

Integration of Horse Manure Vermicompost Doses and Arbuscular Mycorrhizal Fungi to Improve Fruit Quality, and Soil Fertility in Tomato Field Facing Drought Stress

 ,
,  ,
,  and
and
Abstract
1. Introduction
2. Results
2.1. Yield, Fruit Growth, and Root Colonization Changes
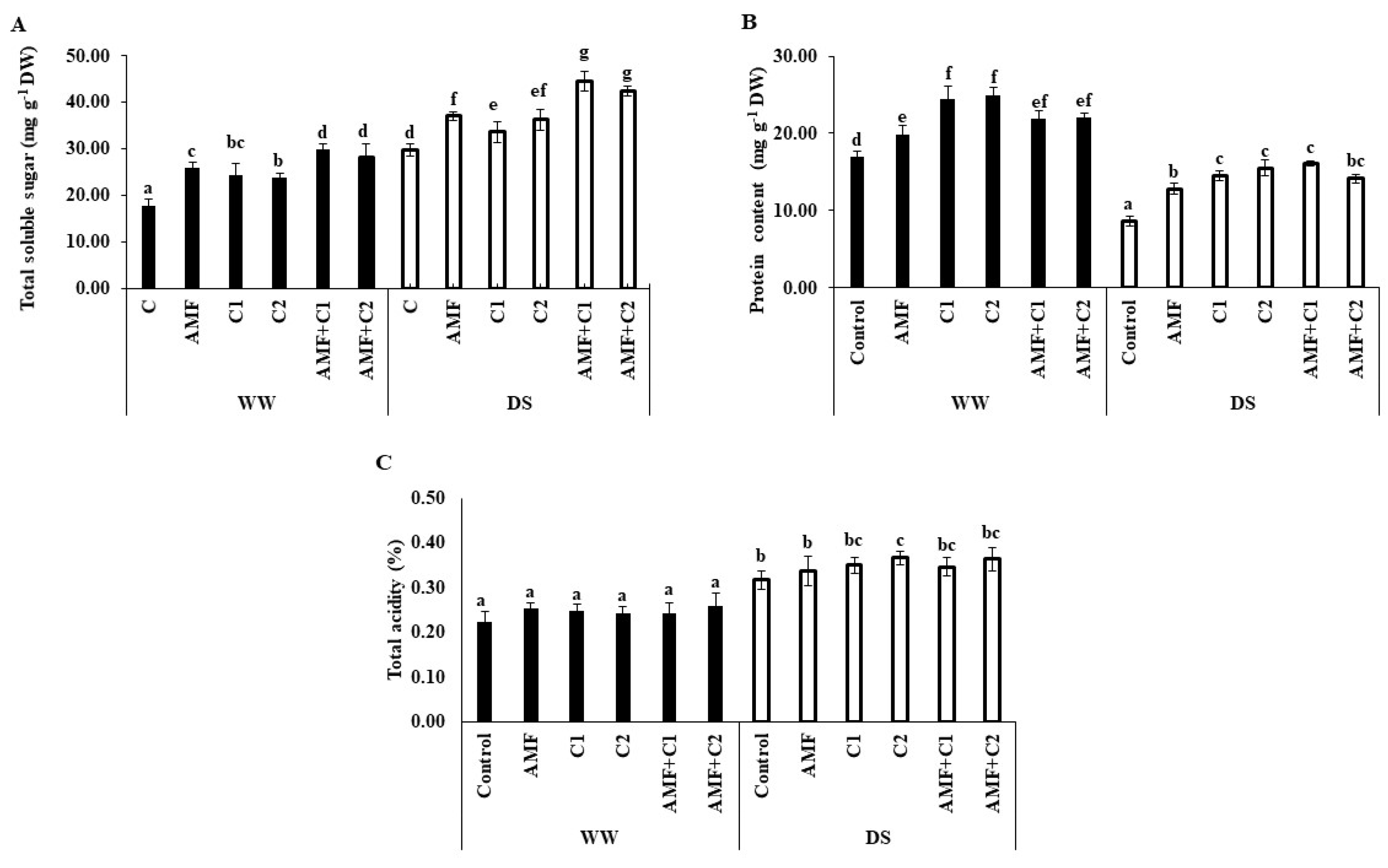
2.2. Osmotic Solutes Changes
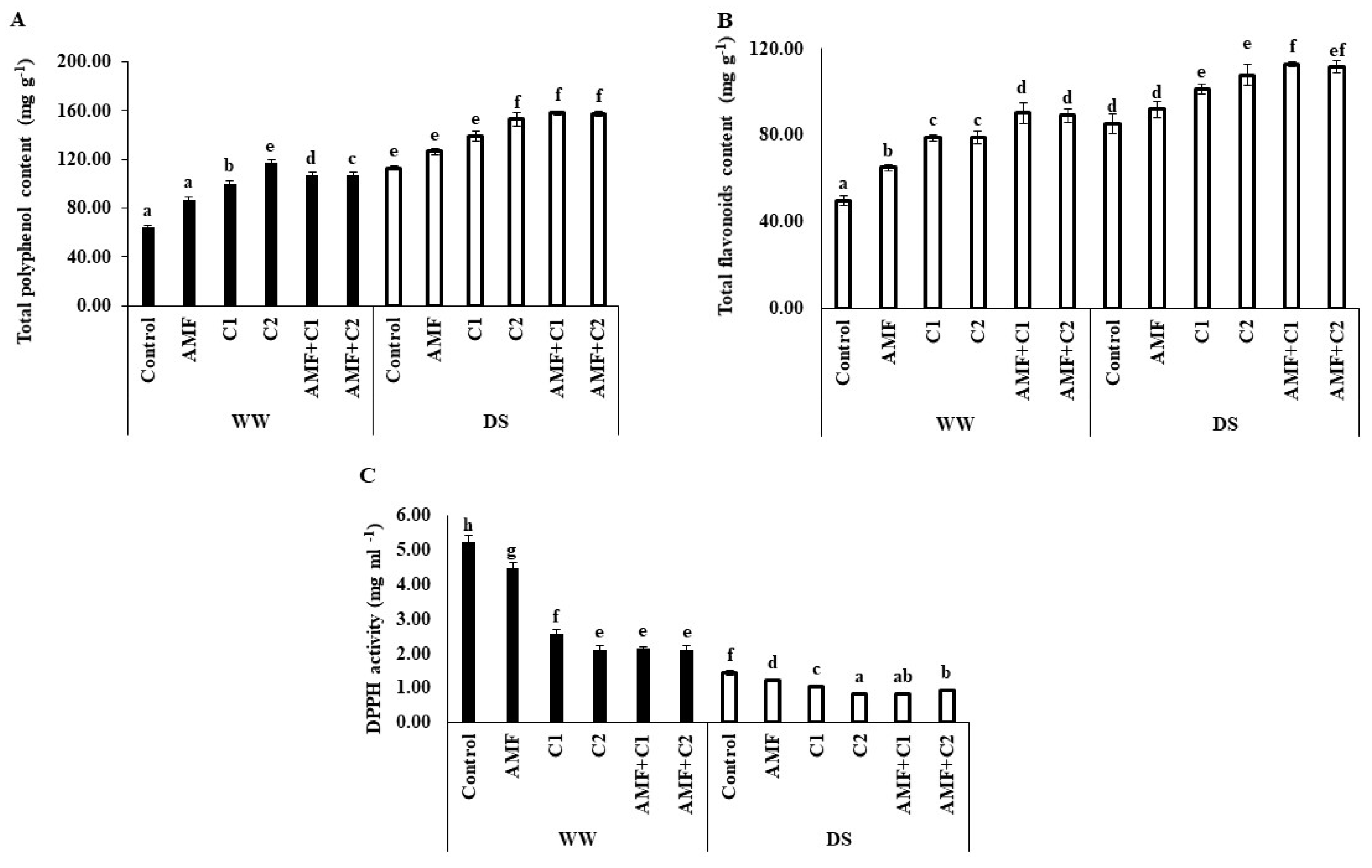
2.3. Carotenoid and Lycopene Content Changes
2.4. Ascorbic Acid and Free Amino Acids Changes
2.5. Phenol, Flavonoid, and DPPH Activity Changes
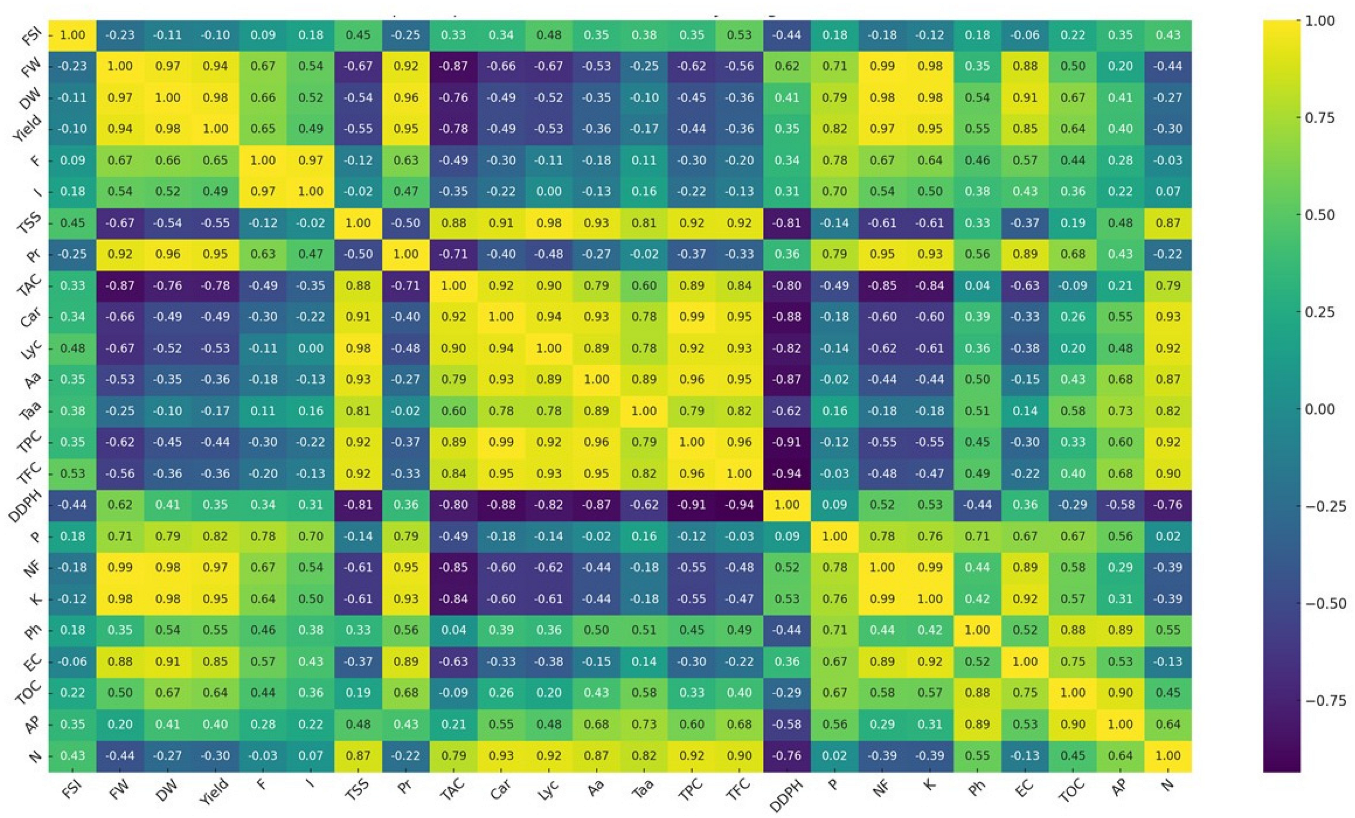
2.6. Plant Nutrient Uptake Changes
2.7. Soil Analysis
2.8. Thermal Mapping of Growth and Biochemical Characteristics of Tomato Fruit Quality
3. Discussion
4. Materials and Methods
4.1. Plant and Biostimulants/Organic Fertilizers Materials
4.2. Field Experimental Design Plant Assay
4.3. Mycorrhization Assessments
4.4. Fruit Yield and Growth Measurement
4.5. Total Soluble Sugar and Protein Content Determination in Fruit Tomatoes
4.6. Total Acidity Determination in Fruits Tomatoes
4.7. Carotenoid and Lycopene Content Determination in Tomato Fruit
4.8. Ascorbic Acid and Free Amino Acids Content Determination in Tomato Fruit
4.9. Assessment of Phenol and Flavonoid Content in Tomato Fruit
4.10. 2,2-Diphenyl-1-picrylhydrazyl Radical Scavenging Activity
4.11. Plant Nutrient Uptake Analysis
4.12. Soil Analysis
4.13. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Abd-Elmabod, S.K.; Muñoz-Rojas, M.; Jordán, A.; Anaya-Romero, M.; Phillips, J.D.; Jones, L.; Zhang, Z.; Pereira, P.; Fleskens, L.; van Der Ploeg, M. Climate Change Impacts on Agricultural Suitability and Yield Reduction in a Mediterranean Region. Geoderma 2020, 374, 114453. [Google Scholar] [CrossRef]
- Gaaloul, N.; Eslamian, S.; Katlance, R. Impacts of Climate Change and Water Resources Management in the Southern Mediterranean Countries. Water Product. J. 2021, 1, 51–72. [Google Scholar]
- El-Sanatawy, A.M.; El-Kholy, A.S.M.; Ali, M.M.A.; Awad, M.F.; Mansour, E. Maize Seedling Establishment, Grain Yield and Crop Water Productivity Response to Seed Priming and Irrigation Management in a Mediterranean Arid Environment. Agronomy 2021, 11, 756. [Google Scholar] [CrossRef]
- Amiri, N.; Lahlali, R.; Amiri, S.; El Jarroudi, M.; Khebiza, M.Y.; Messouli, M. Development of an Integrated Model to Assess the Impact of Agricultural Practices and Land Use on Agricultural Production in Morocco under Climate Stress over the next Twenty Years. Sustainability 2021, 13, 11943. [Google Scholar] [CrossRef]
- Mansour, E.; Desoky, E.-S.M.; Ali, M.M.A.; Abdul-Hamid, M.I.; Ullah, H.; Attia, A.; Datta, A. Identifying Drought-Tolerant Genotypes of Faba Bean and Their Agro-Physiological Responses to Different Water Regimes in an Arid Mediterranean Environment. Agric. Water Manag. 2021, 247, 106754. [Google Scholar] [CrossRef]
- Cook, B.I.; Anchukaitis, K.J.; Touchan, R.; Meko, D.M.; Cook, E.R. Spatiotemporal Drought Variability in the Mediterranean over the Last 900 Years. J. Geophys. Res. Atmos. 2016, 121, 2060–2074. [Google Scholar] [CrossRef] [PubMed]
- Tramblay, Y.; Koutroulis, A.; Samaniego, L.; Vicente-Serrano, S.M.; Volaire, F.; Boone, A.; Le Page, M.; Llasat, M.C.; Albergel, C.; Burak, S. Challenges for Drought Assessment in the Mediterranean Region under Future Climate Scenarios. Earth-Sci. Rev. 2020, 210, 103348. [Google Scholar] [CrossRef]
- Majid, I.; Aziane, N.; Oubbih, J.; Ramah, M.; Chakiri, S. Impact of Climate Change on Cultivated Areas and Crop Yields for Cereals and Pulses in the Zaër Region (Morocco). Ecol. Eng. Environ. Technol. 2024, 3, 14–154. [Google Scholar]
- Wakweya, R.B. Challenges and Prospects of Adopting Climate-Smart Agricultural Practices and Technologies: Implications for Food Security. J. Agric. Food Res. 2023, 14, 100698. [Google Scholar] [CrossRef]
- Lahbouki, S.; Meddich, A.; Ben-Laouane, R.; Outzourhit, A.; Pari, L. Subsurface Water Retention Technology Promotes Drought Stress Tolerance in Field-Grown Tomato. Energies 2022, 15, 6807. [Google Scholar] [CrossRef]
