Abstract
To examine the effects of the recent Acmella radicans invasion on plant community and diversity in invaded habitats, the composition, density, species richness, diversity indices, and evenness index of the soil seed bank community of two different habitats (wasteland and cultivated land) in Yunnan Province, China, were analyzed through field sampling and greenhouse germination tests. A total of 28 species of plants belonging to 15 families and 28 genera, all annual herbs, were found in the soil seed bank. Seed densities and species number in the seed bank tended to be greater in April than in October; cultivated land also featured higher seed densities and species numbers compared to wasteland. With increased A. radicans cover, the seed bank population of A. radicans also significantly increased, but the seed bank populations of many other dominant species (e.g., Ageratum conyzoides and Gamochaeta pensylvanica) and native species (e.g., Laggera crispata and Poa annua) clearly declined. The germination of A. radicans seeds was concentrated during the period from the 4th to the 5th weeks. Vertically, the seed number of A. radicans was significantly different among the 0–5 cm, 5–10 cm and 10–20 cm layers that accounted for 80.7–90.6%, 9.4–16.1% and 0.0–3.2% of the total seed density in wasteland, respectively; and in cultivated land, A. radicans accounted for 56.8–64.9%, 26.7–31.8% and 8.1–13.5% of the total seed density, respectively. With reduced A. radicans cover, the species richness, Simpson index, Shannon–Wiener index, and Pielou indices of the weed community generally increased, and most diversity indices of weed communities in cultivated land were lower than in wasteland under the same cover of A. radicans. The results indicate that the invasion of A. radicans has negatively affected local weed community composition and reduced weed community diversity, and that these negative impacts in cultivated land may be enhanced by human disturbance. Our study was the first to elucidate the influence of A. radicans invasion on soil seed bank community characteristics in invaded habitats, providing a better understanding of its invasion and spread mechanisms in order to aid in developing a scientific basis for the prevention and control of this invader.
1. Introduction
The soil seed bank is usually considered as the natural reserve of all viable seeds in a soil profile and on the soil surface [1]. Many seeds may remain viable though dormant in the soil for years. According to persistence time in the soil, soil seed banks are categorized as transient seed banks (persist for less than 1 year), short persistent seed banks (persist for 1–5 years) and long-term persistent seed banks (persist for more than 5 years) [2]. As a potential source of genetic diversity for plant populations, soil seed banks are vital for vegetation succession, vegetation maintenance, ecosystem restoration, species diversity conservation, and population management strategies [3,4,5]. Therefore, it is of great value to study the species composition, and temporal and spatial dynamics, of soil seed banks to develop comprehensive weed management, sustainable agriculture, and conservation strategies.
Biological invasions have been become one of the main factors leading to biodiversity loss and environment change on a global scale [6,7,8]. The presence of seed banks has been regarded as one of the key components of successful biological invasion for many species [9]. The invasiveness of many exotic plant species is frequently associated with high proliferation and the production of seeds that may persist in the soil for long periods of time [10]. The successful establishment of invasive plants is determined in many cases by the production of a persistent soil seed bank in newly invaded areas, given the role of seed banks as sources of propagules, genetic diversity, and in spreading the risk of germination failure over time [9]. Therefore, studies on the soil seed bank of an invasive plant species can provide valuable information on the invasiveness of a species for planning restoration measures and for developing comprehensive prevention and control measures. This is especially important for invasive alien plant species that are new to a geographic area for which information on seed bank dynamics may be lacking.
Acmella radicans (Jacquin) R.K. Jansen is an annual erect herb in the Asteraceae family that originated in central America and Mexico [11]. This plant has become broadly distributed in Colombia, Bangladesh, Cuba, Curacao, India, Tanzania, and Thailand [11,12,13]. A naturalized population was first recorded in China in 2014 in Anhui Province [14]. In 2017, A. radicans was first found in Changning County, Baoshan City of Yunnan Province in southwestern China during a survey of invasive alien plant species. Currently, A. radicans has become a serious invasive species and has invaded farmland, tea gardens, orchard land, roadsides, and ditches in Baoshan City, Lincang City and Puer City of Yunnan [15]. Acmella radicans has become a dominant species in invaded regions, negatively affecting species richness, species diversity, and the evenness and soil nutrients of local communities. This plant can release allelochemicals such as (E, E)-2,4-decadienal, 2-tridecanone, and caryophyllene oxide to inhibit the seed germination and seedling growth of some major associated weeds and local vegetable crops such as Ageratum conyzoides, Bidens pilosa, Brassica napus, Chloris virgata, and Digitaria sanguinalis [15,16]. Furthermore, A. radicans demonstrates considerable ecological adaptability, occupies a wide range of habitat types, possesses a long flowering and fruiting period (November–March), and exhibits a prolific reproductive capacity (a single plant can produce up to 14,300 seeds) [15]. However, little is known about the soil seed bank in communities invaded by A. radicans.
