
Arecaceae Seeds Constitute a Healthy Source of Fatty Acids and Phenolic Compounds
 , , and
, , and
Abstract
1. Introduction
2. Results
2.1. Fatty Acids
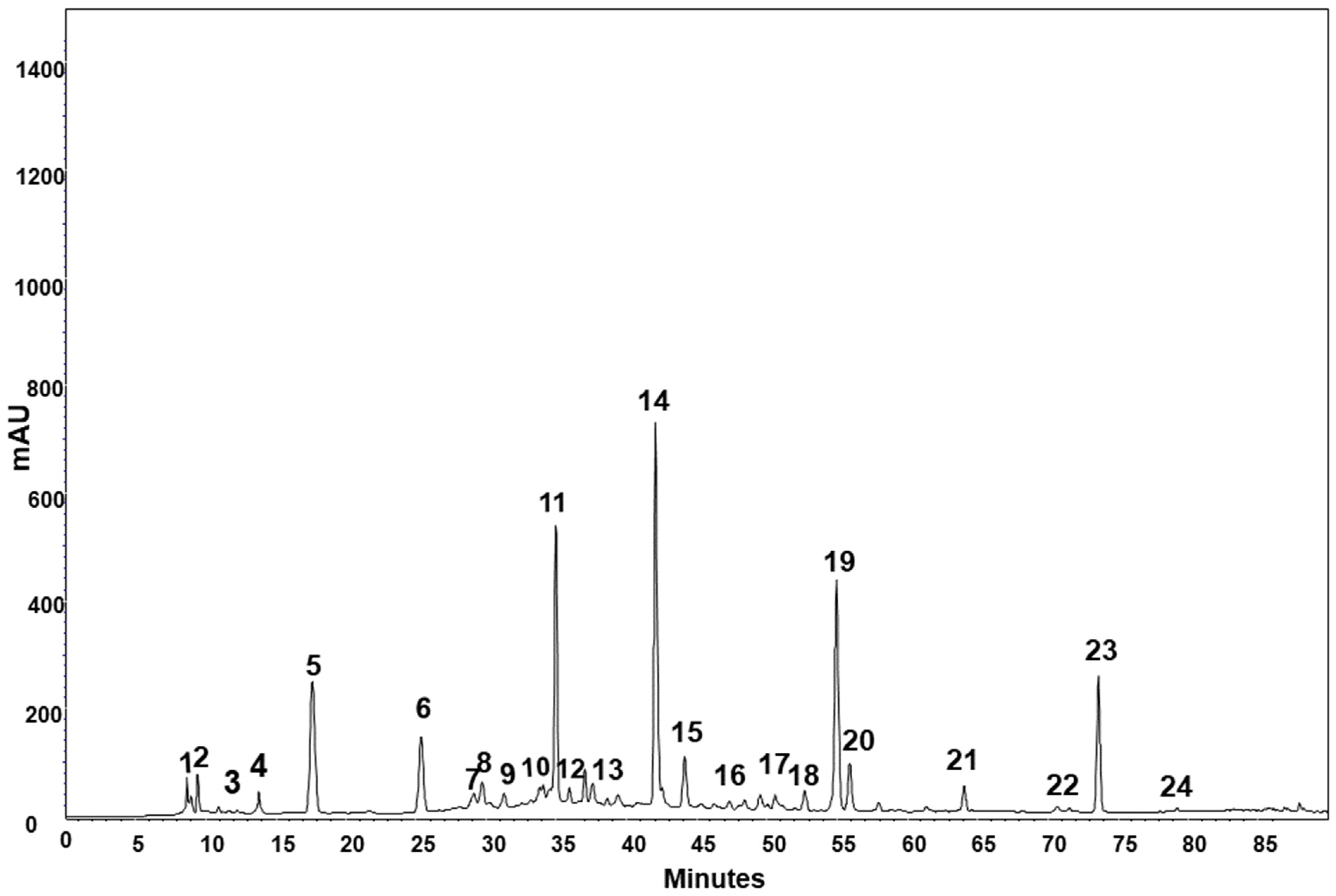
2.2. Phenolic Compounds
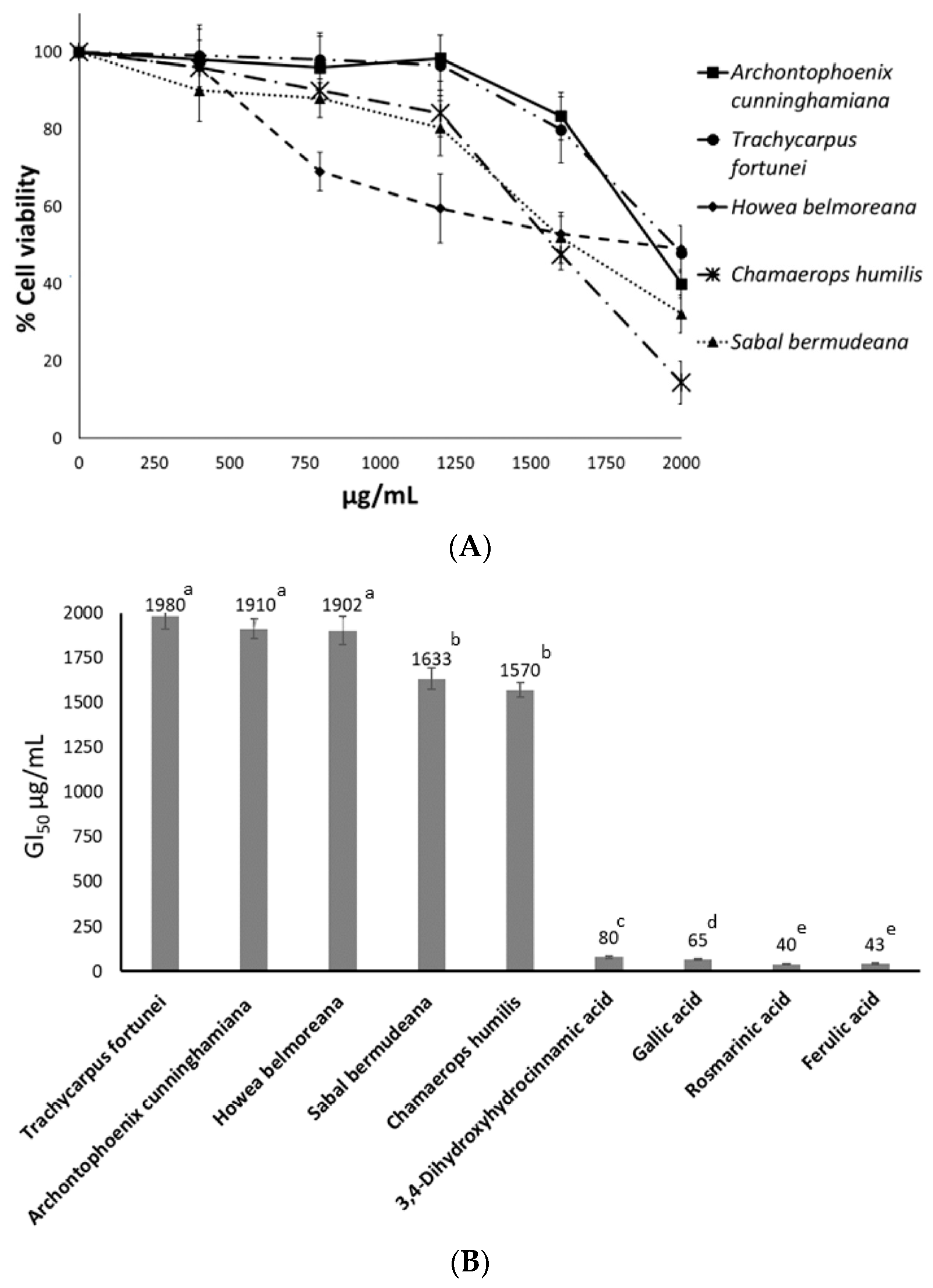
2.3. Antiproliferative Activity against HT-29 Cells
3. Discussion
3.1. Fatty Acids Content
3.2. Phenolic Compounds
3.3. Antiproliferative Activity of Seed Extracts against HT-29 Cells
4. Materials and Methods
4.1. Samples and Chemicals
4.2. FA Analysis
4.3. Identification and Quantification of Phenolic Compounds
4.4. Antiproliferative Assays of Phenolic Extracts on the HT-29 Cell Line
4.5. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Barcelos, E.; Rios, S.D.A.; Cunha, R.N.V.; lopes, R.; Motoike, S.Y.; Ebabiychuk, E.; Eskirycz, A.; Kushnir, S. Oil palm natural diversity and the potential for yield improvement. Front. Plant Sci. 2015, 6, 190. [Google Scholar] [CrossRef] [PubMed]
- Montoya, C.; Cochard, B.; Flori, A.; Cros, D.; Lopes, R.; Cuellar, T.; Espeout, S.; Syaputra, I.; Villeneuve, P.; Pina, M.; et al. Genetic architecture of palm oil fatty acid composition in cultivated oil palm (Elaeis guineensis Jaqc.) compared to its wild relative E. oleifera (H.B.K) Cortés. PLoS ONE 2014, 9, e95412. [Google Scholar] [CrossRef] [PubMed]
- Nehdi, I.A.; Sbihi, H.M.; Tan, C.P.; Rashid, U.; Al-Resayes, S.I. Chemical Composition of Date Palm (Phoenix dactylifera L.) Seed Oil from Six Saudi Arabian Cultivars. J. Food Sci. 2018, 83, 624–630. [Google Scholar] [CrossRef] [PubMed]
- Mancini, A.; Imperlini, E.; Nigro, E.; Montagnese, C.; Daniele, A.; Orrù, S.; Buono, P. Biological and Nutritional Properties of Palm Oil and Palmitic Acid: Effects on Health. Molecules 2015, 20, 17339–17361. [Google Scholar] [CrossRef]
- Guerin, C.; Serret, J.; Montúfar, R.; Vaissayre, V.; Bastos-Siqueira, A.; Durand-Gasselin, T.; Tregear, J.; Morcillo, F.; Dussert, S. Palm seed and fruit lipid composition: Phylogenetic and ecological perspectives. Ann. Bot. 2020, 125, 157–172. [Google Scholar] [CrossRef]
- García-González, C.; Salomón-Torres, R.; Montero-Alpírez, G.; Chávez-Velasco, D.; Ortiz-Uribe, N.; Ruiz-Ortiz, N.S.; Coronado-Ortega, M.A.; Curiel-Alvarez, M.A. Effect of pollen sources on yield oil extraction and fatty acid profile of the date seed (Phoenix dactylifera L.) cultivar Medjool from Mexico. Grasas y Aceites 2019, 70, 315. [Google Scholar] [CrossRef]
- Litchfield, C. Taxonomic patterns in the fat content, fatty acid composition, and triglyceride composition of Palmae seeds. Chem. Phys. Lipids 1970, 4, 96–103. [Google Scholar] [CrossRef]
- Mokbli, S.; Sbihi, H.M.; Nehdi, I.A.; Romdhani-Younes, M.; Tan, C.P.; Al-Resayes, S.I. Characteristics of Chamaerops humilis L. var. humilis seed oil and study of the oxidative stability by blending with soybean oil. J. Food Sci. Technol. 2018, 55, 2170–2179. [Google Scholar] [CrossRef]
- Ndukwe, G.; Ugboaja, A. Acid-Base Catalyzed Transesterification of Archontopheonix cunninghamiana (Bangalow Palm) Seed Oil. J. Appl. Sci. Environ. Manag. 2019, 23, 1717. [Google Scholar] [CrossRef]
- Opute, F.I. The seed lipids of the palm family. J. Am. Oil Chem. Soc. 1979, 56, 528–530. [Google Scholar] [CrossRef]
- Daulatabad, C.D.; Ankalgi, R.E. Component Fatty Acids of Some Indian Seed Oils. Fette Seifen Anstrichm. 1983, 85, 404–406. [Google Scholar] [CrossRef]
- Kotraswamy, K.M.; Shaikh, I.N.; Ankalgi, R.F.; Ankalgi, S.R.; Shaikh, I.N.; Bagwan, U.F. Studies on industrially important Guttiferae and Palmae family. J. Pharmacogn. Phytochem. 2016, 5, 194. [Google Scholar]
- Tutunchi, H.; Ostadrahimi, A.; Saghafi-Asl, M. The Effects of Diets Enriched in Monounsaturated Oleic Acid on the Management and Prevention of Obesity: A Systematic Review of Human Intervention Studies. Adv. Nutr. Int. Rev. J. 2020, 11, 864–877. [Google Scholar] [CrossRef] [PubMed]
- Riley, T.; Petersen, K.; Kris-Etherton, P. Health aspects of high-oleic oils. In High Oleic Oils; Flider, F.J., Ed.; AOCS Press: Urbana, IL, USA, 2022; pp. 201–243. [Google Scholar]
- Huang, L.; Gao, L.; Chen, C. Role of Medium-Chain Fatty Acids in Healthy Metabolism: A Clinical Perspective. Trends Endocrinol. Metab. 2021, 32, 351–366. [Google Scholar] [CrossRef]
- Jadhav, H.B.; Annapure, U.S. Triglycerides of medium-chain fatty acids: A concise review. J. Food Sci. Technol. 2022, 1–10. [Google Scholar] [CrossRef]
- Besbes, S.; Blecker, C.; Deroanne, C.; Bahloul, N.; Lognay, G.; Drira, N.-E.; Attia, H. Date seed oil: Phenolic, tocopherol and sterol profiles. J. Food Lipids 2004, 11, 251–265. [Google Scholar] [CrossRef]
