
Divergent Retention of Sucrose Metabolism Genes after Whole Genome Triplication in the Tomato (Solanum lycopersicum)
 , and
, and
Abstract
:1. Introduction
2. Results
2.1. Identification of Sucrose Metabolism Genes in Solanum Lycopersicum and Vitis vinifera
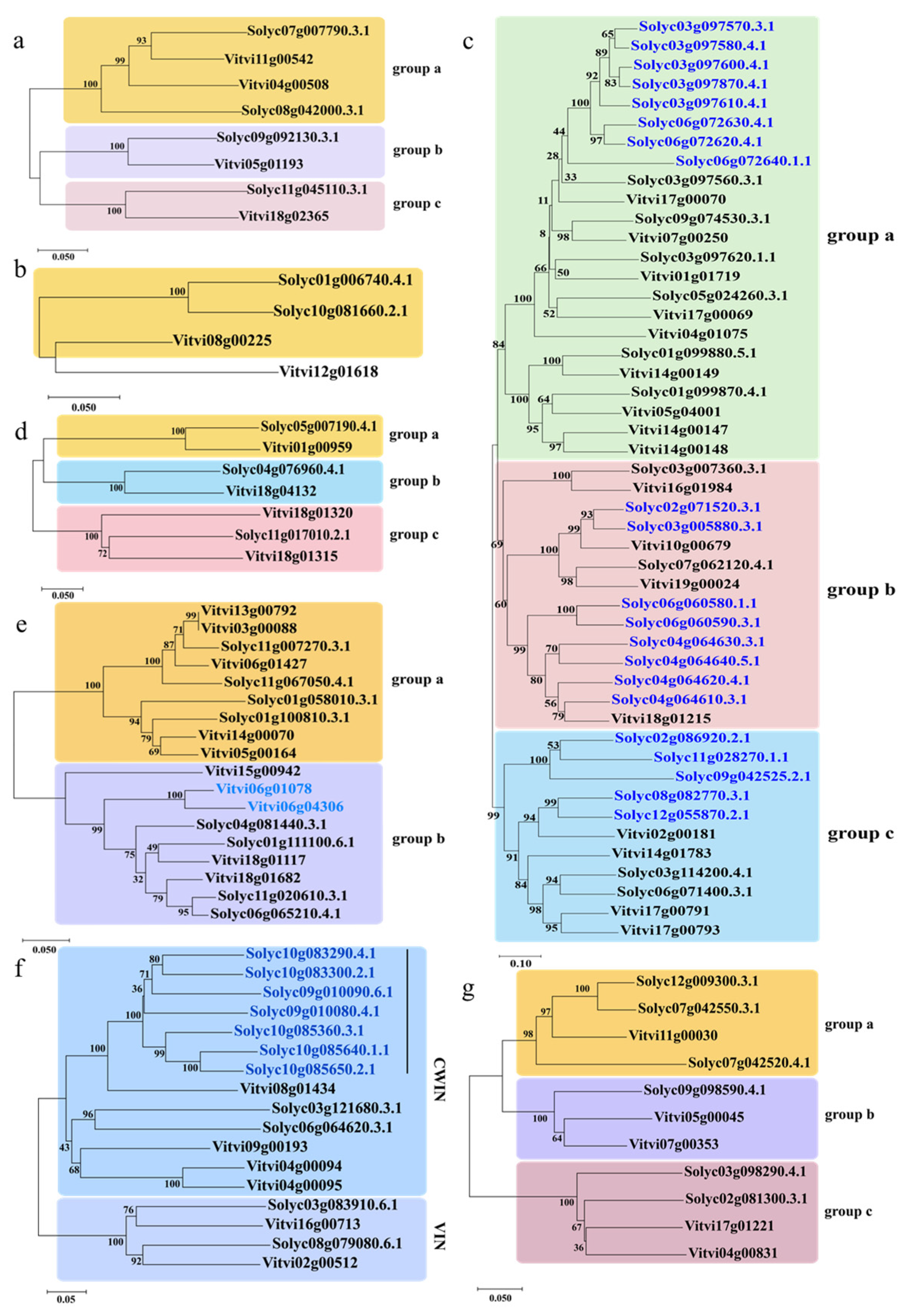
2.2. Phylogenetic Analysis of Sucrose Metabolism Genes in Solanum lycopersicum and Vitis vinifera
2.3. Chromosomal Localization of Sucrose Metabolism Genes in S. lycopersicum and V. vinifera
2.4. Homology Relationship and Synteny Analysis
2.5. Expression Patterns of Sucrose Metabolism Genes in Different Tissues of S. lycopersicum
3. Discussion
4. Materials and Methods
4.1. Identification of Sucrose Metabolism Genes in Solanum lycopersicum and Vitis vinifera
4.2. Phylogenetic Analysis
4.3. Chromosomal Localization and Synteny Analysis
4.4. Transcription Expression Pattern Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Conflicts of Interest
References
- Vargas, W.A.; Salerno, G.L. The Cinderella story of sucrose hydrolysis: Alkaline/neutral invertases, from cyanobacteria to unforeseen roles in plant cytosol and organelles. Plant Sci. 2010, 178, 1–8. [Google Scholar] [CrossRef]
- Vargas, W.; Cumino, A.; Salerno, G.L. Cyanobacterial alkaline/neutral invertases. Origin of sucrose hydrolysis in the plant cytosol? Planta 2003, 216, 951–960. [Google Scholar] [CrossRef] [PubMed]
- Avigad, G.; Dey, P.M. Carbohydrate metabolism: Storage carbohydrates. In Plant Biochemistry; Academic Press: Cambridge, MA, USA, 1997; pp. 143–204. [Google Scholar]
- Ruan, Y.-L. Sucrose metabolism: Gateway to diverse carbon use and sugar signaling. Annu. Rev. Plant Biol. 2014, 65, 33–67. [Google Scholar] [CrossRef]
- Leloir, L.F.; Cardini, C.E. The biosynthesis of sucrose phosphate. J. Biol. Chem. 1955, 214, 157–165. [Google Scholar] [CrossRef] [PubMed]
- Rocher, J.P.; Prioul, J.L.; Lecharny, A.; Reyss, A.; Joussaume, M. Genetic variability in carbon fixation, sucrose-P-synthase and ADP glucose pyrophosphorylase in maize plants of differing growth rate. Plant Physiol. 1989, 89, 416–420. [Google Scholar] [CrossRef] [PubMed]
- Causse, M.; Rocher, J.; Pelleschi, S.; Barrière, Y.; Vienne, D.; Prioul, J. Sucrose phosphate synthase: An enzyme with heterotic activity correlated with maize growth. Crop Sci. 1995, 35, 995–1001. [Google Scholar] [CrossRef]
- Sarquís, J.I.; Gonzalez, H.; Sánchez de Jiménez, E.; Dunlap, J.R. Physiological traits associated with mass selection for improved yield in a maize population. Field Crops Res. 1998, 56, 239–246. [Google Scholar] [CrossRef]
- Zúñiga-Feest, A.; Ort, D.R.; Gutiérrez, A.; Gidekel, M.; Bravo, L.A.; Corcuera, L.J. Light Regulation of sucrose-phosphate synthase activity in the freezing-tolerant grass Deschampsia antarctica. Photosynth. Res. 2005, 83, 75–86. [Google Scholar] [CrossRef]
- Maloney, V.J.; Park, J.-Y.; Unda, F.; Mansfield, S.D. Sucrose phosphate synthase and sucrose phosphate phosphatase interact in Planta and promote plant growth and biomass accumulation. J. Exp. Bot. 2015, 66, 4383–4394. [Google Scholar] [CrossRef]
- Weise, A.; Barker, L.; Kuhn, C.; Lalonde, S.; Buschmann, H.; Frommer, W.B.; Ward, J.M. A new subfamily of sucrose transporters, SUT4, with low affinity/high capacity localized in enucleate sieve elements of plants. Plant Cell 2000, 12, 1345. [Google Scholar] [CrossRef]
- Gabriel-Neumann, E.; Neumann, G.; Leggewie, G.; George, E. Constitutive overexpression of the sucrose transporter SoSUT1 in potato plants increases arbuscular mycorrhiza fungal root colonization under high, but not under low, soil phosphorus availability. J. Plant Physiol. 2011, 168, 911–919. [Google Scholar] [CrossRef] [PubMed]
