
Root Development Monitoring under Different Water Supply Levels in Processing Tomato Plants


 ,
,  ,
, 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
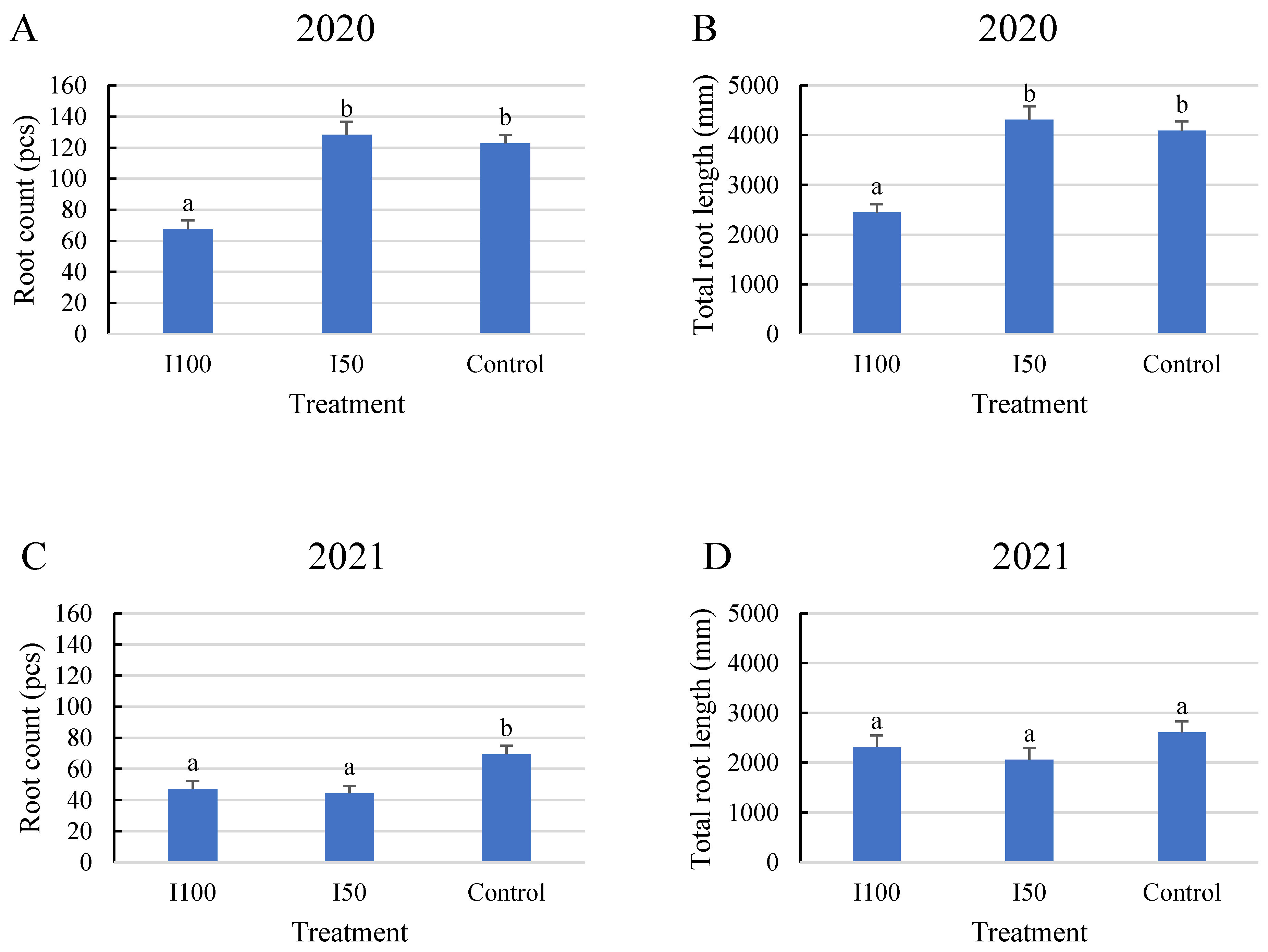
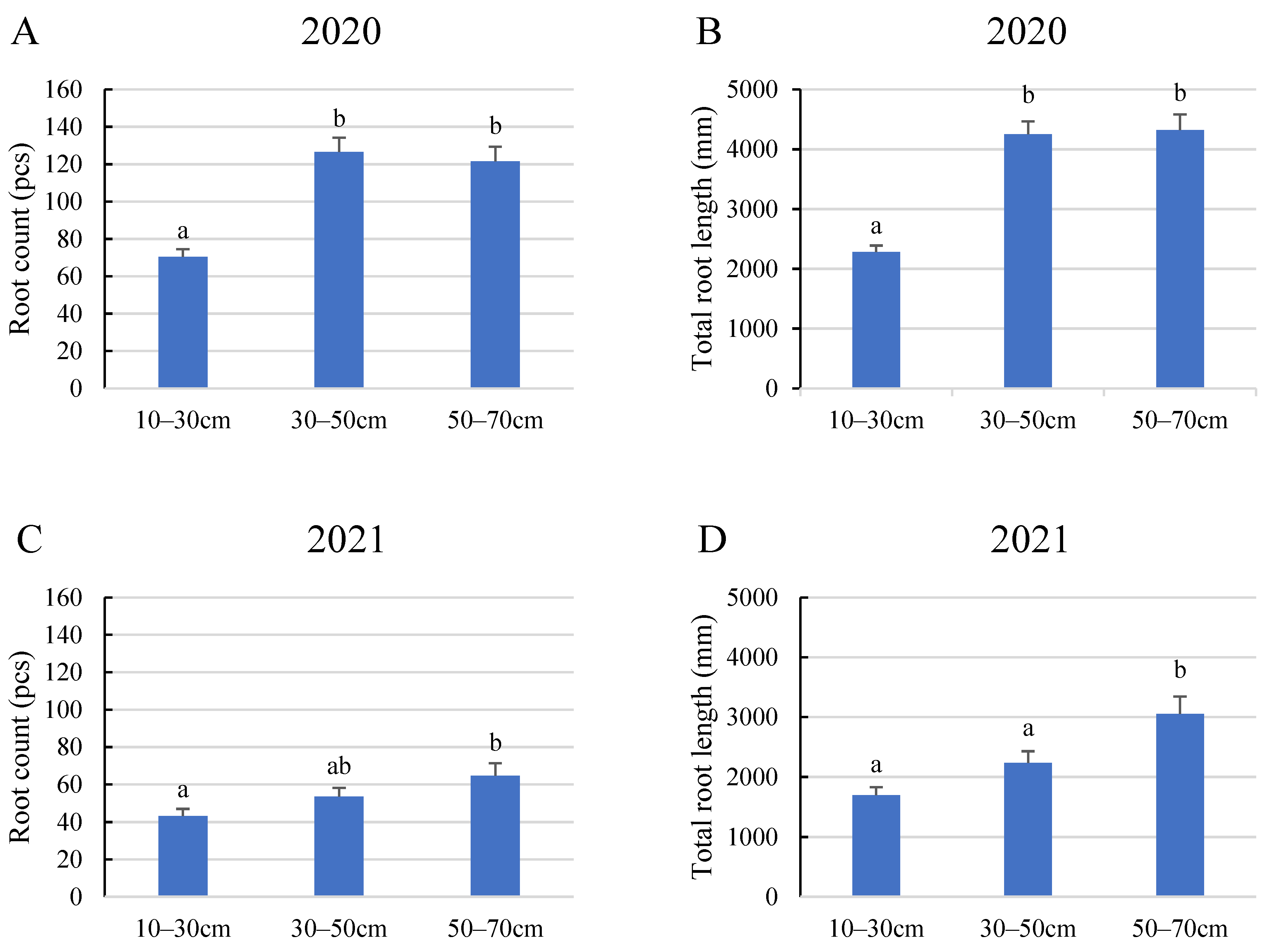
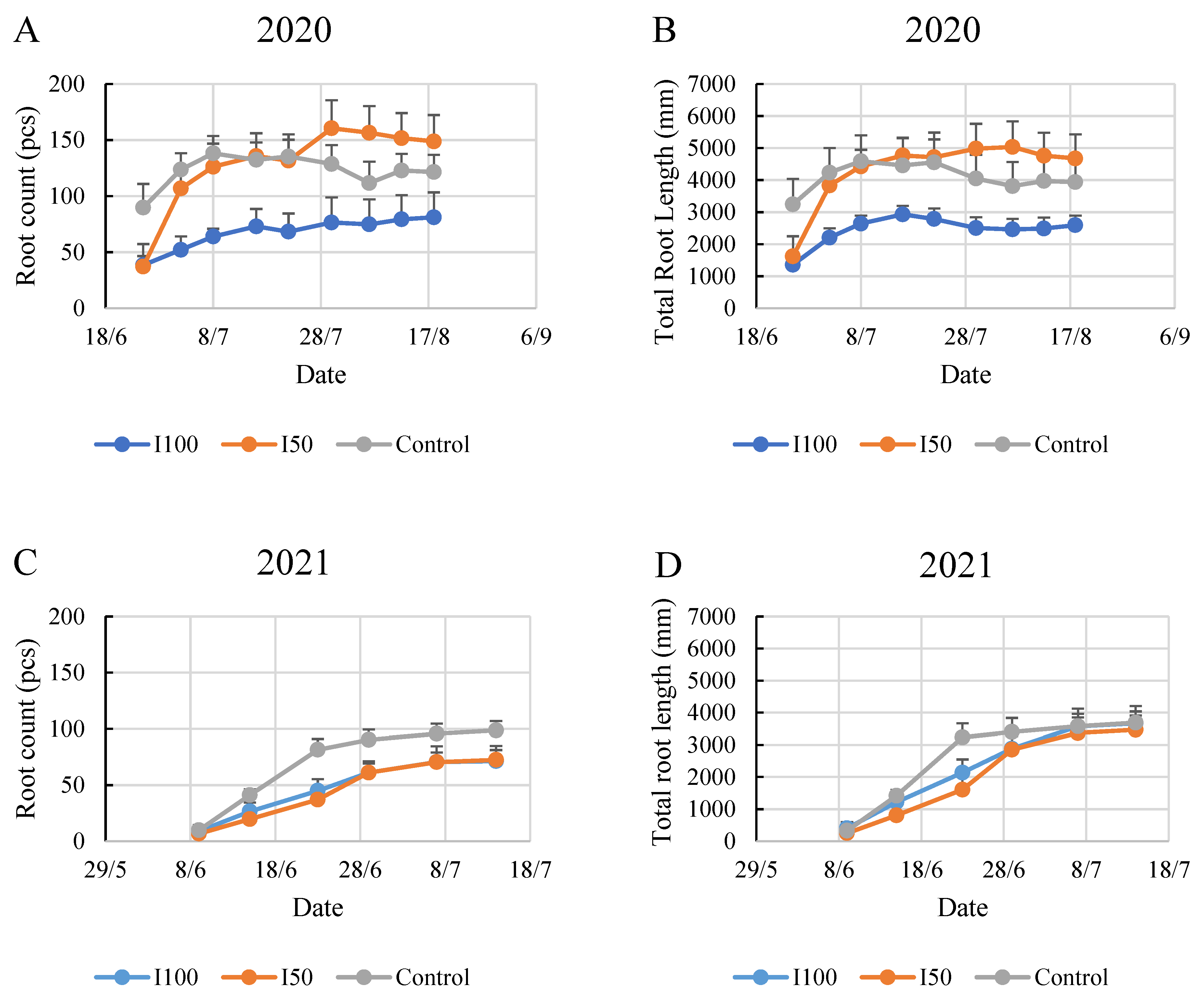
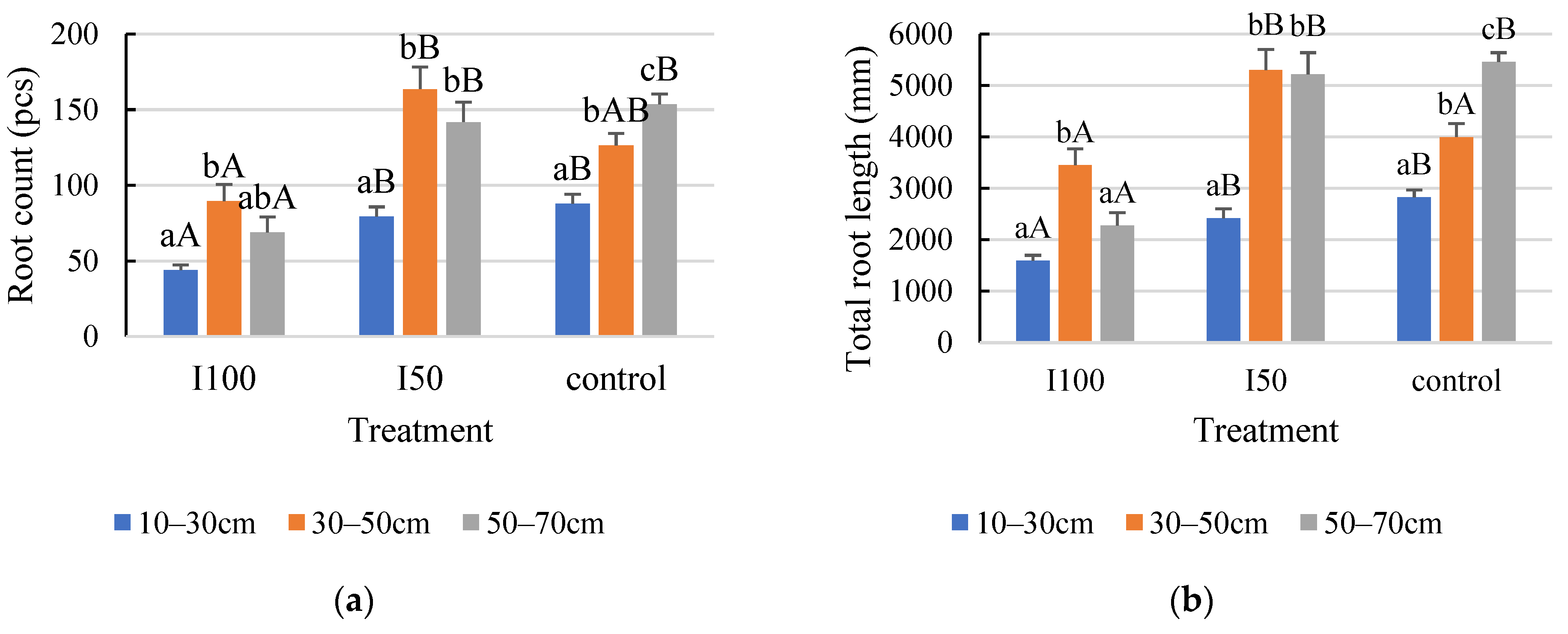
2. Results
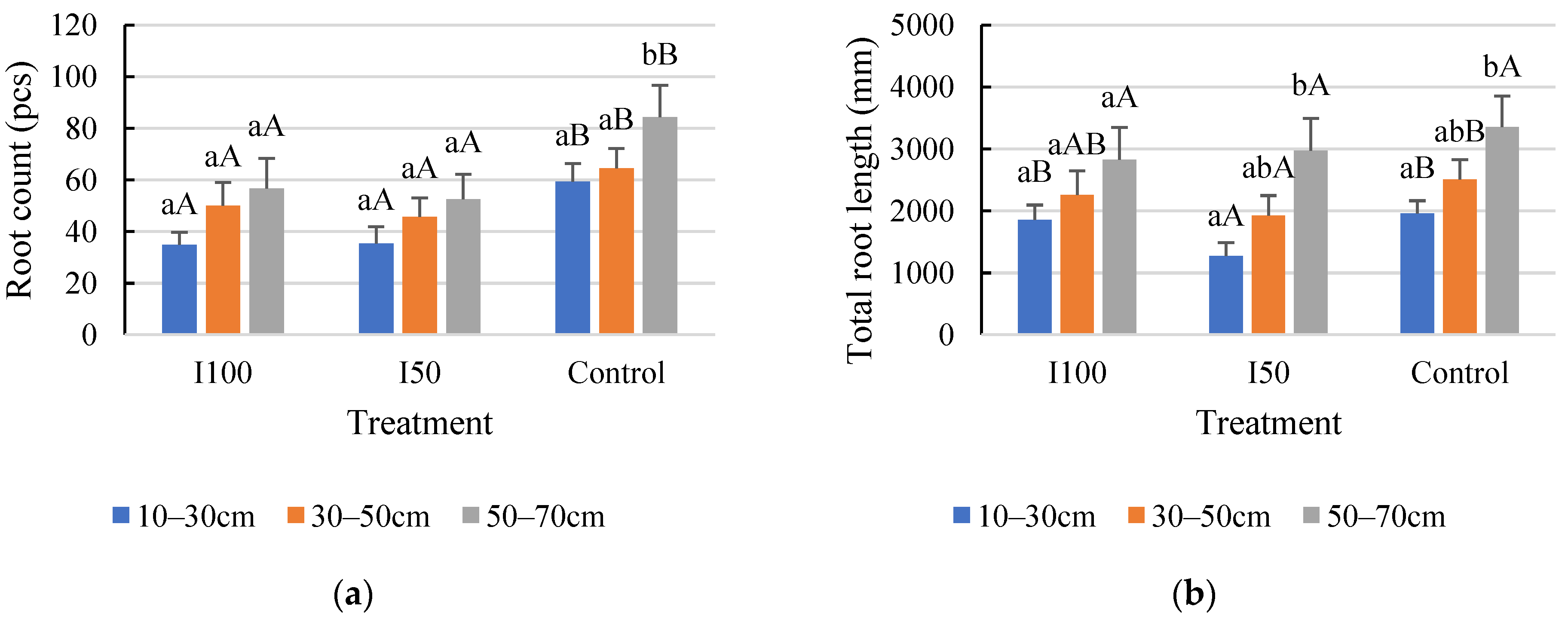
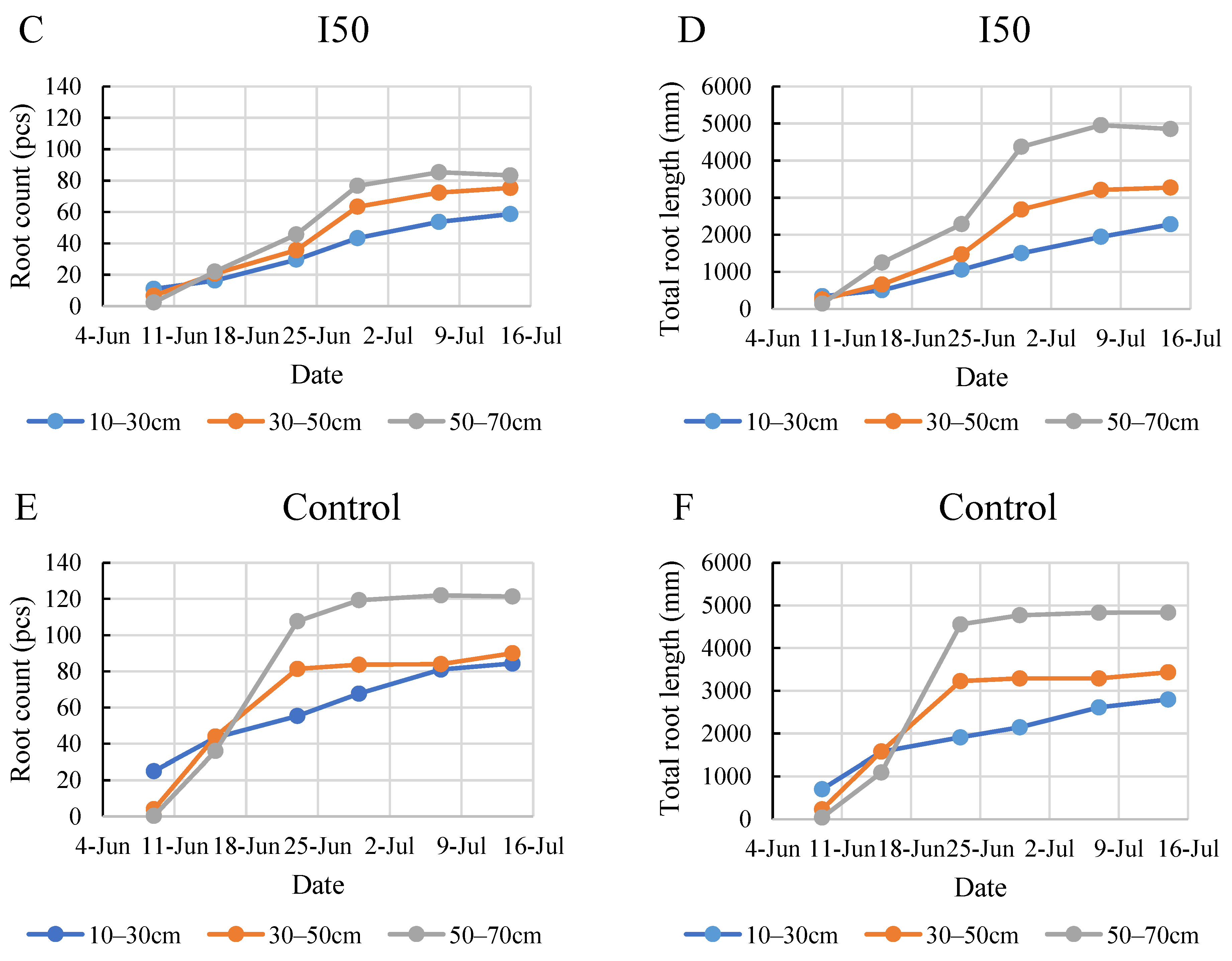
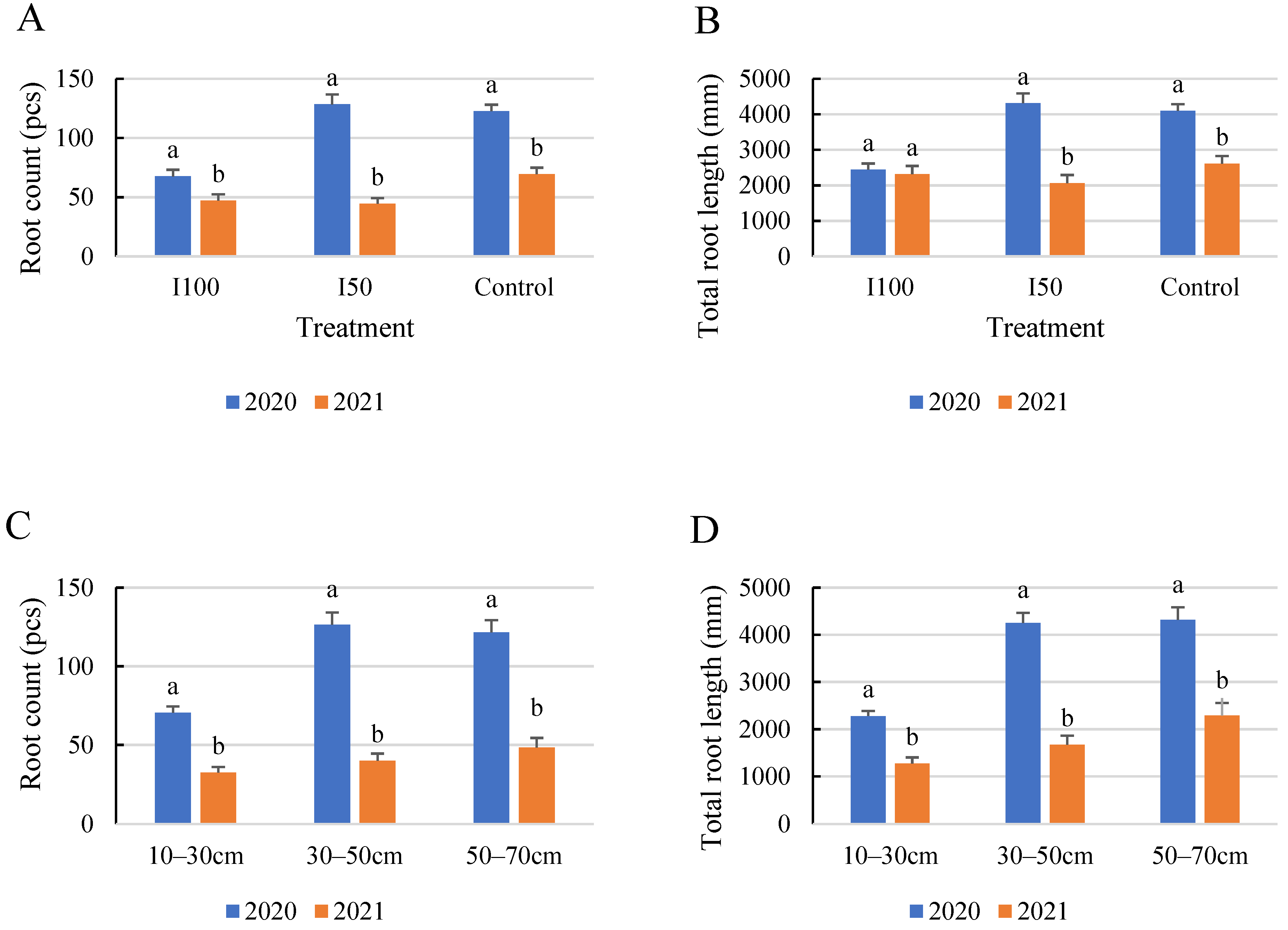
2.1. General Results Regarding Root Count and Root Length
2.2. Evaluation of the Time Scale for the Monitored Root Zone
2.3. Evaluation of the Layer Scale for the Monitored Root Zone
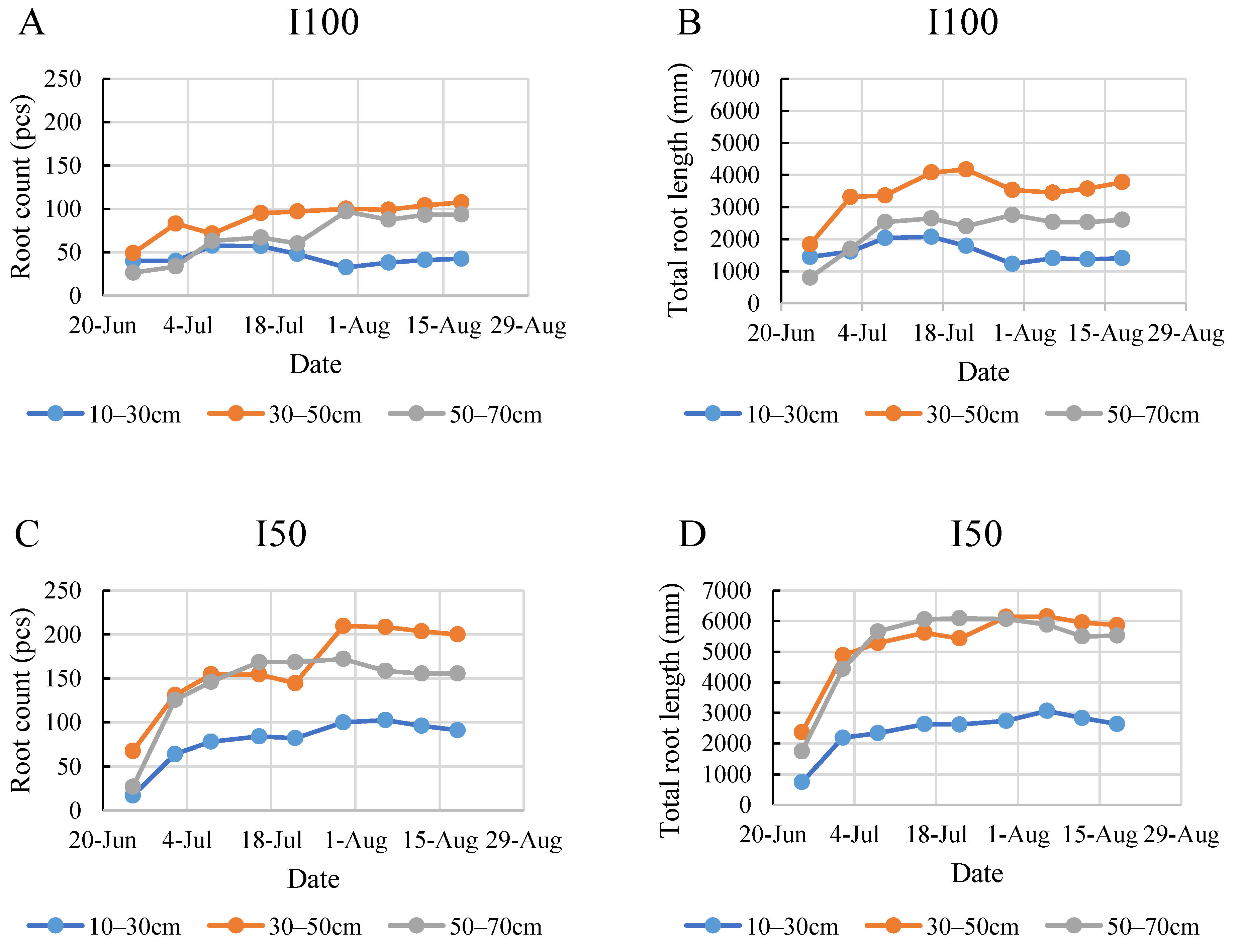
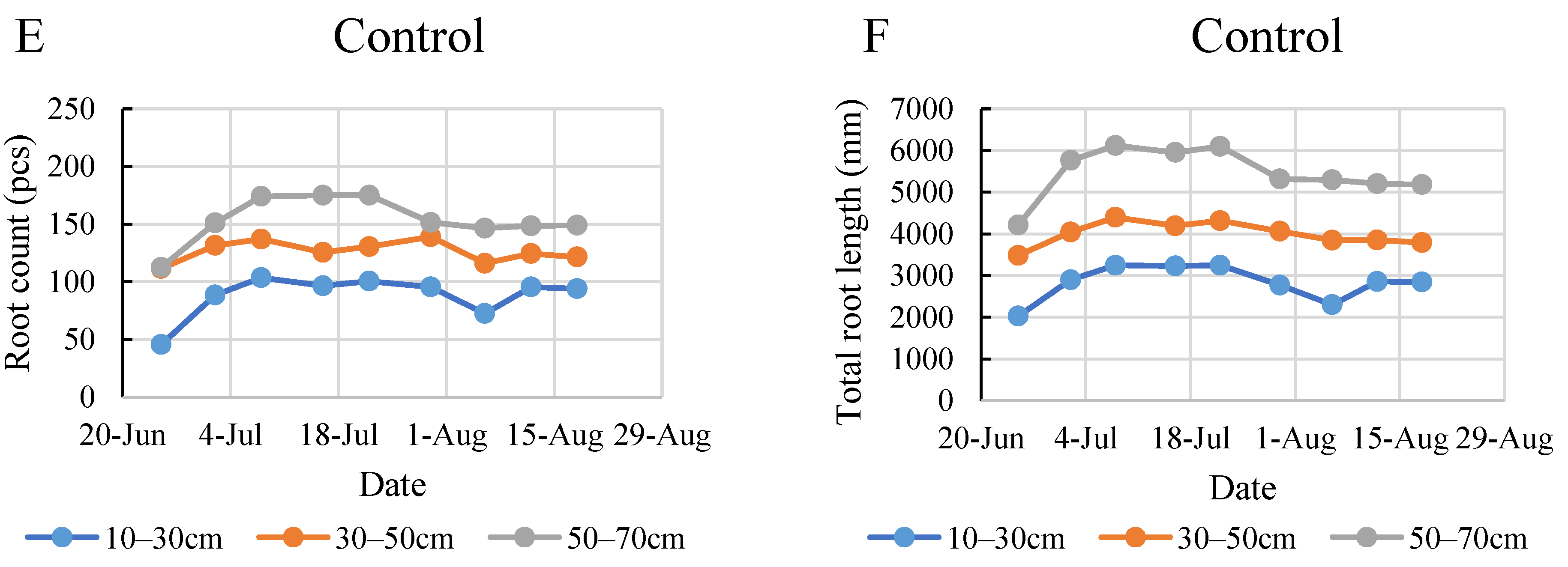
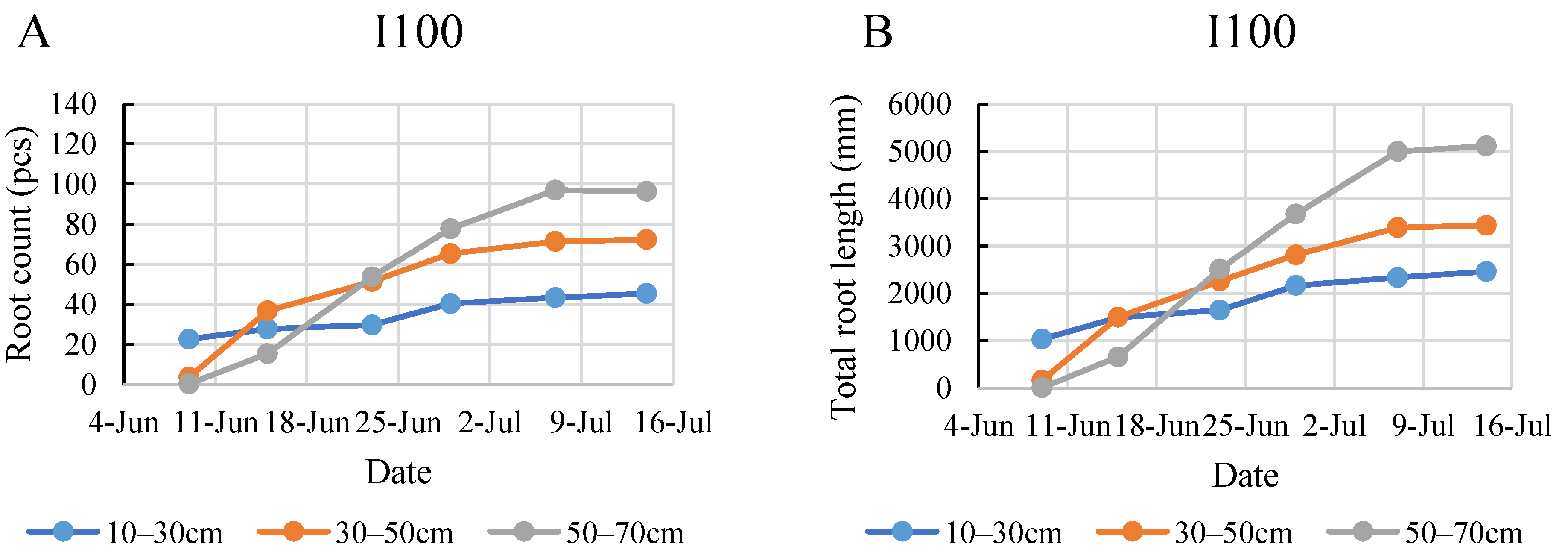
2.4. Root Development during the Monitoring Period
2.5. Comparison of the Root Development in the Two Years
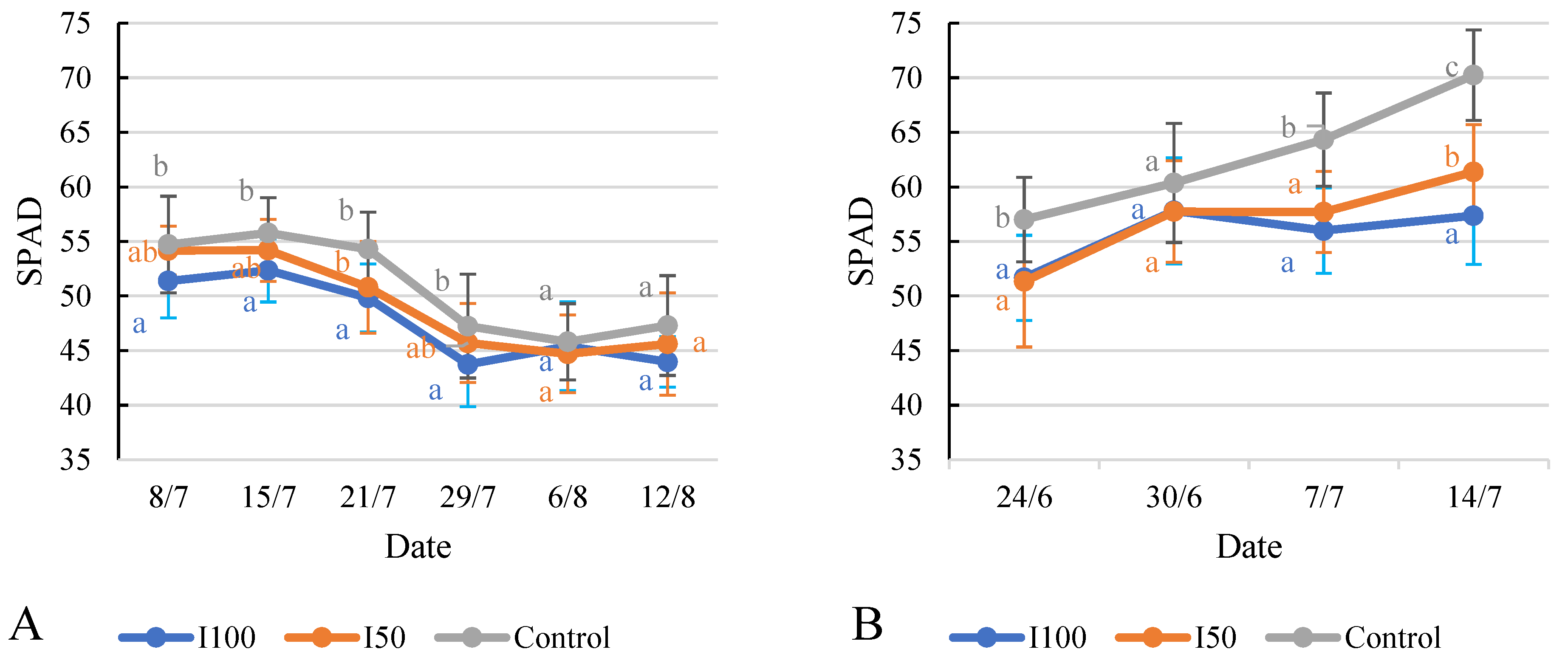
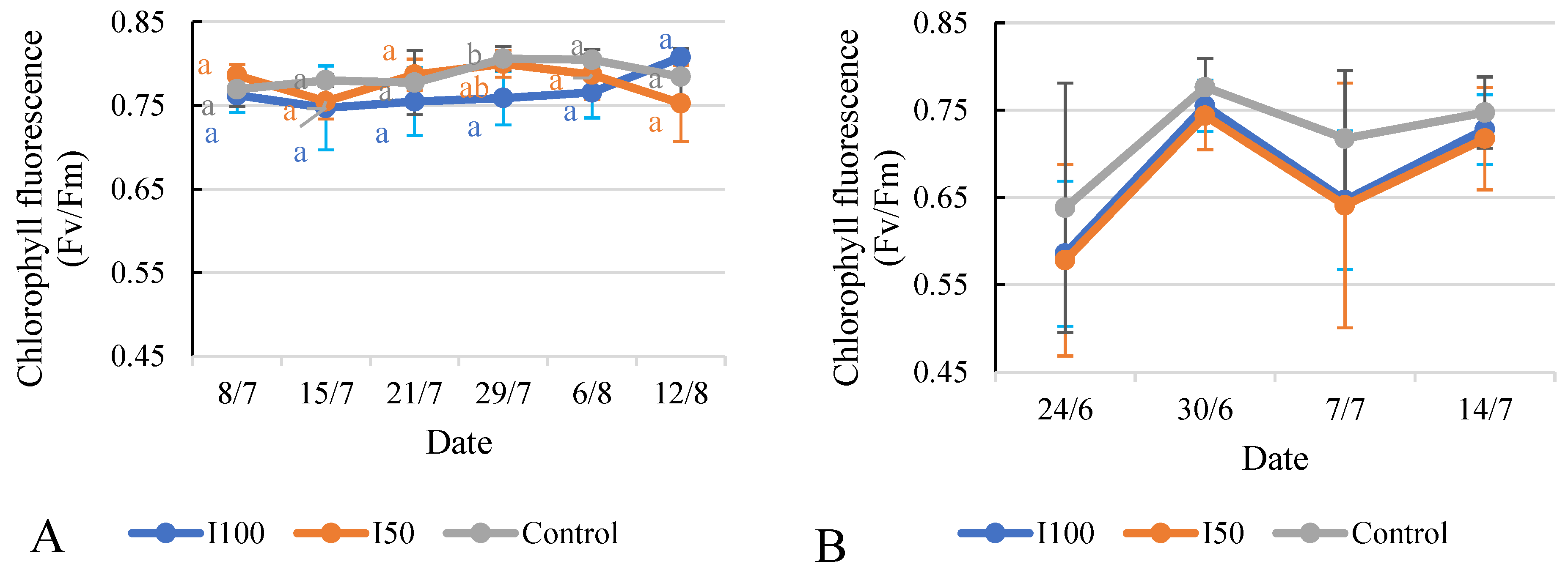
2.6. Effect of Different Treatments on Relative Chlorophyll Content (SPAD) and Chlorophyll Fluorescence (Fv/Fm)
3. Discussion
4. Materials and Methods
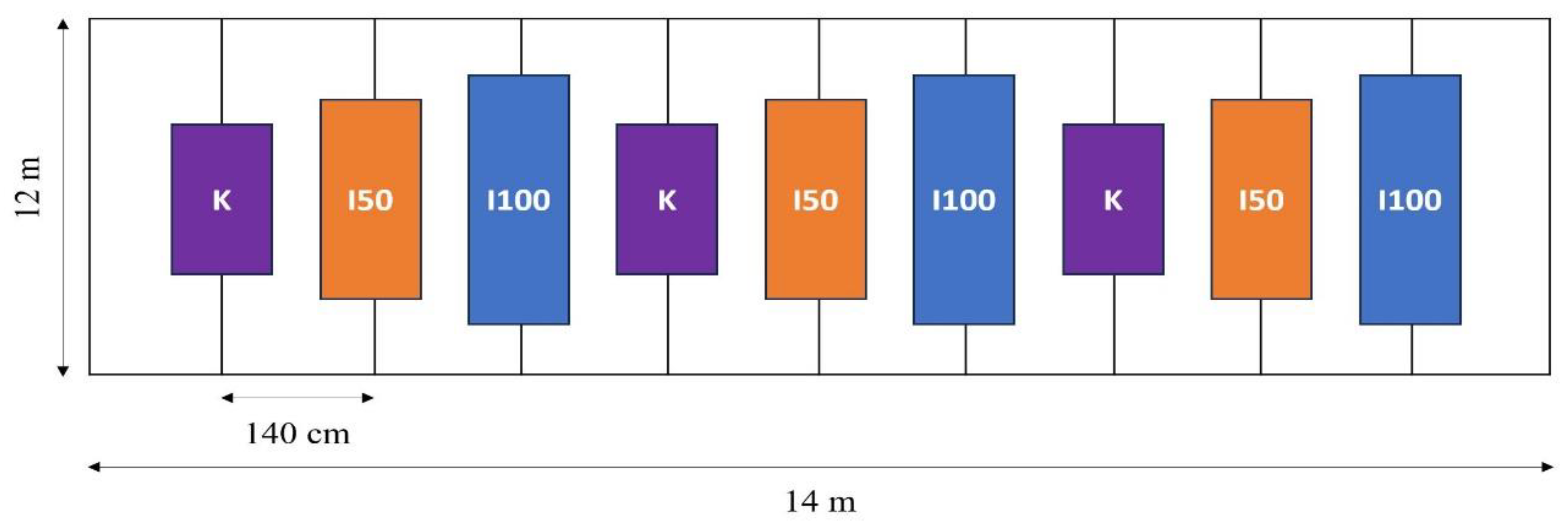
4.1. Plant Material and Experimental Set-Up
4.2. Image Acquisition
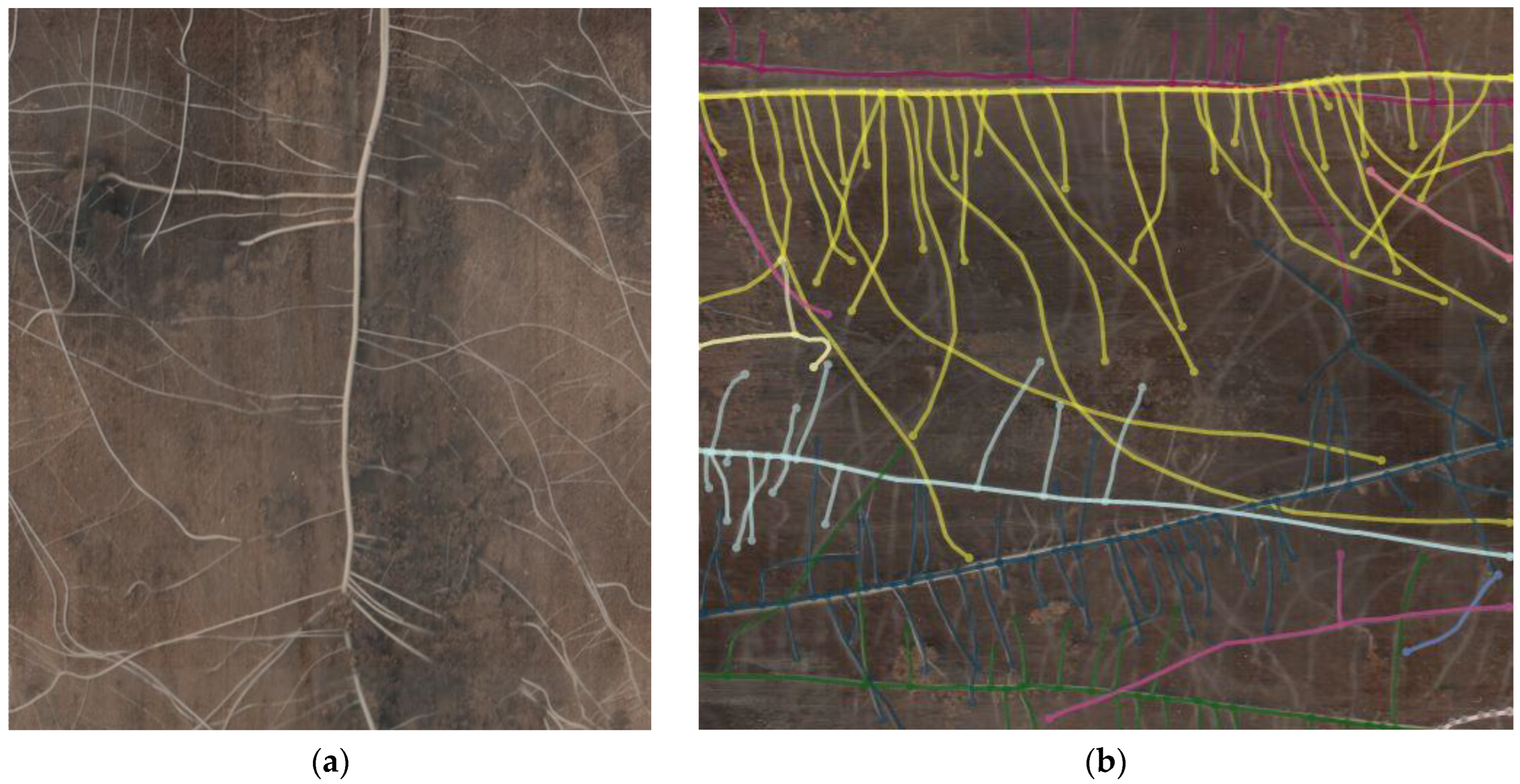
4.3. Image Processing
4.4. Relative Chlorophyll Content and Photosynthetic Activity
4.5. Statistical Evaluation
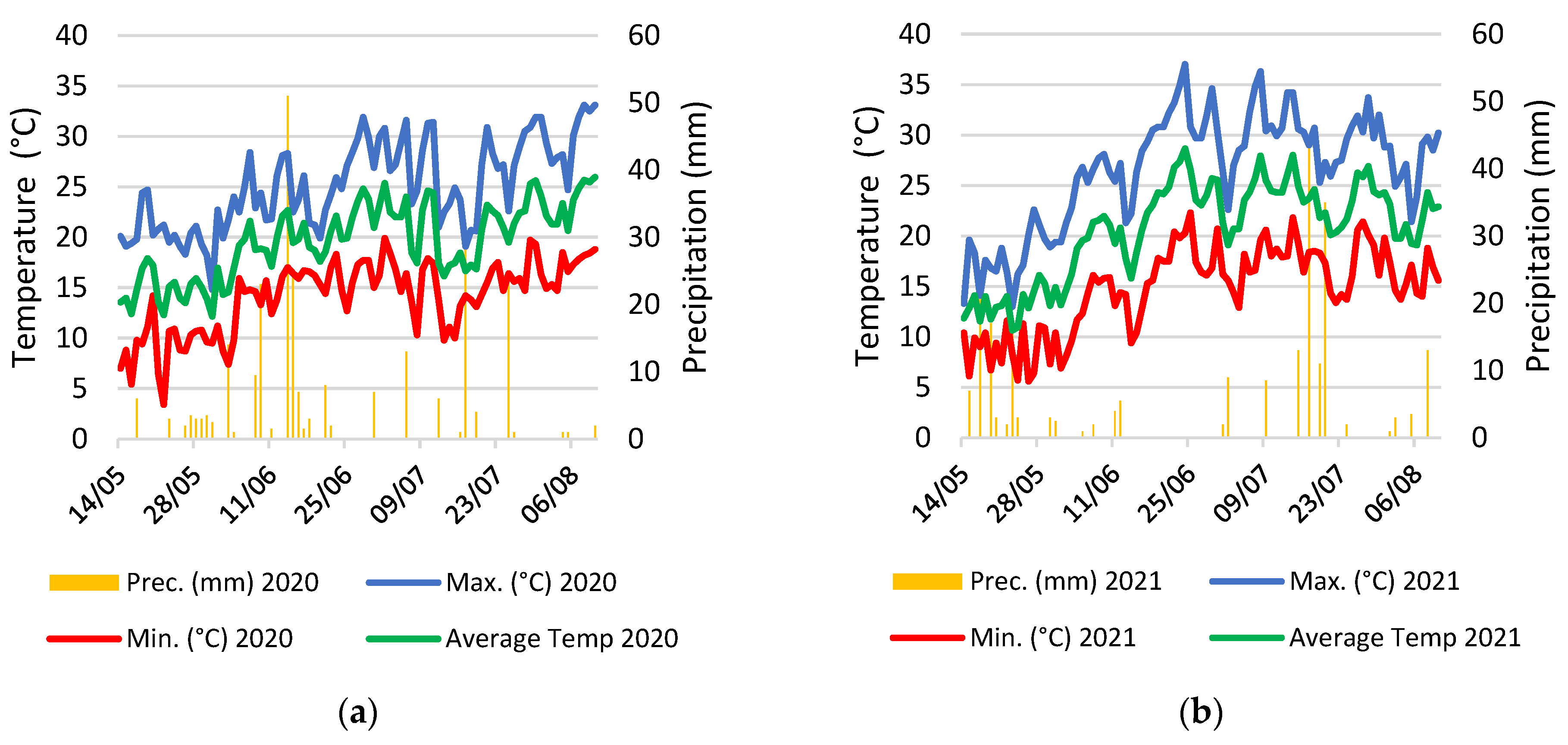
4.6. Meteorological Data
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Salekdeh, G.H.; Reynolds, M.; Bennett, J.; Boyer, J. Conceptual Framework for Drought Phenotyping during Molecular Breeding. Trends Plant Sci. 2009, 14, 488–496. [Google Scholar] [CrossRef]
- Wang, X.; Jiang, D.; Lang, X. Future Extreme Climate Changes Linked to Global Warming Intensity. Sci. Bull. 2017, 62, 1673–1680. [Google Scholar] [CrossRef] [PubMed]
- IPCC. Climate Change 2014: Synthesis Report. Contribution of Working Groups I, II and III to the Fifth Assessment Report of the Intergovernmental Panel on Climate Change; Core Writing Team, Pachauri, R.K., Meyer, L.A., Eds.; IPCC: Geneva, Switzerland, 2014; 151p. [Google Scholar]
- McDowell, N.G.; Beerling, D.J.; Breshears, D.D.; Fisher, R.A.; Raffa, K.F.; Stitt, M. The Interdependence of Mechanisms Underlying Climate-Driven Vegetation Mortality. Trends Ecol. Evol. 2011, 26, 523–532. [Google Scholar] [CrossRef] [PubMed]