In the current study, the composition, density, species richness, diversity indices, and evenness index of soil seed bank communities of A. radicans in two different habitats (wasteland and cultivated land) in Yunnan Province, China, were examined through field sampling and greenhouse tests. Seed bank characterization is important to better elucidate its invasion and spread mechanisms and provide a scientific basis for the forecasting, risk analysis, prevention, and control of A. radicans.
2. Results
2.1. Plant Species and Seed Density
A total of 28 plant species were identified within the study plots, belonging to 28 genera and 15 families (Table 1). Families with the most plant species were Asteraceae (9 species) and Poaceae (6 species), accounting for 32.1% and 21.4% of all species, respectively, and there was only one species for each of the other 13 families. All plants were herbaceous, among which 20 annual plants accounted for 71.4%, 7 perennial plants accounted for 25% and 1 annual/perennial plant constituted 4.6% of all species. In terms of invasiveness, there were 12 invasive alien plant species and 16 native species, accounting for 42.9% and 57.1% of all species, respectively (Table 1).

Table 1.
Basic information of plant species, life form and invasiveness of plant species found in the soil seed bank in study plots located in cultivated and wasteland habitats in Mengtong Township, Changning County.
In terms of the frequency and density of species in the habitat, 12 plant species—Acmella radicans, Ageratum conyzoides, Bidens pilosa, Chloris virgata, Digitaria sanguinalis, Eleusine indica, Gamochaeta pensylvanica, Galinsoga quadriradiata, Oplismenus undulatifolius, Poa annua, Polypogon fugax, and Spermacoce alata—exhibited high population density and dominance (Table 2 and Table 3). As Acmella radicans cover increased, the seed density of Acmella radicans significantly increased; however, seed densities of many other dominant invasive plants, i.e., Ageratum conyzoides, B. pilosa, E. indica, Gamochaeta pensylvanica, Galinsoga quadriradiata, and native plants C. virgata, D. sanguinalis, Laggera crispata, Oxalis corniculata, and P. annua clearly declined. Compared with the total density of different habitats, the seed densities of invasive alien plants, native plants, and A. radicans accounted for 85.7–95.7%, 4.3–14.3% and 10.5–62.4% of the total seed density, respectively (Figure 1). With cover increases in A. radicans, the seed density proportion of invasive plants and A. radicans in both habitats observed was significantly increased, with the proportion significantly higher in cultivated land than in wasteland under the same A. radicans cover (Table 4).
Table 2.
Average seed densities of the soil seed bank within different habitats in April 2022 (seedlings/m2).
Table 3.
Average seed densities of the soil seed bank within different habitats in October 2022 (seedlings/m2).
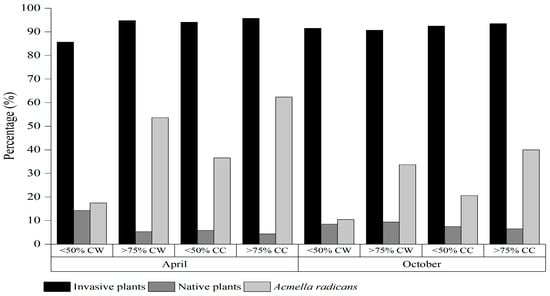
Figure 1.
Percentage of invasive plants, native plants and Acmella radicans of the total density of soil weed communities within different habitats (CW = cover of wasteland and CC = cover of cultivated land).
Table 4.
Kruskal–Wallis test results for the seed density of invasive plants, native plants and Acmella radicans in the soil seed bank within different habitats.
2.2. Soil Seed Germination and Distribution of Acmella radicans
The germination tests showed that the germination process of A. radicans took place over 9 weeks, and that there was no difference in germination characteristics between wasteland and cultivated land. The seeds of A. radicans began to germinate after 2 weeks, with germination concentrated during the period from the 4th to the 5th weeks and ceasing at the 9th week.