- Al-Juhaimi, F.; Ghafoor, K.; Özcan, M.M. Physical and chemical properties, antioxidant activity, total phenol and mineral profile of seeds of seven different date fruit (Phoenix dactylifera L.) varieties. Int. J. Food Sci. Nutr. 2011, 63, 84–89. [Google Scholar] [CrossRef]
- Bijami, A.; Rezanejad, F.; Oloumi, H.; Mozafari, H. Minerals, antioxidant compounds and phenolic profile regarding date palm (Phoenix dactylifera L.) seed development. Sci. Hortic. 2020, 262, 109017. [Google Scholar] [CrossRef]
- Bouhlali, E.D.T.; Derouich, M.; Hmidani, A.; Bourkhis, B.; Khouya, T.; Filali-Zegzouti, Y.; Alem, C. Protective Effect of Phoenix dactylifera L. Seeds against Paracetamol-Induced Hepatotoxicity in Rats: A Comparison with Vitamin C. Sci. World J. 2021, 2021, 6618273. [Google Scholar] [CrossRef]
- John, J.A.; Shahidi, F. Phenolic content, antioxidant and anti-inflammatory activities of seeds and leaves of date palm (Phoenix dactylifera L.). J. Food Bioact. 2019, 5, 120–130. [Google Scholar] [CrossRef]
- Chileh-Chelh, T.; Lyashenko, S.; Lahlou, A.; Belarbi, E.-H.; Rincón-Cervera, M.A.; Rodríguez-García, I.; Urrestarazu-Gavilán, M.; López-Ruiz, R.; Guil-Guerrero, J.L. Buglossoides spp. seeds, a land source of health-promoting n-3 PUFA and phenolic compounds. Food Res. Int. 2022, 157, 111421. [Google Scholar] [CrossRef] [PubMed]
- He, M.; Qin, C.-X.; Wang, X.; Ding, N.-Z. Plant Unsaturated Fatty Acids: Biosynthesis and Regulation. Front. Plant Sci. 2020, 11, 390. [Google Scholar] [CrossRef] [PubMed]
- Pereira-Faria, J.; Arellano, D.B.; Grimaldi, R.; Da Silva, L.C.R.; Vieira, R.F.; Da Silva, D.B.; Agostini-Costa, T.D.S. Caracterização química da amêndoa de coquinho-azedo (Butia capitata var capitata). Rev. Bras. Frutic. 2008, 30, 549–552. [Google Scholar] [CrossRef]
- Moreira, M.; Arrúa, M.P.; Antunes, A.; Fiuza, T.; Costa, B.; Neto, P.W.; Antunes, S. Characterization of Syagrus romanzoffiana oil aiming at biodiesel production. Ind. Crop Prod. 2013, 48, 57–60. [Google Scholar] [CrossRef]
- Tavares, T.; Magalhães, K.; Lorenzo, N.; Nunes, C. Thermal and chemical characterization of fractions from Syagrus romanzoffiana kernel oil. Grasas y Aceites 2021, 72, e420. [Google Scholar] [CrossRef]
- Harkat, H.; Bousba, R.; Benincasa, C.; Atrouz, K.; Gültekin-Özgüven, M.; Altuntas, Ü.; Demircan, E.; Zahran, H.A.; Özçelik, B. Assessment of biochemical composition and antioxidant properties of Algerian date palm (Phoenix dactylifera L.) seed oil. Plants 2022, 11, 381. [Google Scholar] [CrossRef]
- Nehdi, I.A.; Zarrouk, H.; Al-Resayes, S.I. Changes in chemical composition of Phoenix canariensis Hort. Ex Chabaud palm seed oil during the ripening process. Sci. Hortic. 2011, 129, 724–729. [Google Scholar] [CrossRef]
- Essien, E.E.; Antia, B.S.; Etuk, E.I.; David, E.M. Chemical Composition of Livistona chinensis and Areca catechu Palm Nuts. Chem. Nat. Compd. 2021, 57, 1184–1186. [Google Scholar] [CrossRef]
- Matsue, M.; Mori, Y.; Nagase, S.; Sugiyama, Y.; Hirano, R.; Ogai, K.; Ogura, K.; Kurihara, S.; Okamoto, S. Measuring the Antimicrobial Activity of Lauric Acid against Various Bacteria in Human Gut Microbiota Using a New Method. Cell Transplant. 2019, 28, 1528–1541. [Google Scholar] [CrossRef]
- Nitbani, F.O.; Tjitda, P.J.P.; Nitti, F.; Jumina, J.; Detha, A.I.R. Antimicrobial Properties of Lauric Acid and Monolaurin in Virgin Coconut Oil: A Review. ChemBioEng Rev. 2022, 9, 442–461. [Google Scholar] [CrossRef]
- Liu, W.; Luo, X.; Liu, T.; Feng, F. Study on the digestive characteristics of short-and medium-chain fatty acid structural lipid and its rapid intervention on gut microbes: In vivo and in vitro studies. Food Chem. 2022, 380, 131792. [Google Scholar] [CrossRef] [PubMed]
- Knothe, G.; Dunn, R.O. A Comprehensive Evaluation of the Melting Points of Fatty Acids and Esters Determined by Differential Scanning Calorimetry. J. Am. Oil Chem. Soc. 2009, 86, 843–856. [Google Scholar] [CrossRef]
- Zhao, M.; Chiriboga, D.; Olendzki, B.; Xie, B.; Li, Y.; McGonigal, L.J.; Maldonado-Contreras, A.; Ma, Y. Substantial Increase in Compliance with Saturated Fatty Acid Intake Recommendations after One Year Following the American Heart Association Diet. Nutrients 2018, 10, 1486. [Google Scholar] [CrossRef] [PubMed]
- Ardekani, M.R.S.; Khanavi, M.; Hajimahmoodi, M.; Jahangiri, M.; Hadjiakhoondi, A. Comparison of Antioxidant Activity and Total Phenol Contents of some Date Seed Varieties from Iran. Iran. J. Pharm. Res. IJPR 2010, 9, 141–146. [Google Scholar]
- Granger, K.L.; Gallagher, R.S.; Fuerst, E.P.; Alldredge, J.R. Comparison of seed phenolic extraction and assay methods. Methods Ecol. Evol. 2011, 2, 691–698. [Google Scholar] [CrossRef]
