

Aster × chusanensis Growth and Phenolic Acid Composition under Different Cultivation Temperatures



Abstract
1. Introduction
2. Results and Discussion
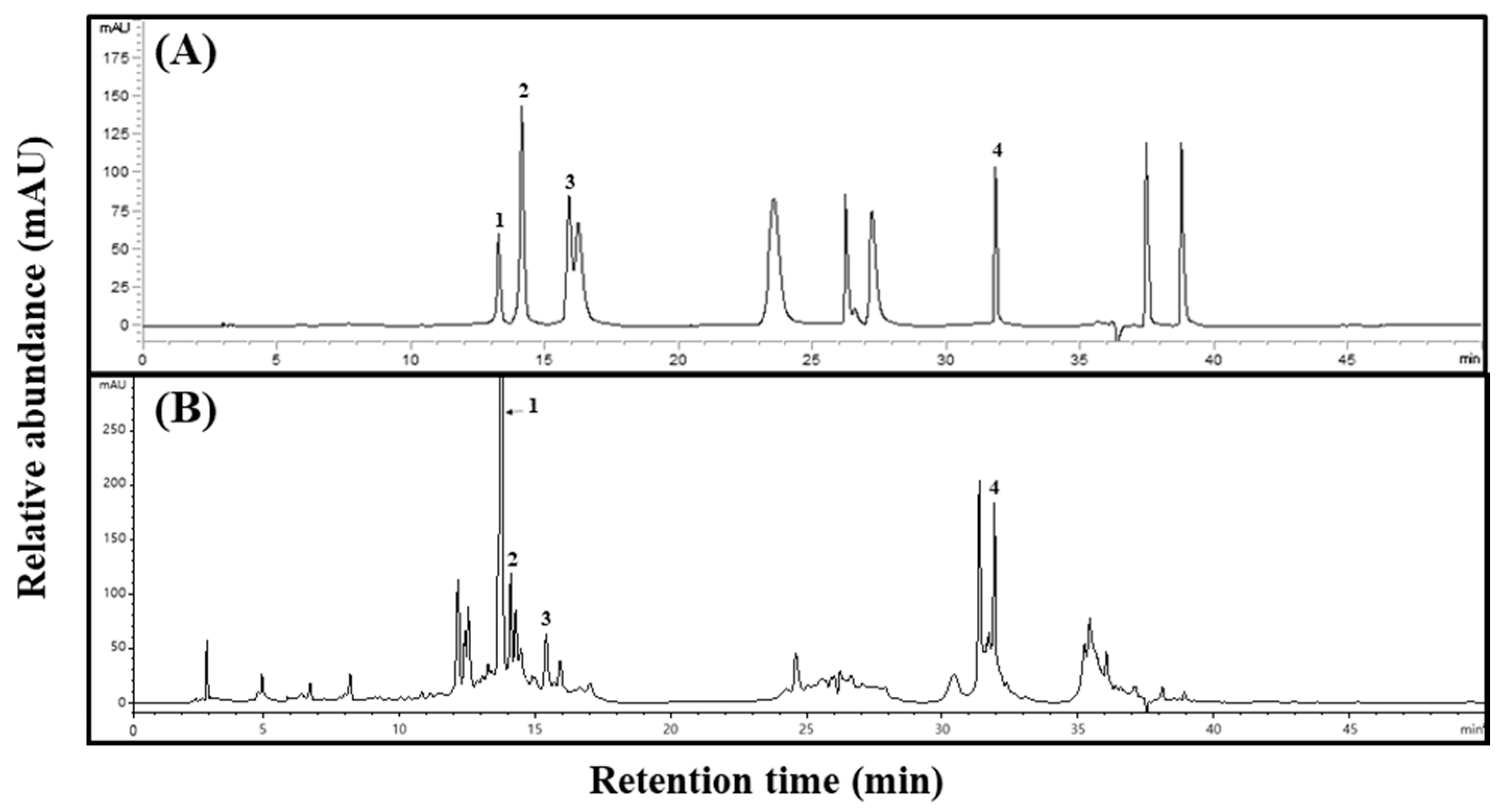
2.1. Phenolic Acid Concentrations of Aster × chusanensis Parts
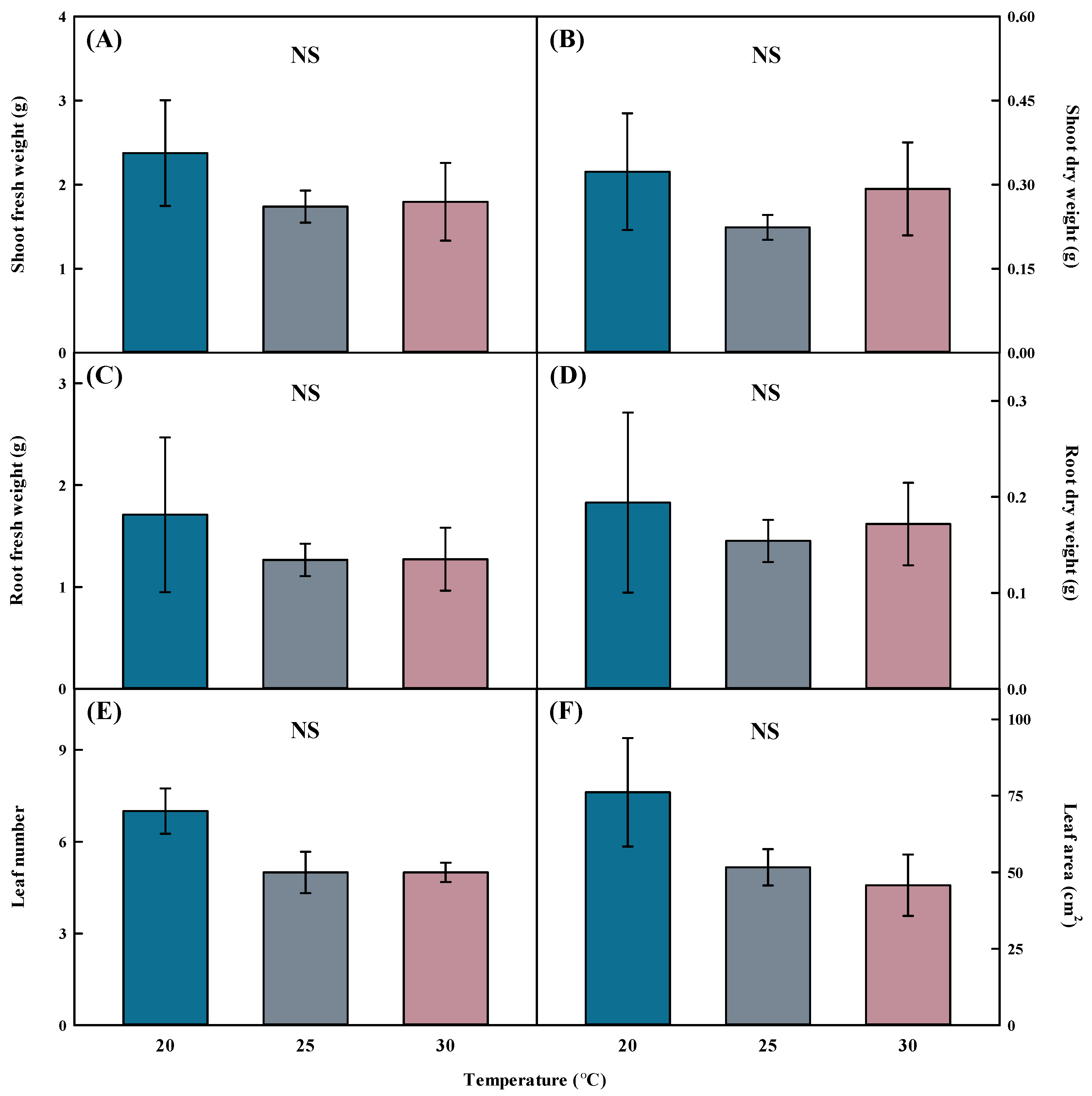
2.2. Growth Characteristic Analysis of Aster × chusanensis under Different Growth Temperatures
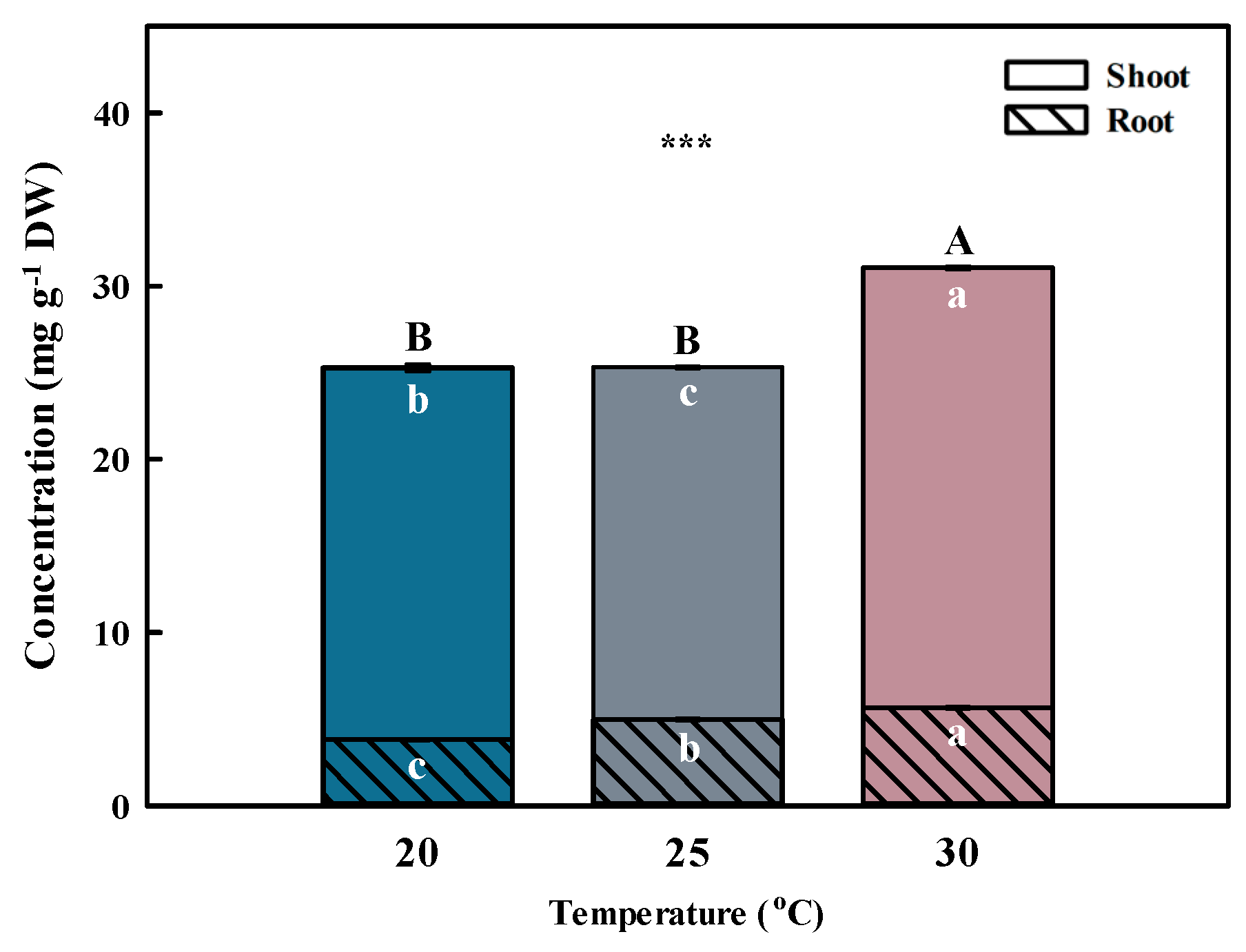
2.3. Phenolic Acid Concentrations in Aster × chusanensis Shoots under Different Growth Temperatures
2.4. Phenolic Acid Concentrations in Aster × chusanensis Roots under Different Growth Temperatures
3. Materials and Methods
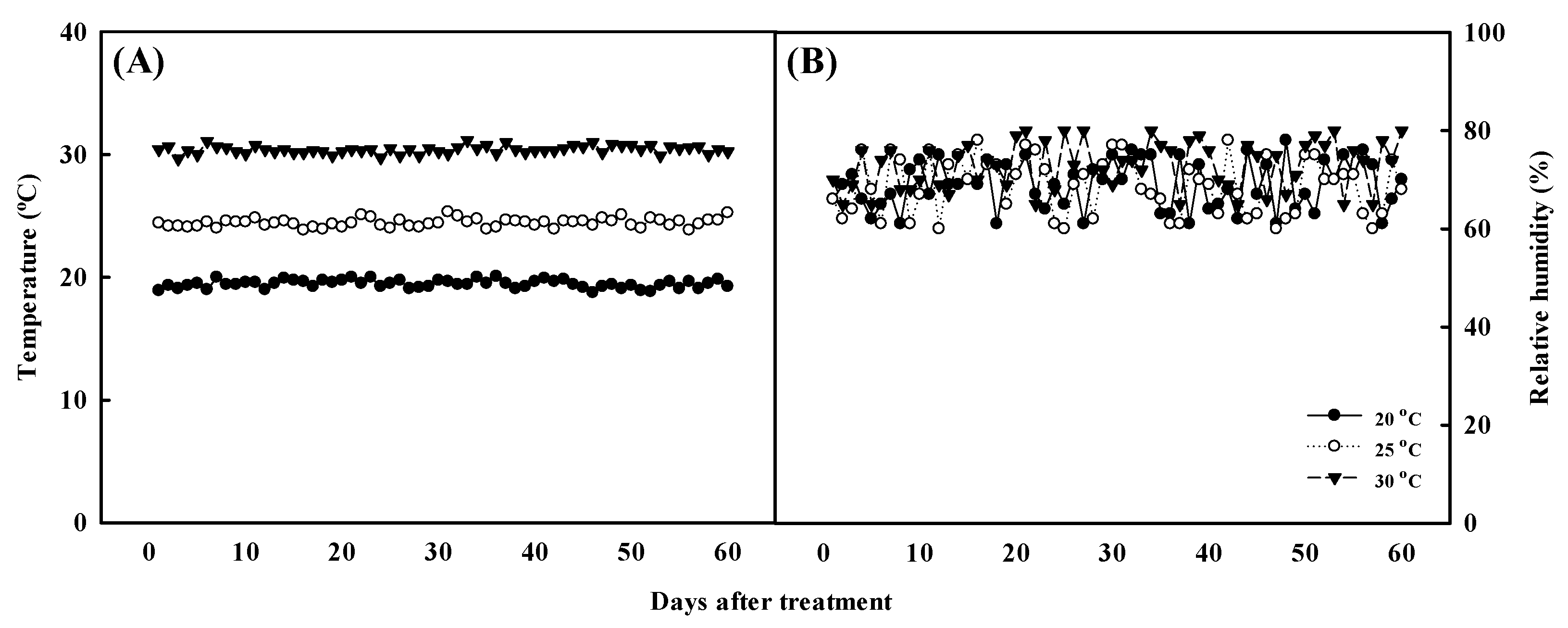
3.1. Plant Material and Growth Temperature Condition
3.2. Growth Characteristic Determination
3.3. Phenolic Acid Analysis
3.4. Chemicals
3.5. Statistical Analysis
4. Conclusions
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Adedeji, O.; Jewoola, O.A. Importance of leaf epidermal characters in the Asteraceae family. Not. Bot. Horti Agrobot. Cluj-Napoca 2008, 36, 7. [Google Scholar]
- Gao, T.; Yao, H.; Song, J.; Zhu, Y.; Liu, C.; Chen, S. Evaluating the feasibility of using candidate DNA barcodes in discriminating species of the large Asteraceae family. BMC Evol. Biol. 2010, 10, 324. [Google Scholar] [CrossRef] [PubMed]
- Shin, Y.H.; Kim, H.J.; Ku, J.J.; Park, K.W.; Choi, K.; Jeong, H.S.; Kang, S.H. The folk plants in northern region of Chungcheongbuk-do. Korean J. Plant Resour. 2012, 25, 707–718. [Google Scholar] [CrossRef]
- Son, M.J.; Jin, C.; Lee, Y.S.; Lee, J.Y.; Kim, H.J. Characterization of caffeoylglucoside derivatives and hypouricemic activity of the ethyl acetate fraction from Aster glehni. Bull. Korean Chem. Soc. 2015, 36, 503–512. [Google Scholar] [CrossRef]