The seed density and distribution of A. radicans were markedly different in different soil depths and habitats (Table 5). Seed densities of A. radicans showed obvious increases with increases in cover of A. radicans in both habitats, and the seed density of A. radicans in cultivated land was significantly higher than in wasteland under the same A. radicans cover. Vertically, the seed number of A. radicans was significantly different in shallower than deeper soil layers. The 0–5 cm, 5–10 cm and 10–20 cm layers accounted for 80.7–90.6%, 9.4–16.1% and 0.0–3.2% of the total seed density in wasteland, respectively; and in cultivated land, A. radicans comprised 56.8–64.9%, 26.7–31.8% and 8.1–13.5% of the total seed density, respectively (Figure 2). It is clear that A. radicans has persistent soil seed banks that play an important role during its invasion and expansion.
Table 5.
Kruskal–Wallis test results for the seed density of Acmella radicans among different soil layers within different habitats.
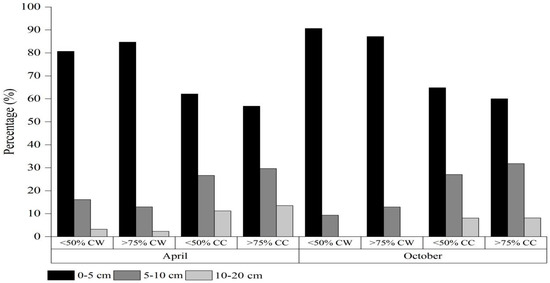
Figure 2.
Percentage of soil seed distribution of Acmella radicans among different soil layers within different habitats (CW = cover of wasteland and CC = cover of cultivated land).
2.3. Effects of Acmella radicans on Soil Plant Species Diversity
Overall, the species richness (S), Simpson index (D), Shannon–Wiener index (H), and Pielou index (J) of weed communities were generally greater with less A. radicans cover, and most diversity indices of weed communities in cultivated land were lower than in wasteland under the same cover of A. radicans (Table 6). The results indicate that the invasion of A. radicans has negatively affected local weed community composition and reduced weed community diversity, and that these negative impacts in cultivated land may be enhanced by human disturbance.
Table 6.
Kruskal–Wallis test results for diversity indices of weed communities within different habitats.
3. Discussion
Increasing numbers of studies have demonstrated that invasive alien plants can reduce species richness, species diversity indices, and evenness indices through strong ecological adaptability, competitiveness and allelopathic inhibition, altering underground and aboveground composition and the structure of plant communities in invaded habitats [17,18,19,20]. As a new invasive species in Yunnan province, A. radicans is already widely distributed in Baoshan, Lincang and Puer cities, causing serious damage to local agricultural production and ecological environments [15]. However, the effect of A. radicans invasion on soil seed bank characteristics was previously unclear. Our current study found that A. radicans has modified the species composition, population density and weed community diversity of the soil seed bank in the invaded area, and human disturbance in cultivated land has aggravated these effects.
The soil seed bank is generally considered to be an important potential seed source for the invasion and spread of invasive alien plants and plays a critical role in vegetation succession and species diversity maintenance [17,18]. Characterizing the seed bank of invasive alien plants informs an understanding of the type, magnitude and rate of change in the seed bank and predicts the recruitment potential of these species from the seed bank [21]. The similarity between soil seed banks and the standing vegetation of invasive plants is usually high at the initial invasion stage and gradually declines over time [17,18]. Our current study found that the species richness of the soil seed bank is clearly higher than that of the aboveground vegetation, while the seed bank population is markedly smaller than the seed rain from the aboveground vegetation in A. radicans invaded habitats. A major factor contributing to this disparity is that a large proportion of the seed rain was transported to other places by wind and other disturbances. A second major factor is related to the invasion history and the storage characteristics of other plant seeds. As the invasion duration and resulting damage increases, invasive alien plants will gradually come to occupy a dominant position in both the local vegetation and the soil seed bank, while reducing the population density of other plants in the community, especially native species [22,23,24,25]. The current study found that as the seed density of A. radicans significantly increased, seed densities of many other dominant invasive plants and native plants clearly declined. The A. radicans seed density corresponded to increases in A. radicans cover, which was partly due to the strong competitiveness and allelopathic inhibition of A. radicans [15,16]. Moreover, with increased cover of A. radicans, the seed density of other invasive plants as well as A. radicans in study plots was markedly increased. The proportion of invasive species in the seed bank was significantly higher in cultivated land than in wasteland under the same A. radicans cover, indicating that A. radicans had higher negative impacts on native species than the other invasive alien species present, and that the weed community in cultivated land was affected by both the stress of A. radicans invasion and human disturbance. Local plant species and community structure are more vulnerable to disturbance than those of invasive alien plants under the same conditions because invasive species usually have greater morphological plasticity and competitiveness [26]. Synergistic impacts may result when adventive invasive alien plant species and disturbances coincide that are conducive to both the invasion of the adventive species and expansion of other invasive species [27].