- Bréda, N.; Huc, R.; Granier, A.; Dreyer, E. Temperate Forest Trees and Stands under Severe Drought: A Review of Ecophysiological Responses, Adaptation Processes and Long-Term Consequences. Ann. For. Sci. 2006, 63, 625–644. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Neményi, A.; Bőcs, A.; Pék, Z.; Helyes, L. Physiological Factors and Their Relationship with the Productivity of Processing Tomato under Different Water Supplies. Water 2019, 11, 586. [Google Scholar] [CrossRef]
- Agbemafle, R.; Danso Owusu-Sekyere, J.; Bart-Plange, A.; Otchere, J.K.; Owusu-Sekyere, J.; Bart-Plange, A.; Otchere, J. Effect of Deficit Irrigation and Storage on Physicochemical Quality of Tomato (Lycopersicon esculentum Mill. Var. Pechtomech) Climate Change View Project Wtaer Pollution View Project Effect of Deficit Irrigation and Storage on Physicochemical Quality of Tomato (Lycopersicon Esculentum Mill. Var. Pechtomech). Food Sci. Qual. Manag. 2014, 34, 113–120. [Google Scholar]
- Liu, J.; Hu, T.; Feng, P.; Yao, D.; Gao, F.; Hong, X. Effect of Potassium Fertilization during Fruit Development on Tomato Quality, Potassium Uptake, Water and Potassium Use Efficiency under Deficit Irrigation Regime. Agric. Water Manag. 2021, 250, 106831. [Google Scholar] [CrossRef]
- Wang, X.; Yun, J.; Shi, P.; Li, Z.; Li, P.; Xing, Y. Root Growth, Fruit Yield and Water Use Efficiency of Greenhouse Grown Tomato Under Different Irrigation Regimes and Nitrogen Levels. J. Plant Growth Regul. 2019, 38, 400–415. [Google Scholar] [CrossRef]
- Patanè, C.; Tringali, S.; Sortino, O. Effects of Deficit Irrigation on Biomass, Yield, Water Productivity and Fruit Quality of Processing Tomato under Semi-Arid Mediterranean Climate Conditions. Sci. Hortic. 2011, 129, 590–596. [Google Scholar] [CrossRef]
- Takács, S.; Pék, Z.; Csányi, D.; Daood, H.G.; Szuvandzsiev, P.; Palotás, G.; Helyes, L. Influence of Water Stress Levels on the Yield and Lycopene Content of Tomato. Water 2020, 12, 2165. [Google Scholar] [CrossRef]
- Célia De Matos Pires, R.; Furlani, P.R.; Ribeiro, R.V.; Junior, D.B.; Sakai, E.; Lourenção, A.L.; Neto, A.T. Irrigation Frequency and Substrate Volume Effects in the Growth and Yield of Tomato Plants under Greenhouse Conditions. Sci. Agric. 2011, 68, 400–405. [Google Scholar] [CrossRef]