- Benabderrazik, K.; Kopainsky, B.; Tazi, L.; Joerin, J.; Six, J. Agricultural Intensification Can No Longer Ignore Water Conservation–A Systemic Modelling Approach to the Case of Tomato Producers in Morocco. Agric. Water Manag. 2021, 256, 107082. [Google Scholar] [CrossRef]
- Asiamah, E.; Arthur, W.; Kyei-Barfour, V.; Sarpong, F.; Ketemepi, H.K. Enhancing the Functional and Physicochemical Properties of Tomato (Solanum lycopersicum L.) Fruit through Polysaccharides Edible Dipping Technique Coating under Various Storage Conditions. Bioact. Carbohydrates Diet. Fibre 2023, 30, 100373. [Google Scholar] [CrossRef]
- Yong, K.T.; Yong, P.H.; Ng, Z.X. Tomato and Human Health: A Perspective from Post-harvest Processing, Nutrient Bio-accessibility, and Pharmacological Interaction. Food Front. 2023, 4, 1702–1719. [Google Scholar] [CrossRef]
- Chakma, R.; Saekong, P.; Biswas, A.; Ullah, H.; Datta, A. Growth, Fruit Yield, Quality, and Water Productivity of Grape Tomato as Affected by Seed Priming and Soil Application of Silicon under Drought Stress. Agric. Water Manag. 2021, 256, 107055. [Google Scholar] [CrossRef]
- Nacoon, S.; Ekprasert, J.; Riddech, N.; Mongkolthanaruk, W.; Jogloy, S.; Vorasoot, N.; Cooper, J.; Boonlue, S. Growth Enhancement of Sunchoke by Arbuscular Mycorrhizal Fungi under Drought Condition. Rhizosphere 2021, 17, 100308. [Google Scholar] [CrossRef]
- Bahadur, A.; Batool, A.; Nasir, F.; Jiang, S.; Mingsen, Q.; Zhang, Q.; Pan, J.; Liu, Y.; Feng, H. Mechanistic Insights into Arbuscular Mycorrhizal Fungi-Mediated Drought Stress Tolerance in Plants. Int. J. Mol. Sci. 2019, 20, 4199. [Google Scholar] [CrossRef] [PubMed]
- Meddich, A. Biostimulants for Resilient Agriculture—Improving Plant Tolerance to Abiotic Stress: A Concise Review. Gesunde Pflanz. 2023, 75, 709–727. [Google Scholar] [CrossRef]
- Tian, H.; Jia, Z.; Liu, W.; Wei, X.; Wang, H.; Bao, G.; Li, J.; Zhou, Q. Effects of Arbuscular Mycorrhizal Fungi on Growth and Nutrient Accumulation of Oat under Drought Conditions. Agronomy 2023, 13, 2580. [Google Scholar] [CrossRef]
- Mitra, D.; Dam, P.; Mondal, R.; Mahakur, B.; Al-Tawaha, A.R.M.; Sangeetha, J.; Thangadurai, D.; Chippalakatti, P. Application of Arbuscular Mycorrhiza Fungi in Agricultural and Horticultural Crops. In Mycorrhizal Technology; Apple Academic Press: Cambridge, MA, USA, 2023; pp. 55–68. [Google Scholar]
- Trinchera, A.; Warren Raffa, D. Weeds: An Insidious Enemy or a Tool to Boost Mycorrhization in Cropping Systems? Microorganisms 2023, 11, 334. [Google Scholar] [CrossRef]
- Wang, M.; Wang, Z.; Guo, M.; Qu, L.; Biere, A. Effects of Arbuscular Mycorrhizal Fungi on Plant Growth and Herbivore Infestation Depend on Availability of Soil Water and Nutrients. Front. Plant Sci. 2023, 14, 1101932. [Google Scholar] [CrossRef]
- Klink, S.; Giesemann, P.; Hubmann, T.; Pausch, J. Stable C and N Isotope Natural Abundances of Intraradical Hyphae of Arbuscular Mycorrhizal Fungi. Mycorrhiza 2020, 30, 773–780. [Google Scholar] [CrossRef] [PubMed]
- Tatewaki, Y.; Higo, M.; Isobe, K. Impacts of Tillage Practices on Growth, Phosphorus Uptake, and Yield of Maize in Controlled and Field-Based Studies in Relation to Arbuscular Mycorrhizal Fungi. Appl. Microbiol. 2023, 3, 358–374. [Google Scholar] [CrossRef]
- Lahbouki, S.; Fernando, A.L.; Rodrigues, C.; Ben-Laouane, R.; Ait-El-Mokhtar, M.; Outzourhit, A.; Meddich, A. Effects of Humic Substances and Mycorrhizal Fungi on Drought-Stressed Cactus: Focus on Growth, Physiology, and Biochemistry. Plants 2023, 12, 4156. [Google Scholar] [CrossRef] [PubMed]
- Wanlin, L.I.; Yan, X. Effects of Polystyrene Microplastics, Simulated Acid Rain and Arbuscular Mycorrhizal Fungi on the Growth of Trifolium Repens and Soil Microbial Community Composition. Pedosphere 2024, 3, 424–437. [Google Scholar]