Invasive alien plants not only change species composition and population density of weed communities in invaded habitats but also affect the species diversity. Zheng et al. [28] reported that the negative correlation between the importance value of Spermacoce latifolia and the species diversity of the community indicated that the invasion and spreading of S. latifolia may have a negative impact on the species diversity of the community. Nan et al. [18] found that the importance value of Oenothera laciniata in the community had a positive correlation with the species diversity of the community but this was not significant. Robertson and Hickman [29] found that as Bothriochloa spp. invasion increases, the native plant community decreased in diversity and abundance, but no effect on native seed bank diversity and density was observed. Our previous study found that A. radicans had negative impacts on species richness, species diversity, and evenness and soil nutrients of local communities [15]. Furthermore, we found that the species richness, Simpson index, Shannon–Wiener index, and Pielou index of the weed community usually increased as the A. radicans cover declined, and most diversity indices of weed communities in cultivated land were lower than in wasteland under the same cover of A. radicans, showing that the invasion of A. radicans has a negative impact on the diversity of local weed communities and that human disturbance in cultivated land may aggravate this trend. In our study area, there were few human activities (never grazed and/or cultivated) since 2002 in the wasteland habitat, indicating that the decline in species diversity of the weed community was directly caused by A. radicans invasion. The more pronounced decline in weed community species diversity in cultivated land was caused by the combination of A. radicans invasion and human disturbance. Other studies have reported that the invasion of exotic plant species and native species suppression were increased by numerous disturbances [30,31].
Invasive alien plants adapt to different environmental conditions and changes with various reproductive strategies during their life history [32]. Soil seed banks may be the most tolerant to a wide range of environmental conditions by comparison to other life history elements because many seeds may be dormant or remain viable in the soil over the long term [2,33]. The soil seed bank is heterogeneously distributed in soil wherein the topsoil layer is usually transient seed and has the highest proportion of the seed bank, while the proportion of the seed bank gradually decreases and becomes more of a persistent seed bank with increases in soil depth [34,35,36]. Acmella radicans is an annual herb, with a five-month flowering and fruiting period, producing up to 14,300 seeds per plant [15]. Its high fecundity is critical to the growth and spread of its populations. The present study showed that A. radicans seeds in the 0–5 cm, 5–10 cm and 10–20 cm layers of cultivated land accounted for 56.8–64.9%, 26.7–31.8% and 8.1–13.5% of the total seed density, respectively. In wasteland, most A. radicans seeds were stored in the 0–5 cm layer, accounting for 80.7–90.6% of the seed bank. The seed densities of A. radicans substantially increased with increases in the cover of A. radicans in both habitats, and the seed density of A. radicans in cultivated land was significantly higher than in wasteland under the same A. radicans cover. It is clear that A. radicans forms a large proportion of its persistent soil seed bank in deep soil in cultivated land, potentially contributing to future recruitment, especially if facilitated by human disturbance. Abundant transient and persistent seed banks of A. radicans are conducive to maintaining relatively high populations or even undergoing large outbreaks under the right conditions. Moreover, the persistent seed bank ensures A. radicans is not threatened with extirpation under adverse environment and human disturbance.
Considering the plant growth, reproduction, and soil seed bank characteristics of A. radicans under natural conditions, it is critical to adopt a long-term and comprehensive plan for its prevention, control and management. As an annual herb, A. radicans completes its life history and sexual reproduction within a single year. Thus, in order to reduce or avoid the seed dispersal of A. radicans, the optimum time for prevention and control is before flowering or at least prior to seed maturation with efficient herbicides or elimination by physical control techniques. In addition, for managing the transient and/or persistent soil seed bank A. radicans, a reduction in human activities or the use of soil herbicides to limit the seed germination after crop planting could be effective. Finally, prevention and control measures such as no-tillage mulching, plastic film mulching, intercropping, and replacement techniques could be combined, which would serve not only to inhibit the number of A. radicans effectively but also produce excellent ecological and economic benefits. Therefore, comprehensive measures must be considered for the prevention and control of A. radicans, incorporating chemical herbicides supplemented by physical control, ecological restoration, and replacement control. It is possible to control or reduce the extent of invasion by A. radicans, but this would require several years of continuous prevention and control measures.
