

Oxidative Stress in Rice (Oryza sativa): Mechanisms, Impact, and Adaptive Strategies



Abstract
1. Introduction
2. Oxidative Stress in Rice
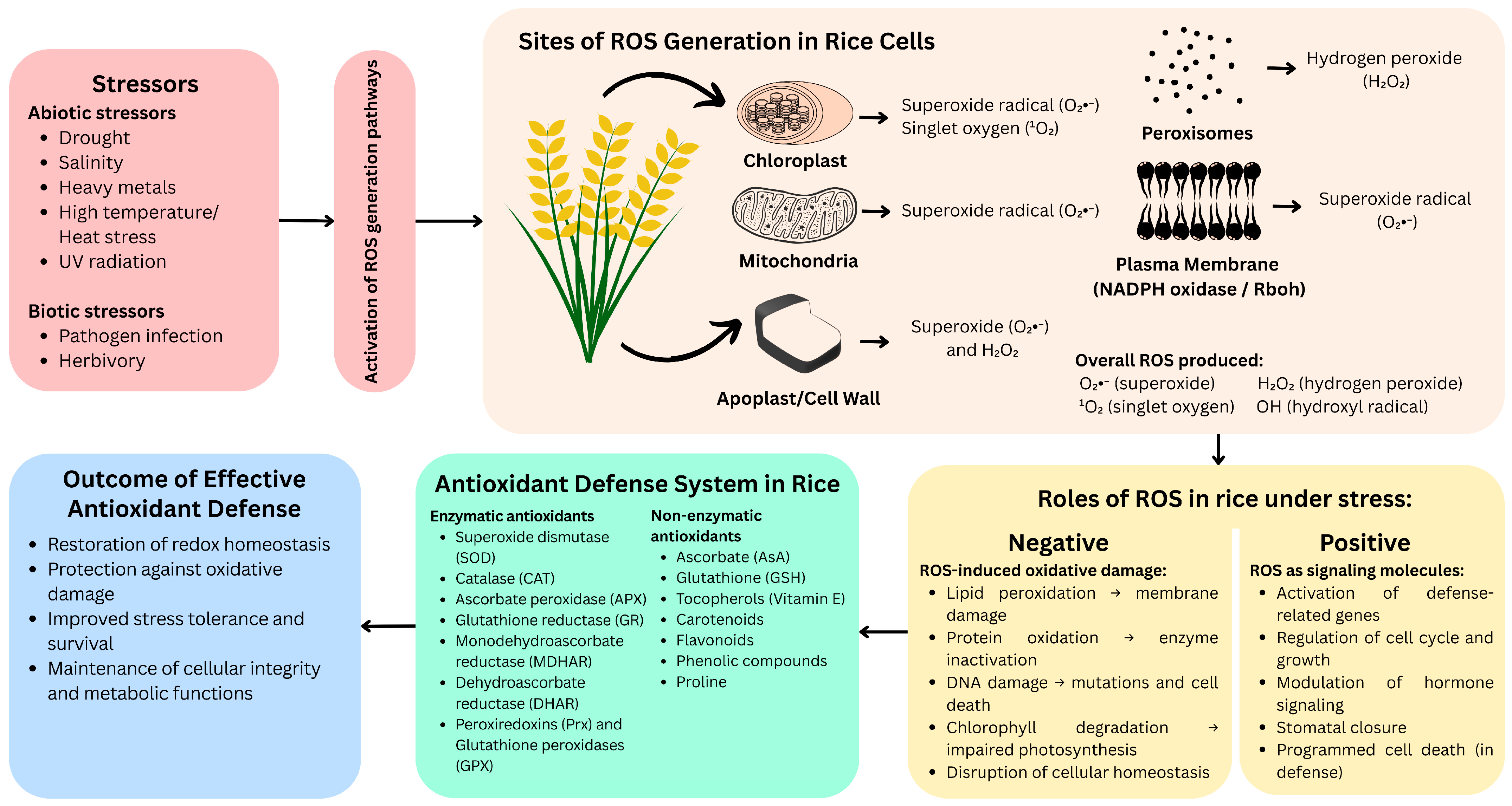
2.1. Sources and Roles of Reactive Oxygen Species (ROS)
2.2. ROS Scavenging and Antioxidant Defense Systems
2.3. Molecular and Genetic Regulation
3. Impact of Oxidative Stress on Rice Growth and Yield
3.1. Effects on Plant Physiology and Biochemistry
3.2. Impact on Grain Yield and Quality
3.3. Variations Among Different Rice Varieties
4. Adaptive Responses to Oxidative Stress in Rice
4.1. Physiological and Biochemical Adaptations
4.2. Molecular and Genetic Adaptations
4.3. Role of Epigenetics and Small RNAs
4.4. Recent Advances in Breeding and Biotechnology for Oxidative Stress Tolerance
5. Challenges and Future Directions
6. Conclusions and Recommendations
- Prioritize research on non-coding RNAs to elucidate their regulatory roles in oxidative stress response pathways, facilitating the development of novel post-transcriptional strategies to enhance stress tolerance.
- Conduct long-term field trials under diverse agro-ecological conditions to validate laboratory findings and ensure the durability and real-world applicability of stress tolerance traits across multiple growing seasons.
- Develop rice varieties with multi-stress tolerance by investigating the interactions between oxidative stress and other abiotic stressors, such as drought and salinity, to produce cultivars capable of thriving under simultaneous environmental stresses.
- Employ a systems biology approach that integrates genomics, proteomics, and metabolomics to deepen understanding of complex stress response networks and identify novel targets for breeding programs.
- Integrate oxidative stress resilience with other key agronomic traits, such as nutrient use efficiency and climate adaptability, within breeding programs aligned with climate-smart agriculture principles to enhance the sustainability of rice production systems.
- Foster interdisciplinary collaboration among researchers, plant breeders, agronomists, and policymakers to scale up the adoption of stress-tolerant rice varieties and implement effective oxidative stress mitigation strategies in support of global food security.
