1. Introduction
The apple is the primary fruit-bearing species in temperate regions and ranks first in terms of global fruit production. Several factors contributed to the spread of apples over the world, including: the nutritional, gustatory, therapeutic, and prophylactic value of fruits; the specific technological properties and agrobiological characteristics of trees; ecological plasticity; and high economic value [
1,
2]. Because of apples’ dietary and sanogenic properties as well as their high nutritional value and health benefits, the aphorism ‘An apple a day keeps the doctor away’ has become a truism [
3,
4]. Apples are a healthy food that contain a wide range of nutrients and non-nutrients. As a result, apples are among the most consumed fruits in many countries and cultures, with a per capita intake of roughly 20–30 kg/year [
5,
6,
7,
8]. By 2025 and 2050, it is anticipated that the average daily consumption of fruits will increase from 204 to 242 g per person worldwide [
9].
Apples are grown on an area of approximately 4 million ha worldwide, with fruit production averaging around 86–87 million metric tonnes per year in recent years [
10]. China is the leading producer of apples worldwide; in the 2019–2020 crop year, apple production amounted to around 41 million tons. Countries with a large apple production countries after China are the United States of America, Turkey, India, and the Russian Federation (these represent the top five countries). Other countries with a large production of apples also include Poland, Italy, France, Germany, Turkey, Argentina, Japan, and Chile. China’s apple production increased rapidly from 4.5 million metric tonnes in 1990 to 40.9 million metric tonnes in 2014, thereby exhibiting a truly remarkable growth in apple acreage and production [
10]. Interestingly, a single cultivar (Fuji) is grown on more than 70% of the apple-dedicated area in China [
11].
The cultivated apple influenced human history, and the apple is considered a symbol of wisdom and love [
12]. Apple varieties have a genetic background with a strong imprint of their ancestors
M. sieversii and
M. sylvestris as well as numerous interspecific and intraspecific hybridizations [
13]. Hybridization can broaden the genetic basis of cultivars by amplifying the heterozygosity of the offspring, but in general, apple breeding using hybridization was based on the principle of ‘crossing the best with the best parent’. Because the best varieties were also the most well-known and widespread in the world, relatively few parents were used in the new breeding works. Consequently, a small number of cultivars were repeatedly used as parents during the apple breeding process [
14,
15]. Thus, the most well-known varieties became the parental forms of most new varieties. Therefore, many of the modern varieties found in the world assortment are related to each other and have common parents or close origins or ancestors; i.e., a common genetic basis [
16]. Among the most well-known and widespread varieties in the world are Golden Delicious and Red Delicious as well as their mutants [
17]. McIntosh, Rome Beauty, Jonathan, Northern Spy, Stayman, York, Cox’s Orange Pippin, Belle de Boskoop, Reinette du Canada, Worcester Pearmain, and James Grieve are other ‘classic’ varieties known and widespread in the world. In recent decades, newer varieties have begun to spread in the culture, including Elstar, Gala, Jonagold, Mutsu, and Pink Lady (all with the Golden Delicious variety as their common parent) or others such as Empire and Fuji (both from Red Delicious), Idared, Granny Smith, Topaz, Pinova, Braeburn, Florina, Arlet, Rubin, Champion, Kanzi, and Jazz [
18]. Newly created varieties have given rise to the possibility of higher market pricing even if many traditional types continue to play a vital role in apple culture around the globe. To control the development and marketing of the new cultivars and raise prices for both cultivar owners and producers, ‘variety clubs’ were created. As a result, many fruit growers were limited in their ability to establish new orchards using certain new cultivars [
19,
20,
21,
22]. However, even for some more recent cultivars such as Crimson Snow, which has gained attention due to promotion and marketing methods as well as its great commercial look and organoleptic attributes, the ancestry is unknown. Based on SSR research, Crimson Snow, a putative descendant of Cripps Pink, was revealed to be a descendant of Delicious [
23].
As with any agricultural species, apple quality and production can vary. The apple can also be impacted by variations in consumer preferences, market demands, the processing industry, farmers’ obstacles and difficulties, and other constraints to sustainable apple production [
11,
24,
25,
26]. The appearance of new diseases or pests or an increase in their aggressiveness might have an impact on fruit yield and quality. Oscillations can occasionally be detected not just across longer time periods but also from one year to the next; for example, because of unfavorable weather during particular periods of the year, climatic events, or other unforeseen circumstances [
27,
28,
29]. In 2017, a single night of frost on April 19 decreased apple production in Europe by 24% [
30]. In China, severe spring frosts in the northern regions in 2020–2021 had a negative impact on fruit quality and quantity because the frost damaged blossoming trees [
31]. Additionally, COVID-19 caused logistical challenges, and the pandemic crisis intensified food insecurity (including the apple sector), causing seasonal labor shortages and issues with supply chains [
32]. Instead, it was projected that apple production would increase in some EU countries and Turkey, where relatively good growing conditions and the introduction of new varieties could result in a sixth consecutive year of rising yield [
33]. Apple breeding must be prospective and anticipatory in order to mitigate the hazards that could result from the many stresses (abiotic and biotic) or the market pressure and the continuously rising expectations of consumers and producers. Apple breeding goals must be both broad and specific in order to produce results that can be sustained in the long term [
34,
35,
36,
37].
Even though remarkable results have been obtained in apple breeding (as evidenced by the multitude of cultivars obtained in various countries around the world), the conservation of genetic resources and the creation of new cultivars are processes that have seen a noticeable slowdown or even stagnation in recent years. The global economic and social situation, the global trend of continuously reducing resources allocated for these purposes, and the risk of biodiversity loss and genetic resource restriction can severely impact the efforts to capitalize on genetic resources and counteract possible risk factors for the cultivated species [
38,
39]. The apple serves as a model species for cultivated plants (especially for perennial ones) due to its extensive use and socioeconomic significance. To highlight the complexity of apple breeding, a study of the effectiveness of selection in F
1 hybrid populations obtained from intraspecific hybridizations was conducted. Depending on the manner of parental form participation and the type of cross-breeding performed, the results of numerous hybridizations were compared. Additionally, it was investigated how the parental forms utilized for various hybrid combinations and the criteria employed in apple breeding could impact selection efficiency. The biological material in the field did not benefit from the maintenance activities because of the drastically reduced funding. Consequently, data were collected under these circumstances, in a natural context (i.e., long-term F
1 hybrid maintenance in the field, absence of tree cutting, fertilization, phytosanitary treatments, etc.).