- Bisht, A.; Garg, N. AMF Species Improve Yielding Potential of Cd Stressed Pigeonpea Plants by Modulating Sucrose-Starch Metabolism, Nutrients Acquisition and Soil Microbial Enzymatic Activities. Plant Growth Regul. 2022, 96, 409–430. [Google Scholar] [CrossRef]
- Leventis, G.; Tsiknia, M.; Feka, M.; Ladikou, E.V.; Papadakis, I.E.; Chatzipavlidis, I.; Papadopoulou, K.; Ehaliotis, C. Arbuscular Mycorrhizal Fungi Enhance Growth of Tomato under Normal and Drought Conditions, via Different Water Regulation Mechanisms. Rhizosphere 2021, 19, 100394. [Google Scholar] [CrossRef]
- Besharati, J.; Shirmardi, M.; Meftahizadeh, H.; Ardakani, M.D.; Ghorbanpour, M. Changes in Growth and Quality Performance of Roselle (Hibiscus sabdariffa L.) in Response to Soil Amendments with Hydrogel and Compost under Drought Stress. South African J. Bot. 2022, 145, 334–347. [Google Scholar] [CrossRef]
- Lahbouki, S.; Ben-Laouane, R.; Outzourhit, A.; Meddich, A. The Combination of Vermicompost and Arbuscular Mycorrhizal Fungi Improves the Physiological Properties and Chemical Composition of Opuntia ficus-indica under Semi-Arid Conditions in the Field. Arid L. Res. Manag. 2022, 284–309. [Google Scholar] [CrossRef]
- Rashtbari, M.; Hossein Ali, A.; Ghorchiani, M. Effect of Vermicompost and Municipal Solid Waste Compost on Growth and Yield of Canola under Drought Stress Conditions. Commun. Soil Sci. Plant Anal. 2020, 51, 2215–2222. [Google Scholar] [CrossRef]
- El Amerany, F.; Rhazi, M.; Wahbi, S.; Taourirte, M.; Meddich, A. The Effect of Chitosan, Arbuscular Mycorrhizal Fungi, and Compost Applied Individually or in Combination on Growth, Nutrient Uptake, and Stem Anatomy of Tomato. Sci. Hortic. 2020, 261, 109015. [Google Scholar] [CrossRef]
- Paymaneh, Z.; Sarcheshmehpour, M.; Mohammadi, H.; Hesni, M.A. Vermicompost and/or Compost and Arbuscular Mycorrhizal Fungi Are Conducive to Improving the Growth of Pistachio Seedlings to Drought Stress. Appl. Soil Ecol. 2023, 182, 104717. [Google Scholar] [CrossRef]
- Pant, A.; Radovich, T.J.K.; Hue, N.V.; Arancon, N.Q. Effects of Vermicompost Tea (Aqueous Extract) on Pak Choi Yield, Quality, and on Soil Biological Properties. Compost Sci. Util. 2011, 19, 279–292. [Google Scholar] [CrossRef]
- Erdal, İ.; Ekinci, K. Effects of Composts and Vermicomposts Obtained from Forced Aerated and Mechanically Turned Composting Method on Growth, Mineral Nutrition and Nutrient Uptake of Wheat. J. Plant Nutr. 2020, 43, 1343–1355. [Google Scholar] [CrossRef]
- Tammam, A.A.; Rabei Abdel Moez Shehata, M.; Pessarakli, M.; El-Aggan, W.H. Vermicompost and Its Role in Alleviation of Salt Stress in Plants–I. Impact of Vermicompost on Growth and Nutrient Uptake of Salt-Stressed Plants. J. Plant Nutr. 2023, 46, 1446–1457. [Google Scholar] [CrossRef]
- Bezabeh, M.W.; Haile, M.; Sogn, T.A.; Eich-Greatorex, S. Yield, Nutrient Uptake, and Economic Return of Faba Bean (Vicia faba L.) in Calcareous Soil as Affected by Compost Types. J. Agric. Food Res. 2021, 6, 100237. [Google Scholar] [CrossRef]
- Lahbouki, S.; Ben-Laouane, R.; Anli, M.; Boutasknit, A.; Ait-Rahou, Y.; Ait-El-Mokhtar, M.; El Gabardi, S.; Douira, A.; Wahbi, S.; Outzourhit, A.; et al. Arbuscular Mycorrhizal Fungi and/or Organic Amendment Enhance the Tolerance of Prickly Pear (Opuntia ficus-indica) under Drought Stress. J. Arid Environ. 2022, 199, 104703. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Ahmad, M.; Zulfiqar, U.; Yaqoob, S.; Hussain, S.; Niazi, N.K.; Gastelbondo, M.; Al-Ashkar, I.; Elshikh, M.S. Vermicompost Application Upregulates Morpho-Physiological and Antioxidant Defense to Conferring Drought Tolerance in Wheat. Plant Stress 2024, 11, 100360. [Google Scholar] [CrossRef]
- Lamaizi, S.; Meddich, A.; Boutasknit, A.; Anli, M.; Lahbouki, S.; El Fels, L.; Ouhdouch, Y.; Hafidi, M. Application of Olive-Mill-Wastewater-Compost in Combination with Symbiotic Microorganisms Improves the Physiological, Biochemical Performance and Tolerance of Tomato (Solanum lycopersicum) Under Drought Stress. Gesunde Pflanz. 2023, 75, 1719–1735. [Google Scholar] [CrossRef]