- Lahoz, I.; Pérez-de-Castro, A.; Valcárcel, M.; Macua, J.I.; Beltrán, J.; Roselló, S.; Cebolla-Cornejo, J. Effect of Water Deficit on the Agronomical Performance and Quality of Processing Tomato. Sci. Hortic. 2016, 200, 55–65. [Google Scholar] [CrossRef]
- EC. EU Agricultural Outlook for Markets, Income and Environment, 2021–2031; European Commission, DG Agriculture and Rural Development: Brussels, Belgium, 2021. [Google Scholar]
- George, S.; Jatoi, S.A.; Siddiqui, S.U. Genotypic differences against PEG simulated drought stress in tomato. Pak. J. Bot. 2013, 45, 1551–1556. [Google Scholar]
- Suminar, E.; Budiarto, R.; Nuraini, A.; Mubarok, S.; Ezura, H. Morpho-physiological responses of iaa9 tomato mutants to different levels of PEG simulated drought stress. Biodiversitas J. Biol. Divers. 2022, 23, 3115–3126. [Google Scholar] [CrossRef]
- Bogoutdinova, L.R.; Baranova, E.N.; Baranova, G.B.; Kononenko, N.V.; Lazareva, E.M.; Smirnova, E.A.; Khaliluev, M.R. Morpho-Biological and Cytological Characterization of Tomato Roots (Solanum lycopersicum L., cv. Rekordsmen) Regenerated under NaCl Salinity in vitro. Cell Tiss. Biol. 2020, 14, 228–242. [Google Scholar] [CrossRef]
- Bogoutdinova, L.R.; Baranova, E.N.; Kononenko, N.V.; Chaban, I.A.; Konovalova, L.N.; Gulevich, A.A.; Lazareva, E.M.; Khaliluev, M.R. Characteristics of Root Cells during In Vitro Rhizogenesis under Action of NaCl in Two Tomato Genotypes Differing in Salt Tolerance. Int. J. Plant Biol. 2023, 14, 104–119. [Google Scholar] [CrossRef]
- Das Choudhury, S.; Samal, A.; Awada, T. Leveraging Image Analysis for High-Throughput Plant Phenotyping. Front. Plant Sci. 2019, 10, 508. [Google Scholar] [CrossRef]
- Atefi, A.; Ge, Y.; Pitla, S.; Schnable, J. Robotic Technologies for High-Throughput Plant Phenotyping: Contemporary Reviews and Future Perspectives. Front. Plant Sci. 2021, 12, 611940. [Google Scholar] [CrossRef]
- Wasaya, A.; Zhang, X.; Fang, Q.; Yan, Z. Root Phenotyping for Drought Tolerance: A Review. Agronomy 2018, 8, 241. [Google Scholar] [CrossRef]
- Donnelly, A.; Yu, R.; Rehberg, C.; Meyer, G.; Young, E.B. Leaf Chlorophyll Estimates of Temperate Deciduous Shrubs during Autumn Senescence Using a SPAD-502 Meter and Calibration with Extracted Chlorophyll. Ann. For. Sci. 2020, 77, 1–12. [Google Scholar] [CrossRef]