- Barakat, A.Z.; Hamed, A.R.; Bassuiny, R.I.; Abdel-Aty, A.M.; Mohamed, S.A. Date palm and saw palmetto seeds functional properties: Antioxidant, anti-inflammatory and antimicrobial activities. J. Food Meas. Charact. 2020, 14, 1064–1072. [Google Scholar] [CrossRef]
- Appaiah, P.; Sunil, L.; Prasanth-Kumar, P.K.; Gopala-Krishna, A.G. Composition of coconut testa, coconut kernel and its oil. J. Am. Oil Chem. Soc. 2014, 91, 917–924. [Google Scholar] [CrossRef]
- Mahayothee, B.; Koomyart, I.; Khuwijitjaru, P.; Siriwongwilaichat, P.; Nagle, M.; Müller, J. Phenolic compounds, antioxidant activity, and medium chain fatty acids profiles of coconut water and meat at different maturity stages. Int. J. Food Prop. 2016, 19, 2041–2051. [Google Scholar] [CrossRef]
- Chang, C.C.; Lee, S.S. Application of HPLC-SPE-NMR in characterization of bioactive natural compounds. In Instrumental Methods for the Analysis and Identification of Bioactive Molecules; Guddadarangavvanahally, K.J., Bhimanagouda, S.P., Pellati, F., Eds.; American Chemical Society: Washington, DC, USA, 2014; Volume 1185, pp. 217–239. [Google Scholar]
- Bouhlali, E.T.; Alem, C.; Ennassir, J.; Benlyas, M.; Mbark, A.N.; Zegzouti, Y.F. Phytochemical compositions and antioxidant capacity of three date (Phoenix dactylifera L) seeds varieties grown in the South East Morocco. J. Saudi Soc. Agric. Sci. 2017, 16, 350–357. [Google Scholar] [CrossRef]
- Sirisena, S.; Zabaras, D.; Ng, K.; Ajlouni, S. Characterization of date (Deglet Nour) seed free and bound polyphenols by high--performance liquid chromatography--mass spectrometry. J. Food Sci. 2017, 82, 333–340. [Google Scholar] [CrossRef]
- Al-Juhaimi, F.; Ôzcan, M.M.; Adiamo, O.Q.; Alsawmahi, O.N.; Ghafoor, K.; Babiker, E.E. Effect of date varieties on physico--chemical properties, fatty acid composition, tocopherol contents, and phenolic compounds of some date seed and oils. J. Food Proc. Preserv. 2018, 42, e13584. [Google Scholar] [CrossRef]
- Hamza, H.; Elfalleh, W.; Nagaz, K. Date Palm Seed Oil (Phoenix dactylifera L.) Green Extraction: Physicochemical Properties, Antioxidant Activities, and Phenolic and Fatty Acid Profiles. J. Food Qual. 2021, 2021, 12308–12333. [Google Scholar] [CrossRef]
- Turki, M.; Barbosa-Pereira, L.; Bertolino, M.; Essaidi, I.; Ghirardello, D.; Torri, L.; Bouzouita, N.; Zeppa, G. Physico-Chemical Characterization of Tunisian Canary Palm (Phoenix canariensis Hort. Ex Chabaud) Dates and Evaluation of Their Addition in Biscuits. Foods 2020, 9, 695. [Google Scholar] [CrossRef] [PubMed]
- Bar, F.A. New chalcanonol glycoside from the seeds of saw palmetto: Antiproliferative and antioxidant effects. Nat. Prod. Res. 2014, 29, 926–932. [Google Scholar] [PubMed]
- Lahlou, A.; Chileh-Chelh, T.; Lyashenko, S.; Rincón-Cervera, M.A.; Rodríguez-García, I.; López-Ruiz, R.; Urrestarazu, M.; Guil-Guerrero, J.L. Arecaceae fruits: Fatty acids, phenolic compounds and in vitro antitumor activity. Food Biosci. 2022, 50, 102181. [Google Scholar] [CrossRef]
- Martínez-Gutiérrez, A.; Fernández-Duran, I.; Marazuela-Duque, A.; Simonet, N.G.; Yousef, I.; Martínez-Rovira, I.; Martínez-Hoyos, J.; Vaquero, A. Shikimic acid protects skin cells from UV-induced senescence through activation of the NAD+-dependent deacetylase SIRT1. Aging 2021, 13, 12308–12333. [Google Scholar] [CrossRef]
- Birková, A.; Hubková, B.; Bolerázska, B.; Mareková, M.; Cizmárová, B. Caffeic acid: A brief overview of its presence, metabolism, and bioactivity. Bioact. Comp. Health Dis. 2020, 3, 74–81. [Google Scholar] [CrossRef]
- Kadar, N.N.M.A.; Ahmad, F.; Teoh, S.L.; Yahaya, M.F. Caffeic Acid on Metabolic Syndrome: A Review. Molecules 2021, 26, 5490. [Google Scholar] [CrossRef]
- Luo, C.; Zou, L.; Sun, H.; Peng, J.; Gao, C.; Bao, L.; Ji, R.; Jin, Y.; Sun, S. A Review of the Anti-Inflammatory Effects of Rosmarinic Acid on Inflammatory Diseases. Front. Pharmacol. 2020, 11, 153. [Google Scholar] [CrossRef]
- Hitl, M.; Kladar, N.; Gavarić, N.; Božin, B. Rosmarinic Acid–Human Pharmacokinetics and Health Benefits. Planta Medica 2020, 87, 273–282. [Google Scholar] [CrossRef]
- Noor, S.; Mohammad, T.; Rub, M.A.; Raza, A.; Azum, N.; Yadav, D.K.; Hassan, I.; Asiri, A.M. Biomedical features and therapeutic potential of rosmarinic acid. Arch. Pharmacal Res. 2022, 45, 205–228. [Google Scholar] [CrossRef] [PubMed]
- Semwal, R.; Joshi, S.K.; Semwal, R.B.; Semwal, D.K. Health benefits and limitations of rutin—A natural flavonoid with high nutraceutical value. Phytochem. Lett. 2021, 46, 119–128. [Google Scholar] [CrossRef]
