
Smart Farming Enhances Bioactive Compounds Content of Panax ginseng on Moderating Scopolamine-Induced Memory Deficits and Neuroinflammation

Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Cultivation and Extraction
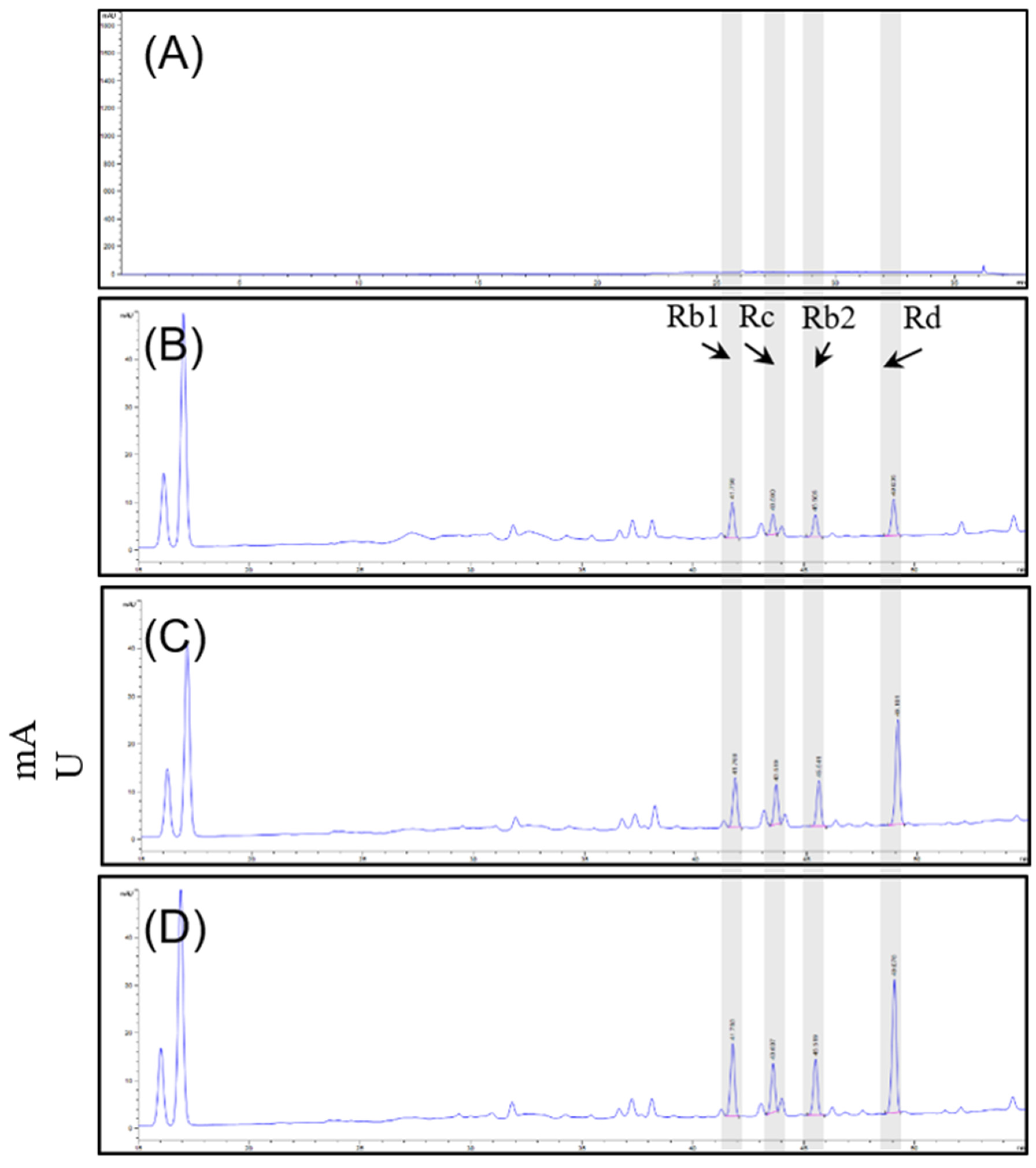
2.2. High-Performance Liquid Chromatography (HPLC) Analysis
2.3. Cell Culture
2.4. Animals and Drug Administration
2.5. Measurement of Nitric Oxide
2.6. Total Protein Isolation from BV-2 Cells and Brain
2.7. Western Blot Analysis
2.8. Novel-Object Recognition Test (NORT)
2.9. Y-Maze Test
2.10. Passive Avoidance Test (PAT)
2.11. Statistics
3. Results
3.1. Compositional Analysis of Ginsenosides of P. ginseng Extract
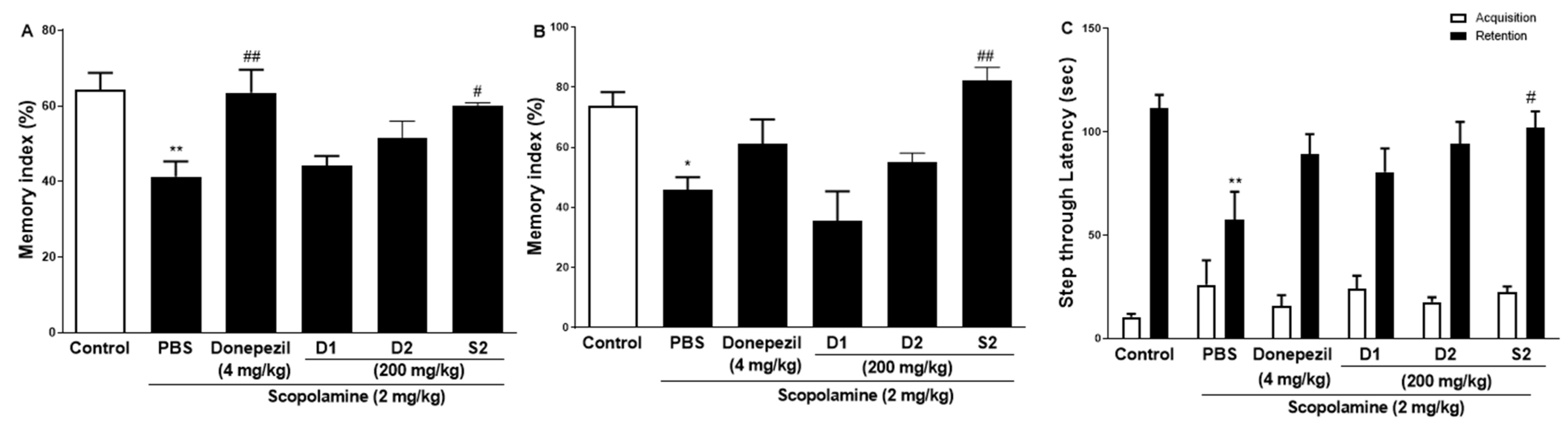
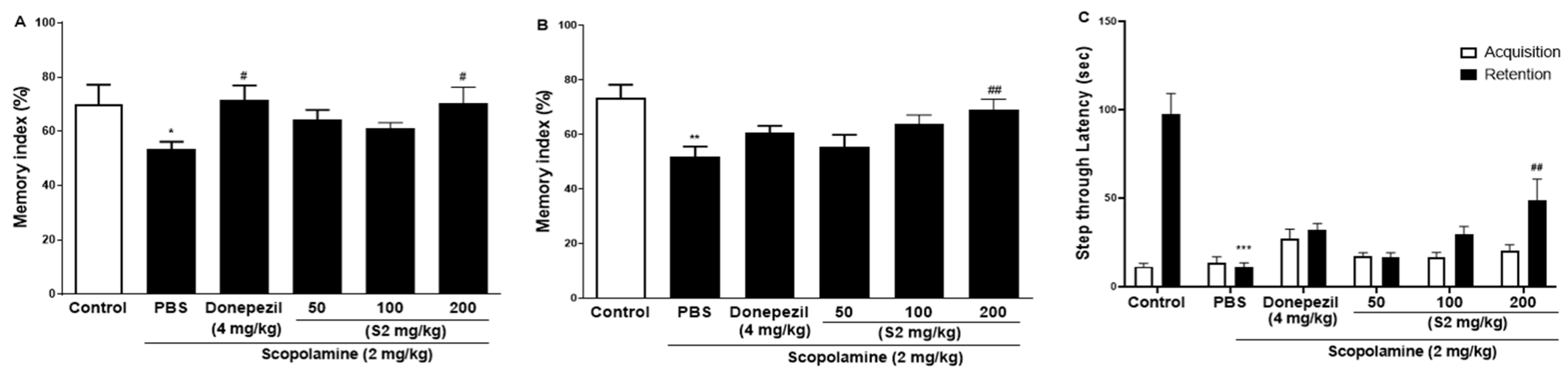
3.2. S2 Extract Enhanced Learning Memory and Cognitive Function in Scopolamine-Induced Memory Impairment
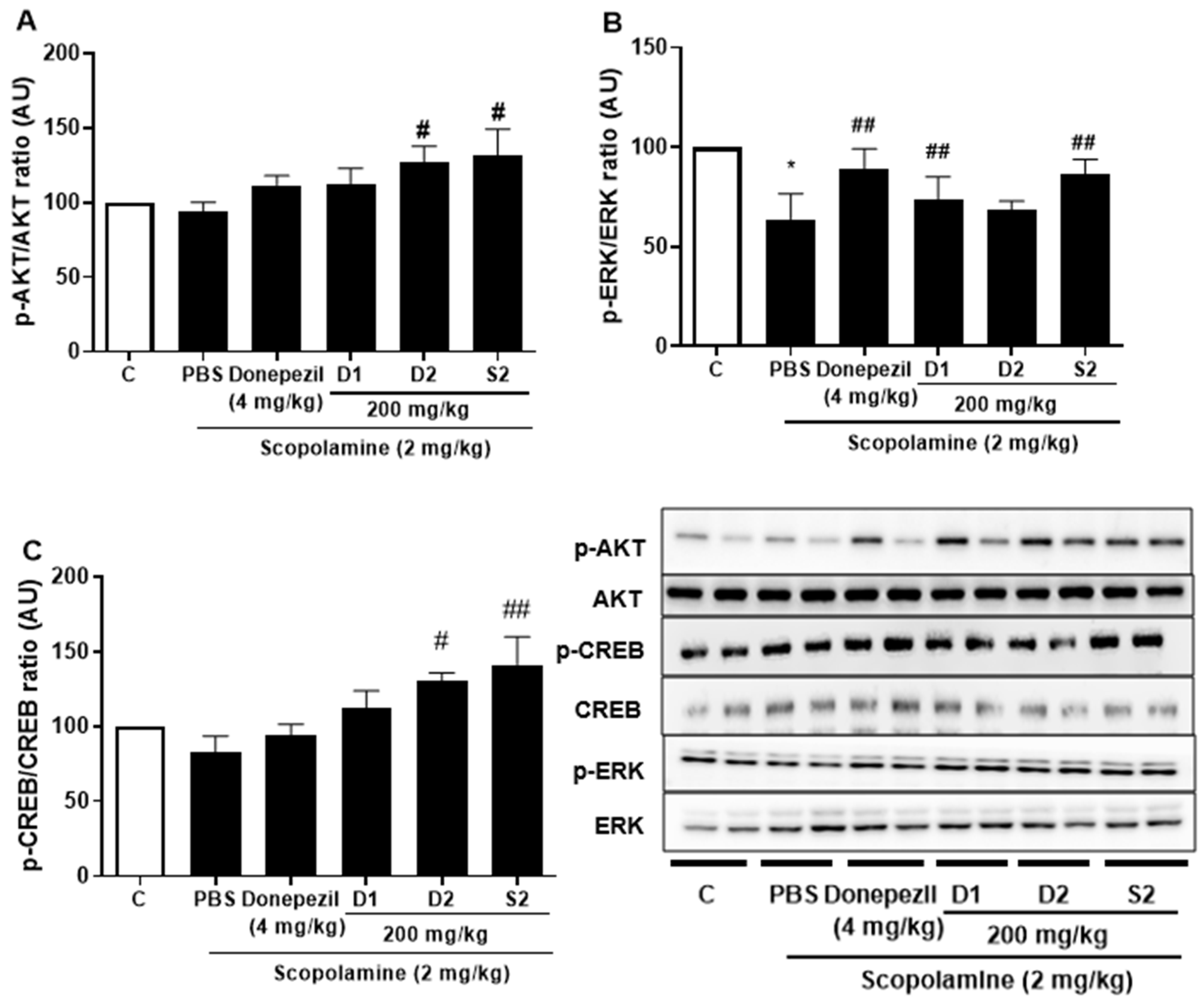
3.3. S2 Extract Reversed Scopolamine-Induced Decreases in p-CREB and AKT/ERK Signaling
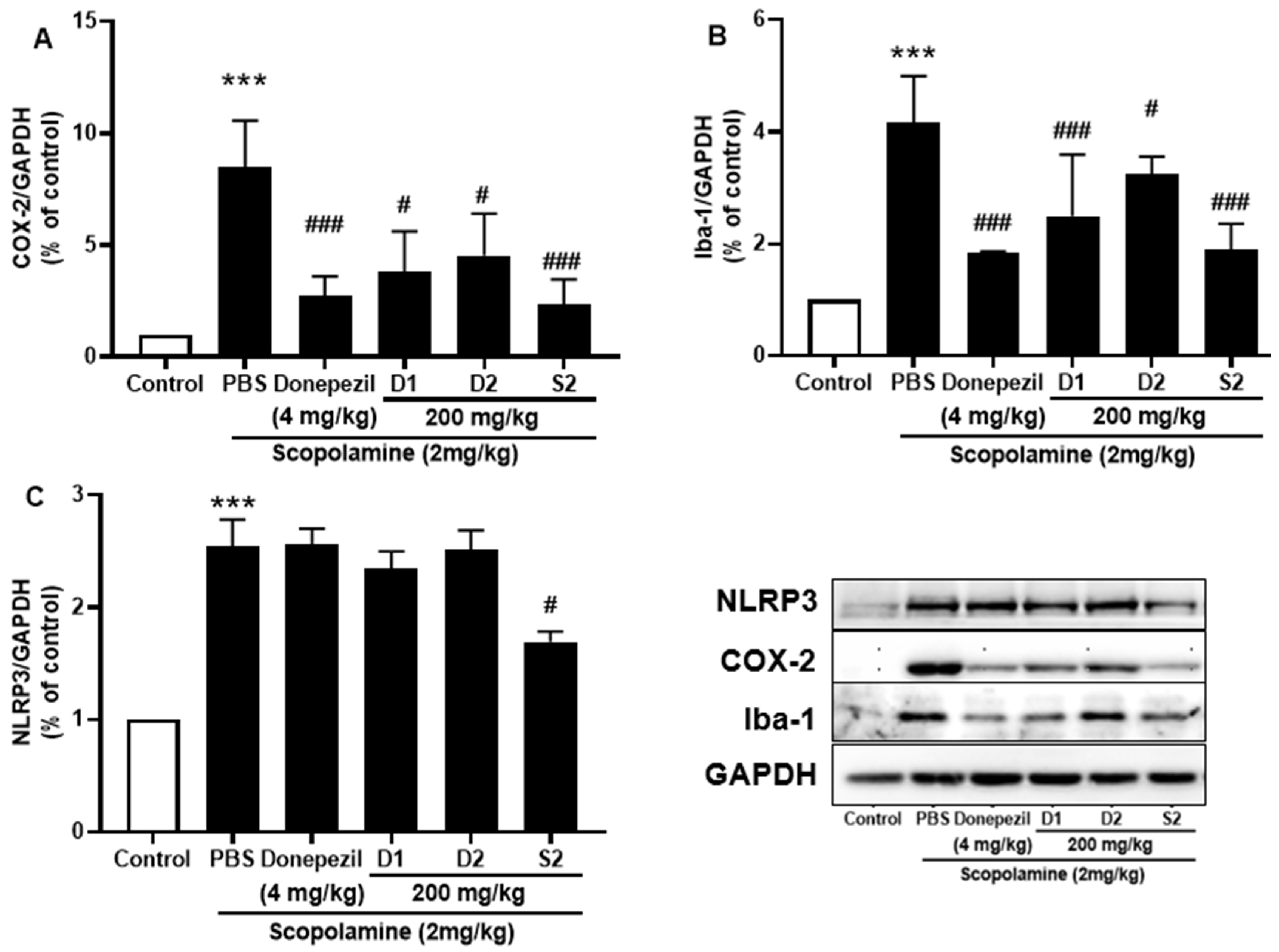
3.4. S2 Extract Attenuated Glial Activation and COX-2 Mediated Neuroinflammation in Scopolamine-Administered Mice
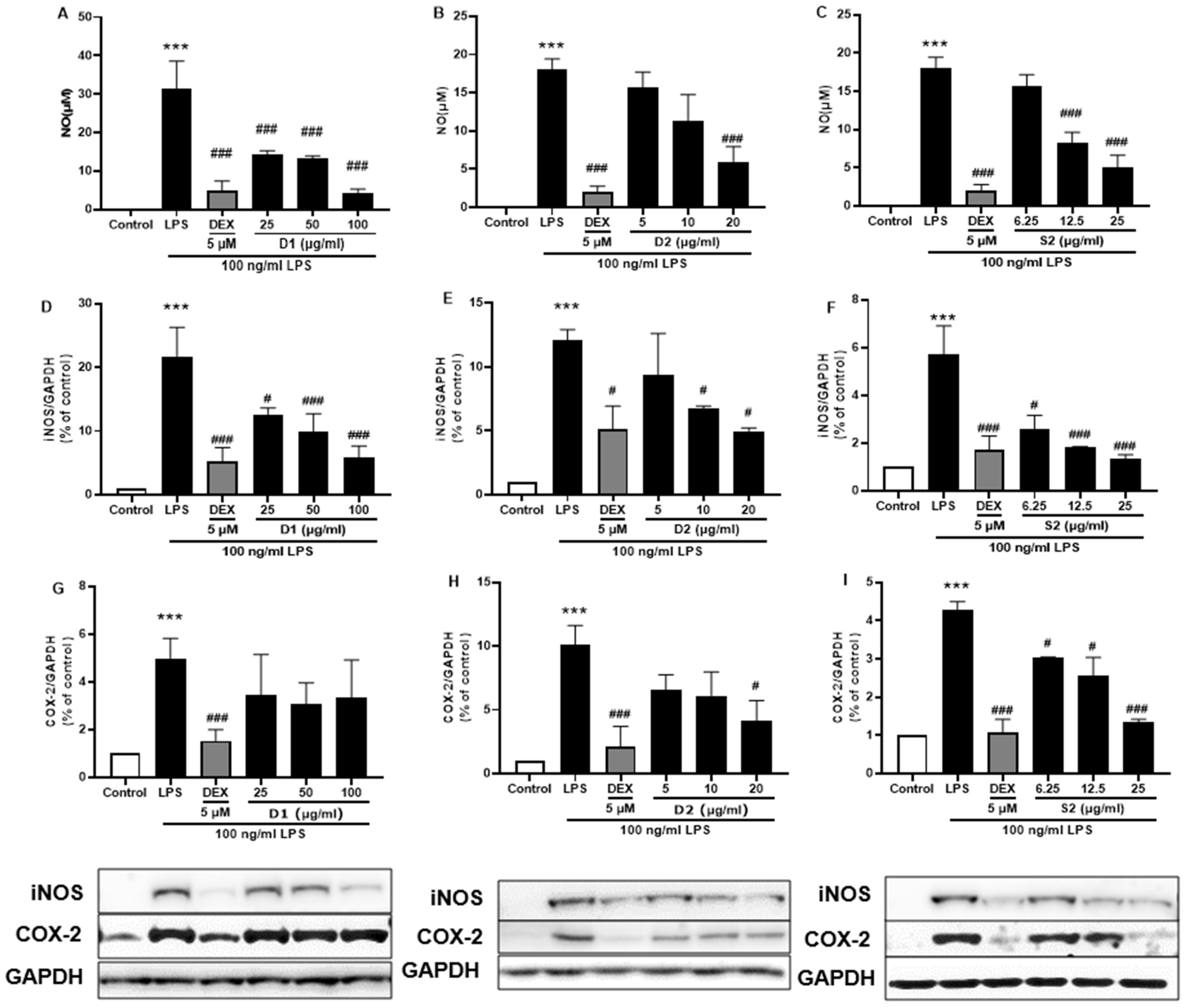
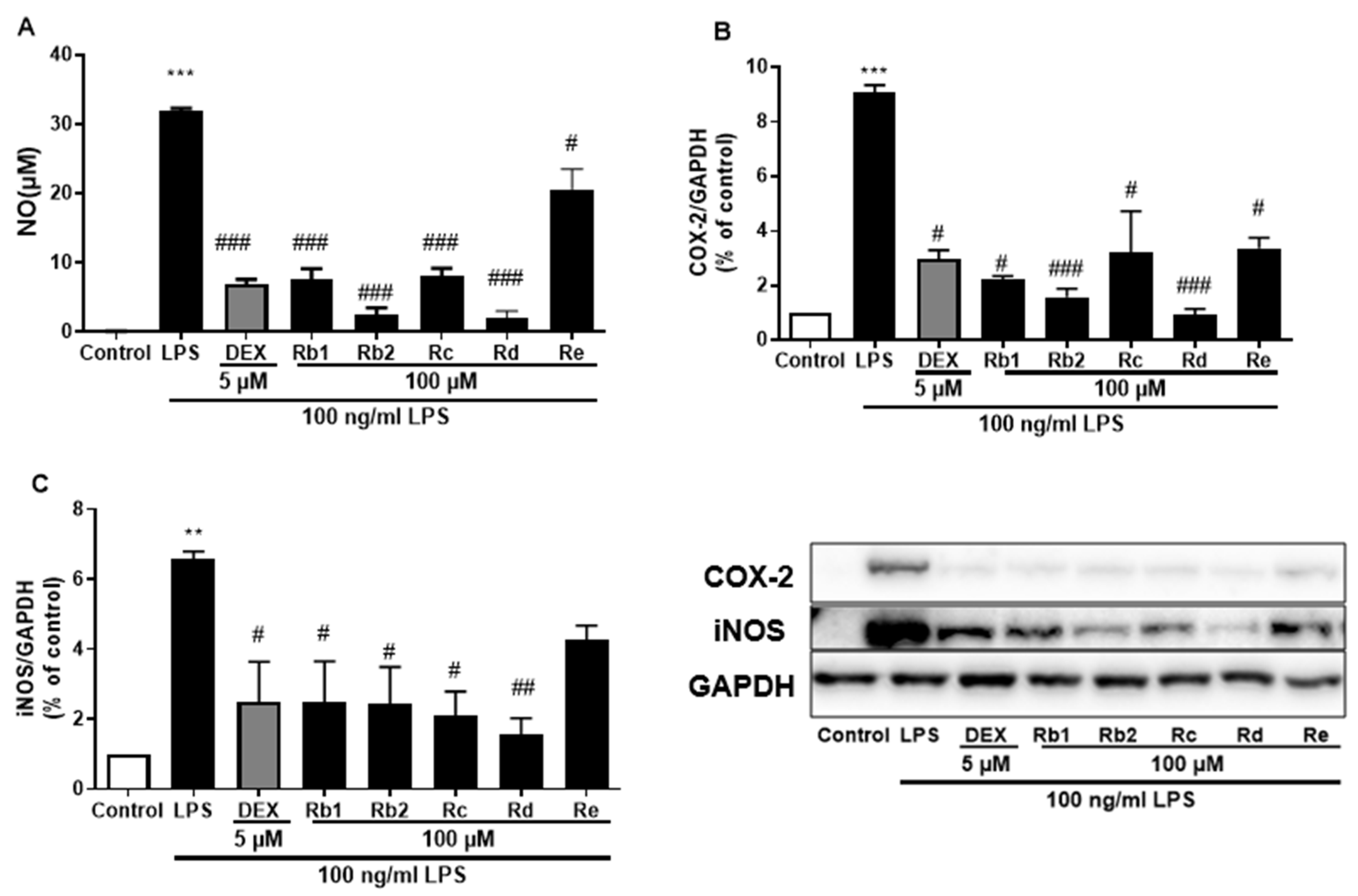
3.5. Ginsenoside Rb2 and Rd in S2 Extract Mainly Attenuates Neuroinflammation in LPS-Induced BV-2 Microglia Cell
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Marciniak, R.; Sheardova, K.; Čermáková, P.; Hudeček, D.; Šumec, R.; Hort, J. Effect of Meditation on Cognitive Functions in Context of Aging and Neurodegenerative Diseases. Front. Behav. Neurosci. 2014, 8, 17. [Google Scholar] [CrossRef]