- Olayiwola, V.A.; Abiodun, F.O. Effects of Arbuscular Mycorrhiza Fungi (Glomus mossae) and Compost on Early Growth Performance of Parkia Biglobosa. Curr. J. Appl. Sci. Technol. 2019, 37, 1–8. [Google Scholar] [CrossRef]
- Benaffari, W.; Boutasknit, A.; Anli, M.; Ait-El-Mokhtar, M.; Ait-Rahou, Y.; Ben-Laouane, R.; Ben Ahmed, H.; Mitsui, T.; Baslam, M.; Meddich, A. The Native Arbuscular Mycorrhizal Fungi and Vermicompost-Based Organic Amendments Enhance Soil Fertility, Growth Performance, and the Drought Stress Tolerance of Quinoa. Plants 2022, 11, 393. [Google Scholar] [CrossRef]
- Ouhaddou, R.; Ech-chatir, L.; Anli, M.; Ben-Laouane, R.; Boutasknit, A.; Meddich, A. Secondary Metabolites, Osmolytes and Antioxidant Activity as the Main Attributes Enhanced by Biostimulants for Growth and Resilience of Lettuce to Drought Stress. Gesunde Pflanz. 2023, 75, 1737–1753. [Google Scholar] [CrossRef]
- Dong, S.; Hu, F.; Alamusa; Ma, Q.; Liu, Z. Arbuscular Mycorrhizal Fungi Enhance the Drought Resistance More Significantly of the Late-successional Psammophytes than That of the Early Ones. Restor. Ecol. 2023, 31, e13832. [Google Scholar] [CrossRef]
- Quiroga, G.; Erice, G.; Aroca, R.; Delgado-Huertas, A.; Ruiz-Lozano, J.M. Elucidating the Possible Involvement of Maize Aquaporins and Arbuscular Mycorrhizal Symbiosis in the Plant Ammonium and Urea Transport under Drought Stress Conditions. Plants 2020, 9, 148. [Google Scholar] [CrossRef] [PubMed]
- Cavagnaro, T.R. Biologically Regulated NutrientSupply Systems. Compost AndArbuscular Mycorrhizas-A Review. Adv. Agron. 2015, 129, 293–321. [Google Scholar] [CrossRef]
- Liu, M.; Shen, Y.; Li, Q.; Xiao, W.; Song, X. Arbuscular Mycorrhizal Fungal Colonization and Soil PH Induced by Nitrogen and Phosphorus Additions Affects Leaf C: N: P Stoichiometry in Chinese Fir (Cunninghamia lanceolata) Forests. Plant Soil 2021, 461, 421–440. [Google Scholar] [CrossRef]
- Li, H.; Zhang, X.; Hou, X.; Du, T. Developmental and Water Deficit-Induced Changes in Hydraulic Properties and Xylem Anatomy of Tomato Fruit and Pedicels. J. Exp. Bot. 2021, 72, 2741–2756. [Google Scholar] [CrossRef]
- Zimmermann, J.; Link, R.M.; Hauck, M.; Leuschner, C.; Schuldt, B. 60-Year Record of Stem Xylem Anatomy and Related Hydraulic Modification under Increased Summer Drought in Ring-and Diffuse-Porous Temperate Broad-Leaved Tree Species. Trees 2021, 35, 919–937. [Google Scholar] [CrossRef]
- Li, H.; Hou, X.; Du, T. Responses of Tomato Fruit Water Balance and Xylem Hydraulic Property of Pedicel and Calyx to Water Deficit and Salinity Stress. Environ. Exp. Bot. 2023, 206, 105195. [Google Scholar] [CrossRef]
- Kaur, S.; Suseela, V. Unraveling Arbuscular Mycorrhiza-Induced Changes in Plant Primary and Secondary Metabolome. Metabolites 2020, 10, 335. [Google Scholar] [CrossRef]
- Hou, L.; Li, M.; Zhang, C.; Liu, N.; Liu, X.; Bo, W.; Pang, X.; Li, Y. Comparative Transcriptomic Analyses of Different Jujube Cultivars Reveal the Co-Regulation of Multiple Pathways during Fruit Cracking. Genes 2022, 13, 105. [Google Scholar] [CrossRef]
- Zhang, X.; Yang, H.; Du, T. Coupled Mechanisms of Water Deficit and Soil Salinity Affecting Tomato Fruit Growth. Agric. Water Manag. 2024, 295, 108747. [Google Scholar] [CrossRef]
- Jiaying, M.; Tingting, C.; Jie, L.; Weimeng, F.; Baohua, F.; Guangyan, L.; Hubo, L.; Juncai, L.; Zhihai, W.; Longxing, T. Functions of Nitrogen, Phosphorus and Potassium in Energy Status and Their Influences on Rice Growth and Development. Rice Sci. 2022, 29, 166–178. [Google Scholar] [CrossRef]
- Püschel, D.; Bitterlich, M.; Rydlová, J.; Bukovská, P.; Sudová, R.; Jansa, J. Benefits in Plant N Uptake via the Mycorrhizal Pathway in Ample Soil Moisture Persist under Severe Drought. Soil Biol. Biochem. 2023, 187, 109220. [Google Scholar] [CrossRef]
- Khalediyan, N.; Weisany, W.; Schenk, P.M. Arbuscular Mycorrhizae and Rhizobacteria Improve Growth, Nutritional Status and Essential Oil Production in Ocimum Basilicum and Satureja Hortensis. Ind. Crops Prod. 2021, 160, 113163. [Google Scholar] [CrossRef]