- Qu, Z.; Liu, A.; Li, P.; Liu, C.; Xiao, W.; Huang, J.; Liu, Z.; Zhang, S. Advances in physiological functions and mechanisms of (−)-epicatechin. Crit. Rev. Food Sci. Nutr. 2020, 61, 211–233. [Google Scholar] [CrossRef]
- Al-Sheddi, E.S. Anticancer potential of seed extract and pure compound from Phoenix dactylifera on human cancer cell lines. Pharmacogn. Mag. 2019, 15, 494. [Google Scholar] [CrossRef]
- Habib, H.M.; El-Fakharany, E.M.; Souka, U.D.; Elsebaee, F.M.; El-Ziney, M.G.; Ibrahim, W.H. Polyphenol-Rich Date Palm Fruit Seed (Phoenix Dactylifera L.) Extract Inhibits Labile Iron, Enzyme, and Cancer Cell Activities, and DNA and Protein Damage. Nutrients 2022, 14, 3536. [Google Scholar] [CrossRef]
- Khan, M.A.; Singh, R.; Siddiqui, S.; Ahmad, I.; Ahmad, R.; Upadhyay, S.; Barkat, A.; Ali, A.M.A.; Zia, Q.; Srivastava, A.; et al. Anticancer potential of Phoenix dactylifera L. seed extract in human cancer cells and pro-apoptotic effects mediated through caspase-3 dependent pathway in human breast cancer MDA-MB-231 cells: An in vitro and in silico investigation. BMC Complement. Med. Ther. 2022, 22, 1–19. [Google Scholar] [CrossRef]
- Lyashenko, S.; González-Fernández, M.J.; Gómez-Mercado, F.; Yunusova, S.; Denisenko, O.; Guil-Guerrero, J.L. Ribes taxa: A promising source of γ-linolenic acid-rich functional oils. Food Chem. 2019, 301, 125309. [Google Scholar] [CrossRef]
- Lyashenko, S.; Fabrikov, D.; González-Fernández, M.J.; Gómez-Mercado, F.; Ruiz, R.L.; Fedorov, A.; de Bélair, G.; Urrestarazu, M.; Rodríguez-García, I.; Álvarez-Corral, M.; et al. Phenolic composition and in vitro antiproliferative activity of Borago spp. seed extracts on HT-29 cancer cells. Food Biosci. 2021, 42, 101043. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
| Nr. | Species | FAs (% of Total FAs) | Total FA g/100 g Dry Seeds | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| 8:0 (CyA) | 10:0 (CA) | 12:0 (LA) | 14:0 (MA) | 16:0 (PA) | 18:0 (SA) | 18:1n-9 (OA) | 18:2n-6 (LA) | |||
| Subfamily Arecoideae | ||||||||||
| Tribe Areceae | ||||||||||
| 1 | Archontophoenix cunninghamiana | 0.4 ± 0.0 h,i | 0.5 ± 0.4 g,h | 21.4 ± 0.5 j,k,l,m | 12.8 ± 1.8 d,e,f | 15.8 ± 2.5 c | 2.2 ± 0.1 h,I,j,k | 14.0 ± 2.2 m | 32.9 ± 2.2 a | 1.3 ± 0.3 m |
| 2 | Chambeyronia macrocarpa | 1.5 ± 0.0 e,f,g | 1.1 ± 0.4 e,f | 52.5 ± 0.4 a | 10.8 ± 1.1 g,h,i | 9.1 ± 0.5 h,I,j,k | 1.3 ± 0.4 m | 15.1 ± 1.7 l,m | 8.7 ± 0.7 p,q | 2.7 ± 1.4 j,k,l,m |
| 3 | Chrysalidocarpus lutescens | n.d | 0.8 ± 0.1 g,h | 44.3 ± 1.8 c | 29.7 ± 0.4 a | 9.7 ± 0.0 f,g,h,i,j | 1.7 ± 0.1 k,l,m | 6.5 ± 0.0 n | 7.4 ± 0.5 q | 3.0 ± 0.5 j,k,l,m |
| 4 | Howea belmoreana | 1.4 ± 0.1 e,f,g,h | 1.7 ± 0.2 e | 40.4 ± 1.2 d,e | 13.2 ± 1.7 d,e,f | 7.4 ± 0.1 l,m,n | 2.7 ± 0.4 f,g,h,i | 21.4 ± 1.7 k | 10.7 ± 1.1 l,m,n | 6.3 ± 0.9 d,e,f,g |
| 5A | H. forsteriana | 1.4 ± 0.0 e,f,g,h | 2.3 ± 0.1 d,e | 43.9 ± 0.4 c,d | 13.9 ± 0.1 d | 6.2 ± 0.0 n,o | 2.6 ± 0.0 g,h,i | 21.2 ± 0.1 k | 7.8 ± 0.4 p,q | 2.6 ± 1.2 k,l,m |
| 5B | H. forsteriana | n.d | n.d | 30.5 ± 0.4 h | 12.2 ± 0.1 e,f,g | 8.6 ± 0.5 i,j,k,l | 4.2 ± 0.1 b | 30.3 ± 0.2 hi | 14.1 ± 0.5 i,j | 6.7 ± 0.4 d,e,f |
| Tribe Chamaedoreeae | ||||||||||
| 6 | Chamaedorea microspadix | 0.5 ± 0.0 h,i | 0.6 ± 0.0 g,h | 8.5 ± 0.2 r | 20.0 ± 0.3 b | 19.2 ± 0.4 b | 2.6 ± 0.1 g,hi | 26.8 ± 0.2 ij | 20.6 ± 0.2 e,f | 4.8 ± 0.1 f,g,h,i,j |
| 7 | C. oblongata | n.d | n.d | n.d | 16.3 ± 0.1 c | 31.2 ± 0.7 a | 6.2 ± 0.5 a | 20.3 ± 0.4 k | 26.3 ± 0.1 c | 4.3 ± 0.7 g,h,i,j,k |
| Tribe Cocoseae | ||||||||||
| 8 | Butia capitata | 14.4 ± 1.3 a | 10.5 ± 0.1 a | 34.9 ± 1.5 f,g | 8.7 ± 0.1 j,k | 5.4 ± 0.1 o | 2.3 ± 0.3 g,h,i,j | 18.8 ± 0.6 k,l | 4.4 ± 0.0 r | 28.0 ± 0.9 b |
| 9 | Cocos nucifera | 7.4 ± 0.7 b | 6.8 ± 0.4 b | 51.0 ± 0.3 b | 16.6 ± 0.6 c | 7.9 ± 0.3 k,l,m | 2.7 ± 0.5 f,g,h | 6.1 ± 0.0 n | 1.1 ± 0.0 s | 68.3 ± 2.9 a |
| 10 | Syagrus romanzofianna | 3.5 ± 2.4 d | 2.6 ± 1.3 d | 25.2 ± 8.0 i | 9.0 ± 0.3 j,k | 9.8 ± 1.0 f,g,h,i | 1.3 ± 0.0 m | 39.5 ± 2.1 e,f,g | 9.3 ± 1.8 n,o,p | 28.1 ± 1.3 b |
| Subfamily Coryphoideae | ||||||||||
| Tribe Caryoteae | ||||||||||
| 11 | Arenga engleri | 5.6 ± 0.4 c | 4.9 ± 0.0 c | 37.4 ± 0.3 e,f | 9.4 ± 0.3 i,j | 12.3 ± 0.3 d,e | 3.3 ± 0.4 c,d,e | 15.6 ± 0.0 l,m | 11.2 ± 0.2 l,m | 8.4 ± 0.3 c,d |
| Tribe Phoeniceae | ||||||||||
| 12 | Phoenix canariensis | n.d | n.d | 13.4 ± 1.2 q | 7.0 ± 1.1 l,m | 13.0 ± 0.8 d | n.d | 45.5 ± 0.2 a,b,c | 20.5 ± 1.9 e,f | 3.7 ± 3.6 h,i,j,k,l |
| 13A | P. dactylifera var. Deglet Nour | 0.4 ± 0.0 h,i | 0.5 ± 0.0 g,h | 22.0 ± 0.4 i,j,k,l | 11.1 ± 1.6 g,h | 9.7 ± 1.3 f,g,h,i,j | 3.2 ± 0.2 d,e,f | 42.5 ± 1.1 b,c,d,e,f | 8.9 ± 0.6 o,p,q | 3.4 ± 0.5 i,j,k,l,m |