- Rummel, N.G.; Butterfield, D.A. Altered metabolism in Alzheimer disease brain: Role of oxidative stress. Antioxid. Redox Signal. 2022, 36, 1289–1305. [Google Scholar] [CrossRef]
- Hampel, H.; Mesulam, M.-M.; Cuello, A.C.; Farlow, M.R.; Giacobini, E.; Grossberg, G.T.; Khachaturian, A.S.; Vergallo, A.; Cavedo, E.; Snyder, P.J.; et al. The cholinergic system in the pathophysiology and treatment of Alzheimer’s disease. Brain 2018, 141, 1917–1933. [Google Scholar] [CrossRef] [PubMed]
- Morris, G.P.; Clark, I.A.; Zinn, R.; Vissel, B. Microglia: A new frontier for synaptic plasticity, learning and memory, and neurodegenerative disease research. Neurobiol. Learn. Mem. 2013, 105, 40–53. [Google Scholar] [CrossRef] [PubMed]
- Richetin, K.; Steullet, P.; Pachoud, M.; Perbet, R.; Parietti, E.; Maheswaran, M.; Eddarkaoui, S.; Bégard, S.; Pythoud, C.; Rey, M.; et al. Tau accumulation in astrocytes of the dentate gyrus induces neuronal dysfunction and memory deficits in Alzheimer’s disease. Nat. Neurosci. 2020, 23, 1567–1579. [Google Scholar] [CrossRef]
- Jahn, H. Memory loss in Alzheimer’s disease. Dialogues Clin. Neurosci. 2022, 15, 445–454. [Google Scholar] [CrossRef]
- Ozben, T.; Ozben, S. Neuro-inflammation and anti-inflammatory treatment options for Alzheimer’s disease. Clin. Biochem. 2019, 72, 87–89. [Google Scholar] [CrossRef]
- Selles, M.C.; Oliveira, M.M.; Ferreira, S.T. Brain Inflammation Connects Cognitive and Non-Cognitive Symptoms in Alzheimer’s Disease. J. Alzheimer’s Dis. 2018, 64, S313–S327. [Google Scholar] [CrossRef]