- Hackel, A.; Schauer, N.; Carrari, F.; Fernie, A.R.; Grimm, B.; Kühn, C. Sucrose transporter LeSUT1 and LeSUT2 inhibition affects tomato fruit development in different ways. Plant J. 2006, 45, 180–192. [Google Scholar] [CrossRef] [PubMed]
- Zheng, Q.-M.; Tang, Z.; Xu, Q.; Deng, X.-X. Isolation, phylogenetic relationship and expression profiling of sugar transporter genes in sweet orange (Citrus sinensis). Plant Cell Tissue Organ Cult. (PCTOC) 2014, 119, 609–624. [Google Scholar] [CrossRef]
- Hu, Z.; Tang, Z.; Zhang, Y.; Niu, L.; Yang, F.; Zhang, D.; Hu, Y. Rice SUT and SWEET transporters. Int. J. Mol. Sci. 2021, 22, 11198. [Google Scholar] [CrossRef]
- Riesmeier, J.W.; Willmitzer, L.; Frommer, W.B. Isolation and characterization of a sucrose carrier cDNA from spinach by functional expression in yeast. EMBO J. 1992, 11, 4705–4713. [Google Scholar] [CrossRef] [PubMed]
- Chen, L.-Q.; Hou, B.-H.; Lalonde, S.; Takanaga, H.; Hartung, M.L.; Qu, X.-Q.; Guo, W.-J.; Kim, J.-G.; Underwood, W.; Chaudhuri, B.; et al. Sugar transporters for intercellular exchange and nutrition of pathogens. Nature 2010, 468, 527–532. [Google Scholar] [CrossRef]
- Chen, L.-Q.; Qu, X.-Q.; Hou, B.-H.; Sosso, D.; Osorio, S.; Fernie, A.R.; Frommer, W.B. Sucrose efflux mediated by SWEET proteins as a key step for phloem transport. Science 2012, 335, 207–211. [Google Scholar] [CrossRef]
- Yuan, M.; Wang, S. Rice MtN3/Saliva/SWEET family genes and their homologs in cellular organisms. Mol. Plant 2013, 6, 665–674. [Google Scholar] [CrossRef]
- Li, X.; Si, W.; Qin, Q.; Wu, H.; Jiang, H. Deciphering evolutionary dynamics of SWEET genes in diverse plant lineages. Sci. Rep. 2018, 8, 13440. [Google Scholar] [CrossRef]
- Zhang, C.; Li, Y.; Wang, J.; Xue, X.; Beuchat, G.; Chen, L. Two evolutionarily duplicated domains individually and post-transcriptionally control SWEET expression for phloem transport. New Phytol. 2021, 232, 1793–1807. [Google Scholar] [CrossRef]
- Seo, P.J.; Park, J.-M.; Kang, S.K.; Kim, S.-G.; Park, C.-M. An Arabidopsis senescence-associated protein SAG29 regulates cell viability under high salinity. Planta 2011, 233, 189–200. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.-L. Signaling role of sucrose metabolism in development. Mol. Plant 2012, 5, 763–765. [Google Scholar] [CrossRef] [PubMed]
- Rolland, F.; Baena-Gonzalez, E.; Sheen, J. Sugar sensing and signaling in plants: Conserved and novel mechanisms. Annu. Rev. Plant Biol. 2006, 57, 675–709. [Google Scholar] [CrossRef] [PubMed]
- Ruan, Y.-L.; Jin, Y.; Yang, Y.-J.; Li, G.-J.; Boyer, J.S. Sugar input, metabolism, and signaling mediated by invertase: Roles in development, yield potential, and response to drought and heat. Mol. Plant 2010, 3, 942–955. [Google Scholar] [CrossRef] [PubMed]
- Yan, W.; Wu, X.; Li, Y.; Liu, G.; Cui, Z.; Jiang, T.; Ma, Q.; Luo, L.; Zhang, P. Cell wall invertase 3 affects cassava productivity via regulating sugar allocation from source to sink. Front. Plant Sci. 2019, 10, 541. [Google Scholar] [CrossRef]
- Beauvoit, B.P.; Colombié, S.; Monier, A.; Andrieu, M.-H.; Biais, B.; Bénard, C.; Chéniclet, C.; Dieuaide-Noubhani, M.; Nazaret, C.; Mazat, J.-P.; et al. Model-assisted analysis of sugar metabolism throughout tomato fruit development reveals enzyme and carrier properties in relation to vacuole expansion. Plant Cell 2014, 26, 3224–3242. [Google Scholar] [CrossRef] [PubMed]
- Lou, Y.; Gou, J.-Y.; Xue, H.-W. PIP5K9, an Arabidopsis Phosphatidylinositol monophosphate kinase, interacts with a cytosolic invertase to negatively regulate sugar-mediated root growth. Plant Cell 2007, 19, 163–181. [Google Scholar] [CrossRef]