- Das, D.; Abbhishek, K.; Banik, P.; Bhattacharya, P. A Valorisation Approach in Recycling of Organic Wastes Using Low-Grade Rock Minerals and Microbial Culture through Vermicomposting. Environ. Chall. 2021, 5, 100225. [Google Scholar] [CrossRef]
- Chen, Q.; Qu, Z.; Ma, G.; Wang, W.; Dai, J.; Zhang, M.; Wei, Z.; Liu, Z. Humic Acid Modulates Growth, Photosynthesis, Hormone and Osmolytes System of Maize under Drought Conditions. Agric. Water Manag. 2022, 263, 107447. [Google Scholar] [CrossRef]
- Siles, J.A.; José, M.; González-Pérez, J.A.; Fernández-Pérez, V.; García-Díaz, C.; Moreno, J.L.; García, C.; Bastida, F. Long-Term Restoration with Organic Amendments Is Clearer Evidenced by Soil Organic Matter Composition than by Changes in Microbial Taxonomy and Functionality. Appl. Soil Ecol. 2024, 198, 105383. [Google Scholar] [CrossRef]
- Okla, M.K.; Rubnawaz, S.; Dawoud, T.M.; Al-Amri, S.; El-Tayeb, M.A.; Abdel-Maksoud, M.A.; Akhtar, N.; Zrig, A.; Abdelgayed, G.; Abdelgawad, H. Laser Light Treatment Improves the Mineral Composition, Essential Oil Production and Antimicrobial Activity of Mycorrhizal Treated Pelargonium graveolens. Molecules 2022, 27, 1752. [Google Scholar] [CrossRef] [PubMed]
- Bi, Y.; Wu, C.; Wang, S.; Gao, X.; Xue, C.; Yang, W.; Li, M.; Xiao, L.; Christie, P. Combined Arbuscular Mycorrhizal Inoculation and Loess Amendment Improve Rooting and Revegetation Post-Mining. Rhizosphere 2022, 23, 100560. [Google Scholar] [CrossRef]
- Kaur, H.; Manna, M.; Thakur, T.; Gautam, V.; Salvi, P. Imperative Role of Sugar Signaling and Transport during Drought Stress Responses in Plants. Physiol. Plant. 2021, 171, 833–848. [Google Scholar] [CrossRef]
- Francesca, S.; Cirillo, V.; Raimondi, G.; Maggio, A.; Barone, A.; Rigano, M.M. A Novel Protein Hydrolysate-Based Biostimulant Improves Tomato Performances under Drought Stress. Plants 2021, 10, 783. [Google Scholar] [CrossRef]
- Soussani, F.E.; Boutasknit, A.; Ben-Laouane, R.; Benkirane, R.; Baslam, M.; Meddich, A. Arbuscular Mycorrhizal Fungi and Compost-Based Biostimulants Enhance Fitness, Physiological Responses, Yield, and Quality Traits of Drought-Stressed Tomato Plants. Plants 2023, 12, 1856. [Google Scholar] [CrossRef]
- Khosravi Shakib, A.; Rezaei Nejad, A.; Khandan Mirkohi, A.; Kalate Jari, S. Vermicompost and Manure Compost Reduce Water-Deficit Stress in Pot Marigold (Calendula Officinalis L. Cv. Candyman Orange). Compost Sci. Util. 2019, 27, 61–68. [Google Scholar] [CrossRef]
- Ganugi, P.; Fiorini, A.; Tabaglio, V.; Capra, F.; Zengin, G.; Bonini, P.; Caffi, T.; Puglisi, E.; Trevisan, M.; Lucini, L. The Functional Profile and Antioxidant Capacity of Tomato Fruits Are Modulated by the Interaction between Microbial Biostimulants, Soil Properties, and Soil Nitrogen Status. Antioxidants 2023, 12, 520. [Google Scholar] [CrossRef]
- Etesami, H.; Jeong, B.R. Contribution of Arbuscular Mycorrhizal Fungi, Phosphate–Solubilizing Bacteria, and Silicon to P Uptake by Plant: A Review. Front. Plant Sci. 2021, 12, 1355. [Google Scholar] [CrossRef]
- Sandmann, G. Diversity and Origin of Carotenoid Biosynthesis: Its History of Coevolution towards Plant Photosynthesis. New Phytol. 2021, 232, 479–493. [Google Scholar] [CrossRef] [PubMed]
- Fanasca, S.; Colla, G.; Maiani, G.; Venneria, E.; Rouphael, Y.; Azzini, E.; Saccardo, F. Changes in Antioxidant Content of Tomato Fruits in Response to Cultivar and Nutrient Solution Composition. J. Agric. Food Chem. 2006, 54, 4319–4325. [Google Scholar] [CrossRef] [PubMed]
- El-Beltagi, H.S.; Mohamed, H.I.; Sofy, M.R. Role of Ascorbic Acid, Glutathione and Proline Applied as Singly or in Sequence Combination in Improving Chickpea Plant through Physiological Change and Antioxidant Defense under Different Levels of Irrigation Intervals. Molecules 2020, 25, 1702. [Google Scholar] [CrossRef] [PubMed]
