Z.; Lin, K.; Hao, J.; Jia, R.; Zhang, Y. Identification of crucial genes and regulatory pathways in alfalfa against fusarium root rot. Plants 2023, 12, 3634. [Google Scholar] [CrossRef]
- Oh, S.K.; Lee, S.; Yu, S.H.; Choi, D. Expression of a novel NAC domain-containing transcription factor (CaNAC1) is preferentially associated with incompatible interactions between chili pepper and pathogens. Planta 2005, 222, 876–887. [Google Scholar] [CrossRef]
- Lee, M.H.; Jeon, H.S.; Kim, H.G.; Park, O.K. An Arabidopsis NAC transcription factor NAC4 promotes pathogen-induced cell death under negative regulation by microrna164. New Phytol. 2017, 214, 343–360. [Google Scholar] [CrossRef]
- Jensen, M.K.; Rung, J.H.; Gregersen, P.L.; Gjetting, T.; Fuglsang, A.T.; Hansen, M.; Joehnk, N.; Lyngkjaer, M.F.; Col-linge, D.B. The HvNAC6 transcription factor: A positive regulator of penetration resistance in barley and Arabidopsis. Plant Mol. Biol. 2007, 65, 137–150. [Google Scholar] [CrossRef] [PubMed]
- Chen, S.P.; Kuo, C.H.; Lu, H.H.; Lo, H.S.; Yeh, K.W. The sweet potato NAC-domain transcription factor IbNAC1 is dynamically coordinated by the activator IbBHLH3 and the repressor IbBHLH4 to reprogram the defense mechanism against wounding. PLoS Genet. 2016, 12, e1006397. [Google Scholar] [CrossRef]
- Yoshii, M.; Shimizu, T.; Yamazaki, M.; Higashi, T.; Miyao, A.; Hirochika, H.; Omura, T. Disruption of a novel gene for a NAC-domain protein in rice confers resistance to rice dwarf virus. Plant J. 2009, 57, 615–625. [Google Scholar] [CrossRef] [PubMed]
- Cenci, A.; Guignon, V.; Roux, N.; Rouard, M. Genomic analysis of NAC transcription factors in banana (Musa acuminata) and definition of NAC orthologous groups for monocots and dicots. Plant Mol. Biol. 2014, 85, 63–80. [Google Scholar] [CrossRef]
- Wang, N.; Zheng, Y.; Xin, H.; Fang, L.; Li, S. Comprehensive analysis of NAC domain transcription factor gene family in Vitis vinifera. Plant Cell Rep. 2013, 32, 61–75. [Google Scholar] [CrossRef]
- Yang, J.; Xu, J.; Xue, J.; Zhu, L. Expression and functional analysis of NAC transcription factors under five diverse growth stages reveal their regulatory roles during wood formation in chinese cedar (Cryptomeria Fortunei hooibrenk). Ind. Crops Prod. 2023, 197, 12. [Google Scholar] [CrossRef]
- Cao, Q.; An, P.; Zhang, S.; Wang, J.; Zhang, H.; Zhang, L. Preliminary analysis of two NAC transcription factor expression patterns in Larix olgensis. J. For. Res. 2022, 33, 601–609. [Google Scholar] [CrossRef]
- Cao, Q.; Hao, J.; Zhang, T.; Liu, L.; Xu, D.; Wang, C.; Zhao, Q.; Zhang, H.; Zhang, L. Isolation and functional analysis of the Larix olgensis LoNAC3 transcription factor gene. BMC Plant Biol. 2024, 24, 881. [Google Scholar] [CrossRef] [PubMed]
- Cai, K.; Song, X.; Yue, W.; Liu, L.; Ge, F.; Wang, J. Identification and functional characterization of abiotic stress tolerance-related platz transcription factor family in barley (Hordeum vulgare L.). Int. J. Mol. Sci. 2024, 25, 191. [Google Scholar] [CrossRef]
- He, L.; Xu, J.; Wang, Y.; Yang, K. Transcription factor ANAC074 binds to NRS1, NRS2, or MYBST1 element in addition to the NACRS to regulate gene expression. Int. J. Mol. Sci. 2018, 19, 3271. [Google Scholar] [CrossRef]
- Lin, R.; Zhao, W.; Meng, X.; Wang, M.; Peng, Y. Rice gene OsNAC19 encodes a novel NAC-domain transcription factor and responds to infection by magnaporthe grisea. Plant Sci. 2007, 172, 120–130. [Google Scholar] [CrossRef]