- Cassab, G.I.; Eapen, D.; Campos, M.E. Root hydrotropism: An update. Am. J. Bot. 2013, 100, 14–24. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, N.; Yamazaki, Y.; Kobayashi, A.; Higashitani, A.; Takahashi, H. Hydrotropism Interacts with Gravitropism by Degrading Amyloplasts in Seedling Roots of Arabidopsis and Radish. Plant Physiol. 2003, 132, 805–810. [Google Scholar] [CrossRef] [PubMed]
- Comas, L.H.; Becker, S.R.; Cruz, V.M.V.; Byrne, P.F.; Dierig, D.A. Root Traits Contributing to Plant Productivity under Drought. Front. Plant Sci. 2013, 4, 442. [Google Scholar] [CrossRef] [PubMed]
- Mao, B.; Takahashi, H.; Takahashi, H.; Fujii, N. Diversity of Root Hydrotropism among Natural Variants of Arabidopsis Thaliana. J. Plant Res. 2022, 135, 799–808. [Google Scholar] [CrossRef]
- Amtmann, A.; Bennett, M.J.; Henry, A. Root Phenotypes for the Future. Plant Cell Environ. 2022, 45, 595–601. [Google Scholar] [CrossRef]
- Hernandez-Espinoza, L.H.; Barrios-Masias, F.H. Physiological and Anatomical Changes in Tomato Roots in Response to Low Water Stress. Sci. Hortic. 2020, 265, 109208. [Google Scholar] [CrossRef]
- Yu, G.R.; Zhuang, J.; Nakayama, K.; Jin, Y. Root Water Uptake and Profile Soil Water as Affected by Vertical Root Distribution. Plant Ecol. 2007, 189, 15–30. [Google Scholar] [CrossRef]
- Zakaria, N.I.; Ismail, M.R.; Awang, Y.; Megat Wahab, P.E.; Berahim, Z. Effect of Root Restriction on the Growth, Photosynthesis Rate, and Source and Sink Relationship of Chilli (Capsicum annuum L.) Grown in Soilless Culture. Biomed Res. Int. 2020, 2020, 2706937. [Google Scholar] [CrossRef]
- Li, Y.; Niu, W.; Cao, X.; Wang, J.; Zhang, M.; Duan, X.; Zhang, Z. Effect of Soil Aeration on Root Morphology and Photosynthetic Characteristics of Potted Tomato Plants (Solanum lycopersicum) at Different NaCl Salinity Levels. BMC Plant Biol. 2019, 19, 331. [Google Scholar] [CrossRef]
- Murchie, E.H.; Lawson, T. Chlorophyll Fluorescence Analysis: A Guide to Good Practice and Understanding Some New Applications. J. Exp. Bot. 2013, 64, 3983–3998. [Google Scholar] [CrossRef]