- Chen, W.N.; Yeong, K.Y. Scopolamine, a toxin-induced experimental model, used for research in Alzheimer’s disease. CNS Neurol. Disord.-Drug Targets (Former. Curr. Drug Targets-CNS Neurol. Disord.) 2020, 19, 85–93. [Google Scholar] [CrossRef]
- San Tang, K. The cellular and molecular processes associated with scopolamine-induced memory deficit: A model of Alzheimer’s biomarkers. Life Sci. 2019, 233, 116695. [Google Scholar] [CrossRef]
- Klinkenberg, I.; Blokland, A. The validity of scopolamine as a pharmacological model for cognitive impairment: A review of animal behavioral studies. Neurosci. Biobehav. Rev. 2010, 34, 1307–1350. [Google Scholar] [CrossRef]
- Xiao, H.; Li, H.; Song, H.; Kong, L.; Yan, X.; Li, Y.; Deng, Y.; Tai, H.; Wu, Y.; Ni, Y. Shenzao jiannao oral liquid, an herbal formula, ameliorates cognitive impairments by rescuing neuronal death and triggering endogenous neurogenesis in AD-like mice induced by a combination of Aβ42 and scopolamine. J. Ethnopharmacol. 2020, 259, 112957. [Google Scholar] [CrossRef]
- Baek, S.; Li, F.; Kim, D.; Kim, S.; Kim, M. Enteromorpha prolifera Extract Improves Memory in Scopolamine-Treated Mice via Downregulating Amyloid-β Expression and Upregulating BDNF/TrkB Pathway. Antioxidants 2020, 9, 620. [Google Scholar] [CrossRef]
- Pak, M.E.; Park, Y.J.; Yang, H.J.; Hwang, Y.-H.; Li, W.; Go, Y. Samhwangsasim-tang attenuates neuronal apoptosis and cognitive decline through BDNF-mediated activation of tyrosin kinase B and p75-neurotrophin receptors. Phytomedicine 2022, 99, 153997. [Google Scholar] [CrossRef]