4. Materials and Methods
4.1. Study Site
The study site was located in Changning County (20°14′ N–25°12′ N, 99°16′ E–100°12′ E), Baoshan City, in the west boundary area of Yunnan Province, southwest China. This area has a subtropical low-latitude mountain plateau monsoon climate, which is characterized by warm winter, cool summer, abundant rainfall, and wet and dry seasons. The annual average temperature of Changning is 14.9 °C, and the annual rainfall is 1259 mm. Recently, the range of Acmella radicans has been expanding rapidly within Changning County, as the plant has invaded cultivated land, orchard land, wastelands, riverbanks, and ditches [15].
4.2. Soil Collection
Two invaded habitats (wasteland and cultivated land) where A. radicans was growing were chosen in Mengtong Township, Changning County. The selected habitats were 5.5 km apart from each other and had similar soil types and ecological conditions but different levels of human disturbance. The wasteland is abandoned without any human activities, and the cultivated land is usually plowed with grown maize over one season and then it is fallowed for six months every year in these years. Plots 4 m × 4 m in size with less than 50 (35–50)% and more than 75 (75–100)% plant cover of A. radicans in each site were separately selected, and a total of 16 plots were randomly established and permanently marked.
The soil samples for analyzing the seed bank were collected in April and October 2022. Acmella radicans plants have completed fruiting, and most of them have fallen to the ground in April, and the natural germination of seed banks of this plant has finished before October. Five sample plots of 10 cm × 10 cm for each plot were randomly chosen at three soil layers (0–5 cm, 5–10 cm and 10–20 cm depths) for the soil collection. The soil samples of five plots at each layer were mixed to provide a composite sample. All the samples were properly marked and then transported to the laboratory. Litter and organic matter (including stones, coarse organic fragments, roots, and rhizomes) were removed from the soil samples; then, the samples were ground and sifted through a 4 mm sieve, which was followed by air-drying at room temperature for further germination tests. The soils from the two locations were similar and identified as yellow earths in two invaded habitats. The soils nutrient levels were measured as follows: pH 4.63–4.67, organic matter 30.96–31.41 g/kg, total N 1.49–1.53 g/kg, total K 1.04–1.21 g/kg, and total P 13.57–13.99 g/kg.
4.3. Germination Tests
Germination tests were conducted in the greenhouse to estimate the seed bank composition and density from 5 June to 13 October 2023. The soil was evenly distributed on flat plates (50 cm × 30 cm × 8 cm), and the soil sample depth was approximately 1–2 cm. The soils were regularly watered to avoiding drying out. After five weeks, the number of seedlings per plate was recorded, and seedlings were identified to species and removed each week. Those plants that could not be identified were transplanted and grown for later species identification. The germination test was terminated at the point when we no longer observed the emergence of new seedlings.
4.4. Statistical Analysis
Based on plant species identification and the seedling counting of soil samples, the species composition, seed density, seed proportion, and weed community diversity were analyzed. We combined the data from the three germination layers in order to estimate the seed density of each species in each soil collected times. Soil seed bank density (seedlings/m2) was calculated from the number of emerged seedlings in each plot with four replicates. The distribution percentage of invasive plants, native plants, and A. radicans was determined. Similarly, the soil seed bank density and distribution percentage A. radicans at each depth (0–5 cm, 5–10 m, 5–10 cm) and that of the whole profile (0–20 cm) were analyzed separately. Species richness, diversity and evenness were estimated as follows: (1) the Simpson diversity index (D) was calculated as D = 1 − ∑Ni (Ni − 1)/N (N − 1), where Ni is the total number of individuals from species i in a plot and N is the total number of individuals from all species in a plot; D ranges from 0 to 1, with 1 being the maximal diversity; (2) the Shannon–Wiener diversity index H [37] was measured as H = −∑Pi lnPi, where Pi is the proportion relative to the total number of species per plot; and (3) the Pielou evenness index (J) [38] was calculated as J = H/lnS, where S is the species richness of each plot. We tested the normality of the soil seed bank data and found the data were not normally distributed. Therefore, the nonparametric Kruskal–Wallis test was used for the identification of significant differences among groups at a 5% level using IBM SPSS22.0 software (Armonk, NY, USA).