- Ruan, Y.-L.; Llewellyn, D.J.; Furbank, R.T. Suppression of sucrose synthase gene expression represses cotton fiber cell initiation, elongation, and seed development. Plant Cell 2003, 15, 952–964. [Google Scholar] [CrossRef]
- Chourey, P.S.; Taliercio, E.W.; Carlson, S.J.; Ruan, Y.L. Genetic evidence that the two isozymes of sucrose synthase present in developing maize endosperm are critical, one for cell wall integrity and the other for starch biosynthesis. Mol. Gen. Genet. 1998, 259, 88–96. [Google Scholar] [CrossRef]
- Zrenner, R.; Salanoubat, M.; Willmitzer, L.; Sonnewald, U. Evidence of the crucial role of sucrose synthase for sink strength using transgenic potato plants (Solanum tuberosum L.). Plant J. 1995, 7, 97–107. [Google Scholar] [CrossRef]
- Coleman, H.D.; Yan, J.; Mansfield, S.D. Sucrose synthase affects carbon partitioning to increase cellulose production and altered cell wall ultrastructure. Proc. Natl. Acad. Sci. USA 2009, 106, 13118–13123. [Google Scholar] [CrossRef] [PubMed]
- Zhang, J. Evolution by gene duplication: An update. Trends Ecol. Evol. 2003, 18, 292–298. [Google Scholar] [CrossRef]
- Wang, X.; Guo, H.; Wang, J.; Lei, T.; Liu, T.; Wang, Z.; Li, Y.; Lee, T.; Li, J.; Tang, H.; et al. Comparative genomic de-convolution of the cotton genome revealed a decaploid ancestor and widespread chromosomal fractionation. New Phytol. 2016, 209, 1252–1263. [Google Scholar] [CrossRef] [PubMed]
- Zeng, L.; Tu, X.-L.; Dai, H.; Han, F.-M.; Lu, B.-S.; Wang, M.-S.; Nanaei, H.A.; Tajabadipour, A.; Mansouri, M.; Li, X.-L.; et al. Whole genomes and transcriptomes reveal adaptation and domestication of pistachio. Genome Biol. 2019, 20, 79. [Google Scholar] [CrossRef] [PubMed]
- Blanc, G.; Wolfe, K.H. Widespread paleopolyploidy in model plant species inferred from age distributions of duplicate genes. Plant Cell 2004, 16, 1667–1678. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Bowers, J.E.; Wang, X.; Ming, R.; Alam, M.; Paterson, A.H. Synteny and collinearity in plant genomes. Science 2008, 320, 486–488. [Google Scholar] [CrossRef] [PubMed]
- Tang, H.; Wang, X.; Bowers, J.E.; Ming, R.; Alam, M.; Paterson, A.H. Unraveling ancient hexaploidy through multiply-aligned angiosperm gene maps. Genome Res. 2008, 18, 1944–1954. [Google Scholar] [CrossRef] [PubMed]
- Paterson, A.H.; Freeling, M.; Tang, H.; Wang, X. Insights from the comparison of plant genome sequences. Annu. Rev. Plant Biol. 2010, 61, 349–372. [Google Scholar] [CrossRef]
- Kondrashov, F.A. Gene duplication as a mechanism of genomic adaptation to a changing environment. Proc. R. Soc. B Biol. Sci. 2012, 279, 5048–5057. [Google Scholar] [CrossRef]
- Qiao, X.; Li, Q.; Yin, H.; Qi, K.; Li, L.; Wang, R.; Zhang, S.; Paterson, A.H. Gene duplication and evolution in recurring polyploidization–diploidization cycles in plants. Genome Biol. 2019, 20, 38. [Google Scholar] [CrossRef]
- Costello, R.; Emms, D.M.; Kelly, S. Gene duplication accelerates the pace of protein gain and loss from plant organelles. Mol. Biol. Evol. 2020, 37, 969–981. [Google Scholar] [CrossRef] [PubMed]
- Birchler, J.A.; Yang, H. The multiple fates of gene duplications: Deletion, hypofunctionalization, subfunctionalization, neofunctionalization, dosage balance constraints, and neutral variation. Plant Cell 2022, 34, 2466–2474. [Google Scholar] [CrossRef] [PubMed]
- Ehrenreich, I.M. Evolution after genome duplication. Science 2020, 368, 1424–1425. [Google Scholar] [CrossRef] [PubMed]
- Jiao, Y.; Li, J.; Tang, H.; Paterson, A.H. Integrated syntenic and phylogenomic analyses reveal an ancient genome duplication in monocots. Plant Cell 2014, 26, 2792–2802. [Google Scholar] [CrossRef] [PubMed]
- Barker, M.S.; Husband, B.C.; Pires, J.C. Spreading winge and flying high: The evolutionary importance of polyploidy after a century of study. Am. J. Bot. 2016, 103, 1139–1145. [Google Scholar] [CrossRef] [PubMed]