- Nájera, C.; Gallegos-Cedillo, V.M.; Ros, M.; Pascual, J.A. LED Lighting in Vertical Farming Systems Enhances Bioactive Compounds and Productivity of Vegetables Crops. Biol. Life Sci. Forum 2022, 16, 24. [Google Scholar] [CrossRef]
- Hasan, M.; Bashir, T.; Ghosh, R.; Lee, S.K.; Bae, H. An Overview of LEDs’ Effects on the Production of Bioactive Compounds and Crop Quality. Molecules 2017, 22, 1420. [Google Scholar] [CrossRef] [PubMed]
- Chiocchio, I.; Barbaresi, A.; Barbanti, L.; Mandrone, M.; Poli, F.; Torreggiani, D.; Trenta, M.; Tassinari, P. Effects of LED supplemental lighting on the growth and metabolomic profile of Taxus baccata cultivated in a smart greenhouse. PLoS ONE 2022, 17, e0266777. [Google Scholar] [CrossRef] [PubMed]
- Huang, X.; Li, N.; Pu, Y.; Zhang, T.; Wang, B. Neuroprotective Effects of Ginseng Phytochemicals: Recent Perspectives. Molecules 2019, 24, 2939. [Google Scholar] [CrossRef]
- Hou, W.; Wang, Y.; Zheng, P.; Cui, R. Effects of Ginseng on Neurological Disorders. Front. Cell. Neurosci. 2020, 14, 55. [Google Scholar] [CrossRef]
- de Oliveira Zanuso, B.; Santos, A.R.D.O.D.; Miola, V.F.B.; Campos, L.M.G.; Spilla, C.S.G.; Barbalho, S.M. Panax ginseng and aging related disorders: A systematic review. Exp. Gerontol. 2022, 161, 111731. [Google Scholar] [CrossRef]
- Muhammad, T.; Ali, T.; Ikram, M.; Khan, A.; Alam, S.I.; Kim, M.O. Melatonin Rescue Oxidative Stress-Mediated Neuroinflammation/ Neurodegeneration and Memory Impairment in Scopolamine-Induced Amnesia Mice Model. J. Neuroimmune Pharmacol. 2019, 14, 278–294. [Google Scholar] [CrossRef] [PubMed]