- Xia, N.; Zhang, G.; Sun, Y.; Zhu, L.; Xu, L.; Chen, X.; Liu, B.; Yu, Y.; Wang, X.; Huang, L.; et al. TaNAC8, a novel NAC transcription factor gene in wheat, responds to stripe rust pathogen infection and abiotic stresses. Physiol. Mol. Plant Pathol. 2010, 74, 394–402. [Google Scholar] [CrossRef]
- Peng, H.; Cheng, H.; Yu, X.; Shi, Q.; Zhang, H.; Li, J.; Ma, H. Characterization of a chickpea (Cicer arietinum L.) NAC family gene, CarNAC5, which is both developmentally-and stress-regulated. Plant Physiol. Biochem. 2009, 47, 1037–1045. [Google Scholar] [CrossRef] [PubMed]
- Meng, C.; Cai, C.; Zhang, T.; Guo, W. Characterization of six novel NAC genes and their responses to abiotic stresses in Gossypium hirsutum L. Plant Sci. 2009, 176, 352–359. [Google Scholar] [CrossRef]
- Ray, S.; Basnet, A.; Bhattacharya, S.; Banerjee, A.; Biswas, K. A comprehensive analysis of NAC gene family in Oryza sativa japonica: A structural and functional genomics approach. J. Biomol. Struct. Dyn. 2023, 41, 856–870. [Google Scholar] [CrossRef]
- Kadier, Y.; Zu, Y.; Dai, Q.; Song, G.; Lin, S.; Sun, Q.; Pan, J.; Lu, M. Genome-wide identification, classification and expression analysis of NAC family of genes in sorghum [Sorghum bicolor (L.) Moench]. Plant Growth Regul. 2017, 83, 301–312. [Google Scholar] [CrossRef]
- Li, X.; Li, N.; Wen, D.; Yu, J.; Hong, J.; Wu, M.; Cheng, L.; Meng, S. Identification of stress responsive NAC genes in Casuarina equisetifolia L. and its expression analysis under abiotic stresses. Agronomy 2024, 14, 535. [Google Scholar] [CrossRef]
- Takasaki, H.; Maruyama, K.; Takahashi, F.; Fujita, M.; Yoshida, T.; Nakashima, K.; Myouga, F.; Toyooka, K.; Yamaguchi-Shinozaki, K.; Shinozaki, K. SNAC-as, stress-responsive NAC transcription factors, mediate ABA-inducible leaf senescence. Plant J. 2015, 84, 1114–1123. [Google Scholar] [CrossRef]
- Han, X.; Feng, Z.; Xing, D.; Yang, Q.; Wang, R.; Qi, L.; Li, G. Two NAC transcription factors from Caragana intermedia altered salt tolerance of the transgenic Arabidopsis. BMC Plant Biol. 2015, 15, 208. [Google Scholar] [CrossRef]
- Zhang, H.; Cui, X.; Guo, Y.; Luo, C.; Zhang, L. Picea wilsonii transcription factor NAC2 enhanced plant tolerance to abiotic stress and participated in Rfcp1-regulated flowering time. Plant Mol. Biol. 2018, 98, 471–493. [Google Scholar] [CrossRef]
- Zhang, S.; Yan, S.; An, P.; Cao, Q.; Wang, C.; Wang, J.; Zhang, H.; Zhang, L. Embryogenic callus induction from immature zygotic embryos and genetic transformation of Larix kaempferi 3x Larix gmelinii 9. PLoS ONE 2021, 16, 26. [Google Scholar] [CrossRef] [PubMed]
- Ruiz-Partida, R.; Rosario, S.M.; Lozano-Juste, J. An update on crop aba receptors. Plants 2021, 10, 1087. [Google Scholar] [CrossRef]
- Mohamed, H.I.; Latif, H.H. Improvement of drought tolerance of soybean plants by using methyl jasmonate. Physiol. Mol. Biol. Plants 2017, 23, 545–556. [Google Scholar] [CrossRef]
- Bu, Q.; Jiang, H.; Li, C.; Zhai, Q.; Zhang, J.; Wu, X.; Sun, J.; Xie, Q.; Li, C. Role of the Arabidopsis thaliana NAC transcription factors ANAC019 and ANAC055 in regulating jasmonic acid-signaled defense responses. Cell Res. 2008, 18, 756–767. [Google Scholar] [CrossRef]