- KIM, T.J. Wild Flowers of Korea; Kugilmedia: Seoul, Republic of Korea, 1996; p. 232. [Google Scholar]
- Lim, Y.; Hyun, J.O.; Kim, Y.D.; Shin, H. Aster chusanensis (Asteraceae), a new species from Korea. J. Plant Biol. 2005, 48, 479–482. [Google Scholar] [CrossRef]
- Shin, H.; Oh, S.H.; Lim, Y.; Hyun, C.W.; Cho, S.H.; Kim, Y.I.; Kim, Y.D. Molecular evidence for hybrid origin of Aster chusanensis, an endemic species of Ulleungdo, Korea. J. Plant Biol. 2014, 57, 174–185. [Google Scholar] [CrossRef]
- De Leonardis, A.; Macciola, V.; Di Rocco, A. Oxidative stabilization of cold-pressed sunflower oil using phenolic compounds of the same seeds. J. Sci. Food Agric. 2003, 83, 523–528. [Google Scholar] [CrossRef]
- Spiridon, I.; Nechita, C.B.; Niculaua, M.; Silion, M.; Armatu, A.; Teacă, C.A.; Bodîrlău, R. Antioxidant and chemical properties of Inula helenium root extracts. Cent. Eur. J. Chem. 2013, 11, 1699–1709. [Google Scholar] [CrossRef]
- Balasundram, N.; Sundram, K.; Samman, S. Phenolic compounds in plants and agri-industrial by-products: Antioxidant activity, occurrence, and potential uses. Food Chem. 2006, 99, 191–203. [Google Scholar] [CrossRef]
- Dos Santos, M.D.; Almeida, M.C.; Lopes, N.P.; De Souza, G.E.P. Evaluation of the anti-inflammatory, analgesic and antipyretic activities of the natural polyphenol chlorogenic acid. Biol. Pharm. Bull. 2006, 29, 2236–2240. [Google Scholar] [CrossRef]
- Mamani-Matsuda, M.; Kauss, T.; Al-Kharrat, A.; Rambert, J.; Fawaz, F.; Thiolat, D.; Moynet, D.; Coves, S.; Malvy, D.; Mossalayi, M.D. Therapeutic and preventive properties of quercetin in experimental arthritis correlate with decreased macrophage inflammatory mediators. Biochem. Pharmacol. 2006, 72, 1304–1310. [Google Scholar] [CrossRef] [PubMed]