- Pariyar, R.; Yoon, C.-S.; Svay, T.; Kim, D.-S.; Cho, H.-K.; Kim, S.Y.; Oh, H.; Kim, Y.-C.; Kim, J.; Lee, H.-S.; et al. Vitis labruscana leaf extract ameliorates scopolamine-induced impairments with activation of Akt, ERK and CREB in mice. Phytomedicine 2017, 36, 8–17. [Google Scholar] [CrossRef] [PubMed]
- Chen, D.-B.; Gao, H.-W.; Peng, C.; Pei, S.-Q.; Dai, A.-R.; Yu, X.-T.; Zhou, P.; Wang, Y.; Cai, B. Quinones as preventive agents in Alzheimer’s diseases: Focus on NLRP3 inflammasomes. J. Pharm. Pharmacol. 2020, 72, 1481–1490. [Google Scholar] [CrossRef]
- Cheon, S.Y.; Koo, B.-N.; Kim, S.Y.; Kam, E.H.; Nam, J.; Kim, E.J. Scopolamine promotes neuroinflammation and delirium-like neuropsychiatric disorder in mice. Sci. Rep. 2021, 11, 8376. [Google Scholar] [CrossRef]
- Trifilieff, P.; Herry, C.; Vanhoutte, P.; Caboche, J.; Desmedt, A.; Riedel, G.; Mons, N.; Micheau, J. Foreground contextual fear memory consolidation requires two independent phases of hippocampal ERK/CREB activation. Learn. Mem. 2006, 13, 349–358. [Google Scholar] [CrossRef]
- Tully, T.; Bourtchouladze, R.; Scott, R.; Tallman, J. Targeting the CREB pathway for memory enhancers. Nat. Rev. Drug Discov. 2003, 2, 267–277. [Google Scholar] [CrossRef]
- Amidfar, M.; de Oliveira, J.; Kucharska, E.; Budni, J.; Kim, Y.-K. The role of CREB and BDNF in neurobiology and treatment of Alzheimer’s disease. Life Sci. 2020, 257, 118020. [Google Scholar] [CrossRef]