Author Contributions
Funding
Conflicts of Interest
References
- Demidchik, V. Mechanisms of oxidative stress in plants: From classical chemistry to cell biology. Environ. Exp. Bot. 2015, 109, 212–228. [Google Scholar] [CrossRef]
- Qamer, Z.; Chaudhary, M.T.; Du, X.; Hinze, L.; Azhar, M.T. Review of oxidative stress and antioxidative defense mechanisms in Gossypium hirsutum L. in response to extreme abiotic conditions. J. Cotton Res. 2021, 4, 9. [Google Scholar] [CrossRef]
- Panda, S.K.; Gupta, D.; Patel, M.; Vyver, C.V.D.; Koyama, H. Functionality of reactive oxygen species (ROS) in plants: Toxicity and control in Poaceae crops exposed to abiotic stress. Plants 2024, 13, 2071. [Google Scholar] [CrossRef] [PubMed]
- Qureshi, M.K. Role of Reactive Oxygen Species and Contribution of New Players in Defense Mechanism under Drought Stress in Rice. Int. J. Agric. Biol. 2018, 20, 1339–1352. [Google Scholar] [CrossRef]
- Syed, M.A.; Ahmed, M.M.E.; Debsharma, S.K.; Jahan, N.; Afrin, W.; Biswas, A.; Afrin, S.; Akter, N.; Akter, M.B. Breeding and omics approaches to understand abiotic stress response in rice. In Omics Approach to Manage Abiotic Stress in Cereals; Roychoudhury, A., Aftab, T., Acharya, K., Eds.; Springer Nature: Singapore, 2022; pp. 341–404. [Google Scholar] [CrossRef]
- Khan, M.I.R.; Kumari, S.; Nazir, F.; Khanna, R.R.; Gupta, R.; Chhillar, H. Defensive role of plant hormones in advancing abiotic stress-resistant rice plants. Rice Sci. 2023, 30, 15–35. [Google Scholar] [CrossRef]
- Dar, M.H.; Bano, D.A.; Waza, S.A.; Zaidi, N.W.; Majid, A.; Shikari, A.B.; Ahangar, M.A.; Hossain, M.; Kumar, A.; Singh, U.S. Abiotic Stress Tolerance-Progress and Pathways of Sustainable Rice Production. Sustainability 2021, 13, 2078. [Google Scholar] [CrossRef]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Mubarik, M.S.; Arif, M.S.; Riaz, M. Abiotic stress-induced oxidative stress in rice. In Advances in Rice Research for Abiotic Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2019; pp. 489–504. [Google Scholar] [CrossRef]
- Jarin, A.S.; Islam, M.M.; Rahat, A.; Ahmed, S.; Ghosh, P.; Murata, Y. Drought stress tolerance in rice: Physiological and biochemical insights. Int. J. Plant Biol. 2024, 15, 692–718. [Google Scholar] [CrossRef]
- Monsur, M.B.; Datta, J.; Rohman, M.M.; Hasanuzzaman, M.; Hossain, A.; Islam, M.S.; Bukhari, M.A.; Jabeen, T.; Mubeen, M.; Nasim, W.; et al. Saline Toxicity and Antioxidant Response in Oryza sativa: An Updated Review. In Managing Plant Production Under Changing Environment; Hasanuzzaman, M., Ahammed, G.J., Nahar, K., Eds.; Springer Nature: Singapore, 2022; pp. 79–102. [Google Scholar] [CrossRef]
- Das, A.; Pal, S.; Chakraborty, N.; Hasanuzzaman, M.; Adak, M.K. Regulation of reactive oxygen species metabolism and oxidative stress signaling by abscisic acid pretreatment in rice (Oryza sativa L.) seedlings through sub1A QTL under salinity. Plant Stress 2024, 11, 100422. [Google Scholar] [CrossRef]
- Guo, Z.; Cai, L.; Liu, C.; Chen, Z.; Guan, S.; Ma, W.; Pan, G. Low-temperature stress affects reactive oxygen species, osmotic adjustment substances, and antioxidants in rice (Oryza sativa L.) at the reproductive stage. Sci. Rep. 2022, 12, 6224. [Google Scholar] [CrossRef]
- Kaur, N.; Kirat, K.; Saini, S.; Sharma, I.; Gantet, P.; Pati, P.K. Reactive oxygen species generating system and brassinosteroids are linked to salt stress adaptation mechanisms in rice. Plant Signal. Behav. 2016, 11, e1247136. [Google Scholar] [CrossRef]
- Liu, J.; Fu, C.; Li, G.; Khan, M.N.; Wu, H. ROS Homeostasis and Plant Salt Tolerance: Plant Nanobiotechnology Updates. Sustainability 2021, 13, 3552. [Google Scholar] [CrossRef]
- Saad-Allah, K.M.; Abdelsalam, I.M. Abiotic stress triggers mitochondrial defense system: A comprehensive review. Iran. J. Plant Physiol. 2020, 10, 3195–3212. [Google Scholar]
- Wang, X.; Zhang, M.; Wang, Y.; Gao, Y.; Li, R.; Wang, G.; Li, W.; Liu, W.; Chen, K. The plasma membrane NADPH oxidase OsRbohA plays a crucial role in developmental regulation and drought-stress response in rice. Physiol. Plant. 2016, 156, 421–443. [Google Scholar] [CrossRef]
- Saeidnejad, A.H. Adjustment of Photosynthetic System Performance under Oxidative Stress. In Handbook of Photosynthesis, 4th ed.; Pessarakli, M., Ed.; CRC Press: Boca Raton, FL, USA, 2024; pp. 685–693. [Google Scholar] [CrossRef]
- Pradhan, A.K.; Jyoti, S.Y.; Shandilya, Z.M.; Rehman, M.; Saikia, D.; Poudel, J.; Kalita, J.; Borborah, K.; Chowra, U.K.; Chutia, J.; et al. Dissecting the molecular basis of drought-induced oxidative stress tolerance in rice. In Molecular Breeding for Rice Abiotic Stress Tolerance and Nutritional Quality; Hossain, M.A., Hassan, L., Md. Ifterkharuddaula, K., Kumar, A., Henry, R., Eds.; Wiley: Hoboken, NJ, USA, 2021; pp. 249–273. [Google Scholar] [CrossRef]
- Liao, M.; Ma, Z.; Kang, Y.; Zhang, B.; Gao, X.; Yu, F.; Yang, P.; Ke, Y. Enhanced disease susceptibility 1 promotes hydrogen peroxide scavenging to enhance rice thermotolerance. Plant Physiol. 2023, 192, 3106–3119. [Google Scholar] [CrossRef] [PubMed]
- Kou, Y.; Qiu, J.; Tao, Z. Every coin has two sides: Reactive oxygen species during rice–Magnaporthe oryzae interaction. Int. J. Mol. Sci. 2019, 20, 1191. [Google Scholar] [CrossRef] [PubMed]
- Hajiboland, R. Reactive Oxygen Species and Photosynthesis. In Oxidative Damage to Plants; Elsevier: Amsterdam, The Netherlands, 2014; pp. 1–63. [Google Scholar] [CrossRef]
- Bashir, F.; Jan, S. Oxidative stress and antioxidant defence systems in response to pesticide stress. In Legumes Under Environmental Stress; Azooz, M.M., Ahmad, P., Eds.; Wiley: Hoboken, NJ, USA, 2015; pp. 103–124. [Google Scholar] [CrossRef]
- Sharma, N.; Nagar, S.; Thakur, M.; Suriyakumar, P.; Kataria, S.; Shanker, A.K.; Landi, M.; Anand, A. Photosystems under high light stress: Throwing light on mechanism and adaptation. Photosynthetica 2023, 61, 250–263. [Google Scholar] [CrossRef] [PubMed]
- Poonam; Bhardwaj, R.; Kaur, R.; Bali, S.; Kaur, P.; Sirhindi, G.; Thukral, A.K.; Ohri, P.; Vig, A.P. Role of various hormones in photosynthetic responses of green plants under environmental stresses. Curr. Protein Pept. Sci. 2015, 16, 435–449. [Google Scholar] [CrossRef]