- Talbi, S.; Rojas, J.A.; Sahrawy, M.; Rodríguez-Serrano, M.; Cárdenas, K.E.; Debouba, M.; Sandalio, L.M. Effect of Drought on Growth, Photosynthesis and Total Antioxidant Capacity of the Saharan Plant Oudeneya Africana. Environ. Exp. Bot. 2020, 176, 104099. [Google Scholar] [CrossRef]
- Rodríguez-Arce, E.; Saldías, M. Antioxidant Properties of Flavonoid Metal Complexes and Their Potential Inclusion in the Development of Novel Strategies for the Treatment against Neurodegenerative Diseases. Biomed. Pharmacother. 2021, 143, 112236. [Google Scholar] [CrossRef]
- Singh, A.; Roychoudhury, A. Role of Phenolic Acids and Flavonoids in the Mitigation of Environmental Stress in Plants. In Biology and Biotechnology of Environmental Stress Tolerance in Plants; Apple Academic Press: Cambridge, MA, USA, 2023; pp. 227–248. [Google Scholar]
- Wang, Y.; Zhang, W.; Liu, W.; Ahammed, G.J.; Wen, W.; Guo, S.; Shu, S.; Sun, J. Auxin Is Involved in Arbuscular Mycorrhizal Fungi-Promoted Tomato Growth and NADP-Malic Enzymes Expression in Continuous Cropping Substrates. BMC Plant Biol. 2021, 21, 48. [Google Scholar] [CrossRef]
- Al Jaouni, S.; Selim, S.; Hassan, S.H.; Mohamad, H.S.H.; Wadaan, M.A.M.; Hozzein, W.N.; Asard, H.; AbdElgawad, H. Vermicompost Supply Modifies Chemical Composition and Improves Nutritive and Medicinal Properties of Date Palm Fruits from Saudi Arabia. Front. Plant Sci. 2019, 10, 424. [Google Scholar] [CrossRef]
- Lahbouki, S.; Anli, M.; El Gabardi, S.; Ait-El-Mokhtar, M.; Ben-Laouane, R.; Boutasknit, A.; Ait-Rahou, Y.; Outzourhit, A.; Wahbi, S.; Douira, A. Evaluation of Arbuscular Mycorrhizal Fungi and Vermicompost Supplementation on Growth, Phenolic Content and Antioxidant Activity of Prickly Pear Cactus (Opuntia ficus-indica). Plant Biosyst. Int. J. Deal. All Asp. Plant Biol. 2021, 156, 882–892. [Google Scholar] [CrossRef]
- Idris, N.S.; Khandaker, M.M.; Rashid, Z.M.; Majrashi, A.; Alenazi, M.M.; Nor, Z.M.; Mohd Adnan, A.F.; Mat, N. Polyphenolic Compounds and Biological Activities of Leaves and Fruits of Syzygium samarangense Cv.‘Giant Green’at Three Different Maturities. Horticulturae 2023, 9, 326. [Google Scholar] [CrossRef]
- Lahbouki, S.; Ech-chatir, L.; Er-Raki, S.; Outzourhit, A.; Meddich, A. Improving Drought Tolerance of Opuntia ficus-indica under Field Using Subsurface Water Retention Technology: Changes in Physiological and Biochemical Parameters. Can. J. Soil Sci. 2022, 102, 888–898. [Google Scholar] [CrossRef]
- Phillips, J.M.; Hayman, D.S. Improved Procedures for Clearing Roots and Staining Parasitic and Vesicular-Arbuscular Mycorrhizal Fungi for Rapid Assessment of Infection. Trans. Br. Mycol. Soc. 1970, 55, 158–161. [Google Scholar] [CrossRef]
- Trouvelot, A.; Kough, J.L. Mesure Du Taux de Mycorhization VA d’un Système Radiculaire. Recherche de Méthode d’estimation Ayant Une Signification Fonctionnelle. In Physiological and Genetical Aspects of Mycorrhizae; Gianinazzi-Pearson, V., Gianinazzi, S., Eds.; INRA: Paris, France, 1986; pp. 217–221. [Google Scholar]
- Tao, H.; Sun, H.; Wang, Y.; Wang, X.; Guo, Y. Effects of Water Stress on Quality and Sugar Metabolism in ‘Gala’Apple Fruit. Hortic. Plant J. 2023, 9, 60–72. [Google Scholar] [CrossRef]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.A.; Smith, F. Colorimetric Method for Determination of Sugars and Related Substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Bradford, M.M. A Rapid and Sensitive Method for the Quantitation of Microgram Quantities of Protein Utilizing the Principle of Protein-Dye Binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Pastori, P.L.; FilgueiraS, R.M.C.; Oster, A.H.; Barbosa, M.G.; Silveira, M.R.S.D.A.; Paiva, L.G.G. Postharvest Quality of Tomato Fruits Bagged with Nonwoven Fabric (TNT). Rev. Colomb. Cienc. Hortícolas 2017, 11, 80–88. [Google Scholar] [CrossRef]
- Zhou, X.; Huang, W.; Kong, W.; Ye, H.; Dong, Y.; Casa, R. Assessment of Leaf Carotenoids Content with a New Carotenoid Index: Development and Validation on Experimental and Model Data. Int. J. Appl. Earth Obs. Geoinf. 2017, 57, 24–35. [Google Scholar] [CrossRef]