2. Results
The average values of the trunk cross-sectional area (TCSA) and the height of the trees in the 25 hybrid combinations produced by direct crosses between parents with different growth vigor in the F
1 apple hybrids showed significant differences. A more accurate evaluation of the outcomes was accomplished by dividing the 25 hybrid combinations into three categories based on the vigor of the maternal parents (weak, medium, and strong vigor) (
Table 1).
The cross-sectional area of the trunk in F1 hybrids on average in the experiment was 54.9 cm2, and in hybrid families it ranged between 33.2 cm2 (Ardelean × Ancuța) and 71.2 cm2 (Ardelean × Prima). The difference between the mean values of the two families with the most extreme TCSA levels resulted in a large range of variation. The average of the hybrids from Ardelean × Prima exceeded twice those of Ardelean × Ancuța, which denoted very strong but completely different influences of Prima and Ancuța as fathers in the transmission of a certain vigor in offspring. The hybrid combinations that produced less vigorous seedlings were: Ancuța × Golden spur, Aromat de vară × Ancuța, Starkrimson × Ancuța, Reinette Baumann × Golden spur, and Roșu de Cluj × Ancuța. Depending on the vigor groups of the maternal parents, the lowest vigor of the F1 hybrids was obtained in the maternal parents with low growth vigor. The coefficients of variation (CV%) for TCSA had high values that oscillated between 15.7% (Kinrei × Jonathan) and 71.5% (Ancuța × Roșu de Cluj). When considering the CV% (small (0–10%), medium (10–20%), high (20–30%), and very high (over 30%)), only two combinations had medium variation, four had high, and the rest had very high.
The height of the hybrids showed a large amplitude, and there were noticeable differences amongst the examined combinations, thereby demonstrating the strong parental influences on the trait. The average height of the trees for all hybrid combinations was 3.8 m; the extreme values were found in the Ardelean × Ancuța (3.0 m) and Ardelean × Feleac (5.1 m) combinations. Elite plants with a smaller tree height, in addition to other desired traits, were more likely to occur in the following hybrid populations: Ardelean × Ancuța, Ancuța × Golden spur, Roșu de Cluj × Kaltherer Böhmer, Roșu de Cluj × Golden spur, Kinrei × Jonathan, Roșu de Cluj × Ancuța, Ancuța × Mutsu, Aromat de vară × Ancuța, and Prima × Feleac.
The vigor of the F
1 hybrids from the five crosses with a common tester revealed obvious differences depending on half-sib families (