| 13B | P. dactylifera var. Medjool | 0.4 ± 0.0 h,i | 0.4 ± 0.1 g,h | 17.5 ± 0.1 n,o,p | 10.9 ± 0.0 g,h,i | 10.5 ± 0.0 f,g | 2.4 ± 0.6 g,h,i,j | 44.7 ± 0.2 a,b,c,d | 10.5 ± 0.0 m,n,o | 6.4 ± 1.8 d,e,f,g |
| 14 | P. reclinata | 0.3 ± 0.0 h,i | 0.4 ± 0.0 g,h | 23.2 ± 0.2 i,j,k | 14.2 ± 0.2 d | 12.8 ± 0.0 d | 2.2 ± 0.0 g,h,i,j,k | 27.5 ± 0.1 h,i,j | 19.0 ± 0.1 f,g | 3.8 ± 0.1 h,i,j,kl |
| Tribe Sabaleae | ||||||||||
| 15 | Sabal bermudana | 0.5 ± 0.0 h,i | 0.6 ± 0.1 g,h | 20.4 ± 0.2 k,l,m,n | 11.6 ± 0.0 f,g | 9.5 ± 0.0 g,h,i,j | 2.3 ± 0.0 g,h,ij | 41.5 ± 0.1 c,d,e,f | 12.1 ± 0.1 k,l | 4.3 ± 0.5 g,h,i,j,k |
| 16 | S. minor | 0.5 ± 0.1 h,i | 0.5 ± 0.0 g,h | 23.9 ± 0.1 i,j,k | 10.8 ± 0.0 g,h,i | 6.6 ± 0.3 m,n,o | 2.1 ± 0.3 i,j,k,l | 38.4 ± 0.4 f,g | 13.9 ± 0.1j | 4.3 ± 0.4 g,h,i,j,k |
| 17 | S. palmetto | n.d | n.d | 15.9 ± 0.2 o,p,q | 11.0 ± 0.1 g,h,i | 9.7 ± 0.4 f,g,h,i,j | 2.3 ± 0.1 g,h,i,j | 47.3 ± 0.5 a | 13.9 ± 0.1j | 5.4 ± 0.1 e,f,g,hi |
| 18 | S. domingensis | 0.2 ± 0.0 i | 0.2 ± 0.0 g,h | 14.7 ± 0.2 p,q | 10.0 ± 0.1 h,i,j | 9.8 ± 0.0 f,g,h,i | 2.4 ± 0.0 g,h,i,j | 45.8 ± 0.0 a,b | 15.7 ± 0.1 h,i | 7.4 ± 0.4 d,e |
| Tribe Trachycarpeae | ||||||||||
| 19A | Chamaerops humilis | n.d | n.d | 18.2 ± 0.1 m,n,o,p | 7.8 ± 0.7 k,l | 9.8 ± 0 f,g,h,i | 3.8 ± 0.2 b,c,d | 41.1 ± 0.1 d,e,f | 18.4 ± 0.4 g | 6.3 ± 0.7 d,e,f,g |
| 19B | C. humilis | n.d | n.d | 13.4 ± 0.1 q | 7.5 ± 1.5 k,l | 12.8 ± 0.5 d | 3.9 ± 0.2 b,c | 39.5 ± 0.1 e,f,g | 24.1 ± 0.4 d | 5.5 ± 0.3 e,f,g,hi |
| 19C | C. humilis | n.d | n.d | 17.2 ± 0.2 n,o,p | 5.6 ± 0.0 m | 10.4 ± 0.1 f,g,h | 2.8 ± 0.1 e,f,g | 46.5 ± 0.5 a,b | 17.7 ± 0.1 g | 5.7 ± 0.1 e,f,g,h |
| 19D | C. humilis | 1.9 ± 0.0 e,f | 2.7 ± 0.0 d | 21.2 ± 0.2 j,k,l,m | 6.7 ± 0.0 l,m | 8.4 ± 0.1 j,k,l | 1.9 ± 0.1 j,k,l | 42.9 ± 0.8 b,c,d,e | 14.2 ± 0.3 h,i,j | 6.1 ± 0.3 e,f,g |
| 20 | Livistona chinensis | n.d | n.d | 18.9 ± 0.3 l,m,n,o | 10.7 ± 0.4 g,h,i | 8.6 ± 0.4 i,j,k,l | 1.6 ± 0.1 l,m | 39.0 ± 0.7 e,f,g | 21.5 ± 0.1 e | 4.4 ± 0.3 g,h,i,j,k |
| 21 | L. fulva | 0.6 ± 0.0 h,i | 0.9 ± 0.0 f,g | 32.7 ± 1.5 g,h | 8.7 ± 0.4 j,k | 9.1 ± 0.4 h,i,j,k | n.d | 26.0 ± 0.6 j | 15.7 ± 0.9 h,i | 8.4 ± 0.3 c,d |
| 22 | L. saribus | 2.4 ± 0.3 d,e | 5.3 ± 1.3 c | 30.1 ± 1.4 h | 13.4 ± 0.6 d,e | 12.9 ± 0.5 d | 3.3 ± 0.4 c,d,e | 12.7 ± 0.6 m | 15.7 ± 0.9 h | 1.2 ± 0.1 m |
| 23 | Trachycarpus fortunei | 0.8 ± 0.0 g,h,i | 0.9 ± 0.0 f,g | 12.9 ± 1.1 q | 8.8 ± 1.0 j,k | 10.9 ± 0.2 e,f | 2.4 ± 0.3 g,h,i,j | 31.3 ± 0.8 h | 29.0 ± 0.9 b | 1.8 ± 0.1 l,m |
| 24 | Washingtonia robusta | 0.9 ± 0.0 f,g,h,i | 0.8 ± 0.0 g | 24.4 ± 0.2 i,j | 12.8 ± 0.0 d,e,f | 7.7 ± 0.0 l,m | 2.8 ± 0.0 e,f,g | 36.7 ± 0.8 g | 13.4 ± 0.2 j,k | 9.9 ± 0.3 c |
| Code | Species | Quinic Acid b | Chelidonic Acid b | Trans-Aconitic Acid b | Gallic Acid | Vanillic Acid | Protocatechuic Acid | Salicylic Acid | 4-OH-Benzoic Acid | DL-p-OH-Phenyllactic Acid |
| Retention time (min) | 8.73 | 9.38 | 11.25 | 13.84 | 14.57 | 18.57 | 25.27 | 27.55 | 29.82 | |
| Subfamily Arecoideae | ||||||||||
| Tribe Areceae | ||||||||||
| 1 | Archontophoenix cunninghamiana | 2.3 ± 0.1 i,j,k | 1.1 ± 0.1 e,f,g | n.d | 4.8 ± 0.8 a | 0.2 ± 0.0 l,m | 9.8 ± 0.8 b,c | 11.9 ± 1.8 b,c | 4.2 ± 0.6 e | 1.8 ± 0.2 g,h |
| 2 | Chambeyronia macrocarpa | 4.3 ± 0.3 f,g,h | 1.2 ± 0.1 e,f | 0.6 ± 0.0 c,d | 0.2 ± 0.0 l | 2.7 ± 0.6 e,f,g | 0.8 ± 0.0 h | 2.9 ± 0.2 i,j,k,l | 2.6 ± 0.5 f,g,h | 1.2 ± 0.1 g,h,i |
| 3 | Dypsis lutescens | 1.5 ± 0.1 i,j,k,l | 0.2 ± 0.0 i,j | 0.7 ± 0.1 c,d | 0.1 ± 0.0 l | 5.2 ± 0.1 d | 7.8 ± 0.7 d,e | 10.0 ± 0.8 c,d,e | 9.2 ± 1.2 a | 15.8 ± 1.2 b |
| 4 | Howea belmoreana | 0.1 ± 0.0 l | 0.3 ± 0.0 h,i,j | 1.1 ± 0.1 c,d | 0.1 ± 0.0 l | 1.2 ± 0.1 h,i,j,k,l,m | 5.6 ± 0.7 f,g | 6.4 ± 0.2 f,g,h | 6.9 ± 0.7 c,d | 15.4 ± 0.2 b |
| 5A | H. forsteriana | 0.1 ± 0.0 l | 0.8 ± 0.2 e,f,g,h,i,g, | n.d | 0.9 ± 0.1 f,g,h,i,j,k | 1.3 ± 0.1 h,i,j,k,l,m | 1.5 ± 0.2 h | 4.5 ± 0.1 g,h,i | 0.9 ± 0.2 l,m | 6.1 ± 0.2 f |
| 5B | H. forsteriana | 0.1 ± 0.0 l | 0.1 ± 0.0 j | n.d | 2.9 ± 0.2 b | 1.8 ± 0.1 f,g,h,i,j,k | 0.9 ± 0.2 h | 3.8 ± 0.3 h,i,j,k | 3.9 ± 0.2 e,f | 1.8 ± 0.2 g,h |
| Tribe Chamaedoreeae | ||||||||||
| 6 | Chamaedorea microspadix | 1.6 ± 0.2 i,j,k,l | 11.9 ± 1.1 a | 0.2 ± 0.0 c,d | 1.3 ± 0.9 c,d,e,f,g,h | 5.2 ± 0.5 d | 0.6 ± 0.0 h | 4.2 ± 0.6 g,h,i,j | 7.3 ± 0.3 b,c | 0.1 ± 0.0 i |
| 7 | C. oblongata | 0.1 ± 0.0 l | 0.2 ± 0.0 i,j | n.d | 0.4 ± 0.0 j,k,l | 0.6 ± 0.0 k,l,m | 0.3 ± 0.0 h | 0.1 ± 0.0 l | 1.3 ± 0.0 i,j,k,l | 2.4 ± 0.3 g |
| Tribe Cocoseae | ||||||||||