- Huang, J.; Xu, W.; Zhai, J.; Hu, Y.; Guo, J.; Zhang, C.; Zhao, Y.; Zhang, L.; Martine, C.; Ma, H.; et al. Nuclear phylogeny and insights into whole genome duplications and reproductive development of Solanaceae plants. Plant Commun. 2023, 4, 100595. [Google Scholar] [CrossRef]
- Wu, S.; Han, B.; Jiao, Y. Genetic contribution of paleopolyploidy to adaptive evolution in angiosperms. Mol. Plant 2020, 13, 59–71. [Google Scholar] [CrossRef]
- Jiao, Y.; Leebens-Mack, J.; Ayyampalayam, S.; Bowers, J.E.; McKain, M.R.; McNeal, J.; Rolf, M.; Ruzicka, D.R.; Wafula, E.; Wickett, N.J.; et al. A genome triplication associated with early diversification of the core eudicots. Genome Biol. 2012, 13, R3. [Google Scholar] [CrossRef]
- Bowers, J.E.; Chapman, B.A.; Rong, J.; Paterson, A.H. Unravelling angiosperm genome evolution by phylogenetic analysis of chromosomal duplication events. Nature 2003, 422, 433–438. [Google Scholar] [CrossRef]
- Blanc, G.; Hokamp, K.; Wolfe, K.H. A recent polyploidy superimposed on older large-scale duplications in the Arabidopsis genome. Genome Res. 2003, 13, 137–144. [Google Scholar] [CrossRef]
- The Potato Genome Sequencing Consortium. Genome sequence and analysis of the tuber crop potato. Nature 2011, 475, 189–195. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Yang, Y.; Liu, C.; Sun, Y.; Zhang, T.; Hou, M.; Huang, S.; Yuan, H. The evolutionary history of the sucrose synthase gene family in higher plants. BMC Plant Biol. 2019, 19, 566. [Google Scholar] [CrossRef] [PubMed]
- The Tomato Genome Consortium. The tomato genome sequence provides insights into fleshy fruit evolution. Nature 2012, 485, 635–641. [Google Scholar] [CrossRef] [PubMed]
- The French–Italian Public Consortium for Grapevine Genome Characterization. The grapevine genome sequence suggests ancestral hexaploidization in major angiosperm phyla. Nature 2007, 449, 463–467. [Google Scholar] [CrossRef] [PubMed]
- Kanayama, Y. Sugar metabolism and fruit development in the tomato. Hortic. J. 2017, 86, 417–425. [Google Scholar] [CrossRef]
- Quinet, M.; Angosto, T.; Yuste-Lisbona, F.J.; Blanchard-Gros, R.; Bigot, S.; Martinez, J.-P.; Lutts, S. Tomato fruit development and metabolism. Front. Plant Sci. 2019, 10, 1554. [Google Scholar] [CrossRef] [PubMed]
- Li, Y.; Wang, H.; Zhang, Y.; Martin, C. Can the world’s favorite fruit, tomato, provide an effective biosynthetic chassis for high-value metabolites? Plant Cell Rep. 2018, 37, 1443–1450. [Google Scholar] [CrossRef] [PubMed]
- Giovannoni, J.J. Genetic regulation of fruit development and ripening. Plant Cell Online 2004, 16, S170–S180. [Google Scholar] [CrossRef]
- Hosmani, P.S.; Flores-Gonzalez, M.; Van De Geest, H.; Maumus, F.; Bakker, L.V.; Schijlen, E.; Van Haarst, J.; Cordewener, J.; Sanchez-Perez, G.; Peters, S.; et al. An improved de novo assembly and annotation of the tomato reference genome using single-molecule sequencing, Hi-C proximity ligation and optical maps. Genomics 2019, 767764. [Google Scholar]
- Alonge, M.; Wang, X.; Benoit, M.; Soyk, S.; Pereira, L.; Zhang, L.; Suresh, H.; Ramakrishnan, S.; Maumus, F.; Ciren, D.; et al. Major impacts of widespread structural variation on gene expression and crop improvement in tomato. Cell 2020, 182, 145–161.e23. [Google Scholar] [CrossRef]
- Su, X.; Wang, B.; Geng, X.; Du, Y.; Yang, Q.; Liang, B.; Meng, G.; Gao, Q.; Yang, W.; Zhu, Y.; et al. A high-continuity and annotated tomato reference genome. BMC Genom. 2021, 22, 898. [Google Scholar] [CrossRef] [PubMed]