- Dalal, V.K.; Tripathy, B.C. Water-stress induced downsizing of light-harvesting antenna complex protects developing rice seedlings from photo-oxidative damage. Sci. Rep. 2018, 8, 5955. [Google Scholar] [CrossRef]
- You, J.; Zong, W.; Hu, H.; Li, X.; Xiao, J.; Xiong, L. A STRESS-RESPONSIVE NAC1-regulated protein phosphatase gene Rice Protein Phosphatase18 modulates drought and oxidative stress tolerance through abscisic acid-independent reactive oxygen species scavenging in rice. Plant Physiol. 2014, 166, 2100–2114. [Google Scholar] [CrossRef]
- Saini, D.; Bharath, P.; Gahir, S.; Raghavendra, A.S. Suppression of photorespiratory metabolism by low O2 and presence of aminooxyacetic acid induces oxidative stress in Arabidopsis thaliana leaves. Physiol. Mol. Biol. Plants 2023, 29, 1851–1861. [Google Scholar] [CrossRef]
- Nongpiur, R.C.; Rawat, N.; Singla-Pareek, S.L.; Pareek, A. OsRR26, a type-B response regulator, modulates salinity tolerance in rice via phytohormone-mediated ROS accumulation in roots and influencing reproductive development. Planta 2024, 259, 96. [Google Scholar] [CrossRef] [PubMed]
- Hisyam, B.; Amirul Ala, M.; Naimah, N.; Sarwar Jah, M. Roles of glycinebetaine on antioxidants and gene function in rice plants under water stress. Asian J. Plant Sci. 2017, 16, 132–140. [Google Scholar] [CrossRef]
- Morita, S. Engineering of Abiotic Stress Tolerance by Modulating Antioxidant Defense Systems. In Advances in Rice Research for Abiotic Stress Tolerance; Elsevier: Amsterdam, The Netherlands, 2019; pp. 755–765. [Google Scholar] [CrossRef]
- Rajput, V.D.; Harish; Singh, R.K.; Verma, K.K.; Sharma, L.; Quiroz-Figueroa, F.R.; Meena, M.; Gour, V.S.; Minkina, T.; Sushkova, S.; et al. Recent Developments in Enzymatic Antioxidant Defence Mechanism in Plants with Special Reference to Abiotic Stress. Biology 2021, 10, 267. [Google Scholar] [CrossRef] [PubMed]
- Rezayian, M.; Niknam, V.; Ebrahimzadeh, H. Oxidative damage and antioxidative system in algae. Toxicol. Rep. 2019, 6, 1309–1313. [Google Scholar] [CrossRef] [PubMed]
- Wang, X.; Liu, H.; Yu, F.; Hu, B.; Jia, Y.; Sha, H.; Zhao, H. Differential activity of the antioxidant defence system and alterations in the accumulation of osmolyte and reactive oxygen species under drought stress and recovery in rice (Oryza sativa L.) tillering. Sci. Rep. 2019, 9, 8543. [Google Scholar] [CrossRef]
- Chatterjee, A.; Dey, T.; Galiba, G.; Kocsy, G.K.; Dey, N.; Kar, R.K. Effect of combination of light and drought stress on physiology and oxidative metabolism of rice plants. Plant Sci. Today 2021, 8, 1245. [Google Scholar] [CrossRef]
- Vighi, I.L.; Benitez, L.C.; Amaral, M.N.; Moraes, G.P.; Auler, P.A.; Rodrigues, G.S.; Deuner, S.; Maia, L.C.; Braga, E.J.B. Functional characterization of the antioxidant enzymes in rice plants exposed to salinity stress. Biol. Plant. 2017, 61, 540–550. [Google Scholar] [CrossRef]
- Kumar, N.; Shankhdhar, S.C.; Shankhdhar, D. Impact of elevated temperature on antioxidant activity and membrane stability in different genotypes of rice (Oryza sativa L.). Indian J. Plant Physiol. 2016, 21, 37–43. [Google Scholar] [CrossRef]
- Rehman, M.; Kundu, B.; Regon, P.; Tanti, B. Biochemical and molecular properties of Boro rice (Oryza sativa L.) cultivars under abiotic stresses. 3 Biotech 2023, 13, 422. [Google Scholar] [CrossRef]
- Wang, L.; Zhang, X.; She, Y.; Hu, C.; Wang, Q.; Wu, L.; You, C.; Ke, J.; He, H. Physiological adaptation mechanisms to drought and rewatering in water-saving and drought-resistant rice. Int. J. Mol. Sci. 2022, 23, 14043. [Google Scholar] [CrossRef]
- Kim, J.-J.; Kim, Y.-S.; Park, S.-I.; Mok, J.-E.; Kim, Y.-H.; Park, H.-M.; Kim, I.-S.; Yoon, H.-S. Cytosolic monodehydroascorbate reductase gene affects stress adaptation and grain yield under paddy field conditions in Oryza sativa L. japonica. Mol. Breed. 2017, 37, 118. [Google Scholar] [CrossRef]
- Faseela, P.; Puthur, J.T. The imprints of the high light and UV-B stresses in Oryza sativa L. ‘Kanchana’ seedlings are differentially modulated. J. Photochem. Photobiol. B 2018, 178, 551–559. [Google Scholar] [CrossRef]
- Yemelyanov, V.V.; Prikaziuk, E.G.; Lastochkin, V.V.; Aresheva, O.M.; Chirkova, T.V. Ascorbate-glutathione cycle in wheat and rice seedlings under anoxia and subsequent reaeration. Vavilov J. Genet. Breed. 2024, 28, 44–54. [Google Scholar] [CrossRef]
- Pandey, S.; Gupta, K.; Gupta, K.; Singh, R.P.; Kumar, A.; Gaur, A.; Panigrahi, J. Role of ascorbate peroxidase in ascorbate-glutathione pathway for stress tolerance. In Annual Plant Reviews Online; Roberts, J.A., Ed.; Wiley: Hoboken, NJ, USA, 2024; Volume 1, pp. 1–48. [Google Scholar] [CrossRef]
- Park, S.-I.; Kim, Y.-S.; Kim, J.-J.; Mok, J.-E.; Kim, Y.-H.; Park, H.-M.; Kim, I.-S.; Yoon, H.-S. Improved stress tolerance and productivity in transgenic rice plants constitutively expressing the Oryza sativa glutathione synthetase OsGS under paddy field conditions. J. Plant Physiol. 2017, 215, 39–47. [Google Scholar] [CrossRef]
- Singh, A.; Banerjee, A.; Roychoudhury, A. Short-term and long-term fluoride stress induce differential molecular and transcriptional regulation and variable ranges of fluoride tolerance in two indica rice (Oryza sativa) varieties. Funct. Plant Biol. 2025, 52, FP23323. [Google Scholar] [CrossRef]
- Kaur, N.; Dhawan, M.; Sharma, I.; Pati, P.K. Interdependency of reactive oxygen species generating and scavenging system in salt sensitive and salt tolerant cultivars of rice. BMC Plant Biol. 2016, 16, 131. [Google Scholar] [CrossRef] [PubMed]
- Pan, R.; Zhang, Z.; Li, Y.; Zhu, S.; Anwar, S.; Huang, J.; Zhang, C.; Yin, L. Stage-specific effects of silver nanoparticles on physiology during the early growth stages of rice. Plants 2024, 13, 3454. [Google Scholar] [CrossRef]
- Refli; Purwestri, Y.A. The response of antioxidant genes in rice (Oryza sativa L.) seedling Cv. Cempo Ireng under drought and salinity stresses. AIP Conf. Proc. 2016, 1744, 020047. [Google Scholar] [CrossRef]
- Yuan, H.; Qian, J.; Wang, C.; Shi, W.; Chang, H.; Yin, H.; Xiao, Y.; Wang, Y.; Li, Q. Exogenous Melatonin Enhances Rice Blast Disease Resistance by Promoting Seedling Growth and Antioxidant Defense in Rice. Int. J. Mol. Sci. 2025, 26, 1171. [Google Scholar] [CrossRef]