- Kim, H.H.; Park, G.H.; Park, K.S.; Lee, J.Y.; An, B.J. Anti-oxidant and Anti-inflammation Activity of Fractions from Aster glehni Fr. Schm. Microbiol. Biotechnol. Lett. 2010, 38, 434–441. [Google Scholar]
- Nugroho, A.; Kim, M.H.; Choi, J.; Choi, J.S.; Jung, W.T.; Lee, K.T.; Park, H.J. Phytochemical studies of the phenolic substances in Aster glehni extract and its sedative and anticonvulsant activity. Arch. Pharmacal Res. 2012, 35, 423–430. [Google Scholar] [CrossRef] [PubMed]
- Kang, D.H.; Han, E.H.; Jin, C.; Kim, H.J. Inhibitory Effects of Ethanolic Extracts from Aster glehni on Xanthine Oxidase and Concentration Determination of Bioactive Components Using HPLC-UV. J. Korean Soc. Food Sci. Nutr. 2016, 45, 1610–1616. [Google Scholar] [CrossRef]
- He, J.; Yao, L.; Pecoraro, L.; Liu, C.; Wang, J.; Huang, L.; Gao, W. Cold stress regulates accumulation of flavonoids and terpenoids in plants by phytohormone, transcription process, functional enzyme, and epigenetics. Crit. Rev. Biotechnol. 2023, 43, 680–697. [Google Scholar] [CrossRef] [PubMed]
- Hassan, M.U.; Chattha, M.U.; Khan, I.; Chattha, M.B.; Barbanti, L.; Aamer, M.; Iqbal, M.M.; Nawaz, M.; Mahmood, A.; Ali, A.; et al. Heat stress in cultivated plants: Nature, impact, mechanisms, and mitigation strategies—A review. Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2021, 155, 211–234. [Google Scholar] [CrossRef]
- Razzak, M.A.; Asaduzzaman, M.; Tanaka, H.; Asao, T. Effects of supplementing green light to red and blue light on the growth and yield of lettuce in plant factories. Sci. Hortic. 2022, 305, 111429. [Google Scholar] [CrossRef]