- Peng, S.; Zhang, Y.; Zhang, J.; Wang, H.; Ren, B. ERK in Learning and Memory: A Review of Recent Research. Int. J. Mol. Sci. 2010, 11, 222–232. [Google Scholar] [CrossRef]
- Zhao, X.; Li, S.; Gaur, U.; Zheng, W. Artemisinin improved neuronal functions in Alzheimer’s disease animal model 3xtg mice and neuronal cells via stimulating the ERK/CREB signaling pathway. Aging Dis. 2020, 11, 801. [Google Scholar] [CrossRef]
- Lv, J.; Lu, C.; Jiang, N.; Wang, H.; Huang, H.; Chen, Y.; Li, Y.; Liu, X. Protective effect of ginsenoside Rh2 on scopolamine-induced memory deficits through regulation of cholinergic transmission, oxidative stress and the ERK-CREB-BDNF signaling pathway. Phytother. Res. 2021, 35, 337–345. [Google Scholar] [CrossRef]
- Akiyama, H.; Barger, S.; Barnum, S.; Bradt, B.; Bauer, J.; Cole, G.M.; Cooper, N.R.; Eikelenboom, P.; Emmerling, M.; Fiebich, B.L. Inflammation and Alzheimer’s disease. Neurobiol. Aging 2000, 21, 383–421. [Google Scholar] [CrossRef] [PubMed]
- Iqbal, S.; Shah, F.A.; Naeem, K.; Nadeem, H.; Sarwar, S.; Ashraf, Z.; Imran, M.; Khan, T.; Anwar, T.; Li, S. Succinamide Derivatives Ameliorate Neuroinflammation and Oxidative Stress in Scopolamine-Induced Neurodegeneration. Biomolecules 2020, 10, 443. [Google Scholar] [CrossRef] [PubMed]
- Cheon, S.Y.; Koo, B.-N.; Kim, S.Y.; Kam, E.H.; Nam, J.; Kim, E.J. Scopolamine-induced delirium promotes neuroinflammation and neuropsychiatric disorder in mice. Res. Sq. 2021, 11, 8376. [Google Scholar] [CrossRef]
- Wang, Z.; Zhang, S.; Xiao, Y.; Zhang, W.; Wu, S.; Qin, T.; Yue, Y.; Qian, W.; Li, L. NLRP3 inflammasome and inflammatory diseases. Oxidative Med. Cell. Longev. 2020, 2020, 4063562. [Google Scholar] [CrossRef]