- Karwa, S.; Arya, S.S.; Maurya, S.; Pal, M. Physiological characterization of reproductive stage heat stress tolerance in contrasting rice genotypes. Plant Physiol. Rep. 2020, 25, 157–162. [Google Scholar] [CrossRef]
- Thuesombat, P.; Hannongbua, S.; Ekgasit, S.; Chadchawan, S. Effects of silver nanoparticles on hydrogen peroxide generation and antioxidant enzyme responses in rice. J. Nanosci. Nanotechnol. 2016, 16, 8030–8043. [Google Scholar] [CrossRef]
- Wang, Y.; Deng, C.; Ai, P.; Cui, X.; Zhang, Z. ALM1, encoding a Fe-superoxide dismutase, is critical for rice chloroplast biogenesis and drought stress response. Crop J. 2021, 9, 1018–1029. [Google Scholar] [CrossRef]
- Kumar, V.; Chaudhary, P.; Prasad, A.; Dogra, V.; Kumar, A. Jasmonic acid limits Rhizoctonia solani AG1-IA infection in rice by modulating reactive oxygen species homeostasis. Plant Physiol. Biochem. 2023, 196, 520–530. [Google Scholar] [CrossRef] [PubMed]
- Ren, N.; Zhang, G.; Yang, X.; Chen, J.; Ni, L.; Jiang, M. MAPKKK28 functions upstream of the MKK1-MPK1 cascade to regulate abscisic acid responses in rice. Plant Cell Environ. 2024, 47, 5140–5157. [Google Scholar] [CrossRef]
- Huang, S.; Hu, L.; Zhang, S.; Zhang, M.; Jiang, W.; Wu, T.; Du, X. Rice OsWRKY50 mediates ABA-dependent seed germination and seedling growth, and ABA-independent salt stress tolerance. Int. J. Mol. Sci. 2021, 22, 8625. [Google Scholar] [CrossRef]
- Yan, L.; Wang, B.; Li, J.; Song, Z.; Chen, M.; Xu, Y.; Xu, B.; Yang, B.; Li, J.; Liu, J.; et al. A novel SAPK10-WRKY87-ABF1 biological pathway synergistically enhances abiotic stress tolerance in transgenic rice (Oryza sativa). Plant Physiol. Biochem. 2021, 168, 252–262. [Google Scholar] [CrossRef]
- Hong, Y.; Liu, Q.; Cao, Y.; Zhang, Y.; Chen, D.; Lou, X.; Cheng, S.; Cao, L. The OsMPK15 negatively regulates Magnaporthe oryzae and Xoo disease resistance via SA and JA signaling pathway in rice. Front. Plant Sci. 2019, 10, 752. [Google Scholar] [CrossRef]
- Bo, C.; Chen, H.; Luo, G.; Li, W.; Zhang, X.; Ma, Q.; Cheng, B.; Cai, R. Maize WRKY114 gene negatively regulates salt-stress tolerance in transgenic rice. Plant Cell Rep. 2020, 39, 135–148. [Google Scholar] [CrossRef]
- Yuan, X.; Wang, H.; Cai, J.; Bi, Y.; Li, D.; Song, F. Rice NAC transcription factor ONAC066 functions as a positive regulator of drought and oxidative stress response. BMC Plant Biol. 2019, 19, 278. [Google Scholar] [CrossRef]
- Li, J.; Zhang, M.; Yang, L.; Mao, X.; Li, J.; Li, L.; Wang, J.; Liu, H.; Zheng, H.; Li, Z.; et al. OsADR3 increases drought stress tolerance by inducing antioxidant defense mechanisms and regulating OsGPX1 in rice (Oryza sativa L.). Crop J. 2021, 9, 1003–1017. [Google Scholar] [CrossRef]
- Bao, H.; Cui, Y.; Zheng, X.; Luo, C.; Li, Y.; Chen, L. Decoding the role of OsPRX83 in enhancing osmotic stress tolerance in rice through ABA-dependent pathways and ROS scavenging. Plant Signal. Behav. 2024, 19, 2391134. [Google Scholar] [CrossRef]
- Yoo, S.J.; Kim, S.H.; Kim, M.J.; Ryu, C.M.; Kim, Y.C.; Cho, B.H.; Yang, K.Y. Involvement of the OsMKK4-OsMPK1 cascade and its downstream transcription factor OsWRKY53 in the wounding response in rice. Plant Pathol. J. 2014, 30, 168–177. [Google Scholar] [CrossRef] [PubMed]
- Xiao, X.; Tang, Z.; Li, X.; Hong, Y.; Li, B.; Xiao, W.; Gao, Z.; Lin, D.; Li, C.; Luo, L.; et al. Overexpressing OsMAPK12-1 inhibits plant growth and enhances resistance to bacterial disease in rice. Funct. Plant Biol. 2017, 44, 694. [Google Scholar] [CrossRef] [PubMed]
- Xiao, J.; Cheng, H.; Li, X.; Xiao, J.; Xu, C.; Wang, S. Rice WRKY13 regulates cross talk between abiotic and biotic stress signaling pathways by selective binding to different cis-elements. Plant Physiol. 2013, 163, 1868–1882. [Google Scholar] [CrossRef] [PubMed]
- Wang, B.; Zhong, Z.; Wang, X.; Han, X.; Yu, D.; Wang, C.; Song, W.; Zheng, X.; Chen, C.; Zhang, Y. Knockout of the OsNAC006 transcription factor causes drought and heat sensitivity in rice. Int. J. Mol. Sci. 2020, 21, 2288. [Google Scholar] [CrossRef]
- Hong, Y.; Zhang, H.; Huang, L.; Li, D.; Song, F. Overexpression of a stress-responsive NAC transcription factor gene ONAC022 improves drought and salt tolerance in rice. Front. Plant Sci. 2016, 7, 4. [Google Scholar] [CrossRef]
- Chen, Y.; Xia, P. NAC transcription factors as biological macromolecules responded to abiotic stress: A comprehensive review. Int. J. Biol. Macromol. 2025, 308, 142400. [Google Scholar] [CrossRef]
- Welner, D.H.; Deeba, F.; Lo Leggio, L.; Skriver, K. NAC transcription factors: From structure to function in stress-associated networks. In Plant Transcription Factors; Yuan, L., Zhang, D., Eds.; Elsevier: Amsterdam, The Netherlands, 2016; pp. 199–212. [Google Scholar] [CrossRef]
- Pinto, S.D.S.; Souza, A.E.D.; Oliva, M.A.; Pereira, E.G. Oxidative damage and photosynthetic impairment in tropical rice cultivars upon exposure to excess iron. Sci. Agric. 2016, 73, 217–226. [Google Scholar] [CrossRef]
- Ma, J.; Lv, C.; Xu, M.; Hao, P.; Wang, Y.; Shen, W.; Gao, Z.; Chen, G.; Lv, C. Analysis of chlorophyll a fluorescence and proteomic differences of rice leaves in response to photooxidation. Acta Physiol. Plant. 2017, 39, 46. [Google Scholar] [CrossRef]
- Liu, Y.; Gao, J.; Zhong, M.; Chen, L.; Zhang, W. Effects of phosphorus and potassium supply on photosynthetic nitrogen metabolism, nitrogen absorption, and nitrogen utilization of hydroponic rice. Agronomy 2024, 14, 1726. [Google Scholar] [CrossRef]
- Zhong, C.; Cao, X.; Hu, J.; Zhu, L.; Zhang, J.; Huang, J.; Jin, Q. Nitrogen Metabolism in Adaptation of Photosynthesis to Water Stress in Rice Grown under Different Nitrogen Levels. Front. Plant Sci. 2017, 8, 1079. [Google Scholar] [CrossRef] [PubMed]
- Salgotra, R.K.; Chauhan, B.S. Ecophysiological responses of rice (Oryza sativa L.) to drought and high temperature. Agronomy 2023, 13, 1877. [Google Scholar] [CrossRef]
- Sen, A.; Puthur, J.T. Halo and UV-B priming influences various physiological and importantly yield parameters of Oryza sativa var. Vyttila 6. N. Z. J. Crop Hortic. Sci. 2021, 49, 1–16. [Google Scholar] [CrossRef]
- Rathore, R.S.; Mishra, M.; Pareek, A.; Singla-Pareek, S.L. A glutathione-independent DJ-1/Pfp1 domain containing glyoxalase III, OsDJ-1C, functions in abiotic stress adaptation in rice. Planta 2024, 259, 81. [Google Scholar] [CrossRef]