Table A1) due to the maternal parents because the paternal parent was the same in each combined testcross. Among the HRS selections and varieties used in the hybridization with Feleac as a tester, DSF 7/68 and DSF 3/80 had the highest combining ability for tree vigor assessed as TCSA (the latter for both TCSA and hybrid height). According to current breeding goals, hybrid combinations with the maternal parents X-5-71, X-17-19, and X-3-8 are promising for the selection of new elite plants with low growth vigor. A contrasting situation was found in the Prima cultivar used as the maternal parent. Its progenies were distinguished by a robust appearance and trees with thick trunks but a lower height than the hybrids of all other combinations. TCSA as the mean of hybrid combinations with a Mutsu tester was much lower than with a Feleac tester. The reduced vigor transmitted from Mutsu to their offspring was also confirmed for the F
1 hybrids’ heights. The testcross with the lowest tester vigor (Starkrimson) was found to have the highest mean TCSA value.
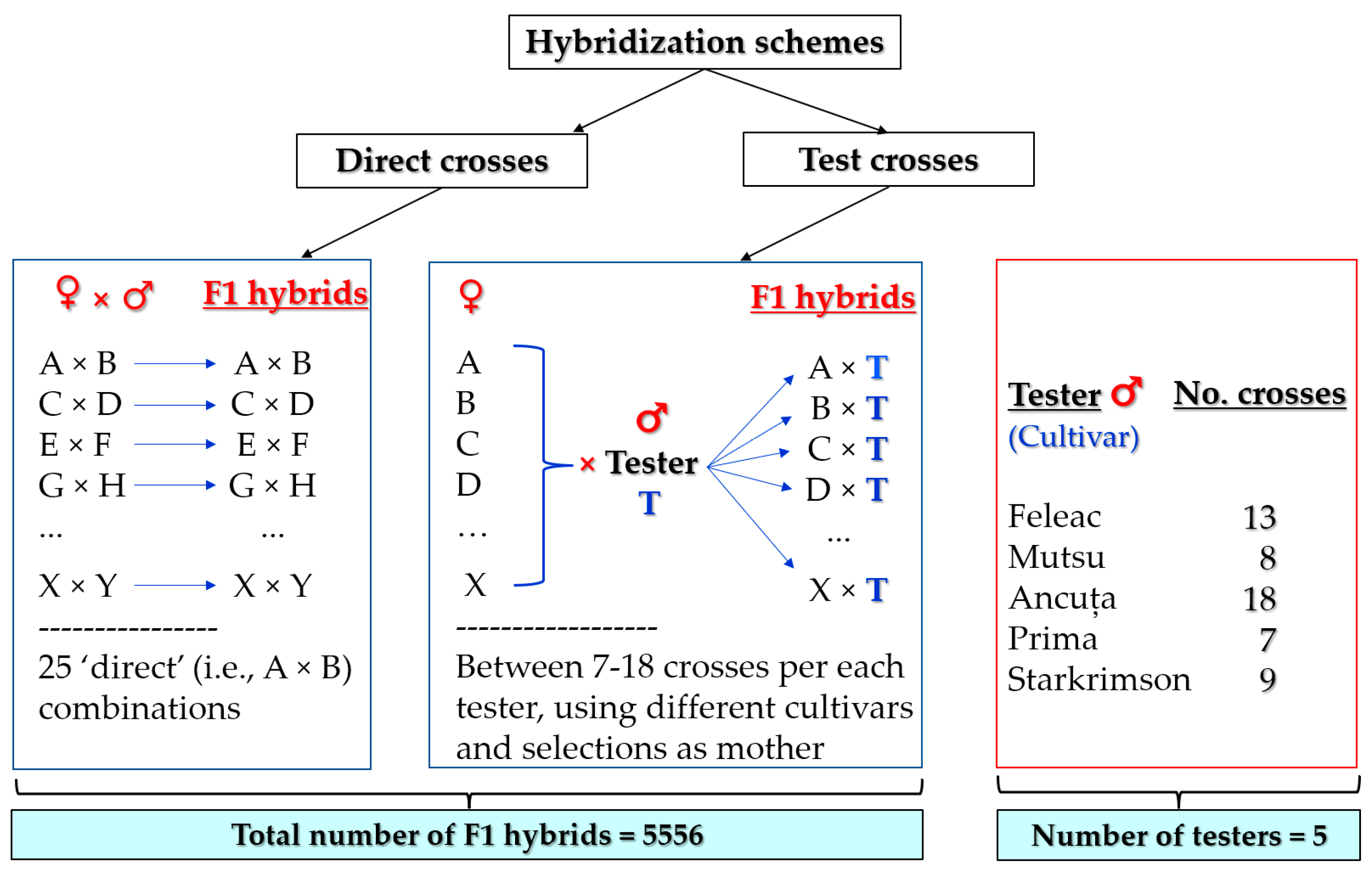
For tree vigor, a total of 5556 F
1 hybrids originated from direct and testcross hybridizations with parents of different levels of vigor were arbitrarily classified into three classes (
Table 2). Overall, the following proportions were obtained for the TCSA: 16.8% weak, 64.7% medium, and 18.6% strong. By dividing the tree height of the F
1 hybrids into weak, medium, and strong, the proportions obtained in the three categories were 15.8%, 53.1%, and 31.1%.
When the parents were divided into three vigor categories to evaluate the combining ability for all crosses, it was discovered that genetic effects of the general combining ability (GCA) were generally more significant than those of the specific combining ability (SCA) both for TCSA and tree height (
Table 3). In accordance with the stated breeding objectives, it is important to create new cultivars with low vigor, and it was confirmed that offspring with low vigor were more frequently produced from parents with low growth vigor. Thus, the parents with weak vigor had the highest positive GCA value for TCSA and tree height. According to the vigor of the parents and how they were combined, additive gene effects played a significant role in the transmission and fixation of vigor in the F
1 hybrids, albeit in a variety of manners (i.e., in some combinations such as strong × weak vigor parents, negative GCA values were recorded).
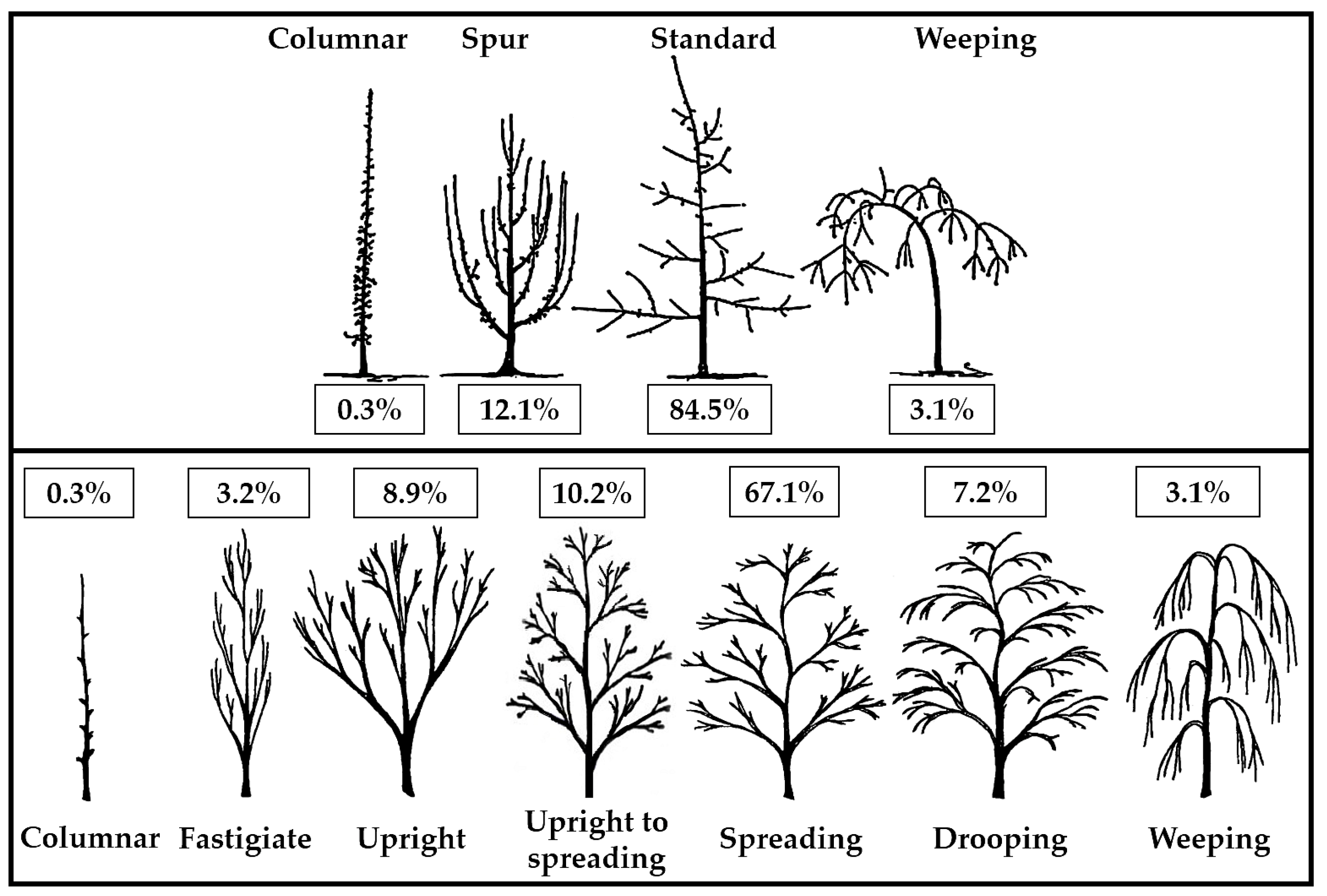
The growth and ramification of the trees in some hybrids were rather simply framed in one of the four architectural ‘ideotypes’ established by Lespinasse et al. [
40].