- Bessada, S.M.; Barreira, J.C.; Oliveira, M.B.P. Asteraceae species with most prominent bioactivity and their potential applications: A review. Ind. Crops Prod. 2015, 76, 604–615. [Google Scholar] [CrossRef]
- Gai, F.; Karamać, M.; Janiak, M.A.; Amarowicz, R.; Peiretti, P.G. Sunflower (Helianthus annuus L.) plants at various growth stages subjected to extraction—Comparison of the antioxidant activity and phenolic profile. Antioxidants 2020, 9, 535. [Google Scholar] [CrossRef] [PubMed]
- Naveed, M.; Hejazi, V.; Abbas, M.; Kamboh, A.A.; Khan, G.J.; Shumzaid, M.; Ahmad, F.; Babazadeh, D.; FangFang, X.; Modarresi-Ghazani, F.; et al. Chlorogenic acid (CGA): A pharmacological review and call for further research. Biomed. Pharmacother. 2018, 97, 67–74. [Google Scholar] [CrossRef]
- Manuja, R.; Sachdeva, S.; Jain, A.; Chaudhary, J. A comprehensive review on biological activities of p-hydroxy benzoic acid and its derivatives. Int. J. Pharm. Sci. Rev. Res. 2013, 22, 109–115. [Google Scholar]
- Sharma, N.; Tiwari, N.; Vyas, M.; Khurana, N.; Muthuraman, A.; Utreja, P. An overview of therapeutic effects of vanillic acid. Plant Arch. 2020, 20, 3053–3059. [Google Scholar]
- Megersa, H.G. Propagation methods of selected horticultural crops by specialized organs: Review. J. Hortic. 2017, 4, 198. [Google Scholar] [CrossRef]
- Im, H.J.; Yang, J.C.; Lee, D.J.; Na, C.S.; Che, S.H. Influence of Cytokinins on Callus and Shoot Induction of Aster × chusanensis Y.; Lim, JO Hyun, YD Kim & H. Shin. Flower Res. J. 2023, 31, 260–266. [Google Scholar]