5. Conclusions
Our results indicated that the invasion of A. radicans has changed the species composition, population density and weed community diversity of soil seed bank in the invaded area, and that human disturbance in cultivated land aggravated this trend. As A. radicans cover increased, the seed density of A. radicans significantly increased, but the seed densities of many other dominant invasive plants and native plants clearly declined. The soil seed germination of A. radicans occurred over 9 weeks and was concentrated during the period from the 4th to the 5th weeks. Seeds of A. radicans in the 0–5 cm, 5–10 cm and 10–20 cm soil layers of cultivated land accounted for 56.8–64.9%, 26.7–31.8% and 8.1–13.5% of the total seed density, respectively, but in wasteland, most seeds were stored in the 0–5 cm layer with A. radicans seeds accounting for 80.7–90.6% of the seed bank. The species richness, Simpson index, Shannon–Wiener index, and Pielou index of the weed community were generally increased with declining A. radicans cover, and most diversity indices of weed communities in cultivated land were lower than in wasteland under the same cover of A. radicans. The relationship between the soil seed bank and aboveground vegetation, influence of different environmental factors on seed germination, and effect of disturbance on the sexual reproduction characteristics of A. radicans require further research to help develop effective management strategies.
Author Contributions
S.S. and F.Z. (Fudou Zhang) conceived and designed the experiments; X.W., K.Y., F.Z. (Fengping Zheng), G.X., Z.F., Y.Y., S.Y. and G.J. performed the experiments; X.W. analyzed the data; S.S. and D.R.C. wrote the draft. All authors have read and agreed to the published version of the manuscript.
Funding
This research was supported by the National Key R&D Program of China (2021YFC2600400), Key Research and Development Program of Yunnan Province (202103AF140007 and 202203AE140008), and Ten Thousand Talent Program (Young Top-Notch Talent) of Yunnan Province (YNWR-QNBJ-2018-201).
Data Availability Statement
Data are contained within the article.
Acknowledgments
We wish to thank Randi Wu from the Agriculture and Life Sciences College of Kunming University for his great experimental support.
Conflicts of Interest
The authors declare no conflicts of interest. The founding sponsors had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, and in the decision to publish the results.
References
- Yu, S.; Jiang, G. The research development of soil seed bank and several hot topics. Acta Phytoecol. Sin. 2003, 27, 552–560. [Google Scholar] [CrossRef]
- Yu, S.; Chen, H.; Lang, N. The classification systems of soil seed banks and seed persistence in soil. Acta Ecol. Sin. 2007, 27, 2099–2108. [Google Scholar]
- Liu, R.; Zhan, J.; Shi, Z.; Chen, L. Soil seed bank and its correlations with aboveground vegetation and environmental factors in water level fluctuating zone of Danjiangkou Reservoir, Central China. Chin. J. Appl. Ecol. 2013, 24, 801–808. [Google Scholar]
- Chu, H.; Zhang, C.; Dong, Q.; Shang, Z.; Degen, A.; Yang, X.; Yu, Y.; Yang, Z.; Zhang, Y. The effect of grazing intensity and season on the soil seed bank and its relation with above-ground vegetation on the alpine steppe. Agr. Ecosyst. Environ. 2019, 285, 106622. [Google Scholar] [CrossRef]
- Hou, Z.; Xie, Y.; Yu, X.; Li, F. Characteristics of soil seed banks in different water level areas after returning farmland into lake in Qingshanyuan of Dongting Lake. Chin. J. Appl. Ecol. 2009, 20, 1323–1328. [Google Scholar]