- Roldán-Gutiérrez, J.M.; de Castro, M.D.L. Lycopene: The Need for Better Methods for Characterization and Determination. TrAC Trends Anal. Chem. 2007, 26, 163–170. [Google Scholar] [CrossRef]
- Adrian, J.; Peiró Esteban, J.M. Análisis Nutricional de los Alimentos; Editorial Acribia SA: Madrid, Spain, 2000; ISBN 8420009199. [Google Scholar]
- Lee, Y.P.; Takahashi, T. An Improved Colorimetric Determination of Amino Acids with the Use of Ninhydrin. Anal. Biochem. 1966, 14, 71–77. [Google Scholar] [CrossRef]
- Singleton, V.L.; Rossi, J.A. Colorimetry of Total Phenolics with Phosphomolybdic-Phosphotungstic Acid Reagents. Am. J. Enol. Vitic. 1965, 16, 144–158. [Google Scholar] [CrossRef]
- Tohidi, B.; Rahimmalek, M.; Arzani, A. Essential Oil Composition, Total Phenolic, Flavonoid Contents, and Antioxidant Activity of Thymus Species Collected from Different Regions of Iran. Food Chem. 2017, 220, 153–161. [Google Scholar] [CrossRef] [PubMed]
- Aruwa, C.E.; Amoo, S.O.; Kudanga, T. Extractable and Macromolecular Antioxidants of Opuntia ficus-indica Cladodes: Phytochemical Profiling, Antioxidant and Antibacterial Activities. South Afr. J. Bot. 2019, 125, 402–410. [Google Scholar] [CrossRef]
- Watts, S.; Halliwell, L. Essential Environmental Science: Methods & Techniques; Psychology Press: London, UK, 1996; ISBN 0415132460. [Google Scholar]
- Watts, S.; Halliwell, L. Appendix 3–Detailed Field and Chemical Methods for Soil. In Essential Environmental Science, Methods & Techniques; Routledge: London, UK, 1996; Volume 548, pp. 475–505. [Google Scholar]
- Wolf, B. A Comprehensive System of Leaf Analyses and Its Use for Diagnosing Crop Nutrient Status. Commun. Soil Sci. Plant Anal. 1982, 13, 1035–1059. [Google Scholar] [CrossRef]
- Aubert, G. Methodes d’Analyses Des Sols: Documents de Travail Tous Droits Reserves; Centre Régional de Documentation Pédagogique: Marseille, France, 1978. [Google Scholar]
- Olsen, S.R.; Sommers, L.E. Phosphorus in Methods of Soil Analysis Part 2. Chem. Microbiol. Prop. Agron. Monogr. 1982, 9, 421–422. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Yield/Plant | TD (cm) | LD (cm) | FSI | FW (g/Plant) | DW (g/Plant) | F % | I% | ||
|---|---|---|---|---|---|---|---|---|---|
| WW | Control | 21.33 ± 1.52 e | 4.91 ± 0.52 cd | 8.36 ± 0.77 f-h | 1.70 ± 0.04 e | 644.67 ± 22.32 f | 18.35 ± 1.75 d | 37.84 ± 9.56 bc | 22.43 ± 1.14 b |
| AMF | 25.33 ± 2.13 f | 5.26 ± 0.51 ef | 8.69 ± 0.24 gh | 1.65 ± 0.07 de | 674.67 ± 19.22 g | 20.03 ± 1.68 e | 72.16 ± 16.36 g | 54.11 ± 3.52 g | |
| C1 | 31.67 ± 1.54 g | 4.95 ± 0.27 de | 8.92 ± 0.12 h | 1.80 ± 0.03 f | 689.67 ± 22.57 gh | 22.74 ± 2.09 ef | 48.16 ± 8.36 de | 26.67 ± 0.96 bc | |
| C2 | 38.00 ± 1.36 i | 5.03 ± 0.22 e | 9.06 ± 0.09 h | 1.80 ± 0.03 f | 702.00 ± 14.25 h | 24.02 ± 1.41 f | 47.55 ± 7.25 d | 27.82 ± 1.35 c | |
| AMF+C1 | 37.00 ± 1.42 hi | 6.88 ± 0.44 g | 9.27 ± 0.12 ij | 1.34 ± 0.05 ab | 704.00 ± 11.65 h | 23.55 ± 2.23 ef | 69.13 ± 15.78 g | 45.76 ± 3.66 f | |
| AMF+C2 | 34.67 ± 1.26 gh | 6.93 ± 0.63 g | 9.47 ± 0.17 j | 1.36 ± 0.05 ab | 703.33 ± 12.47 h | 24.89 ± 1.45 g | 61.37 ± 9.45 fg | 40.85 ± 2.32 e | |
| DS | Control | 10.00 ± 1.46 a | 2.71 ± 0.52 a | 4.02 ± 0.32 a | 1.48 ± 0.07 c | 416.00 ± 13.66 a | 10.28 ± 1.08 a | 22.39 ± 6.38 a | 14.19 ± 0.68 a |
| AMF | 14.33 ± 2.06 b | 3.43 ± 0.22 b | 5.52 ± 0.08 b | 1.60 ± 0.03 d | 447.67 ± 9.52 b | 12.61 ± 0.95 b | 51.65 ± 8.28 e | 34.19 ± 2.36 d | |
| C1 | 16.33 ± 2.36 cd | 4.08 ± 0.12 bc | 5.85 ± 0.30 cd | 1.43 ± 0.03 bc | 478.33 ± 25.66 c | 14.76 ± 1.31 bc | 31.84 ± 17.56 b | 20.85 ± 1.27 b | |
| C2 | 16.78 ± 1.45 d | 4.21 ± 0.02 c | 6.03 ± 0.36 d | 1.43 ± 0.04 bc | 495.11 ± 20.52 d | 15.37 ± 0.86 c | 34.66 ± 5.42 b | 23.62 ± 1.09 b | |
| AMF+C1 | 16.00 ± 1.76 c | 4.22 ± 0.28 c | 6.01 ± 0.15 d | 1.42 ± 0.06 b | 489.67 ± 16.34 cd | 15.19 ± 1.28 c | 48.5 ± 8.32 de | 33.58 ± 1.55 d | |