| 8 | Butia capitata | 0.2 ± 0.0 l | 0.5 ± 0.0 f,g,h,i | 1.1 ± 0.1 c,d | 0.7 ± 0.0 g,h,i,j,k,l | 0.8 ± 0.3 j,k,l,m | 1.7 ± 0.3 h | 3.2 ± 0.2 h,i,j,k,l | 2.5 ± 0.3 g,h,i,j | 0.9 ± 0.1 h,i |
| 9 | Cocos nucifera | 0.4 ± 0.0 l,k | 0.9 ± 0.0 e,f,g,h,i | 1.7 ± 0.1 b,c,d | 1.9 ± 0.0 c,d | 0.7 ± 0.0 k,l,m | 1.3 ± 0.2 h | 0.9 ± 0.0 j,k,l | 3.2 ± 0.2 e,f,g,h | 0.4 ± 0.0 i |
| 10 | Syagrus romanzofianna | 0.3 ± 0.0 l | 0.6 ± 0.0 f,g,h,i | 0.1 ± 0.0 d | 0.5 ± 0.0 i,j,k,l | n.d | 4.9 ± 0.3 g | 11.3 ± 0.3 c,d | 0.4 ± 0.0 m | 14.8 ± 0.1 b,c |
| Subfamily Coryphoideae | ||||||||||
| Tribe Caryoteae | ||||||||||
| 11 | Arenga engleri | 0.8 ± 0.0 k,l | 0.3 ± 0.0 h,i,j | 1.4 ± 0.1 b,c,d | 1.0 ± 0.2 e,f,g,h,i,j | 13.1 ± 0.9 b | 0.2 ± 0.0 h | n.d | 0.1 ± 0.0 m | 11.6 ± 0.9 d |
| Tribe Phoeniceae | ||||||||||
| 12 | Phoenix canariensis | 8.6 ± 1.3 d | 0.4 ± 0.0 g,h,i,j | n.d | 1.7 ± 0.4 c,d,e | 0.1 ± 0.0 m | 7.1 ± 0.9 e,f | 8.5 ± 2.2 c,d,e,f | 0.3 ± 0.0 m | 5.7 ± 0.4 f |
| 13A | P. dactylifera var. Deglet Nour | 1.5 ± 0.3 i,j,k,l | 0.3 ± 0.0 h,i,j | n.d | 0.4 ± 0.0 g,k,l | 22.0 ± 1.0 a | 4.4 ± 0.1 g | n.d | n.d | 6.2 ± 0.3 f |
| 13B | P. dactylifera var. Medjool | 1.8 ± 0.2 i,j,k,l | 0.2 ± 0.0 i,j | n.d | 1.9 ± 0.2 c,d | 1.3 ± 0.1 h,i,j,k,l,m | 0.9 ± 0.1 h | n.d | 1.2 ± 0.0 j,k,l,m | 2.1 ± 0.3 g,h |
| 14 | P. reclinata | 0.9 ± 0.0 k,l | n.d | n.d | 0.9 ± 0.0 f,g,h,i,j,k | 3.2 ± 0.0 e | 0.1 ± 0.0 h | n.d | n.d | 0.3 ± 0.0 i |
| Tribe Sabaleae | ||||||||||
| 15 | Sabal bermudana | 6.9 ± 0.7 d,e | 0.3 ± 0.0 h,i,j | n.d | 0.7 ± 0.1 g,h,i,j,k,l | 2.4 ± 1.2 e,f,g,h | 8.5 ± 0.7 c,d,e | 8.9 ± 0.4 c,d,e,f | 8.5 ± 0.9 a,b | n.d |
| 16 | S. minor | 3.3 ± 0.0 g,h,i | 0.1 ± 0.0 j | 0.7 ± 0.0 c,d | 1.4 ± 0.1 c,d,e,f,g | 3.0 ± 0.1 e,f | 9.3 ± 0.6 c,d | 15.1 ± 1.5 b | 4.1 ± 0.1 e | 13.8 ± 0.1 c |
| 17 | S. palmetto | 16.2 ± 2.9 b | 0.2± 0.0 i,j | 1.9 ± 0.2 c,b,d | 1.0 ± 0.0 e,f,g,h,i,j | 7.2 ± 0.2 c | 15.7 ± 0.2 a | 33.5 ± 2.9 a | 1.9 ± 0.1 h,i,j,k,l | 17.1 ± 0.4 a |
| 18 | S. domingensis | 1.1 ± 0.0 j,k,l | 0.9 ± 0.0 e,f,g,h,i | 0.4 ± 0.0 c,d | 1.6 ± 0.1 c,d,e,f | n.d | 5.7 ± 0.3 f,g | 4.1 ± 0.3 g,h,i,j | n.d | n.d |
| Tribe Trachycarpeae | ||||||||||
| 19A | Chamaerops humilis | 12.9 ± 1.3 c | 2.3 ± 0.4 d | n.d | 1.3 ± 0.0 c,d,e,f,g,h | 2.4 ± 0.1 e,f,g,h | 11.1 ± 0.7 b | 6.5 ± 0.5 fgh | 9.8 ± 1.2 a | n.d |
| 19B | C. humilis | 19.9 ± 0.8 a | 2.2 ± 0.2 d | 0.4 ± 0.0 c,d | 1.4 ± 0.1 c,d,e,f,g | 2.3 ± 0.0 e,f,g,h,i | 16.1 ± 2.5 a | 8.4 ± 0.4 d,e,f | 1.0 ± 0.0 k,l,m | 5.6 ± 0.4 f |
| 19C | C. humilis | 5.6 ± 0.4 e,f | 1.0 ± 0.0 e,f,g,h | 2.3 ± 2.8 b,c | 2.0 ± 0.1 c | 1.6 ± 0.1 g,h,i,j,k | 4.9 ± 0.2 g | 7.4 ± 0.8 e,f,g | 7.4 ± 0.3 b,c | 5.2 ± 0.3 f |
| 19D | C. humilis | 5.7 ± 0.3 e,f | 1.4 ± 0.1 e | 0.6 ± 0.0 c,d | 1.2 ± 0.1 d,e,f,g,h,i | 2.1 ± 0.8 e,f,g,h,i | 1.5 ± 0.0 h | 10.3 ± 1.9 c,d,e | 2.6 ± 0.3 f,g,h,i | 8.8 ± 0.5 e |
| 20 | Livistona chinensis | 1.0 ± 0.0 j,k,l | 9.7 ± 0.7 b | 4.3 ± 0.5 a | 0.6 ± 0.0 h,i,j,k,l | 2.0 ± 0.0 e,f,g,h,i,j | 6.9 ± 0.3 e,f | 10.5 ± 3.1 c,d,e | 5.9 ± 0.3 d | n.d |
| 21 | L. fulva | 1.2 ± 0.0 j,k,l | 6.3 ± 0.1 c | 3.1 ± 0.1 b | 1.1 ± 0.1 e,f,g,h,i,j | 1.7 ± 0.0 g,h,i,j,k | 1.4 ± 0.1 h | 3.6 ± 0.1 h,i,j,k | 3.5 ± 0.2 e,f,g | n.d |
| 22 | L. saribus | 0.5 ± 0.0 k,l | 0.2 ± 0.0 i,j | 0.1 ± 0.0 d | 0.1 ± 0.0 l | 2.0 ± 0.0 e,f,g,h,i,j | 1.7 ± 0.0 h | 0.6 ± 0.0 k,l | 2.3 ± 0.1 g,h,i,j,k | n.d |
| 23 | Trachycarpus fortunei | 5.0 ± 0.2 e,f,g | 0.4 ± 0.0 g,h,i,j | 1.6 ± 0.2 b,c,d | 0.2 ± 0 l | 1.1 ± 0.1 i,j,k,l,m | 0.2 ± 0.0 h | 0.5 ± 0.0 k,l | 0.4 ± 0.0 m | n.d |
| 24 | Washingtonia robusta | 2.9 ± 0.2 h,i,j | 0.1 ± 0.0 j | 0.7 ± 0.1 c,d | 1.9 ± 0.3 c,d | 1.4 ± 0.0 h,i,j,k,l | 0.7 ± 0.0 h | 0.5 ± 0.0 k,l | 0.5 ± 0.0 m | n.d |
| Code | Species | 3,4-Dihydroxyhydrocinnamic Acid | Chlorogenic Acid | Caffeic Acid | (-)-Catechin | Syringic Acid | Dactylifric Acidc | trans-Coumaric Acid | Ferulic Acid | Sinapic Acid |
| Retention time (min) | 30.56 | 35.21 | 36.32 | 38.53 | 39.10 | 42.76 | 44.64 | 47.72 | 49.38 | |
| Subfamily Arecoideae | ||||||||||
| Tribe Areceae | ||||||||||
| 1 | Archontophoenix cunninghamiana | 0.8 ± 0.0 b,c | 1.8 ± 0.3 a | 27.9 ± 1.6 d | 11.6 ± 1.4 g | 1.6 ± 0.2 g,h,i,j | 1.8 ± 0.0 j | 0.4 ± 0.0 f,g | 8.5 ± 0.6 a | 0.5 ± 0.0 f,g |
| 2 | Chambeyronia macrocarpa | 2.1 ± 0.3 a | 0.9 ± 0.2 c,d,e | 38.5 ± 0.9 c | 0.8 ± 0.0 j,k | 16.1 ± 1.0 b | 0.6 ± 0.0 j | 2.6 ± 2.3 d | 1.4 ± 0.0 h,i,j,k | n.d |
| 3 | Dypsis lutescens | n.d | 0.9 ± 0.1 c,d,e | 53.5 ± 3.6 b | 1.4 ± 0.2 i,j,k | 0.6 ± 0.1 i,j | 0.4 ± 0.0 j | 0.6 ± 0.0 f,g | 2.4 ± 0.1 e | 0.4 ± 0.1 f,g |
| 4 | Howea belmoreana | 0.1 ± 0.0 d | 0.4 ± 0.2 f,g,h | 1.2 ± 0.0 h,i,j,k | 17.1 ± 0.4 d | 0.1 ± 0.0 j | 1.8 ± 0.2 j | 0.4 ± 0.0 f,g | 2.2 ± 0.2 e,f | 7.7 ± 0.9 b |