- Sreekanth, D.; Pawar, D.V.; Kumar, R.; Ratnakumar, P.; Sondhia, S.; Singh, P.K.; Mishra, J.S.; Chander, S.; Mukkamula, N.; Kiran Kumar, B. Biochemical and physiological responses of rice as influenced by Alternanthera paronychioides and Echinochloa colona under drought stress. Plant Growth Regul. 2024, 103, 119–137. [Google Scholar] [CrossRef]
- Wang, B.; Xue, P.; Zhang, Y.; Zhan, X.; Wu, W.; Yu, P.; Chen, D.; Fu, J.; Hong, Y.; Shen, X.; et al. OsCPK12 phosphorylates OsCATA and OsCATC to regulate H₂O₂ homeostasis and improve oxidative stress tolerance in rice. Plant Commun. 2024, 5, 100780. [Google Scholar] [CrossRef] [PubMed]
- Tourky, S.M.N.; Shukry, W.M.; Hossain, M.A.; Siddiqui, M.H.; Pessarakli, M.; Elghareeb, E.M. Cobalt enhanced the drought-stress tolerance of rice (Oryza sativa L.) by mitigating the oxidative damage and enhancing yield attributes. S. Afr. J. Bot. 2023, 159, 191–207. [Google Scholar] [CrossRef]
- Zafar, S.A.; Hameed, A.; Ashraf, M.; Khan, A.S.; Qamar, Z.; Li, X.; Siddique, K.H.M. Agronomic, physiological and molecular characterisation of rice mutants revealed the key role of reactive oxygen species and catalase in high-temperature stress tolerance. Funct. Plant Biol. 2020, 47, 440. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, X.; Li, M.; Shi, C.; Jiang, M. Simulation Model for Assessing High-Temperature Stress on Rice. Agronomy 2024, 14, 900. [Google Scholar] [CrossRef]
- Song, Y.; Zheng, C.; Li, S.; Chen, J.; Jiang, M. Chitosan-magnesium oxide nanoparticles improve salinity tolerance in rice (Oryza sativa L.). ACS Appl. Mater. Interfaces 2023, 15, 20649–20660. [Google Scholar] [CrossRef]
- Kim, Y.-S.; Park, S.-I.; Kim, J.-J.; Shin, S.-Y.; Kwak, S.-S.; Lee, C.-H.; Park, H.-M.; Kim, Y.-H.; Kim, I.-S.; Yoon, H.-S. Over-expression of dehydroascorbate reductase improves salt tolerance, environmental adaptability and productivity in Oryza sativa. Antioxidants 2022, 11, 1077. [Google Scholar] [CrossRef] [PubMed]
- Kasajima, I. Difference in oxidative stress tolerance between rice cultivars estimated with chlorophyll fluorescence analysis. BMC Res. Notes 2017, 10, 168. [Google Scholar] [CrossRef] [PubMed]
- Srivastava, S.; Da Silva, R.; Sharma, P.K. Impact of hypersalinity on two salt-stressed varieties of rice (Oryza sativa): A comparative study. Russ. J. Plant Physiol. 2022, 69, 60. [Google Scholar] [CrossRef]
- Swapna, S.; Shylaraj, K.S. Screening for osmotic stress responses in rice varieties under drought condition. Rice Sci. 2017, 24, 253–263. [Google Scholar] [CrossRef]
- Nupur, J.A.; Hannan, A.; Islam, A.U.; Sagor, G.H.M.; Robin, A.H.K. Root development and anti-oxidative response of rice genotypes under polyethylene glycol induced osmotic stress. Plant Breed. Biotechnol. 2020, 8, 151–162. [Google Scholar] [CrossRef]
- Mostofa, M.G.; Saegusa, D.; Fujita, M.; Tran, L.-S.P. Hydrogen Sulfide Regulates Salt Tolerance in Rice by Maintaining Na+/K+ Balance, Mineral Homeostasis and Oxidative Metabolism Under Excessive Salt Stress. Front. Plant Sci. 2015, 6, 01055. [Google Scholar] [CrossRef]
- Hasanuzzaman, M.; Raihan, M.R.H.; Siddika, A.; Rahman, K.; Nahar, K. Supplementation with Ascophyllum nodosum extracts mitigates arsenic toxicity by modulating reactive oxygen species metabolism and reducing oxidative stress in rice. Ecotoxicol. Environ. Saf. 2023, 255, 114819. [Google Scholar] [CrossRef]
- Cao, X.; Wu, L.; Wu, M.; Zhu, C.; Jin, Q.; Zhang, J. Abscisic acid mediated proline biosynthesis and antioxidant ability in roots of two different rice genotypes under hypoxic stress. BMC Plant Biol. 2020, 20, 198. [Google Scholar] [CrossRef]
- Salsinha, Y.C.F.; Nurbaiti, S.; Sebastian, A.; Indradewa, D.; Purwestri, Y.A.; Rachmawati, D. Proline-related gene expressions contribute to physiological changes of East Nusa Tenggara (Indonesia) local rice cultivars during drought stress. Biodiversitas J. Biol. Divers. 2022, 23, 734. [Google Scholar] [CrossRef]
- Lv, B.-S.; Li, X.-W.; Ma, H.-Y.; Yang, H.-Y.; Wei, L.-X.; Lv, H.-Y.; Jiang, C.-J.; Liang, Z.-W. Different modes of proline accumulation in response to saline-alkaline stress factors in rice (Oryza sativa L.). Res. Crops 2014, 15, 14. [Google Scholar] [CrossRef]
- Binodh, A.K.; Kathiresan, P.K.; Thankappan, S.; Senthil, A. Acclimatization of non-cultivated rice landraces to early moisture stress mediated by enzymatic antioxidants and osmolyte accumulation. Biocatal. Agric. Biotechnol. 2023, 47, 102623. [Google Scholar] [CrossRef]
- Melandri, G.; AbdElgawad, H.; Floková, K.; Jamar, D.C.; Asard, H.; Beemster, G.T.S.; Ruyter-Spira, C.; Bouwmeester, H.J. Drought tolerance in selected aerobic and upland rice varieties is driven by different metabolic and antioxidative responses. Planta 2021, 254, 13. [Google Scholar] [CrossRef] [PubMed]
- Mostofa, M.G.; Hossain, M.A.; Fujita, M. Trehalose pretreatment induces salt tolerance in rice (Oryza sativa L.) seedlings: Oxidative damage and co-induction of antioxidant defense and glyoxalase systems. Protoplasma 2015, 252, 461–475. [Google Scholar] [CrossRef] [PubMed]
- Xu, N.; Chu, Y.; Chen, H.; Li, X.; Wu, Q.; Jin, L.; Wang, G.; Huang, J. Rice transcription factor OsMADS25 modulates root growth and confers salinity tolerance via the ABA-mediated regulatory pathway and ROS scavenging. PLoS Genet. 2018, 14, e1007662. [Google Scholar] [CrossRef]
- Solórzano, E.; Corpas, F.J.; González-Gordo, S.; Palma, J.M. Reactive oxygen species (ROS) metabolism and nitric oxide (NO) content in roots and shoots of rice (Oryza sativa L.) plants under arsenic-induced stress. Agronomy 2020, 10, 1014. [Google Scholar] [CrossRef]
- Shan, L.; Xu, Y.; Wu, D.; Hu, J.; Yu, T.; Dang, C.; Fang, Y.; Zhang, X.; Tian, Q.; Xue, D. Effects of salicylic acid on growth, physiology, and gene expression in rice seedlings under salt and drought stress. Plant Stress 2024, 11, 100413. [Google Scholar] [CrossRef]
- Rizwan, M.; Mostofa, M.G.; Ahmad, M.Z.; Imtiaz, M.; Mehmood, S.; Adeel, M.; Dai, Z.; Li, Z.; Aziz, O.; Zhang, Y.; et al. Nitric oxide induces rice tolerance to excessive nickel by regulating nickel uptake, reactive oxygen species detoxification and defense-related gene expression. Chemosphere 2018, 191, 23–35. [Google Scholar] [CrossRef]