- Ampofo, J.; Ngadi, M.; Ramaswamy, H.S. The impact of temperature treatments on elicitation of the phenylpropanoid pathway, phenolic accumulations and antioxidative capacities of common bean (Phaseolus vulgaris) sprouts. Food Bioprocess Technol. 2020, 13, 1544–1555. [Google Scholar] [CrossRef]
- Pazouki, L.; Kanagendran, A.; Li, S.; Kännaste, A.; Memari, H.R.; Bichele, R.; Niinemets, Ü. Mono-and sesquiterpene release from tomato (Solanum lycopersicum) leaves upon mild and severe heat stress and through recovery: From gene expression to emission responses. Environ. Exp. Bot. 2016, 132, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Zhu, L.; Bloomfield, K.J.; Hocart, C.H.; Egerton, J.J.; O’Sullivan, O.S.; Penillard, A.; Weerasinghe, L.K.; Atkin, O.K. Plasticity of photosynthetic heat tolerance in plants adapted to thermally contrasting biomes. Plant Cell Environ. 2018, 41, 1251–1262. [Google Scholar] [CrossRef] [PubMed]
- Martinez-Medina, A.; Flors, V.; Heil, M.; Mauch-Mani, B.; Pieterse, C.M.; Pozo, M.J.; Ton, J.; van Dam, N.M.; Conrath, U. Recognizing plant defense priming. Trends Plant Sci. 2016, 21, 818–822. [Google Scholar] [CrossRef]
- Beckers, G.J.; Conrath, U. Priming for stress resistance: From the lab to the field. Curr. Opin. Plant Biol. 2007, 10, 425–431. [Google Scholar] [CrossRef]
- Theocharis, A.; Bordiec, S.; Fernandez, O.; Paquis, S.; Dhondt-Cordelier, S.; Baillieul, F.; Clément, C.; Barka, E.A. Burkholderia phytofirmans PsJN primes Vitis vinifera L. and confers a better tolerance to low nonfreezing temperatures. Mol. Plant Microbe Interact. 2012, 25, 241–249. [Google Scholar] [CrossRef]
- Allakhverdiev, S.I.; Kreslavski, V.D.; Klimov, V.V.; Los, D.A.; Carpentier, R.; Mohanty, P. Heat stress: An overview of molecular responses in photosynthesis. Photosynth. Res. 2008, 98, 541–550. [Google Scholar] [CrossRef] [PubMed]