- Qin, C.; Yu, C.; Shen, Y.; Fang, X.; Chen, L.; Min, J.; Cheng, J.; Zhao, S.; Xu, M.; Luo, Y.; et al. Whole-genome sequencing of cultivated and wild peppers provides insights into Capsicum domestication and specialization. Proc. Natl. Acad. Sci. USA 2014, 111, 5135–5140. [Google Scholar] [CrossRef] [PubMed]
- Panchy, N.; Lehti-Shiu, M.; Shiu, S.-H. Evolution of gene duplication in plants. Plant Physiol. 2016, 171, 2294–2316. [Google Scholar] [CrossRef] [PubMed]
- Wan, H.; Wu, L.; Yang, Y.; Zhou, G.; Ruan, Y.-L. Evolution of sucrose metabolism: The dichotomy of invertases and beyond. Trends Plant Sci. 2018, 23, 163–177. [Google Scholar] [CrossRef]
- Duan, Y.; Yang, L.; Zhu, H.; Zhou, J.; Sun, H.; Gong, H. Structure and expression analysis of sucrose phosphate synthase, sucrose synthase and invertase gene families in Solanum lycopersicum. Int. J. Mol. Sci. 2021, 22, 4698. [Google Scholar] [CrossRef]
- Feng, C.-Y.; Han, J.-X.; Han, X.-X.; Jiang, J. Genome-wide identification, phylogeny, and expression analysis of the SWEET gene family in tomato. Gene 2015, 573, 261–272. [Google Scholar] [CrossRef]
- Ruhlmann, J.M.; Kram, B.W.; Carter, C.J. Cell wall invertase 4 is required for nectar production in Arabidopsis. J. Exp. Bot. 2010, 61, 395–404. [Google Scholar] [CrossRef]
- Tian, H.; Ma, L.; Zhao, C.; Hao, H.; Gong, B.; Yu, X.; Wang, X. Antisense repression of sucrose phosphate synthase in transgenic muskmelon alters plant growth and fruit development. Biochem. Biophys. Res. Commun. 2010, 393, 365–370. [Google Scholar] [CrossRef]
- Chong, J.; Piron, M.-C.; Meyer, S.; Merdinoglu, D.; Bertsch, C.; Mestre, P. The SWEET family of sugar transporters in grapevine: VvSWEET4 is involved in the interaction with Botrytis cinerea. J. Exp. Bot. 2014, 65, 6589–6601. [Google Scholar] [CrossRef]
- Li, J.; Qin, M.; Qiao, X.; Cheng, Y.; Li, X.; Zhang, H.; Wu, J. A new insight into the evolution and functional divergence of SWEET transporters in chinese white pear (Pyrus bretschneideri). Plant Cell Physiol. 2017, 58, 839–850. [Google Scholar] [CrossRef]
- Anur, R.M.; Mufithah, N.; Sawitri, W.D.; Sakakibara, H.; Sugiharto, B. Overexpression of sucrose phosphate synthase enhanced sucrose content and biomass production in transgenic sugarcane. Plants 2020, 9, 200. [Google Scholar] [CrossRef] [PubMed]
- Ru, L.; Chen, B.; Li, Y.; Wills, R.B.H.; Lv, Z.; Lu, G.; Yang, H. Role of sucrose phosphate synthase and vacuolar invertase in postharvest sweetening of immature sweetpotato tuberous roots (Ipomoea batatas (L.) Lam Cv ‘Xinxiang’). Sci. Hortic. 2021, 282, 110007. [Google Scholar] [CrossRef]
- Blanc, G.; Wolfe, K.H. Functional divergence of duplicated genes formed by polyploidy during Arabidopsis evolution. Plant Cell 2004, 16, 1679–1691. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Zhang, Y.; Xu, N.; Zhang, H.; Fan, Y.; Rui, C.; Han, M.; Malik, W.A.; Wang, Q.; Sun, L.; et al. Genome-wide identification of CK gene family suggests functional expression pattern against Cd2+ stress in Gossypium hirsutum L. Int. J. Biol. Macromol. 2021, 188, 272–282. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Li, M.; Wu, X.; Wang, J. The gene structure and expression level changes of the GH3 gene family in Brassica napus relative to its diploid ancestors. Genes 2019, 10, 58. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.-L.; Li, Y.; Fan, Y.-F.; Li, Z.; Yoshida, K.; Wang, J.-Y.; Ma, X.-K.; Wang, N.; Mitsuda, N.; Kotake, T.; et al. Wolfberry genomes and the evolution of Lycium (Solanaceae). Commun. Biol. 2021, 4, 671. [Google Scholar] [CrossRef] [PubMed]
- Cannon, S.B.; Mitra, A.; Baumgarten, A.; Young, N.D.; May, G. The roles of segmental and tandem gene duplication in the evolution of large gene families in Arabidopsis thaliana. BMC Plant Biol. 2004, 4, 10. [Google Scholar] [CrossRef]