- Yoon, C.; Huh, M.; Kang, S.-G.; Park, J.; Lee, C. Implement smart farm with IoT technology. In Proceedings of the 2018 20th International Conference on advanced communication technology (ICACT), Chuncheon, Republic of Korea, 11–14 February 2018; pp. 749–752. [Google Scholar]
- O’Grady, M.J.; O’Hare, G.M. Modelling the smart farm. Inf. Process. Agric. 2017, 4, 179–187. [Google Scholar] [CrossRef]
- Lee, T.K.; Lee, J.Y.; Cho, Y.-J.; Kim, J.-E.; Kim, S.Y.; Park, J.H.Y.; Yang, H.; Lee, K.W. Optimization of the extraction process of high levels of chlorogenic acid and ginsenosides from short-term hydroponic-cultured ginseng and evaluation of the extract for the prevention of atopic dermatitis. J. Ginseng Res. 2021, 46, 367–375. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | D1 Extract | D2 Extract | S2 Extract | |
|---|---|---|---|---|
| Ginsinoside 16 content (mg/g) | Rg1 | 8.26 | 7.59 | 8.57 |
| Rf | 1.43 | 1.34 | 1.73 | |
| Rh1 | 0.91 | 0.98 | 0.80 | |
| Rg2 | 2.10 | 1.66 | 2.07 | |
| Rb1 | 5.85 | 8.28 | 11.22 ** | |
| Rc | 3.16 | 6.13 | 7.39 ** | |
| Rb2 | 3.16 | 6.43 | 7.73 ** | |
| Rb3 | 0.16 | 1.07 | 1.32 | |
| Rd | 4.82 | 13.60 ** | 17.22 *** | |
| F2 | 12.57 ** | 3.07 | 8.39 | |
| Rg3 | N.D | N.D | N.D | |
| Rk1 | N.D | N.D | N.D | |
| Rg5 | N.D | N.D | N.D | |
| Rh2 | N.D | N.D | N.D | |
| Re | 34.94 | 29.12 | 34.95 | |
| F1 | 1.58 | 2.46 | 1.92 | |
| Ginsenosides (mg/g) | 78.96 | 81.73 | 103.32 ** | |