- Saha, I.; Dolui, D.; Ghosh, A.; Adak, M.K. In-built antioxidation capacity of sub1A QTL in rice (Oryza sativa L.) and its modulation by exogenous application of polyamine and nitric oxide. J. Exp. Biol. Agric. Sci. 2020, 8, 210–223. [Google Scholar] [CrossRef]
- Lakra, N.; Nutan, K.K.; Das, P.; Anwar, K.; Singla-Pareek, S.L.; Pareek, A. A nuclear-localized histone-gene binding protein from rice (OsHBP1b) functions in salinity and drought stress tolerance by maintaining chlorophyll content and improving the antioxidant machinery. J. Plant Physiol. 2015, 176, 36–46. [Google Scholar] [CrossRef]
- Xu, K.; Chen, S.; Li, T.; Ma, X.; Liang, X.; Ding, X.; Liu, H.; Luo, L. OsGRAS23, a rice GRAS transcription factor gene, is involved in drought stress response through regulating expression of stress-responsive genes. BMC Plant Biol. 2015, 15, 141. [Google Scholar] [CrossRef]
- Sujitha, D.; Kumar, H.G.J.; Thapliayal, G.; Pal, G.; Vanitha, P.A.; Uttarkar, A.; Patil, M.; Reddy, B.H.R.; Niranjan, V.; Rayalcheruvu, U.; et al. Transcription factors controlling the expression of oxidative stress associated genes in rice (Oryza sativa L.). Plant Biotechnol. Rep. 2023, 17, 835–851. [Google Scholar] [CrossRef]
- Hoang, T.; Tran, T.; Nguyen, T.; Williams, B.; Wurm, P.; Bellairs, S.; Mundree, S. Improvement of salinity stress tolerance in rice: Challenges and opportunities. Agronomy 2016, 6, 54. [Google Scholar] [CrossRef]
- Dasgupta, P.; Prasad, P.; Bag, S.K.; Chaudhuri, S. Dynamicity of histone H3K27ac and H3K27me3 modifications regulate the cold-responsive gene expression in Oryza sativa L. ssp. Indica. Genomics 2022, 114, 110433. [Google Scholar] [CrossRef]
- Tseng, B.; Huang, C.; King, Y.; Wu, M.; Hsieh, C.; Hsieh, K.; Hsing, Y.; Jeng, S. Hydrogen peroxide regulates the Osa-miR156-OsSPL2/OsTIFY11b module in rice. Plant Cell Environ. 2023, 46, 2507–2522. [Google Scholar] [CrossRef]
- Cheng, X.; He, Q.; Tang, S.; Wang, H.; Zhang, X.; Lv, M.; Liu, H.; Gao, Q.; Zhou, Y.; Wang, Q.; et al. The miR172/IDS1 signaling module confers salt tolerance through maintaining ROS homeostasis in cereal crops. New Phytol. 2021, 230, 1017–1033. [Google Scholar] [CrossRef] [PubMed]
- Pandey, A.K.; Gedda, M.R.; Verma, A.K. Effect of arsenic stress on expression pattern of a rice specific miR156j at various developmental stages and their allied co-expression target networks. Front. Plant Sci. 2020, 11, 752. [Google Scholar] [CrossRef]
- Xia, H.; Huang, W.; Xiong, J.; Tao, T.; Zheng, X.; Wei, H.; Yue, Y.; Chen, L.; Luo, L. Adaptive epigenetic differentiation between upland and lowland rice ecotypes revealed by methylation-sensitive amplified polymorphism. PLoS ONE 2016, 11, e0157810. [Google Scholar] [CrossRef]
- Kumar, M.; Lee, S.-C.; Kim, J.-Y.; Kim, S.-J.; Aye, S.S.; Kim, S.-R. Over-expression of dehydrin gene, OsDhn1, improves drought and salt stress tolerance through scavenging of reactive oxygen species in rice (Oryza sativa L.). J. Plant Biol. 2014, 57, 383–393. [Google Scholar] [CrossRef]
- Monterisi, S.; Zhang, L.; Garcia-Perez, P.; Alzate Zuluaga, M.Y.; Ciriello, M.; El-Nakhel, C.; Buffagni, V.; Cardarelli, M.; Colla, G.; Rouphael, Y.; et al. Integrated multi-omic approach reveals the effect of a Graminaceae-derived biostimulant and its lighter fraction on salt-stressed lettuce plants. Sci. Rep. 2024, 14, 10710. [Google Scholar] [CrossRef]
- Lai, Y.; Ma, J.; Zhang, X.; Xuan, X.; Zhu, F.; Ding, S.; Shang, F.; Chen, Y.; Zhao, B.; Lan, C.; et al. High-quality chromosome-level genome assembly and multi-omics analysis of rosemary (Salvia rosmarinus) reveals new insights into the environmental and genome adaptation. Plant Biotechnol. J. 2024, 22, 1833–1847. [Google Scholar] [CrossRef]
- Zhang, Y.; Zhou, Y.; Zhu, W.; Liu, J.; Cheng, F. Non-coding RNAs fine-tune the balance between plant growth and abiotic stress tolerance. Front. Plant Sci. 2022, 13, 965745. [Google Scholar] [CrossRef] [PubMed]
- Bahuguna, R.N.; Tamilselvan, A.; Muthurajan, R.; Solis, C.A.; Jagadish, S.V.K. Mild preflowering drought priming improves stress defences, assimilation and sink strength in rice under severe terminal drought. Funct. Plant Biol. 2018, 45, 827. [Google Scholar] [CrossRef] [PubMed]

{kind=link}
| Stress Condition | Mechanism of ROS Production | Impact | References |
|---|---|---|---|
| Drought | Increased ROS production in chloroplasts, mitochondria, and peroxisomes due to enhanced photorespiration and impaired antioxidant defenses. | This leads to oxidative damage due to elevated ROS levels, disrupting photosynthesis and metabolic balance. | [4,18] |
| Salinity | Induces ROS production in mitochondria, contributing to oxidative stress and programmed cell death (PCD). | ROS accumulation damages cellular components, leading to growth inhibition and cell death. | [14] |
| Heat | Heat stress triggers ROS production, regulated by proteins like OsEDS1, which promote hydrogen peroxide scavenging to balance ROS homeostasis. | Excess ROS causes cellular damage, reduced yield, and heat-induced oxidative stress. | [19] |
| Pathogens | ROS generated by NADPH oxidases like OsRbohB during rice–Magnaporthe oryzae interactions as a defense response. The pathogen scavenges host-derived ROS to facilitate infection. | Host ROS combat pathogens, but excessive ROS can also lead to tissue damage and aid pathogen infection. | [20] |
| Antioxidant Defense Mechanisms | Function | References |
|---|---|---|
| Enzymatic Antioxidants | ||
| Superoxide Dismutase (SOD) | Converts superoxide radicals to hydrogen peroxide | [29,30] |
| Catalase (CAT) | Decomposes hydrogen peroxide to water and oxygen | [29] |
| Ascorbate Peroxidase (APX) | Reduces hydrogen peroxide using ascorbate | [29,30] |
| Glutathione Peroxidase (GPX) | Reduces hydrogen peroxide and organic hydroperoxides | [29,31] |
| Glutathione Reductase (GR) | Regenerates reduced glutathione | [29,31] |
| Non-Enzymatic Antioxidants | ||
| Glutathione (GSH) | Directly scavenges ROS and regenerates other antioxidants | [29,31] |
| Ascorbate (Vitamin C) | Scavenges ROS and acts as a substrate for APX | [29,31] |
| Phenolic Compounds, Flavonoids, Tocopherols | Contribute to overall antioxidant capacity | [32] |
| Regulatory Element | Key Findings | Impact on Stress Tolerance | References |