| AMF+C2 | 15.00 ± 2.43 bc | 4.09 ± 0.24 bc | 6.55 ± 0.27 e | 1.60 ± 0.05 d | 511.67 ± 35.75 de | 15.30 ± 1.12 c | 42.86 ± 8.67 cd | 28.71 ± 0.68 c |
| Water Regime | Treatments | Phosphorus (%) | Nitrogen (%) | Potassium (%) |
|---|---|---|---|---|
| WW | Control | 0.06 ± 0.01 c | 1.26 ± 0.16 d | 2.77 ± 0.41 f |
| AMF | 0.12 ± 0.02 d | 1.37 ± 0.12 ef | 2.68 ± 0.36 g | |
| C1 | 0.11 ± 0.01 de | 1.44 ± 0.18 fg | 3.12 ± 0.49 hi | |
| C2 | 0.14 ± 0.03 ef | 1.53 ± 0.27 h | 3.24 ± 0.52 ig | |
| AMF+C1 | 0.16 ± 0.02 f | 1.50 ± 0.12 h | 3.45 ± 0.29 g | |
| AMF+C2 | 0.14 ± 0.03 ef | 1.48 ± 0.11 gh | 3.36 ± 0.48 g | |
| DS | Control | 0.05 ± 0.00 a | 0.86 ± 0.19 a | 1.07 ± 0.29 a |
| AMF | 0.08 ± 0.00 cd | 0.88 ± 0.22 ab | 1.15 ± 0.32 b | |
| C1 | 0.05 ± 0.01 bc | 0.95 ± 0.19 b | 1.44 ± 0.24 c | |
| C2 | 0.10 ± 0.02 de | 0.99 ± 0.16 bc | 1.78 ± 0.16 de | |
| AMF+C1 | 0.12 ± 0.01 e | 1.06 ± 0.18 c | 1.86 ± 0.16 e | |
| AMF+C2 | 0.06 ± 0.00 c | 1.03 ± 0.11 c | 1.75 ± 0.35 de |
| Water Regime | WW | DS | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Treatments | Control | AMF | C1 | C2 | AMF+C1 | AMF+C2 | Control | AMF | C1 | C2 | AMF+C1 | AMF+C2 |
| pH | 7.74 ± 0.45 a | 7.80 ± 0.33 ab | 7.84 ± 0.52 bc | 7.93 ± 0.82 c | 7.86 ± 0.63 bc | 7.91 ± 0.28 c | 7.74 ± 0.51 a | 7.84 ± 0.76 bc | 7.81 ± 0.38 a–c | 7.86 ± 0.34 bc | 7.88 ± 0.28 bc | 7.88 ± 0.31 bc |
| EC (mS cm−1) | 1.75 ± 0.36 cd | 1.72 ± 0.54 cd | 1.82 ± 0.41 cd | 1.79 ± 0.34 cd | 1.83 ± 0.35 d | 1.81 ± 0.41 cd | 1.52 ± 0.57 a | 1.55 ± 0.45 ab | 1.67 ± 0.13 bc | 1.65 ± 0.28 bc | 1.72 ± 0.35 cd | 1.72 ± 0.21 cd |
| TOC (%) | 0.92 ± 0.12 b | 1.15 ± 0.31 c | 1.35 ± 0.28 e–g | 1.48 ± 0.19 gh | 1.43 ± 0.25 fg | 1.52 ± 0.41 h | 0.88 ± 0.11 a | 0.95 ± 0.08 b | 1.25 ± 0.19 d | 1.21 ± 0.25 d | 1.39 ± 0.14 e–g | 1.41 ± 0.32 e–g |
| AP (mg/kg) | 26.74 ± 3.36 a | 43.3 ± 4.15 c | 84.36 ± 3.86 ef | 89.19 ± 4.18 fg | 102.75 ± 5.85 i | 91.95 ± 6.22 gh | 33.84 ± 3.68 b | 53.48 ± 5.32 c | 72.19 ± 5.85 d | 88.32 ± 4.86 fg | 99.40 ± 5.85 hi | 99.39 ± 5.56 hi |
| N (g/kg) | 0.70 ± 0.09 a | 0.98 ± 0.12 cd | 0.84 ± 0.09 b | 1.05 ± 0.15 de | 1.02 ± 0.19 de | 1.08 ± 0.11 e | 0.85 ± 0.06 b | 1.12 ± 0.25 e–g | 1.27 ± 0.19 fg | 1.30 ± 0.21 g | 1.34 ± 0.21 g | 1.34 ± 0.14 g |
| Treatments | Water Regime | |
|---|---|---|
| Control | well-watered | Plants non-inoculated and non-amended |
| AMF | water-stressed | Plants inoculated with AMF (200 g/plot) |
| C1 | Plants amended with vermicompost at 0.6 kg/plot (5 t ha−1) | |
| C2 | Plants amended with vermicompost at 1.2 kg/plot (10 t ha−1) | |
| AMF+C1 | Combined application of AMF and C1 | |
| AMF+C2 | Combined application of AMF and C2 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Lahbouki, S.; Hashem, A.; Kumar, A.; Abd_Allah, E.F.; Meddich, A. Integration of Horse Manure Vermicompost Doses and Arbuscular Mycorrhizal Fungi to Improve Fruit Quality, and Soil Fertility in Tomato Field Facing Drought Stress. Plants 2024, 13, 1449. https://doi.org/10.3390/plants13111449
Lahbouki S, Hashem A, Kumar A, Abd_Allah EF, Meddich A. Integration of Horse Manure Vermicompost Doses and Arbuscular Mycorrhizal Fungi to Improve Fruit Quality, and Soil Fertility in Tomato Field Facing Drought Stress. Plants. 2024; 13(11):1449. https://doi.org/10.3390/plants13111449
Chicago/Turabian StyleLahbouki, Soufiane, Abeer Hashem, Ajay Kumar, Elsayed Fathi Abd_Allah, and Abdelilah Meddich. 2024. "Integration of Horse Manure Vermicompost Doses and Arbuscular Mycorrhizal Fungi to Improve Fruit Quality, and Soil Fertility in Tomato Field Facing Drought Stress" Plants 13, no. 11: 1449. https://doi.org/10.3390/plants13111449
APA StyleLahbouki, S., Hashem, A., Kumar, A., Abd_Allah, E. F., & Meddich, A. (2024). Integration of Horse Manure Vermicompost Doses and Arbuscular Mycorrhizal Fungi to Improve Fruit Quality, and Soil Fertility in Tomato Field Facing Drought Stress. Plants, 13(11), 1449. https://doi.org/10.3390/plants13111449