Figure 1 depicts the obvious predominance of plants with normal growth and fruiting (84.52%). The spur type accounted for 12.14% of the total, 3.07% of the weeping type, and only 0.27% of the compact-columnar type. The spur proportion was divided between the fastigiate (3.2%) and upright (8.9%) ideotypes and the standard between the upright to spreading (10.2%), spreading (67.1%), and drooping (7.2%) ideotypes using the UPOV classification for edible [
41] and ornamental cultivars [
42].
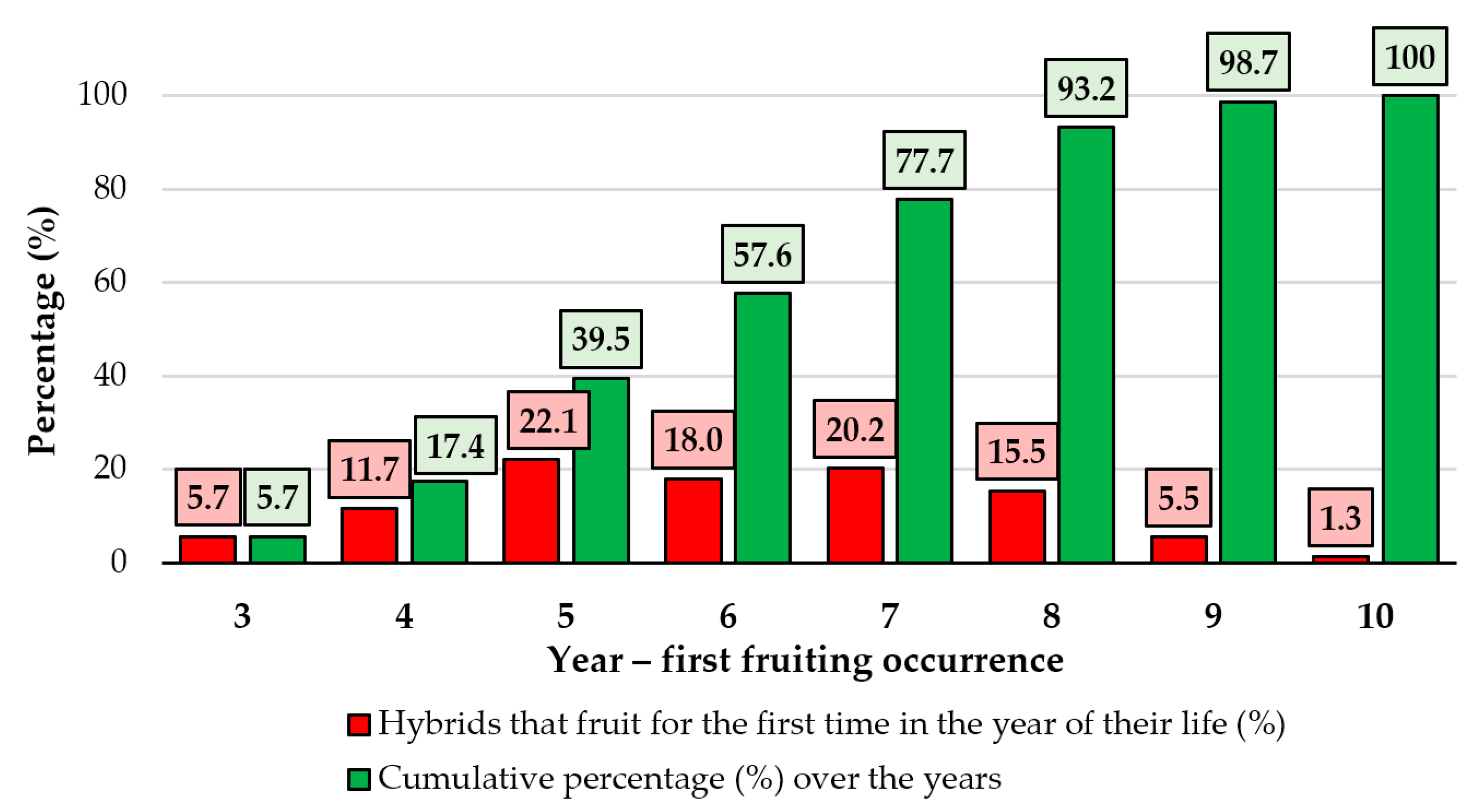
In the first two years of their lives, none of the hybrids attained the fruiting period. A relatively small percentage of seedlings (5.7%) produced their first fruit in the third year (
Figure 2).
The percentage of hybrids that produced fruit increased gradually in consecutive years and reached 11.7% in the fourth year, 22.1% in the fifth, a slightly lower percentage (18.0%) in the sixth, 20.2% in the seventh, 15.5% in the eighth, 5.5% in the ninth, and 1.3% in the tenth. The relationship between the cumulative percentage of hybrids and the year of fruiting indicated the length of the juvenile stage in apple hybrids (seedlings on their own roots). It should be noted that only hybrids that began bearing fruit within the first 10 years of life were examined. However, some of the hybrids analyzed did not bear fruit at the end of the study period (probably due to an extremely long juvenile period or due to sterility). In the direct crosses, hybrids with a high number of fruits per tree were recorded in the following combinations: Reinette Baumann × Golden spur, Cluj Red × Kaltherer Böhmer, Roșu de Cluj × Ancuța, Aromat de vară × Melba, and Golden Delicious × Ancuța (
Table 4).