- Kalaji, H.M.; Jajoo, A.; Oukarroum, A.; Brestic, M.; Zivcak, M.; Samborska, I.A.; Cetner, M.D.; Łukasik, I.; Goltsev, V.; Ladle, R.J. Chlorophyll a Fluorescence as a Tool to Monitor Physiological Status of Plants under Abiotic Stress Conditions. Acta Physiol. Plant 2016, 38, 102. [Google Scholar] [CrossRef]
- Scholander, P.F.; Bradstreet, E.D.; Hemmingsen, E.A.; Hammel, H.T. Sap Pressure in Vascular Plants. Science 1965, 148, 339–346. [Google Scholar] [CrossRef] [PubMed]
- Singh, C.; Rai, P.; BK, R. Genetic Engineering for Drought Resistance in Rice. Agric. Res. Technol. 2017, 11. [Google Scholar] [CrossRef]
- Katerji, N.; Campi, P.; Mastrorilli, M. Productivity, evapotranspiration, and water use efficiency of corn and tomato crops simulated by AquaCrop under contrasting water stress conditions in the Mediterranean region. Agric. Water Manag. 2013, 130, 14–26. [Google Scholar] [CrossRef]
- Nemeskéri, E.; Helyes, L. Physiological Responses of Selected Vegetable Crop Species to Water Stress. Agronomy 2019, 9, 447. [Google Scholar] [CrossRef]
- Nicolas, M.E.; Lambers, H.; Simpson, R.J.; Dalling, M.J. Effect of Drought on Metabolism and Partitioning of Carbon in Two Wheat Varieties Differing in Drought-Tolerance. Ann. Bot. 1985, 55, 727–742. [Google Scholar] [CrossRef]
- Lee, D.K.; Jung, H.; Jang, G.; Jeong, J.S.; Kim, Y.S.; Ha, S.H.; Do Choi, Y.; Kim, J.K. Overexpression of the OsERF71 Transcription Factor Alters Rice Root Structure and Drought Resistance. Plant Physiol. 2016, 172, 575–588. [Google Scholar] [CrossRef]
- Fara, S.J.; Teixeira Delazari, F.; Silva Gomes, R.; Araújo, W.L.; da Silva, D.J.H. Stomata Opening and Productiveness Response of Fresh Market Tomato under Different Irrigation Intervals. Sci. Hortic. 2019, 255, 86–95. [Google Scholar] [CrossRef]
- Vadez, V.; Rao, J.S.; Bhatnagar-Mathur, P.; Sharma, K.K. DREB1A Promotes Root Development in Deep Soil Layers and Increases Water Extraction under Water Stress in Groundnut. Plant Biol. 2013, 15, 45–52. [Google Scholar] [CrossRef]
- Koevoets, I.T.; Venema, J.H.; Elzenga, J.T.M.; Testerink, C. Roots Withstanding Their Environment: Exploiting Root System Architecture Responses to Abiotic Stress to Improve Crop Tolerance. Front. Plant Sci. 2016, 7, 1335. [Google Scholar] [CrossRef]