- Ye, C.-Y.; Wu, D.; Mao, L.; Jia, L.; Qiu, J.; Lao, S.; Chen, M.; Jiang, B.; Tang, W.; Peng, Q.; et al. The genomes of the allohexaploid Echinochloa crus-galli and its progenitors provide insights into polyploidization-driven adaptation. Mol. Plant 2020, 13, 1298–1310. [Google Scholar] [CrossRef]
- Chen, S.; Hajirezaei, M.; Peisker, M.; Tschiersch, H.; Sonnewald, U.; Börnke, F. Decreased sucrose-6-phosphate phosphatase level in transgenic tobacco inhibits photosynthesis, alters carbohydrate partitioning, and reduces growth. Planta 2005, 221, 479–492. [Google Scholar] [CrossRef]
- Bezrutczyk, M.; Hartwig, T.; Horschman, M.; Char, S.N.; Yang, J.; Yang, B.; Frommer, W.B.; Sosso, D. Impaired phloem loading in zmsweet13a,b,c sucrose transporter triple knock-out mutants in Zea mays. New Phytol. 2018, 218, 594–603. [Google Scholar] [CrossRef]
- Liu, X.; Zhang, Y.; Yang, C.; Tian, Z.; Li, J. AtSWEET4, a hexose facilitator, mediates sugar transport to axial sinks and affects plant development. Sci. Rep. 2016, 6, 24563. [Google Scholar] [CrossRef] [PubMed]
- Mizuno, H.; Kasuga, S.; Kawahigashi, H. The sorghum SWEET gene family: Stem sucrose accumulation as revealed through transcriptome profiling. Biotechnol. Biofuels 2016, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- Zeng, X.; Luo, Y.; Vu, N.T.Q.; Shen, S.; Xia, K.; Zhang, M. CRISPR/Cas9-mediated mutation of OsSWEET14 in rice cv. Zhonghua11 confers resistance to Xanthomonas oryzae pv. oryzae without yield penalty. BMC Plant Biol. 2020, 20, 313. [Google Scholar] [CrossRef] [PubMed]
- Zhou, A.; Ma, H.; Feng, S.; Gong, S.; Wang, J. A novel sugar transporter from Dianthus spiculifolius, DsSWEET12, affects sugar metabolism and confers osmotic and oxidative stress tolerance in Arabidopsis. Int. J. Mol. Sci. 2018, 19, 497. [Google Scholar] [CrossRef] [PubMed]
- Salts, Y.; Sobolev, I.; Chmelnitsky, I.; Shabtai, S.; Barg, R. Genomic structure and expression of Lestd1, a seven-transmembrane-domain protein-encoding gene specifically expressed in tomato pollen. Isr. J. Plant Sci. 2005, 53, 79–88. [Google Scholar] [CrossRef]
- Force, A.; Lynch, M.; Pickett, F.B.; Amores, A.; Yan, Y.; Postlethwait, J. Preservation of duplicate genes by complementary, degenerative mutations. Genetics 1999, 151, 1531–1545. [Google Scholar] [CrossRef]
- Van Hoof, A. Conserved functions of yeast genes support the duplication, degeneration and complementation model for gene duplication. Genetics 2005, 171, 1455–1461. [Google Scholar] [CrossRef]
- Kühn, C.; Grof, C.P. Sucrose transporters of higher plants. Curr. Opin. Plant Biol. 2010, 13, 287–297. [Google Scholar] [CrossRef]
- Amor, Y.; Haigler, C.H.; Johnson, S.; Wainscott, M.; Delmer, D.P. A membrane-associated form of sucrose synthase and its potential role in synthesis of cellulose and callose in plants. Proc. Natl. Acad. Sci. USA 1995, 92, 9353–9357. [Google Scholar] [CrossRef]
- Jiang, Y.; Guo, W.; Zhu, H.; Ruan, Y.-L.; Zhang, T. Overexpression of GhSusA1 increases plant biomass and improves cotton fiber yield and quality: Role of a novel sucrose synthase gene in cotton. Plant Biotechnol. J. 2012, 10, 301–312. [Google Scholar] [CrossRef]
- Stein, O.; Granot, D. An overview of sucrose synthases in plants. Front. Plant Sci. 2019, 10, 95. [Google Scholar] [CrossRef]
- Fallahi, H.; Scofield, G.N.; Badger, M.R.; Chow, W.S.; Furbank, R.T.; Ruan, Y.-L. Localization of sucrose synthase in developing seed and siliques of Arabidopsis thaliana reveals diverse roles for SUS during development. J. Exp. Bot. 2008, 59, 3283–3295. [Google Scholar] [CrossRef] [PubMed]
- Akashi, H. Gene expression and molecular evolution. Curr. Opin. Genet. Dev. 2001, 11, 660–666. [Google Scholar] [CrossRef]