These hybrid populations have the potential for successful selection of elite plants with profuse fruiting, but this trait must also be coupled to the size of the fruits and other advantageous qualities. The coefficient of variation per experiment (70.9%) demonstrated that the number of fruits per tree in F1 apple hybrids was an extremely variable trait. Within the families, the CV% was very high and had limits between 31.5% (Roșu de Cluj × Ancuța) and 92.6% (Kinrei × Jonathan).
Hybrids with large fruits were obtained in the following families: Aromat de vară × Mutsu, Roșu de Cluj × Kaltherer Böhmer, Ardelean × Prima, Aromat de vară × Reinette Baumann, Aromat de vară × Melba, Kinrei × Ardelean, and Golden Delicious × Ancuța. The coefficient of variation (CV%) fluctuated strongly within the hybrid families between 16.2 and 69.3%.
The number of fruits on the tree in the testcrosses (
Table A2) varied widely, and the coefficient of variation was very high (81.3%). In the case of Mutsu and Prima as testers, the F value did not reveal significant differences. Therefore, it was concluded that the number of fruits per tree in the two half-sibling families was not due to true differences between the maternal parents but to errors. The Ancuta tester highlighted DSF 7/68, X–6–3, Roșu de Cluj, X–17–16, and Golden Delicious mothers, all of which had a good combining ability for the number of fruits per plant. They can provide biological material conducive to effective selection in the direction of choosing individuals with good prospects in the breeding process and at the same time can be used as suitable parents for high productivity in new hybridization programs. The variation limits of the average scores for the number of fruits per hybrid when Starkrimson was used as the paternal tester oscillated between 1.14 and 7.75, thereby indicating the significant differences between the maternal parents for their general combining ability. It resulted in a high CV% value (74.6%) for all half-sibling families with Starkrimson as the tester.
For the average number of days required for fruit ripening of F
1 hybrids from families resulting from direct hybridizations, a relatively large range of variation was recorded (
Table 5). The amplitude was between 126.7 days (Mutsu × Roşu de Cluj) and 186.0 days (Ancuța × Golden spur). The minimum value was recorded in offspring from two winter varieties since many hybrids had fruit ripening toward the end of August to the beginning of September. The high standard deviation may indicate that the early ripening of the fruits in this cross probably had a subjective causality determined by the triploidy of the maternal parent. The mean of all hybrid families produced by crossing winter-ripening parents (160.6 days) illustrated that within them, an effective selection could be found to obtain some winter varieties. The hybrid combinations that can provide a useful biological material for the selection of individuals with a very early ripening of the fruits proved to be Ardelean × Clar alb and Prima × Ardelean. The coefficient of variation for the number of days required for fruit ripening in the F
1 apple hybrids had a relatively low value compared to other characteristics (i.e., the cross-sectional area of the trunk).
Fruit quality was highlighted in 21 hybrid combinations; some had a significant proportion of offspring with an appropriate taste for the selection. As a result, the hybrid combinations of Prima × Feleac, Ardelean × Feleac, and Prima × Ardelean stood out. Mutsu × Roșu de Cluj, Ardelean × Prima, Roșu de Cluj × Feleac, Golden Delicious × Ancuţa, Roșu de Cluj × Ancuţa, and Kinrei × Jonathan represented other hybrid families with good-tasting fruits that may be of interest for selection.
The coefficient of variation for fruit taste in the analyzed families was relatively high, thereby indicating a large diversity of fruit taste in each family. These values suggested the presence of a wide range of genotypes that ranged from low-quality- to high-quality-tasting fruit hybrids. Some hybrids produced fruits with taste comparable to the cultivars in the assortment.
In hybridizations with the five testers, the number of days required by F
1 hybrids to reach physiological fruit maturity varied significantly (
Table A3) both among families that had a common tester and within each hybrid family.
The testcross hybridization of 13 apple varieties and selections with the Feleac variety used as paternal tester highlighted the late maturing hybrids from the combination III-VI-5-26 × Feleac, which had an average maturation period of 201.4 days. A late offspring (174.0 mean days of ripening) was produced when a summer variety (Aromat de vară) and a winter variety (Mutsu as tester) were crossed. It was interesting that by crossing with a winter selection (X–6–64), offspring with an early fruit ripening (110.0 days) were obtained. The hybrids with the latest ripening when Ancuța was used as a tester resulted from the crossing with the DSF 3/80 selection (200.8 days), and a large part of the descendants completed the fruit ripening in storage. As a paternal tester, Prima, which is an autumn variety, induced a relatively late fruit ripening in the progeny (average of 154.7 days of experiment); even though the seven genotypes used as maternal parents contained one (X–13–10) with summer ripening and three (X–21–20, Ardelean and X–5–52) with autumn ripening. Using Starkrimson as a paternal tester (winter variety), an average of 174.3 days was achieved, which was higher than the averages produced by the other testers (Feleac, Mutsu, Ancuța, and Prima).
When Feleac was used as paternal tester, the differences between the mean scores for the fruit taste of the hybrids in the 13 families were not statistically significant (
Table A3). Hybrids with pleasant-tasting fruits were obtained in crosses with the Prima, Ardelean, Roșu de Cluj, X–17–19, III–VI–5–26, DSF 3/86, DSF 7/68, and X–9–70 varieties. The CV% of the taste of the fruits in the ensemble families with Feleac as a tester was 33.1%, and within the hybrid families it was between 21.0–70.7%. Ancuta as a tester revealed a good general combining ability for progenies with good fruit taste for the following genotypes: X–9–69, DSF 7/68, DSF 3/41, X–6–3, DSF 5/22, X–9–70, Golden Delicious, Roșu de Cluj, and DSF 5/67. When using the Prima variety as a tester, the possibility of identifying hybrids with very-good-tasting fruits was high in combinations with X–13–10 and X–5–65. Instead, as for fruit size, the worst results for fruit taste were obtained when crossing with the X–21–20 selection. As a tester, Starkrimson identified DSF 3/58, 218/2, DSF 5/45, and X–6–3 as good parents for fruit quality. In Starkrimson half-siblings, it was observed that the higher the average score in a hybrid family (indicating a high proportion of individuals with good-tasting fruit), the lower the variation in the trait, thereby indicating the relative uniformity of individuals with the trait.