- Jiang, H.; Li, H.; Bu, Q.; Li, C. The RHA2a-interacting proteins ANAC019 and ANAC055 may play a dual role in regulating aba response and jasmonate response. Plant Signal. Behav. 2009, 4, 464–466. [Google Scholar] [CrossRef]
- Duval, I.; Lachance, D.; Giguere, I.; Bomal, C.; Morency, M.; Pelletier, G.; Boyle, B.; MacKay, J.J.; Seguin, A. Large-scale screening of transcription factor-promoter interactions in spruce reveals a transcriptional network involved in vascular development. J. Exp. Bot. 2014, 65, 2319–2333. [Google Scholar] [CrossRef] [PubMed]
- Simpson, S.D.; Nakashima, K.; Narusaka, Y.; Seki, M.; Shinozaki, K.; Yamaguchi-Shinozaki, K. Two different novel cis-acting elements of ERD1, a ClpA homologous Arabidopsis gene function in induction by dehydration stress and dark-induced senescence. Plant J. 2003, 33, 259–270. [Google Scholar] [CrossRef]
- Nuruzzaman, M.; Sharoni, A.M.; Kikuchi, S. Roles of NAC transcription factors in the regulation of biotic and abiotic stress responses in plants. Front. Microbiol. 2013, 4, 248. [Google Scholar] [CrossRef] [PubMed]
- Bhadouriya, S.L.; Suresh, A.; Gupta, H.; Mehrotra, S.; Gupta, D.; Mehrotra, R. In silico analysis of ccgac and catgtg cis-regulatory elements across genomes reveals their roles in gene regulation under stress. Curr. Genom. 2021, 22, 353–362. [Google Scholar] [CrossRef] [PubMed]
- Ma, J.; Yuan, M.; Sun, B.; Zhang, D.; Zhang, J.; Li, C.; Shao, Y.; Liu, W.; Jiang, L. Evolutionary divergence and biased expression of NAC transcription factors in hexaploid bread wheat (Triticum aestivum L.). Plants 2021, 10, 382. [Google Scholar] [CrossRef]
- Tamura, K.; Peterson, D.; Peterson, N.; Stecher, G.; Nei, M.; Kumar, S. MEGA5: Molecular evolutionary genetics analysis using maximum likelihood, evolutionary distance, and maximum parsimony methods. Mol. Biol. Evol. 2011, 28, 2731–2739. [Google Scholar] [CrossRef]
- Gietz, D.; St Jean, A.; Woods, R.A.; Schiestl, R.H. Improved method for high efficiency transformation of intact yeast cells. Nucleic Acids Res. 1992, 20, 1425. [Google Scholar] [CrossRef]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]




Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cao, Q.; Du, J.; Yin, M.; Wang, C.; Zhang, T.; Zhao, Q.; Liu, L.; Zhang, H.; Zhang, L. Isolation and Expression Pattern Analysis of Larix olgensis LoNAC5: LoNAC5 Acts as a Positive Regulator of Drought and Salt Tolerance. Plants 2025, 14, 1527. https://doi.org/10.3390/plants14101527
Cao Q, Du J, Yin M, Wang C, Zhang T, Zhao Q, Liu L, Zhang H, Zhang L. Isolation and Expression Pattern Analysis of Larix olgensis LoNAC5: LoNAC5 Acts as a Positive Regulator of Drought and Salt Tolerance. Plants. 2025; 14(10):1527. https://doi.org/10.3390/plants14101527
Chicago/Turabian StyleCao, Qing, Junjie Du, Mengxu Yin, Chen Wang, Tiantian Zhang, Qingrong Zhao, Lu Liu, Hanguo Zhang, and Lei Zhang. 2025. "Isolation and Expression Pattern Analysis of Larix olgensis LoNAC5: LoNAC5 Acts as a Positive Regulator of Drought and Salt Tolerance" Plants 14, no. 10: 1527. https://doi.org/10.3390/plants14101527
APA StyleCao, Q., Du, J., Yin, M., Wang, C., Zhang, T., Zhao, Q., Liu, L., Zhang, H., & Zhang, L. (2025). Isolation and Expression Pattern Analysis of Larix olgensis LoNAC5: LoNAC5 Acts as a Positive Regulator of Drought and Salt Tolerance. Plants, 14(10), 1527. https://doi.org/10.3390/plants14101527