- Ashraf, M.A.; Iqbal, M.; Rasheed, R.; Hussain, I.; Riaz, M.; Arif, M.S. Environmental stress and secondary metabolites in plants: An overview. In Plant Metabolites and Regulation under Environmental Stress; Academic Press: Cambridge, MA, USA, 2018; pp. 153–167. [Google Scholar]
- Luo, H.B.; Ma, L.; Xi, H.F.; Duan, W.; Li, S.H.; Loescher, W.; Wang, J.F.; Wang, L.J. Photosynthetic responses to heat treatments at different temperatures and following recovery in grapevine (Vitis amurensis L.) leaves. PLoS ONE 2011, 6, e23033. [Google Scholar] [CrossRef] [PubMed]
- Lee, H.Y.; Cho, D.Y.; Kim, D.H.; Park, J.H.; Jeong, J.B.; Jeon, S.H.; Lee, J.H.; Ko, E.J.; Cho, K.M. Examining the Alterations in Metabolite Constituents and Antioxidant Properties in Mountain-Cultivated Ginseng (Panax ginseng CA Meyer) Organs during a Two-Month Maturation Period. Antioxidants 2024, 13, 612. [Google Scholar] [CrossRef] [PubMed]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Phenolic Acid | Leaf | Stem | Root | Sig |
|---|---|---|---|---|
| Chlorgenic acid | 28,215.82 ± 3711.68 z a y | 1593.63 ± 21.20 b | 1164.48 ± 25.75 b | ** |
| p-hydroxybenzoic acid | 498.64 ± 1.33 a | 155.10 ± 0.58 b | 147.83 ± 0.79 c | *** |
| Vanilic acid | 354.12 ± 5.94 a | ND x | 207.26 ± 31.12 b | ** |
| Benzoic acid | 5231.75 ± 271.42 a | 560.58 ± 0.67 c | 1601.37 ± 60.51 b | *** |
| Total | 34,300.31 ± 3435.43 a | 2309.29 ± 19.96 b | 3120.91 ± 118.15 b | ** |
| Time (min) | Mobile Phase A (%) | Mobile Phase B (%) |
|---|---|---|
| 0 | 100 | 0 |
| 3 | 97 | 3 |
| 5 | 95 | 5 |
| 8 | 90 | 10 |
| 10 | 85 | 15 |
| 15 | 95 | 5 |
| 17 | 92 | 8 |
| 19 | 90 | 10 |
| 20 | 85 | 15 |