In the ensemble of analyzed hybrids with edifying results for fruit development and ripening both from direct hybridizations and testcrosses (5484 hybrids), the following proportions were obtained for the ripening period of the fruits: summer 12.7%, autumn 53.5%, and winter 33.8% (
Table 6).
The important genetic additive effects as well as the non-additive genetic effects in the transmission, fixation, and phenotypic manifestation of fruit ripening in F
1 apple hybrids were highlighted by combining all hybridizations in a diallel system with three groups of parents according to the season of fruit ripening (summer, autumn, and winter) (
Table 7). The results showed that if parents with a certain fruit maturity were used, the additivity effects contributed significantly to obtain offspring with the same maturity. The general combining ability (GCA) value for the summer cultivars was significantly positive (+1.7457 ***), which indicated that they were the most suitable parents for obtaining offspring in which those with a summer ripening of fruits predominated. The same situation was recorded in the autumn and winter parents. Instead, due to significantly negative GCA values, the additivity of polygenes acted against producing hybrids with summer fruit ripening when the parents were represented by autumn and winter varieties.
Interaction effects of dominance and epistasis were significantly positive in crosses with parental forms of the summer × summer type (CSC = +1.1043 **). Contrary, in summer × autumn and summer × winter hybridizations, the non-additive effects significantly prevented the obtaining of offspring with a summer fruit ripening. Thus, significant additive and non-additive effects were involved in the genetic inheritance of the summer ripening of fruits (the CSC constancy had high positive values in maternal parents with summer ripening of fruits). Therefore, summer fruit ripening was determined either by the combined effect of additivity and dominance and epistasis interactions or solely by the no-additive interactions. These latter effects, although partially non-transmissible, could decisively influence the trait in F1 hybrids, especially in summer × summer crosses.
Apple scab (
Venturia inaequalis (Cke.) Wint.) attack on F
1 apple hybrids from 22 direct hybrid combinations highlighted relatively low values of the average scores recorded for most hybrid combinations (
Table 8). However, significant differences between the tested families were identified. The most sensitive hybrids to the apple scab attack were those from the combinations Mutsu × Roşu de Cluj and Ancuţa × Starkrimson. Instead, the hybrids from the crosses Ancuţa × Mutsu, Ancuţa × Roşu de Cluj, and Prima × Feleac showed a proper response to the apple scab attack. Descendants with a certain tolerance to the disease were also obtained in the following combinations: Aromat de vară × Reinette Baumann, Prima × Ardelean, Ardelean × Feleac, Roşu de Cluj × Feleac, Aromat de vară × Mutsu, and Reinette Baumann × Golden spur. Obviously, the chances of selection for the identification of promising elites were increased in hybrid populations that stood out due to resistant or tolerant trees to apple scab attack.
The hybrids of Aromat de vară × Mutsu, Aromat de vară × Ancuta, and Ancuta × Starkrimson provided an appropriate response to the powdery mildew (Podosphaera leucotricha (Ell. et Ev.) Salm.) attack. Within the hybrid families, the coefficient of variation of the notes for disease response ranged from small-medium (CV% of 10.2% in Roşu de Cluj × Ancuța) to very high (CV% of 52.8% in Ardelean × Prima). The CV% of powdery mildew noted in the experiment represented by the 22 hybrid combinations was 38.1%.
The average notes between half-sibling families based on the tester did not differ significantly for the apple scab attack in the testcrosses, but there were significant differences in each of the five types of testcrosses (
Table A4). Each tester highlighted genotypes with a higher general combining ability to produce F
1 hybrids with a suitable response to apple scab. Thus, in the hybridizations with Feleac, the following stood out: X–17–19, Prima, X–9–19, and Roşu de Cluj (1.35). With Mutsu as a tester, less sensitive hybrids to scab fungus were recorded when the mothers were Ancuța, Aromat de vară, and X–5–71, while the Ancuța tester highlighted the X–17–16 and X–21–20 selections. Prima as the tester highlighted X–5–65 and X–21–20 selections, and Starkrimson two other selections: III–II–17–25 and DSF 1/54.
The testcross schemes were also useful in identifying hybrid families in which the selection for resistance or tolerance to powdery mildew could be more efficient. Feleac as a tester indicated the hybrid populations in which the mothers were X–5–71, X–3–8, X–9–70, DSF 7/68, and III–VI–5–26. Mutsu highlighted Aromat de vară, 218/2, X–6–73, and DSF 3/70; Ancuța as the tester highlighted DSF 5/67, X–17–16, and X–9–70. Prima identified the following genotypes: X–5–65, X–21–20, X–6–24, and X–5–52; while Starkrimson as the tester highlighted 218/2, DSF 3/58, and X–6–3.
The main genetic parameters analyzed within half-sibling families depending on testcrosses are presented in
Table 9. There were clear differences between the mean values obtained in the F
1 hybrids in the half-sibling families depending on the trait and tester used. The genotypic coefficient of variation (GCV) registered large oscillations depending on the tester for the following traits: fruit size—between 2.6 (Feleac) and 41.8 (Prima); fruit taste—between 8.1 (Mutsu) and 49.8 (Prima); and response to powdery mildew attack—between 9.8 (Mutsu) and 41.0 (Prima). For the response of the F
1 hybrids to apple scab attack, because the Prima cultivar has a monogenic resistance to disease (
Vf gene; i.e.,
Rvi6), the GCV value is only indicative and presented in parentheses. For the same reason, heritability coefficients were also not calculated.
There were differences between heritability in the broad sense (H2) and heritability in the narrow sense (h2) depending on the trait and the tester. For H2, a greater variation in the genotype’s contribution to the trait’s phenotypic expression was observed as follows: fruit size—between 0.531–0.946 (Feleac and Prima); taste of the fruits—between 0.566–0.975, and response to powdery mildew attack—between 0.554–0.891 (Mutsu and Prima). Greater uniformity of H2 values among testers was identified for fruit ripening and tree vigor (TCSA and tree height).