|---|---|---|---|
| MAPK Pathway (OsMPK15) | OsMPK15 knock-out mutants exhibit increased ROS production, leading to enhanced pathogen resistance | Balances ROS levels, promotes disease resistance, and maintains cellular homeostasis. | [56] |
| WRKY Transcription Factors (WRKY114) | Overexpression of WRKY114 decreases salt-stress tolerance and reduces sensitivity to ABA | Regulates stress responses, but overactivation can lead to maladaptive responses. | [57] |
| NAC Transcription Factors (ONAC066) | ONAC066 activates OsDREB2A, enhancing drought and oxidative stress tolerance | Positively regulates stress-related genes, reduces ROS accumulation, and enhances tolerance. | [58] |
| Zinc Finger Protein (OsADR3) | OsADR3 enhances drought tolerance by increasing ROS scavenging and ABA sensitivity | Improves ROS scavenging and stress tolerance, particularly under drought conditions. | [59] |
| Peroxidase Gene (OsPRX83) | OsPRX83 enhances osmotic and oxidative stress tolerance via ABA-dependent pathways | Contributes to tolerance of osmotic stress and oxidative damage. | [60] |
| General Stress-Related Genes | Transgenic approaches (SOD and APX overexpression) have improved stress tolerance in lab settings | Enhance antioxidant activity and stress resilience. | [30] |
| Physiological Process | Effects of Oxidative Stress | Impact | References |
|---|---|---|---|
| Photosynthesis | Oxidative damage to chloroplast structures, reducing chlorophyll content and energy conversion efficiency, especially under excess iron and intense light. | Compromised photosynthesis leads to reduced crop productivity. | [8,68,69] |
| Respiration | Increased ROS production during photorespiration under drought conditions, causing lipid peroxidation and protein oxidation in chloroplasts and mitochondria. | Cellular damage affects respiration efficiency and disrupts the plant’s energy balance, limiting growth, and development. | [4] |
| Nutrient Uptake | Phosphorus and potassium deficiencies impair nitrogen metabolism; oxidative damage increases by water stress, limiting the plant’s ability to absorb and utilize nutrients effectively. | Impaired nutrient uptake reduces plant growth, particularly under drought and heat stress conditions. | [70,71] |
| Stomatal Conductance | Decreases stomatal conductance and water retention, reducing turgor pressure under drought and high temperatures. | Lower stomatal conductance leads to decreased water uptake and reduced growth, particularly during prolonged drought stress. | [72] |
| Lipid Peroxidation (Marker) | Malondialdehyde accumulation signals oxidative damage in cellular membranes, reflecting lipid peroxidation and the biochemical toll on the plant tissues. | Lipid peroxidation indicates the extent of oxidative damage, affecting cell integrity and overall plant health. | [8,73] |
| Stress Factor | Impact on Yield | Impact on Quality | Mitigation |
|---|---|---|---|
| Drought | Reduced grain size and number [75] compromises the energy balance and growth of rice plants [4]. | Negatively impacts the nutrient uptake [70,71]. | The application of cobalt has shown promise [77], and the development of stress-tolerant rice varieties has been identified as a sustainable approach [72,78]. |
| High Temperature | During critical stages, particularly flowering, results in significant yield losses [79]. | Poor starch deposition and increased chalkiness during the grain-filling period lead to decreased nutritional value [76]. | Genetic improvements aimed at enhancing stress tolerance [18] and applying antioxidants to improve the effects of oxidative stress [80]. |
| Salinity | Contributes to a reduction in overall yield, due to the adverse effects on physiological processes [81]. | The quality of rice is compromised by oxidative damage to proteins and lipids, which undermines its nutritional properties [81]. | The use of CMgO nanoparticles has emerged as a promising mitigation strategy for salinity stress [80]. |
| Rice Variety | Oxidative Stress Tolerance Traits | Examples |
|---|---|---|
| Traditional Varieties | Exhibit natural resilience to oxidative stress due to evolutionary adaptations like better membrane integrity and low ion leakage, enabling them to maintain photosystem II function under stress [82]. Higher levels of unsaturated fatty acids improve membrane fluidity, especially under salinity-induced stress [83]. | Co13 [82], Korgut [83] |
| Modern Varieties | Selected for enhanced oxidative stress tolerance with improvements in antioxidant enzyme activities, membrane stability, and drought recovery [84]. | Swarnaprabha, Kattamodan [84] |
| Genetically Engineered Varieties | Modified to introduce specific stress-tolerant genes, such as SUB1, which improve antioxidant defenses (ascorbate peroxidase and peroxidase activity), thus increasing resilience to oxidative stress and maintaining higher yields under adverse conditions [85]. | Binadhan-11, BRRI dhan52 [85] |
| Adaptive Mechanism | Function in Stress Response | References |
|---|---|---|
| Osmolyte Accumulation | Accumulation of osmolytes such as proline and glycine betaine aids in osmotic adjustment, protects cellular components from ionic toxicity, and scavenges ROS. These compounds help maintain membrane integrity and balance the cellular redox environment under oxidative stress. | [87,88] |
| Proline Synthesis | Proline acts as both an osmoprotectant and an antioxidant. Proline synthesis genes (OsP5CS and OsP5CR) are upregulated, while proline catabolic genes (OsPRODH) are downregulated during oxidative stress. This regulation helps rice maintain water absorption and minimize physiological damage. | [89,90] |
| Antioxidant Enzyme Activity | Antioxidant enzymes such as superoxide dismutase (SOD), catalase (CAT), and peroxidase (POD) neutralize ROS, reducing oxidative damage to cellular structures. Increased enzyme activity correlates with higher stress tolerance. | [91,92] |
| Calcium-Dependent Protein Kinases (CPKs) | CPKs regulate antioxidant enzyme activity, improving the plant’s ability to cope with oxidative stress by modulating cellular ROS scavenging. | [76] |
| Adaptive Mechanism | Function in Oxidative Stress Response | Examples | References |
|---|---|---|---|