- Marouelli, W.A.; Silva, W.L.C. Water Tension Thresholds for Processing Tomatoes under Drip Irrigation in Central Brazil. Irrig. Sci. 2007, 25, 411–418. [Google Scholar] [CrossRef]
- He, M.; Dijkstra, F.A. Drought Effect on Plant Nitrogen and Phosphorus: A Meta-Analysis. New Phytol. 2014, 204, 924–931. [Google Scholar] [CrossRef] [PubMed]
- Khapte, P.S.; Kumar, P.; Burman, U.; Kumar, P. Deficit Irrigation in Tomato: Agronomical and Physio-Biochemical Implications. Sci. Hortic. 2019, 248, 256–264. [Google Scholar] [CrossRef]
- Manschadi, A.M.; Christopher, J.; Devoil, P.; Hammer, G.L. The Role of Root Architectural Traits in Adaptation of Wheat to Water-Limited Environments. Funct. Plant Biol. 2006, 33, 823–837. [Google Scholar] [CrossRef] [PubMed]
- Guida, G.; Sellami, M.H.; Mistretta, C.; Oliva, M.; Buonomo, R.; De Mascellis, R.; Patanè, C.; Rouphael, Y.; Albrizio, R.; Giorio, P. Agronomical, Physiological and Fruit Quality Responses of Two Italian Long-Storage Tomato Landraces under Rain-Fed and Full Irrigation Conditions. Agric. Water Manag. 2017, 180, 126–135. [Google Scholar] [CrossRef]
- Li, X.; Šimůnek, J.; Shi, H.; Yan, J.; Peng, Z.; Gong, X. Spatial Distribution of Soil Water, Soil Temperature, and Plant Roots in a Drip-Irrigated Intercropping Field with Plastic Mulch. Eur. J. Agron. 2017, 83, 47–56. [Google Scholar] [CrossRef]
- Schneider, J.R.; Caverzan, A.; Chavarria, G. Water Deficit Stress, ROS Involvement, and Plant Performance. Arch. Agron. Soil Sci. 2019, 65, 1160–1181. [Google Scholar] [CrossRef]
- Lum, M.S.; Hanafi, M.M.; Rafii, Y.M.; Akmar, A.S.N. Effect of drought stress on growth, proline and antioxidant enzyme activities of upland rice. JAPS J. Anim. Plant Sci. 2014, 24, 1487–1493. [Google Scholar]
- Mirzaee, M.; Ghanati, F. Effects of Drought Stress on the Lipid Peroxidation and Antioxidant Enzyme Activities in Two Canola (Brassica napus L.) Cultivars Phytochemistry View Project Signaling Pathways Involved in Secondary Metabolites Biosynthesis View Project. J. Agric. Sci. Technol. 2013, 15, 593–602. [Google Scholar]
- Jha, S.K.; Gao, Y.; Liu, H.; Huang, Z.; Wang, G.; Liang, Y.; Duan, A. Root Development and Water Uptake in Winter Wheat under Different Irrigation Methods and Scheduling for North China. Agric. Water Manag. 2017, 182, 139–150. [Google Scholar] [CrossRef]