The genetic effects of additivity played a different role depending on both the tester and the trait analyzed. Thus, the non-additive genetic effects of dominance and epistasis had a more consistent role for TCSA with Starkrimson (h2 = 0.408), for tree height with Mutsu (0.416), for fruit size and fruit taste with Prima (0.384 and 0.410, respectively), for fruit ripening with Starkrimson (0.427), and for apple scab and powdery mildew attack with Starkrimson (0.341 and 0.376, respectively). Expected response to selection had relatively high values with Starkrimson for TCSA and the number of fruits per tree and with Prima for fruit size, fruit taste, and response to powdery mildew attack.
The percentage of F
1 apple hybrids that presented the analyzed traits expressed at an optimal level for selection differed strongly depending on the character of interest for apple breeding as well as on the type of hybridization in which the estimation was performed (
Table 10).
A relatively low percentage (4.6%) was obtained for fruit flavor in direct hybridizations, but an unexpectedly high percentage for the trees’ tolerance to apple scab attack was obtained. In the case of the testers, the percentage of F1 hybrids with very good fruit taste was also very low, especially with Starkrimson (1.4%) but also with Ancuța and Prima and slightly higher with Feleac (9.2%). Overall, a higher proportion of hybrids appropriate for selection were obtained under the hybridization categories for winter ripe fruits, apple scab tolerance, and powdery mildew. Among the testers noted were Ancuța and Feleac (for the ‘large fruit’ trait), Starkrimson and Prima (for tolerance to apple scab), and Mutsu (for weak vigor of the trees).
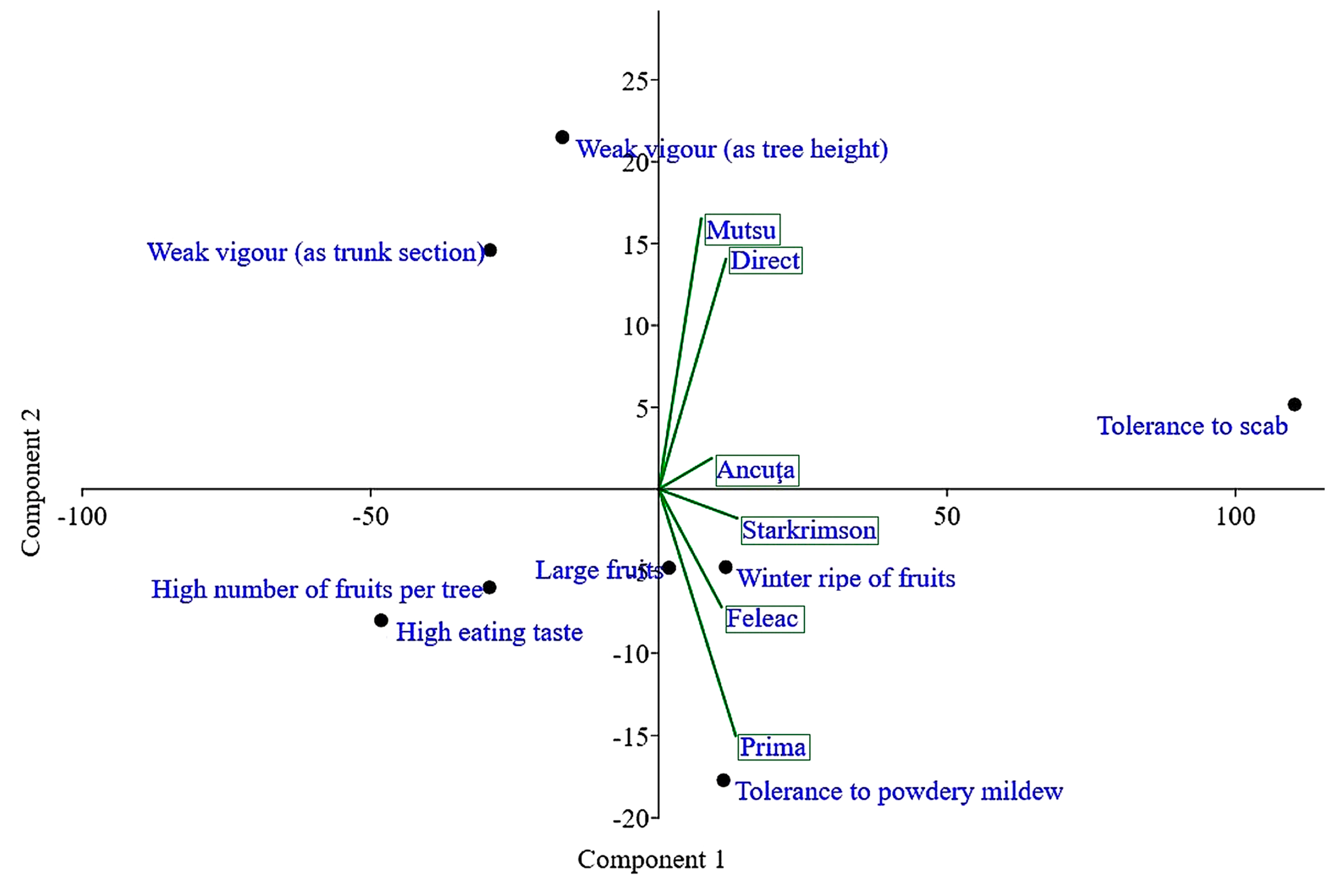
A principal component analysis (PCA), a multivariate technique for analyzing quantitative data, highlighted a close link between hybridizations when utilizing Mutsu as the tester and direct hybridizations (
Figure 3). Ancuța was also positioned in the same quadrant (quadrant I, top right) but at a certain distance. In addition, the other testers were on the right side of the vertical axis of the PCA but in quadrant II (lower right). The Prima cultivar was found in the area that was the farthest from the other places (particularly in relation to Mutsu).
Tolerance to apple scab was the only trait that was placed in a different quadrant from the others. This trait was the most different from the others (quadrant I). In opposition to the adequate response of trees to apple scab attack, the main characteristics of fruit production and fruit quality appeared, namely the number of fruits per tree, large fruits, and high eating taste. On the other diagonal, in quadrant IV there was reduced tree vigor (trunk section area and tree height, both elements of tree growth that were located close) in an inverse relationship with winter ripeness of fruits and tree tolerance to powdery mildew attack. Of the two main components of the PCA, PC1 captured a very large part of the total variation (90.5%) but PC2 only a small part (6.2%).