| Submergence and Oxidative Stress Tolerance | Primarily involved in submergence tolerance, sub1A indirectly enhances antioxidation pathways, suggesting overlapping stress responses to submergence and oxidative stress. | Sub1A provides dual tolerance to both submergence and oxidative stress in rice cultivars. | [98] |
| Salinity and Drought Tolerance | Saltol QTL and OsHBP1b transcription factor enhance salinity and drought tolerance by modulating antioxidant enzyme activities and reducing ROS levels. | Saltol QTL enhances rice’s resilience to drought and salinity, offering better stress tolerance. | [99] |
| Transcriptional Regulation | Enhances drought and oxidative stress tolerance by regulating stress-responsive genes and reducing H2O2 accumulation, minimizing oxidative damage. | OsGRAS23 transcription factor improves rice’s adaptive responses under drought and oxidative stress. | [100] |
| Antioxidant Enzyme Activity | Overexpression of OsPRX83 increases osmotic and oxidative stress tolerance by boosting ROS scavenging and antioxidant defenses. | OsPRX83 overexpression leads to increased oxidative stress resistance in genetically modified rice. | [60] |
| Regulatory Components | These regulatory components maintain ROS homeostasis and activate stress resistance mechanisms, fine-tuning the plant’s response to oxidative stress. | Regulatory networks involving kinases and transcription factors modulate ROS balance under stress. | [30] |
| Polyamine and Proline Synthesis | Polyamines act as ROS scavengers and stabilize membrane integrity, providing protection against oxidative damage. | Polyamines contribute to antioxidant activity and stress tolerance. | [98] |
| Differential Gene Expression | Transcription factors modulate the plant’s response to oxidative stress, with variations across different rice genotypes revealing genetic diversity in stress tolerance. | Diverse genetic responses in rice to oxidative stress show potential for breeding stress-tolerant varieties. | [101] |
| Adaptive Mechanism | Function in Oxidative Stress Response | Examples |
|---|---|---|
| RNA-directed DNA Methylation (RdDM) | Regulates gene expression by silencing transposable elements and maintaining genome stability under stress, crucial for development and stress adaptation [81,102]. | Silencing of transposable elements to maintain genome stability during stress [81]. |
| Histone Modifications | Facilitates chromatin remodeling, controlling access to stress-responsive genes, allowing for rapid responses to oxidative stress [103]. | Genome-wide changes in H3K27 modifications enable transcription factor access under stress [103]. |
| MicroRNAs (miRNAs) | miRNAs regulate gene expression under oxidative stress by modulating ROS scavenging and homeostasis [104,105]. | miR156 targets OsSPL2 and OsTIFY11b to manage ROS and oxidative stress [104]; miR172 scavenges ROS under salt stress [105]. |
| Developmental Stage-Specific miRNA Expression | miRNAs exhibit stage-specific expression, modulating both stress response and development, reflecting their dual role in rice’s growth phases [106]. | Osa-miR156j is differentially expressed across developmental stages, influencing both stress responses and development [106]. |
| Transgenerational Epigenetic Inheritance | Stress-induced DNA methylation changes may be inherited across generations, allowing for rice plants to pass on stress tolerance traits epigenetically [107]. | Transgenerational inheritance of stress tolerance traits via DNA methylation [107]. |
| Approach | Function in Improving Oxidative Stress Tolerance | Examples/Findings |
|---|---|---|
| Gene Overexpression | Enhances drought, salinity, and oxidative stress tolerance by improving ROS scavenging ability. | Transgenic rice overexpressing OsDhn1 shows increased stress resilience [108]. |
| Marker-Assisted Selection (MAS) | Improves stress-resilient rice varieties by incorporating genes related to submergence and salinity tolerance. | SUB1A for submergence tolerance and SalTol for salinity tolerance integrated into MAS [85,99]. |
| CRISPR/Cas9 Gene Editing | Allows for targeted gene modifications for oxidative stress tolerance; a precision tool for enhancing rice resilience to stress factors. | CRISPR/Cas9 edits stress-responsive genes for improved tolerance [108]. |
| Transcriptomics | Reveals expression profiles of genes involved in salinity and oxidative stress tolerance, aiding genomic breeding efforts. | Identified Rbohs and brassinosteroid biosynthesis genes are important for stress tolerance [45]. |
| Proteomics and Metabolomics | Identifies key enzymes and antioxidants involved in oxidative stress management, providing biochemical markers for breeding. | NADP-malic enzyme, glutathione, and ascorbate are critical for oxidative stress control [11]. |
| Abscisic Acid (ABA) Treatment | Enhances stress tolerance by activating antioxidant enzyme activities and maintaining cellular homeostasis under stress conditions. | ABA treatments boost antioxidant enzyme activity for improved stress resilience [11]. |
| Multi-Omics Integration | Provides comprehensive insights into stress-related genes, proteins, and metabolic pathways for a holistic understanding of stress tolerance. | Master regulators (e.g., transcription factors) identified for genetic manipulation [101]. |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2025 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tavu, L.E.J.; Redillas, M.C.F.R. Oxidative Stress in Rice (Oryza sativa): Mechanisms, Impact, and Adaptive Strategies. Plants 2025, 14, 1463. https://doi.org/10.3390/plants14101463
Tavu LEJ, Redillas MCFR. Oxidative Stress in Rice (Oryza sativa): Mechanisms, Impact, and Adaptive Strategies. Plants. 2025; 14(10):1463. https://doi.org/10.3390/plants14101463
Chicago/Turabian StyleTavu, Lady Edlenill J., and Mark Christian Felipe R. Redillas. 2025. "Oxidative Stress in Rice (Oryza sativa): Mechanisms, Impact, and Adaptive Strategies" Plants 14, no. 10: 1463. https://doi.org/10.3390/plants14101463
APA StyleTavu, L. E. J., & Redillas, M. C. F. R. (2025). Oxidative Stress in Rice (Oryza sativa): Mechanisms, Impact, and Adaptive Strategies. Plants, 14(10), 1463. https://doi.org/10.3390/plants14101463