- Simberloff, D.; Martin, J.L.; Genovesi, P.; Maris, V.; Wardle, D.A.; Aronson, J.; Courchamp, F.; Galil, B.; García-Berthou, E.; Pascal, M.; et al. Impacts of biological invasions: What’s what and the way forward. Trends Ecol. Evol. 2013, 28, 58–66. [Google Scholar] [CrossRef]
- Blackburn, T.M.; Essl, F.; Evans, T.; Hulme, P.E.; Jeschke, J.M.; Kühn, I.; Kumschick, S.; Marková, Z.; Mrugała, A.; Nentwig, W.; et al. A unified classification of alien species based on the magnitude of their environmental impacts. PLoS Biol. 2014, 12, e1001850. [Google Scholar] [CrossRef]
- Pyšek, P.; Jarošík, V.; Hulme, P.E.; Pergl, J.; Hejda, M.; Schaffner, U.; Vilà, M. A global assessment of invasive plant impacts on resident species, communities and ecosystems: The interaction of impact measures, invading species’ traits and environment. Glob. Chang. Biol. 2012, 18, 1725–1737. [Google Scholar] [CrossRef]
- Gioria, M.; Le Roux, J.J.; Hirsch, H.; Moravcová, L.; Pyšek, P. Characteristics of the soil seed bank of invasive and non-invasive plants in their native and alien distribution range. Biol. Invasions 2019, 21, 2313–2332. [Google Scholar] [CrossRef]
- Marques, A.R.; Costa, C.F.; Atman, A.P.F.; Garcia, Q.S. Germination characteristics and seedbank of the alien species Leucaena leucocephala (Fabaceae) in Brazilian forest: Ecological implications. Weed Res. 2014, 54, 576–583. [Google Scholar] [CrossRef]
- Bagga, J.; Deshmukh, U.B. Acmella radicans (Jacquin) R.K. Jansen (Asteraceae)—A new distributional plant record for Jharkhand State (India). J. New Biol. Rep. 2018, 7, 24–27. [Google Scholar]
- Panyadee, P.; Inta, A. Taxonomy and ethnobotany of Acmella (Asteraceae) in Thailand. Biodiversitas 2022, 23, 2177–2186. [Google Scholar] [CrossRef]
- Rahman, M.M.; Khan, S.A.; Hossain, G.M.; Jakaria, M.; Rahim, M.A. Acmella radicans (Jacq.) R.K. Jansen (Asteraceae)—A new angiosperm record. J. Biol. Sci. 2016, 5, 87–93. [Google Scholar] [CrossRef]
- Wang, Z.; Yan, X.; Li, H.; Ma, J. Acmella radicans var. debilis (Kunth) R.K. Jansen (Asteraceae), a newly naturalized plant in China. J. Trop. Subtrop. Bot. 2015, 23, 643–646. [Google Scholar] [CrossRef]
- Yang, K.; Wu, X.; Zheng, F.; Fan, Z.; Wu, R.; Xu, G.; Yang, Y.; Zhang, F.; Yang, S.; Shen, S. Allelopathic effects of aqueous extracts of the invasive plant Acmella radicans on seed germination and seedling growth of four weeds. Acta Agrestia Sin. 2023, 31, 3757–3765. [Google Scholar] [CrossRef]
- Yang, K.; Yang, Y.; Wu, X.; Zheng, F.; Xu, G.; Yang, S.; Jin, G.; Clements, D.R.; Shen, S.; Zhang, F. Allelopathic potential and chemical composition of essential oil from the invasive plant Acmella radicans. Agronomy 2024, 14, 342. [Google Scholar] [CrossRef]
- Shen, S.; Xu, G.; Zhang, F.; Li, T.; Jin, G.; Zhang, Y. Characteristics of the seed banks and seedling banks of Mikania micrantha-invaded soils different in type of habitat. J. Eco. Rural Environ. 2013, 29, 483–488. [Google Scholar]
- Nan, K.; Wu, Q.; Hu, R.; Chen, S.; Ding, B. Seasonal dynamics of species composition of community and soil seed bank invaded by Oenothera laciniata Hill in Wenzhou, Zhejiang. J. Trop. Subtrop. Bot. 2009, 17, 535–542. [Google Scholar]
- Wang, S.; Gao, X.; Wang, J.; Dang, W. Characteristics of soil seed banks of crofton weed and their effects on seedlings. Chin. J. Plant Ecol. 2009, 33, 380–386. [Google Scholar] [CrossRef]
- Li, M.; Li, L.; Hao, G.; Kang, B.; Gao, Y.; Li, H. Effects of Amaranthus palmeri on soil seed bank community characteristics in abandoned cultivated land. J. Ecol. Rural Environ. 2023, 39, 1188–1195. [Google Scholar] [CrossRef]
- Gioria, M.; Pyšek, P. The legacy of plant invasions: Changes in the soil seed bank of invaded plant communities. BioScience 2016, 66, 40–53. [Google Scholar] [CrossRef]