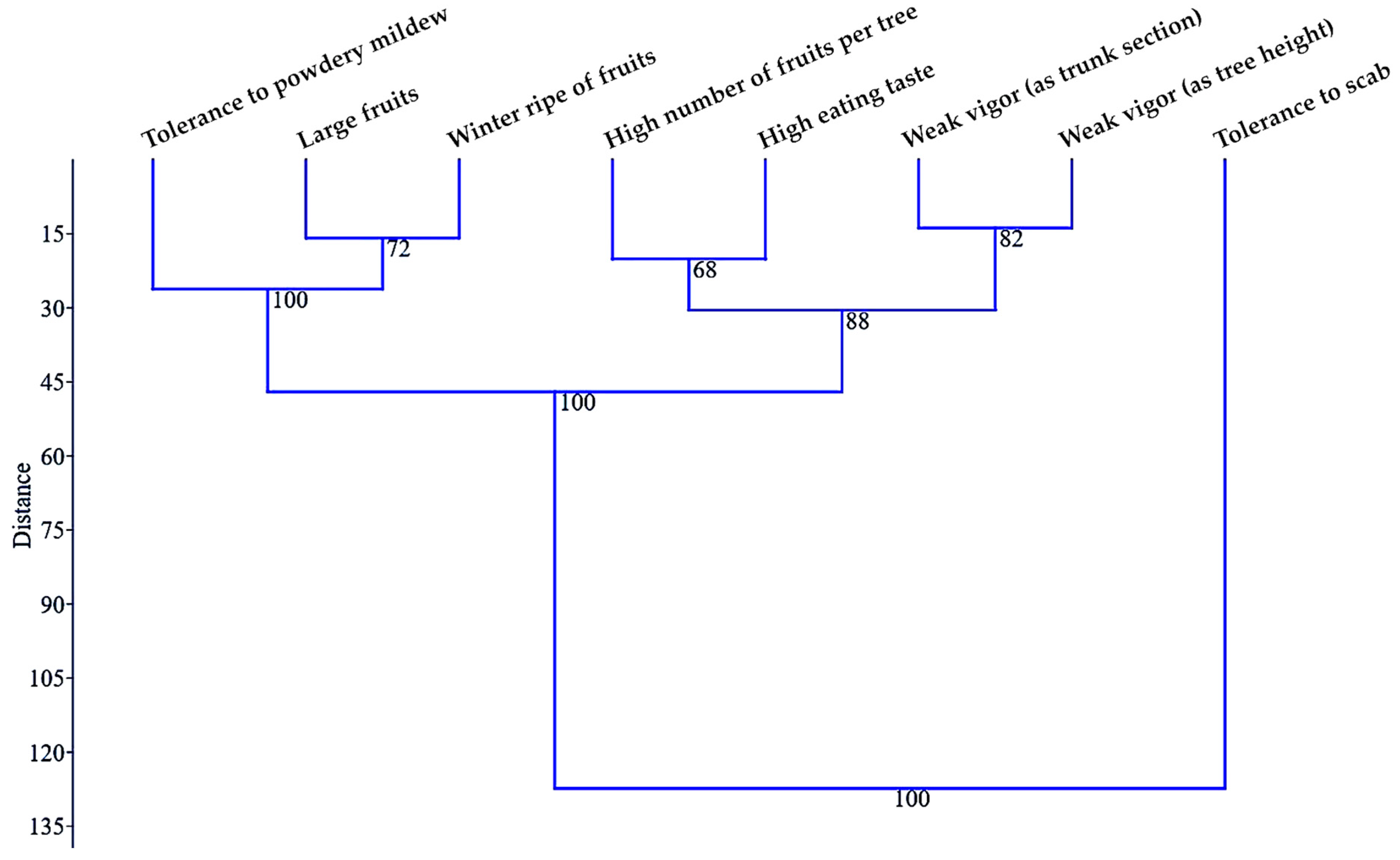
A UPGMA dendrogram (
Figure 4) confirmed the distance of tolerance to apple scab attack from the rest of the analyzed characters, which was highlighted previously by PCA. The tolerance to apple scab formed as a distinct cluster a singular character that was different from the other cluster, on which there were two subclusters: one with three characteristics and the others with four characteristics grouped by two.
If the arrangement of the vigor elements of the trees together in a common subcluster was to be expected, the positioning of a high number of fruits per tree and high eating taste in a common subcluster appeared quite surprising. The close relationship revealed by the multivariate analysis between these two traits may suggest that hybrids with a larger fruit load in the crown of the trees may produce fruits with a better taste. Similarly, another cluster with two close characters could indicate that the winter ripening of fruits can be associated with larger fruits. These two characteristics would be quite closely related to tolerance to powdery mildew attack.
Extreme variation existed in the minimum of progeny required to produce a hybrid with quantitative traits of agronomic interest (
Table 11). Regardless, this computed amount (if these traits behave independently) appeared extremely high depending on the type of hybridization (direct vs. testcross), the tester, and the cumulative traits of interest.
The average for the entire experiment was at least 122 offspring for the two traits that were crucial in the selection of elite plants (high eating taste and large fruits) and increased to 692 progenies if two more traits of great interest in apple breeding were added (tolerance to apple scab and powdery mildew attacks). If we added a fifth trait (high number of fruits per tree), we reached 6383 hybrids, and then the exponential growth continued to impressive numbers with the addition of the next desired traits.
Thus, if we combined the intended fruit ripening period (a breeding goal in HRS was late ripening to produce ‘winter’ cultivars) with the lowered tree vigor (another goal was to produce cultivars suitable for crop intensification), we obtained 259,509 hybrids. Interestingly, if instead of weak vigor as trunk cross-sectional area we included weak vigor as tree height, the minimum number of hybrids was reduced to 166,835. Analysis indicated that among the testers, Feleac and Prima produced the best results for the first five extremely relevant traits (high eating taste, large fruits, tolerances to apple scab and powdery mildew attack, and high number of fruits per tree).





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}