- Long, M.; Betrán, E.; Thornton, K.; Wang, W. The origin of new genes: Glimpses from the young and old. Nat. Rev. Genet. 2003, 4, 865–875. [Google Scholar] [CrossRef] [PubMed]
- Prince, V.E.; Pickett, F.B. Splitting pairs: The diverging fates of duplicated genes. Nat. Rev. Genet. 2002, 3, 827–837. [Google Scholar] [CrossRef] [PubMed]
- Velt, A.; Frommer, B.; Blanc, S.; Holtgräwe, D.; Duchêne, É.; Dumas, V.; Grimplet, J.; Hugueney, P.; Kim, C.; Lahaye, M.; et al. An improved reference of the grapevine genome reasserts the origin of the PN40024 highly homozygous genotype. G3 Genes Genomes Genet. 2023, 13, jkad067. [Google Scholar] [CrossRef] [PubMed]
- Zhang, L.; Chen, F.; Zhang, X.; Li, Z.; Zhao, Y.; Lohaus, R.; Chang, X.; Dong, W.; Ho, S.Y.W.; Liu, X.; et al. The water lily genome and the early evolution of flowering plants. Nature 2020, 577, 79–84. [Google Scholar] [CrossRef]
- Kumar, S.; Stecher, G.; Li, M.; Knyaz, C.; Tamura, K. MEGA X: Molecular evolutionary genetics analysis across computing platforms. Mol. Biol. Evol. 2018, 35, 1547–1549. [Google Scholar] [CrossRef]
- Chao, J.; Li, Z.; Sun, Y.; Aluko, O.O.; Wu, X.; Wang, Q.; Liu, G. MG2C: A user-friendly online tool for drawing genetic maps. Mol. Hortic. 2021, 1, 16. [Google Scholar] [CrossRef]
- Hanada, K.; Zou, C.; Lehti-Shiu, M.D.; Shinozaki, K.; Shiu, S.-H. Importance of lineage-specific expansion of plant tandem duplicates in the adaptive response to environmental stimuli. Plant Physiol. 2008, 148, 993–1003. [Google Scholar] [CrossRef]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]
- Hurst, L.D. The Ka/Ks ratio: Diagnosing the form of sequence evolution. Trends Genet. 2002, 18, 486. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Solanum lycopersicum | Vitis vinifera | Ka | Ks | Ka/Ks | |
|---|---|---|---|---|---|
| SPS | Solyc07g007790 | Vitvi11g00542 | 0.11362809 | 1.33703922 | 0.08498486 |
| Solyc07g007790 | Vitvi04g00508 | 0.17194748 | 1.39563813 | 0.123203481 | |
| Solyc08g042000 | Vitvi04g00508 | 0.19964061 | 1.04116391 | 0.19174753 | |
| SPP | Solyc01g006740 | Vitvi08g00225 | 0.16184532 | 1.33939481 | 0.120834664 |
| Solyc10g081660 | Vitvi08g00225 | 0.16741032 | 1.06108889 | 0.157772192 | |
| SWEET | Solyc02g086920 | Vitvi14g01783 | 0.40602836 | 1.40050661 | 0.289915347 |
| Solyc02g086920 | Vitvi17g00791 | 0.41698207 | 2.39161681 | 0.174351537 | |
| Solyc01g099870 | Vitvi14g00147 | 0.35410311 | 1.36055722 | 0.260263296 | |
| Solyc01g099870 | Vitvi05g04001 | 0.2686224 | 1.48840512 | 0.180476669 | |
| Solyc01g099880 | Vitvi14g04044 | 0.66660488 | N | N | |
| Solyc02g071520 | Vitvi10g00679 | 0.21747408 | 1.14595337 | 0.189775683 | |
| Solyc03g005880 | Vitvi10g00679 | 0.19707583 | 1.95513806 | 0.10079893 | |
| Solyc03g007360 | Vitvi16g01984 | 0.21263783 | 2.28626265 | 0.093006738 | |
| Solyc03g097560 | Vitvi01g01719 | 0.32878582 | N | N | |
| Solyc03g097560 | Vitvi17g00069 | 0.37389825 | 2.41720521 | 0.154682045 | |
| Solyc03g114200 | Vitvi14g01783 | 0.39521583 | N | N | |
| Solyc03g114200 | Vitvi17g00791 | 0.26071019 | 1.43942492 | 0.181121076 | |
| Solyc04g064610 | Vitvi18g01215 | 0.17285757 | 2.93820554 | 0.058831 | |
| Solyc05g024260 | Vitvi01g01719 | 0.36347455 | 2.05818095 | 0.176599899 | |
| Solyc06g071400 | Vitvi14g01783 | 0.36411637 | 3.26503459 | 0.111519912 | |
| Solyc06g071400 | Vitvi17g00791 | 0.23992208 | 3.88509738 | 0.061754458 | |
| Solyc07g062120 | Vitvi19g00024 | 0.24415894 | 1.29049766 | 0.189197504 | |
| Solyc08g082770 | Vitvi02g00181 | 0.3428003 | 2.44187365 | 0.140384128 | |
| Solyc09g074530 | Vitvi07g00250 | 0.26903608 | N | N | |
| Solyc06g072620 | Vitvi17g00069 | 0.30971683 | 1.9635318 | 0.157734566 | |
| SUT | Solyc05g007190 | Vitvi01g00959 | 0.16589023 | 1.15491739 | 0.143638179 |