- Li, Q.; Dong, B.; Qiao, Y.; Liu, M.; Zhang, J. Root Growth, Available Soil Water, and Water-Use Efficiency of Winter Wheat under Different Irrigation Regimes Applied at Different Growth Stages in North China. Agric. Water Manag. 2010, 97, 1676–1682. [Google Scholar] [CrossRef]
- Sharma, S.P.; Leskovar, D.I.; Crosby, K.M.; Volder, A.; Ibrahim, A.M.H. Root Growth, Yield, and Fruit Quality Responses of Reticulatus and Inodorus Melons (Cucumis melo L.) to Deficit Subsurface Drip Irrigation. Agric. Water Manag. 2014, 136, 75–85. [Google Scholar] [CrossRef]
- Zotarelli, L.; Scholberg, J.M.; Dukes, M.D.; Muñoz-Carpena, R.; Icerman, J. Tomato Yield, Biomass Accumulation, Root Distribution and Irrigation Water Use Efficiency on a Sandy Soil, as Affected by Nitrogen Rate and Irrigation Scheduling. Agric. Water Manag. 2009, 96, 23–34. [Google Scholar] [CrossRef]
- Xu, C.; Tao, H.; Tian, B.; Gao, Y.; Ren, J.; Wang, P. Limited-irrigation improves water use efficiency and soil reservoir capacity through regulating root and canopy growth of winter wheat. Field Crops Res. 2016, 196, 268–275. [Google Scholar] [CrossRef]
- Wu, B.; Zhang, L.; Tian, J.; Zhang, G.; Zhang, W. Fine root dynamics, longevity, and canopy characteristics of cotton under varying water and nitrogen levels. Plant Soil. 2023, 482, 191–209. [Google Scholar] [CrossRef]
- Luo, H.; Xu, H.; Chu, C.; He, F.; Fang, S. High Temperature Can Change Root System Architecture and Intensify Root Interactions of Plant Seedlings. Front. Plant Sci. 2020, 11, 160. [Google Scholar] [CrossRef] [PubMed]
- Chakraborty, P.K.; Banerjee, S.; Nath, R.; Samanta, S. Assessing Congenial Soil Temperature and Its Impact on Root Growth, Grain Yield of Summer Rice under Varying Water Stress Condition in Lower Gangetic Plain of India. J. Saudi Soc. Agric. Sci. 2022, 21, 98–107. [Google Scholar] [CrossRef]
- Karlova, R.; Boer, D.; Hayes, S.; Testerink, C. Root Plasticity under Abiotic Stress. Plant Physiol. 2021, 187, 1057–1070. [Google Scholar] [CrossRef]
- Ogweno, J.O.; Song, X.S.; Hu, W.H.; Shi, K.; Zhou, Y.H.; Yu, J.Q. Detached Leaves of Tomato Differ in Their Photosynthetic Physiological Response to Moderate High and Low Temperature Stress. Sci. Hortic. 2009, 123, 17–22. [Google Scholar] [CrossRef]