| 5A | H. forsteriana | n.d | 0.1 ± 0.1 h | 0.4 ± 0.0 j,k | 3.0 ± 0.1 h,i,j,k | 12.8 ± 0.9 c,d | 0.4 ± 0.0 j | 0.8 ± 0.1 e,f,g | 5.8 ± 0.3 b | 0.1 ± 0.0 g |
| 5B | H. forsteriana | 0.1 ± 0.0 d | 0.2 ± 0.0 h | 2.5 ± 0.1 h,i,j,k | 1.0 ± 0.1 j,k | 2.6 ± 0.3 f,g,h | 1.0 ± 0.0 j | 0.7 ± 0.1 f,g | 4.5 ± 0.6 c | 0.5 ± 0.1 f,g |
| Tribe Chamaedoreeae | ||||||||||
| 6 | Chamaedorea microspadix | 2.3 ± 0.0 a | 0.2 ± 0.0h | 0.9 ± 0.0 i,j,k | 3.1 ± 0.4 h,i,j,k | 0.3 ± 0.0 i,j | 4.3 ± 0.2 i,j | 0.6 ± 0.0 f,g | 0.2 ± 0.0 p | 0.5 ± 0.0 f,g |
| 7 | C. oblongata | 1.0 ± 0.0 b | 0.1 ± 0.0h | 0.4 ± 0.0 j,k | 28.9 ± 0.0 a | 1.3 ± 0.2 h,i,j | 0.9 ± 0.0 j | 1.0 ± 0.0 e,f,g | 0.3 ± 0.0 o,p | 0.2 ± 0.0 f,g |
| Tribe Cocoseae | ||||||||||
| 8 | Butia capitata | n.d | 0.3 ± 0.0 g,h | 0.6 ± 0.0 i,j,k | 13.5 ± 0.7 f,g | 2.7 ± 0.2 f,g,h | 3.0 ± 0.2 i,j | 1.9 ± 0.2 d,e,f | 5.0 ± 0.3 c | 1.2 ± 0.1 d,e,f |
| 9 | Cocos nucifera | n.d | 1.8 ± 0.3 a | 0.1 ± 0.0 k | 17.9 ± 0.5 c,d | 1.5 ± 0.3 g,h,i,j | 1.9 ± 0.2 j | 0.9± 0.0 e,f,g | 1.8 ± 0.0 e,f,g,h | 0.9 ± 0.0 e,f,g |
| 10 | Syagrus romanzofianna | n.d | 0.8 ± 0.0 d,e,f | n.d | 15.1 ± 1.3 d,e | 3.2 ± 0.2 f,g | 2.0 ± 0.2 i,j | 0.1 ± 0.0 g | 4.5 ± 0.3c | 1.4 ± 0.1 c,d,e,f |
| Subfamily Coryphoideae | ||||||||||
| Tribe Caryoteae | ||||||||||
| 11 | Arenga engleri | n.d | 0.8 ± 0.2 c,c,d,e,f | 3.7 ± 0.9 h,i | 20.1 ± 2.5 c | 8.1 ± 1.2 e | 1.6 ± 0.2 j | 1.3 ± 0.0 d,e,f,g | 0.6 ± 0.0 m,n,o,p | 0.5 ± 0.0 f,g |
| Tribe Phoeniceae | ||||||||||
| 12 | Phoenix canariensis | 0.2 ± 0.0 d | 0.5 ± 0.0 e,f,g,h | 16.2 ± 2.0 e | 4.2 ± 0.2 h,i | 3.8 ± 0.7 f | 0.4 ± 0.0 j | 28.1 ± 1.3 a | 3.2 ± 0.2 d | 2.5 ± 0.3 c |
| 13A | P. dactylifera var. Deglet Nour | 0.3 ± 0.0 d | 0.2 ± 0.0 h | 1.2 ± 0.0 h,i,j,k | 25.1 ± 3.5 b | 11.4 ± 0.2 d | 27.4 ± 3.1 d,e | 11.8 ± 0.3 c | 0.9 ± 0.0 j,k,l,m,n,o | 8.1 ± 0.6 b |
| 13B | P. dactylifera var. Medjool | 0.7 ± 0.0 c | 1.6 ± 0.2 a,b | 2.6 ± 1.1 h,i,j,k | 20.7 ± 1.9c | 19.3 ± 2.1 a | 31.2 ± 1.1 c,d | 27.2 ± 0.0 a | 1.7 ± 0.3 f,g,h,i | 0.9 ± 0.0 e,f,g |
| 14 | P. reclinata | 0.2 ± 0.0 d | 0.7 ± 0.0 d,e,f,g | 1.9 ± 0.2 h,i,j,k | 12.5 ± 1.4 f,g | 13.5 ± 0.3 c | 29.1 ± 0.1 d,e | 20.9 ± 0.8 b | 0.2 ± 0.0 p | 0.5 ± 0.1 f,g |
| Tribe Sabaleae | ||||||||||
| 15 | Sabal bermudana | n.d | 1.6 ± 0.0 a,b | 4.2 ± 0.0 g,h | 4.8 ± 0.6 h | 2.4 ± 0.1 f,g,h | 79.7 ± 12.1 a | 0.6 ± 0.1 f,g | n.d | 1.8 ± 0.1 c,d,e |
| 16 | S. minor | n.d | 0.5 ± 0.0 e,f,g,h | 16.1 ± 0.6 e | 3.6 ± 0.5 h,i,j | 1.0 ± 0.1 h,i,j | 38.1 ± 3.1 b,c | 2.3 ± 0.1 d,e | 1.1 ± 0.2 i,j,k,l,m,n | 1.9 ± 0.3 c,d,e |
| 17 | S. palmetto | n.d | 0.4 ± 0.0 f,g,h | 15.1 ± 1.2 e | 1.5 ± 0.2 i,j,k | 1.5 ± 0.1 g,h,i,j | 22.3 ± 2.1 e,f | 0.3 ± 0.0 g | 1.2 ± 0.3 h,i,j,k,l,m | n.d |
| 18 | S. domingensis | n.d | 1.1 ± 0.0 c,d | 14.5 ± 1.5 e | 2.6 ± 0.3 h,i,j,k | n.d | 21.7 ± 0.1 e,f | 0.9 ± 0.0 e,f,g | 0.7 ± 0.0 l,m,n,o,p | n.d |
| Tribe Trachycarpeae | ||||||||||
| 19A | Chamaerops humilis | n.d | 0.2 ± 0.0 h | 0.5 ± 0.0 j,k | 0.9 ± 0.1 j,k | n.d | 1.2 ± 0.1 j | 0.9 ± 0.2 e,f,g | 1.3 ± 0.1 h,i,j,k,l | n.d |
| 19B | C. humilis | n.d | 0.1 ± 0.1 h | 0.4 ± 0.0 j,k | 1.4 ± 0.2 i,j,k | n.d | 0.1 ± 0.0 j | 0.1 ± 0.0 g | 1.5 ± 0.2 g,h,i,j | 15.1 ± 1.7 a |
| 19C | C. humilis | n.d | 0.3 ± 0.0 gh | 0.8 ± 0.1 i,j,k | 3.0 ± 0.2 h,i,j,k | n.d | 0.5 ± 0.0 j | 0.3 ± 0.0 g | 0.6 ± 0.2 m,n,o,p | 0.2 ± 0.0 f,g |
| 19D | C. humilis | n.d | 1.2 ± 0.6 b,c | 2.2 ± 0.1 h,i,j,k | 0.4 ± 0.0 k | 1.6 ± 0.3 g,h,i,j | 0.1 ± 0.0 j | 0.4 ± 0.0 f,g | 2.1 ± 0.2 e,f,g, | 2.4 ± 0.1 c,d |
| 20 | Livistona chinensis | n.d | 0.5 ± 0.0 e,f,g,h | 66.1 ± 2.3 a | 2.6 ± 0.2 h,i,j,k | 1.3 ± 0.0 h,i,j | 39.1 ± 0.4 b | 0.4 ± 0.0 f,g | 0.8 ± 0.0 k,l,m,n,o,p | 0.2 ± 0.0 f,g |
| 21 | L. fulva | n.d | 0.3 ± 0.1 g,h | 8.0 ± 0.3 f | 1.3 ± 0.2 i,j,k | 2.0 ± 0.0 g,h,i | 12.4 ± 0.6 g,h | 0.2 ± 0.0 g | 0.5 ± 0.0 n,o,p | n.d |
| 22 | L. saribus | n.d | 0.5 ± 0.1 e,f,g,h | 8.9 ± 0.1 f | 1.5 ± 0.3 i,j,k | 0.2 ± 0.0 j | 18.0 ± 0.1 f,g | 1.0 ± 0.0 e,f,g | 0.2 ± 0.0 p | n.d |
| 23 | Trachycarpus fortunei | n.d | 0.1 ± 0.0 h | 3.4 ± 0.3 h,i,j | 2.1 ± 0.3 h,i,j,k | 1.8 ± 0.3 g,h,i,j | 12.8 ± 0.3 g,h | 0.5 ± 0.0 f,g | 0.6 ± 0.0 m,n,o,p | n.d |
| 24 | Washingtonia robusta | n.d | 0.4 ± 0.0 f,g,h | 7.3 ± 0.4 f,g | 0.4 ± 0.0 k | 1.6 ± 0.6 g,h,i,j | 9.0 ± 0.2 h,i | 1.0 ± 0.0 e,f,g | 0.3 ± 0.0 o,p | 0.4 ± 0.0 f,g |
| Code | Species | Eriodictyol d | Rutin | Rosmarinic Acid | 2-OH-4-methoxybenzoic Acid | Naringenin | Quercetin | Luteolin | Kaempferol | Total Phenolics (mg/100 g) |
| Retention time (min) | 54.26 | 57.35 | 58.65 | 62.87 | 65.96 | 68.76 | 72.28 | 76.73 | ||
| Subfamily Arecoideae | ||||||||||