| Solyc04g076960 | Vitvi18g04132 | 0.16269373 | 1.61605975 | 0.10067309 | |
| CIN | Solyc01g100810 | Vitvi14g00070 | 0.14473684 | 1.32718543 | 0.10905548 |
| Solyc01g111100 | Vitvi18g01117 | 0.1469987 | 1.71248171 | 0.085839576 | |
| Solyc01g111100 | Vitvi03g00088 | 0.11761239 | 1.2100907 | 0.097193039 | |
| Solyc04g081440 | Vitvi18g01117 | 0.12531074 | 1.23472624 | 0.101488682 | |
| Solyc11g007270 | Vitvi13g00792 | 0.13692573 | 1.09844259 | 0.124654428 | |
| Solyc11g067050 | Vitvi13g00792 | 0.16899417 | 1.54111736 | 0.109656911 | |
| Solyc11g067050 | Vitvi06g01427 | 0.18299346 | 1.20043492 | 0.152439299 | |
| Solyc06g065210 | Vitvi18g01682 | 0.06598712 | 1.43375694 | 0.046023927 | |
| Solyc01g058010 | Vitvi05g00164 | 0.17749537 | 1.58143294 | 0.112237049 | |
| CWIN | Solyc09g010080 | Vitvi08g01434 | 0.25575845 | 1.64857908 | 0.155138722 |
| Solyc10g083290 | Vitvi08g01434 | 0.23448308 | 1.73611241 | 0.135062155 | |
| VIN | Solyc03g083910 | Vitvi16g00713 | 0.267711178 | 1.948821335 | 0.137370816 |
| Solyc03g083910 | Vitvi02g00512 | 0.276151171 | 2.657257382 | 0.103923381 | |
| Solyc08g079080 | Vitvi02g00512 | 0.29190687 | 2.080093594 | 0.140333527 | |
| SUS | Solyc12g009300 | Vitvi11g00030 | 0.10168938 | 1.58385903 | 0.064203557 |
| Solyc07g042520 | Vitvi11g00030 | 0.17376854 | 1.49401818 | 0.116309523 |
| Solanum lycopersicum | Solanum lycopersicum | Ka | Ks | Ka/Ks | |
|---|---|---|---|---|---|
| SPS | Solyc01g006740 | Solyc10g081660 | 0.062151704 | 0.65616436 | 0.094719719 |
| SWEET | Solyc02g086920 | Solyc03g114200 | 0.466864795 | 3.05815308 | 0.152662336 |
| Solyc02g086920 | Solyc06g071400 | 0.485013419 | 3.28256218 | 0.147754526 | |
| Solyc02g071520 | Solyc03g005880 | 0.108893908 | 0.89339854 | 0.121887269 | |
| Solyc03g097610 | Solyc06g072630 | 0.201591662 | 0.89821045 | 0.224437004 | |
| Solyc02g086920 | Solyc03g114200 | 0.466864795 | 3.05815308 | 0.152662336 | |
| Solyc03g114200 | Solyc06g071400 | 0.205827634 | 1.19510213 | 0.172225979 | |
| CIN | Solyc11g007270 | Solyc11g067050 | 0.201332217 | 1.87098003 | 0.107607892 |
| Acid INV | Solyc09g010080 | Solyc10g083290 | 0.158937563 | 1.07235908 | 0.148213006 |
| Solyc10g083290 | Solyc10g085650 | 0.183792572 | 1.14966465 | 0.159866246 | |
| Solyc10g083300 | Solyc10g085640 | 0.203293349 | 1.55918308 | 0.130384527 | |
| Solyc03g083910 | Solyc08g079080 | 0.308055691 | 2.07739243 | 0.148289599 | |
| SUS | Solyc07g042520 | Solyc12g009300 | 0.193685014 | 2.37941318 | 0.081400328 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xu, Y.; Yao, Z.; Cheng, Y.; Ruan, M.; Ye, Q.; Wang, R.; Zhou, G.; Liu, J.; Liu, C.; Wan, H. Divergent Retention of Sucrose Metabolism Genes after Whole Genome Triplication in the Tomato (Solanum lycopersicum). Plants 2023, 12, 4145. https://doi.org/10.3390/plants12244145
Xu Y, Yao Z, Cheng Y, Ruan M, Ye Q, Wang R, Zhou G, Liu J, Liu C, Wan H. Divergent Retention of Sucrose Metabolism Genes after Whole Genome Triplication in the Tomato (Solanum lycopersicum). Plants. 2023; 12(24):4145. https://doi.org/10.3390/plants12244145
Chicago/Turabian StyleXu, Yang, Zhuping Yao, Yuan Cheng, Meiying Ruan, Qingjing Ye, Rongqing Wang, Guozhi Zhou, Jia Liu, Chaochao Liu, and Hongjian Wan. 2023. "Divergent Retention of Sucrose Metabolism Genes after Whole Genome Triplication in the Tomato (Solanum lycopersicum)" Plants 12, no. 24: 4145. https://doi.org/10.3390/plants12244145
APA StyleXu, Y., Yao, Z., Cheng, Y., Ruan, M., Ye, Q., Wang, R., Zhou, G., Liu, J., Liu, C., & Wan, H. (2023). Divergent Retention of Sucrose Metabolism Genes after Whole Genome Triplication in the Tomato (Solanum lycopersicum). Plants, 12(24), 4145. https://doi.org/10.3390/plants12244145