- Mishra, K.B.; Iannacone, R.; Petrozza, A.; Mishra, A.; Armentano, N.; La Vecchia, G.; Trtílek, M.; Cellini, F.; Nedbal, L. Engineered Drought Tolerance in Tomato Plants Is Reflected in Chlorophyll Fluorescence Emission. Plant Sci. 2012, 182, 79–86. [Google Scholar] [CrossRef]
- Arbex de Castro Vilas Boas, A.; Page, D.; Giovinazzo, R.; Bertin, N.; Fanciullino, A.-L. Combined Effects of Irrigation Regime, Genotype, and Harvest Stage Determine Tomato Fruit Quality and Aptitude for Processing into Puree. Front. Plant Sci. 2017, 8, 1725. [Google Scholar] [CrossRef] [PubMed]
- Fanciullino, A.L.; Sinkora, M.; Breniere, T.; Conejero, G.; Page, D.; Bertin, N. Improving crop sustainability and fresh and processed fruit quality through integrated analyses along the food chain. In Proceedings of the XXXI International Horticultural Congress (IHC2022): International Symposium on Integrative Approaches to Product Quality in 1353, Angers, France, 14–20 August 2022; pp. 163–174. [Google Scholar]
- Takács, S.; Csengeri, E.; Pék, Z.; Bíró, T.; Szuvandzsiev, P.; Palotás, G.; Helyes, L. Performance Evaluation of Aquacrop Model in Processing Tomato Biomass, Fruit Yield and Water Stress Indicator Modelling. Water 2021, 13, 3587. [Google Scholar] [CrossRef]
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
M’hamdi, O.; Égei, M.; Pék, Z.; Ilahy, R.; Nemeskéri, E.; Helyes, L.; Takács, S. Root Development Monitoring under Different Water Supply Levels in Processing Tomato Plants. Plants 2023, 12, 3517. https://doi.org/10.3390/plants12203517
M’hamdi O, Égei M, Pék Z, Ilahy R, Nemeskéri E, Helyes L, Takács S. Root Development Monitoring under Different Water Supply Levels in Processing Tomato Plants. Plants. 2023; 12(20):3517. https://doi.org/10.3390/plants12203517
Chicago/Turabian StyleM’hamdi, Oussama, Márton Égei, Zoltán Pék, Riadh Ilahy, Eszter Nemeskéri, Lajos Helyes, and Sándor Takács. 2023. "Root Development Monitoring under Different Water Supply Levels in Processing Tomato Plants" Plants 12, no. 20: 3517. https://doi.org/10.3390/plants12203517
APA StyleM’hamdi, O., Égei, M., Pék, Z., Ilahy, R., Nemeskéri, E., Helyes, L., & Takács, S. (2023). Root Development Monitoring under Different Water Supply Levels in Processing Tomato Plants. Plants, 12(20), 3517. https://doi.org/10.3390/plants12203517