| Ginsenoside Content (mg/g) | NO Inhibition Efficacy (%) | COX-2 Inhibition Efficacy (%) | Relative Increase Rate (S2/D2, %) | |||
|---|---|---|---|---|---|---|
| D1 | D2 | S2 | ||||
| Rb1 | 5.85 | 8.28 | 11.22 | 62.13% | 75.75% | 135.51% |
| Rb2 | 3.16 | 6.43 | 7.73 | 62.96% | 82.67% | 120.22% |
| Rc | 3.16 | 6.13 | 7.39 | 68.28% | 64.49% | 120.55% |
| Rd | 4.82 | 13.6 | 17.22 | 76.05% | 89.63% | 126.62% |
| Re | 34.94 | 29.12 | 34.95 | 35.31% | 62.94% | 120.02% |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huang, T.; Lee, S.; Lee, T.; Yun, S.; Kim, Y.; Yang, H. Smart Farming Enhances Bioactive Compounds Content of Panax ginseng on Moderating Scopolamine-Induced Memory Deficits and Neuroinflammation. Plants 2023, 12, 640. https://doi.org/10.3390/plants12030640
Huang T, Lee S, Lee T, Yun S, Kim Y, Yang H. Smart Farming Enhances Bioactive Compounds Content of Panax ginseng on Moderating Scopolamine-Induced Memory Deficits and Neuroinflammation. Plants. 2023; 12(3):640. https://doi.org/10.3390/plants12030640
Chicago/Turabian StyleHuang, Tianqi, Sangbin Lee, Teamin Lee, Seungbeom Yun, Yongduk Kim, and Hyunok Yang. 2023. "Smart Farming Enhances Bioactive Compounds Content of Panax ginseng on Moderating Scopolamine-Induced Memory Deficits and Neuroinflammation" Plants 12, no. 3: 640. https://doi.org/10.3390/plants12030640
APA StyleHuang, T., Lee, S., Lee, T., Yun, S., Kim, Y., & Yang, H. (2023). Smart Farming Enhances Bioactive Compounds Content of Panax ginseng on Moderating Scopolamine-Induced Memory Deficits and Neuroinflammation. Plants, 12(3), 640. https://doi.org/10.3390/plants12030640