| 22 | 80 | 20 |
| 26 | 85 | 15 |
| 27 | 80 | 20 |
| 28 | 70 | 30 |
| 32 | 60 | 40 |
| 36 | 40 | 60 |
| 37 | 70 | 30 |
| 38 | 60 | 40 |
| 40 | 50 | 50 |
| 45 | 40 | 60 |
| 55 | 20 | 80 |
| 60 | 10 | 90 |
| 65 | 0 | 100 |
| Phenolic Acid | LOD (µg mL) | LOQ (µg mL) |
|---|---|---|
| /S) | /S) | |
| Chlorogenic acid | 33.594 | 101.801 |
| p-hydroxybenzoic acid | 42.326 | 128.261 |
| Vanillic acid | 41.386 | 125.413 |
| Benzoic acid | 3.773 | 11.433 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Sim, H.-S.; Kwon, H.J.; Jang, S.-N.; Lee, G.O.; Kang, I.-J.; Yang, G.-S.; Nam, G.-H.; Park, J.E.; Byun, H.Y.; You, Y.-H.; et al. Aster × chusanensis Growth and Phenolic Acid Composition under Different Cultivation Temperatures. Plants 2024, 13, 1855. https://doi.org/10.3390/plants13131855
Sim H-S, Kwon HJ, Jang S-N, Lee GO, Kang I-J, Yang G-S, Nam G-H, Park JE, Byun HY, You Y-H, et al. Aster × chusanensis Growth and Phenolic Acid Composition under Different Cultivation Temperatures. Plants. 2024; 13(13):1855. https://doi.org/10.3390/plants13131855
Chicago/Turabian StyleSim, Han-Sol, Hyuk Joon Kwon, Seong-Nam Jang, Ga Oun Lee, In-Je Kang, Gyu-Sik Yang, Gi-Heum Nam, Ji Eun Park, Ha Yeon Byun, Young-Hyun You, and et al. 2024. "Aster × chusanensis Growth and Phenolic Acid Composition under Different Cultivation Temperatures" Plants 13, no. 13: 1855. https://doi.org/10.3390/plants13131855
APA StyleSim, H.-S., Kwon, H. J., Jang, S.-N., Lee, G. O., Kang, I.-J., Yang, G.-S., Nam, G.-H., Park, J. E., Byun, H. Y., You, Y.-H., & Son, K.-H. (2024). Aster × chusanensis Growth and Phenolic Acid Composition under Different Cultivation Temperatures. Plants, 13(13), 1855. https://doi.org/10.3390/plants13131855