- Wang, R.; Dong, H.; Liu, T.; Zhao, W.; Wang, H.; Ma, Q.; Liu, Y. Characteristics of soil seed banks and their contribution to aboveground population of invasive weeds Ambrosia artemisiifolia and A. trifid. J. Shihezi Univ. 2021, 39, 72–79. [Google Scholar] [CrossRef]
- Dang, W.; Gao, X.; Wang, J.; Li, A. Soil seed bank traits in an area invaded by Eupatorium adenophorum. Biodivers. Sci. 2008, 16, 126–132. [Google Scholar] [CrossRef]
- Shen, Y.; Liu, W. Persistent soil seed bank of Eupatorium adenophorum. Chin. J. Plant Ecol. 2004, 28, 768–772. [Google Scholar] [CrossRef]
- Salomé-Díaz, J.; Golubov, J.; Eguiarte, L.E.; Búrquez, A. Difference in germination traits between congeneric native and exotic species may affect invasion. Plants 2024, 13, 478. [Google Scholar] [CrossRef]
- Geng, Y.; Zhang, W.; Li, B.; Chen, J. Phenotypic plasticity and invasiveness of alien plants. Biodivers. Sci. 2004, 12, 447–455. [Google Scholar] [CrossRef]
- Zenni, R.D.; da Cunha, W.L.; Musso, C.; de Souza, J.V.; Nardoto, G.B.; Miranda, H.S. Synergistic impacts of co-occurring invasive grasses cause persistent effects in the soil-plant system after selective removal. Funct. Ecol. 2020, 34, 1102–1112. [Google Scholar] [CrossRef]
- Zheng, S.; Dai, L.; Lin, P.; Chen, X.; Ding, B. Species composition of community invaded by Spermacoce latifolia Aubl. and seasonal dynamics of its soil seed bank. J. Zhejiang Univ. (Agric. Life Sci.) 2009, 35, 677–685. [Google Scholar] [CrossRef]
- Robertson, S.G.; Hickman, K.R. Aboveground plant community and seed bank composition along an invasion gradient. Plant Ecol. 2012, 213, 1461–1475. [Google Scholar] [CrossRef]
- Shen, S.; Xu, G.; Clements, D.R.; Jin, G.; Liu, S.; Zhang, F.; Yang, Y.; Chen, A.; Kato-Noguchi, H. Effects of invasive plant Mikania micrantha on plant community and diversity in farming systems. Asian J. Plant Sci. 2015, 14, 27–33. [Google Scholar] [CrossRef]
- Lembrechtsa, J.J.; Pauchard, A.; Lenoird, J.; Nuñeze, M.A.; Geronf, C.; Vena, A.; Bravo-Monasteriob, P.; Tenebg, E.; Nijsa, I.; Ann Milbau, A. Disturbance is the key to plant invasions in cold environments. Proc. Natl. Acad. Sci. USA 2016, 113, 14061–14066. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Lu, B.; Lu, X.; Huang, S. On reproductive strategies of invasive plants and their impacts on native plants. Biodivers. Sci. 2018, 26, 457–467. [Google Scholar] [CrossRef]
- Angert, A.L.; Huxman, T.E.; Chesson, P.; Venable, D.L. Functional tradeoffs determine species coexistence via the storage effect. Proc. Natl. Acad. Sci. USA 2009, 106, 11641–11645. [Google Scholar] [CrossRef] [PubMed]
- Qian, J.; Liu, Z.; Hatier, J.H.B.; Liu, B. The vertical distribution of soil seed bank and its restoration implication in an active sand dune of northeastern Inner Mongolia, China. Land Degrad. Dev. 2016, 27, 305–315. [Google Scholar] [CrossRef]
- Feledyn-Szewczyk, B.; Smagacz, J.; Kwiatkowski, C.A.; Harasim, E.; Woźniak, A. Weed flora and soil seed bank composition as affected by tillage system in three-year crop rotation. Agriculture 2020, 10, 186. [Google Scholar] [CrossRef]
- Skuodienė, R.; Matyžiūtė, V.; Aleinikovienė, J.; Frercks, B.; Repšienė, R. Seed bank community under different-intensity agrophytocenoses on Hilly Terrain in Lithuania. Plants 2023, 12, 1084. [Google Scholar] [CrossRef]
- Simpson, E.H. Measurement of diversity. Nature 1949, 163, 688. [Google Scholar] [CrossRef]
- Ma, K.; Liu, Y. Measurement of biotic community diversity I α diversity (Part 2). Biodivers. Sci. 1994, 2, 231–239. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).