| Tribe Areceae | ||||||||||
| 1 | Archontophoenix cunninghamiana | 2.9 ± 0.2 e,f,g | 22.9 ± 3.3 d | 41.8 ± 1.6 a | 5.1 ± 0.4 d | 9.5 ± 1.6 a | 1.8 ± 0.1 d | 3.1 ± 0.5 e | 22.8 ± 2.2 a | 197.5 ± 5.6 b |
| 2 | Chambeyronia macrocarpa | 1.4 ± 0.3 h,i,j,k,l | 1.7 ± 0.1 k,l,m | 6.6 ± 0.6 c | 3.7 ± 0.2 e | 0.6 ± 0.0 e,f | 0.3 ± 0.0 i,j,k | 0.4 ± 0.0 f | 0.1 ± 0.0 e | 88.2 ± 2.9 f,g |
| 3 | Dypsis lutescens | 3.7 ± 0.3 e | 2.4 ± 0.2 j,k,l,m | 3.0 ± 0.0 d,e,f | 1.7 ± 0.0 h,i | 0.1 ± 0.0 f | 0.7 ± 0.1 f,g,h | 0.1 ± 0.0 f | 0.7 ± 0.0 d,e | 120.7 ± 4.1 e |
| 4 | Howea belmoreana | 6.3 ± 0.5 c | 26.1 ± 2.3 d | 0.1 ± 0.0 j | 0.3 ± 0.0 j | 0.7 ± 0.0 e,f | 1.7 ± 0.0 d | 0.2 ± 0.0 f | 0.5 ± 0.0 e | 96.1 ± 2.8 f |
| 5A | H. forsteriana | 20.7 ± 1.5 a | 15.3 ± 1.4 e,f | 0.3 ± 0.0 i,j | 0.3 ± 0.0 j | 0.4 ± 0.0 e,f | 0.7 ± 0.0 f,g,h | 0.4 ± 0.0 f | 0.3 ± 0.0 e | 77.0 ± 2.3 h |
| 5B | H. forsteriana | 9.0 ± 0.8 b | 35.4 ± 2.9 c | 0.4 ± 0.0 h,i,j | 0.9 ± 0.0 i,j | 0.2 ± 0.0 f | 0.4 ± 0.0 h,i,j,k | 0.3 ± 0.0 f | 0.5 ± 0.0 e | 75.3 ± 3.1 h |
| Tribe Chamaedoreeae | ||||||||||
| 6 | Chamaedorea microspadix | 0.5 ± 0 k,l,m | 1.7 ± 0.2 k,l,m | 1.4 ± 0.3 g,h,i,j | 2.3 ± 0.5 f,g,h | 2.3 ± 0.1 c,d | 0.3 ± 0.0 i,j,k | 0.6 ± 0.0 f | 0.1 ± 0.0 e | 40.3 ± 1.4 n,o |
| 7 | C. oblongata | 0.2 ± 0 m | 0.3 ± 0.0 m | 1.9 ± 0.1 e,f,g,h | n.d | 1.3 ± 0.2 d,e | 0.2 ± 0.0 j,k | 0.4 ± 0.0 f | 0.2 ± 0.0 e | 43.7 ± 0.4 lmno |
| Tribe Cocoseae | ||||||||||
| 8 | Butia capitata | 3.6 ± 0.3 e | 3.5 ± 0.1 j,k,l,m | 0.4 ± 0.0 h,i,j | 0.9 ± 0.0 i,j | 0.9 ± 0.0 e,f | 0.6 ± 0.0 f,g,h,i | 1.2 ± 0.3 e,f | 0.2 ± 0.0 e | 49.3 ± 1.1 k,l,m,n,o |
| 9 | Cocos nucifera | 1.9 ± 0.4 g,h,i | 5.9 ± 0.4 i,j | 1.1 ± 0.0 h,i,j | 0.2 ± 0.0 j | 0.3 ± 0.0 e,f | 5.0 ± 0.3 a | 0.4 ± 0.0 f | 4.5 ± 0.0 b | 54.5 ± 1.0 j,k,l |
| 10 | Syagrus romanzofianna | 2.9 ± 0.3 e,f,g | 2.7 ± 0.0 j,k,l,m | 0.5 ± 0.0 h,i,j | 1.8 ± 0.2 g,h,i | 4.3 ± 0.4 b | 2.3 ± 0.2 c | 1.4 ± 0.2 e,f | 2.1 ± 0.3 c | 77.0 ± 1.6 h |
| Subfamily Coryphoideae | ||||||||||
| Tribe Caryoteae | ||||||||||
| 11 | Arenga engleri | 3.1 ± 0.3 e,f | 11.8 ± 1.2 f,g,h | 3.4 ± 0.4 d,e | 11.9 ± 0.8 a | n.d | 0.2 ± 0.0 j,k | 0.3 ± 0.0 f | 0.2 ± 0.0 e | 93.6 ± 3.5 f |
| Tribe Phoeniceae | ||||||||||
| 12 | Phoenix canariensis | 0.7 ± 0.0 j,k,l,m | 2.3 ± 0.2 j,k,l,m | 0.3 ± 0.0 i,j | n.d | n.d | 0.6 ± 0.0 f,g,h,i | 1.7 ± 0.1 e,f | 0.3 ± 0.0 e | 88.4 ± 3.5 f,g |
| 13A | P. dactylifera var. Deglet Nour | 1.0 ± 0.1 i,j,k,l,m | 4.6 ± 0.8 j,k,l | 3.5 ± 0.9 d | 1.2 ± 0.2 i,j | 3.0 ± 0.2 c | 0.3 ± 0.0 i,j,k | 0.5 ± 0.0 f | 0.6 ± 0.0 e | 134.1 ± 5.0 d |
| 13B | P. dactylifera var. Medjool | 0.4 ± 0.0 l,m | 11.9 ± 1.9 f,g,h | 14.3 ± 1.7 b | 0.4 ± 0.0 j | 0.5 ± 0.0 e,f | 0.9 ± 0.0 f | 0.7 ± 0.0 f | 1.9 ± 0.3 c,d | 144.3 ± 4.2 d |
| 14 | P. reclinata | 0.6 ± 0.0 k,l,m | 0.6 ± 0.0 m | 1.8 ± 0.0 f,g,h,i | 0.2 ± 0.0 j | 0.1 ± 0.0 f | 0.5 ± 0.0 g,h,i,j | 0.3 ± 0.0 f | 0.9 ± 0.0 c,d,e | 89.0 ± 1.7 f,g |
| Tribe Sabaleae | ||||||||||
| 15 | Sabal bermudana | 2.9 ± 0.1 e,f,g | 10.6 ± 0.9 g,h | 3.5 ± 0.6 d | 1.2 ± 0.3 i,j | 0.3 ± 0.0 e,f | 1.7 ± 0.2 d | 14.5 ± 0.0 c | 0.4 ± 0.0 e | 159.2 ± 12.3 c |
| 16 | S. minor | 3.4 ± 0.6 e | 49.6 ± 1.9 b | 5.6 ± 0.1 c | 2.8 ± 0.7 e,f,g | 2.8 ± 0.3 c | 0.8 ± 0.1 f,g | 24.6 ± 1.5 b | 1.1 ± 0.2 c,d,e | 202.0 ± 4.5 b |
| 17 | S. palmetto | 3.2 ± 0.3 e,f | 58.2 ± 2.1 a | 5.7 ± 0.0 c | 3.0 ± 0.2 e,f | 0.3 ± 0.0 e,f | 0.9 ± 0.0 f | 55.6 ± 3.8 a | 0.4 ± 0.0 e | 246.0 ± 5.8 a |
| 18 | S. domingensis | 2.2 ± 0.3 f,g,h | 14.3 ± 1.2 e,f,g | 3.4 ± 0.6 d,e | 5.6 ± 0.4 d | 0.1 ± 0.0 f | 1.3 ± 0.0 e | 11.4 ± 0.1 d | 1.3 ± 0.2 c,d,e | 91.5 ± 2.2 f |
| Tribe Trachycarpeae | ||||||||||
| 19A | Chamaerops humilis | 1.8 ± 0.1 g,h,i,j | 4.9 ± 0.0 i,j,k | 1.7 ± 0.1 f,g,h,i | 12.0 ± 0.3 a | 0.2 ± 0.0 f | 0.3 ± 0.0 i,j,k | 0.2 ± 0.0 f | 0.1 ± 0.0 e | 57.3 ± 1.5 i,j,k |
| 19B | C. humilis | 0.2 ± 0.0 m | 13.5 ± 0.7 e,f,g | 5.2 ± 0.4 c | 7.2 ± 0.4 c | 0.3 ± 0.0 e,f | 0.4 ± 0.0 h,i,j,k | 0.2 ± 0.0 f | 0.3 ± 0.0 e | 80.8 ± 3.2 g,h |
| 19C | C. humilis | 0.5 ± 0.0 k,l,m | 8.5 ± 0.6 h,i | 2.8 ± 0.3 d,e,f,g | 2.4 ± 0.1 f,g,h | 0.6 ± 0.0 e,f | 0.1 ± 0.0 k | 0.7 ± 0.0 f | 0.6 ± 0.0 e | 50.4 ± 1.2 k,l,m,n |
| 19D | C. humilis | 0.7 ± 0.0 j,k,l,m | 5.5 ± 0.5 i,j | 1.4 ± 0.3 g,h,i,j | 7.0 ± 0.5 c | 0.2 ± 0.0 f | 0.2 ± 0.0 j,k | 0.1 ± 0.0 f | 0.2 ± 0.0 e | 52.2 ± 2.4 j,k,l,m |
| 20 | Livistona chinensis | 1.6 ± 0.2 h,i,j,k | 0.1 ± 0.0 m | 0.8 ± 0.0 h,i,j | n.d | n.d | 0.3 ± 0.0 i,j,k | 0.2 ± 0.0 f | 0.3 ± 0.0 e | 140.2 ± 3.9 d |
| 21 | L. fulva | 0.6 ± 0.3 k,l,m | 0.6 ± 0.0 m | 0.9 ± 0.1 h,i,j | n.d | n.d | 0.4 ± 0.0 h,i,j,k | 0.1 ± 0.0 f | 0.4 ± 0.0 e | 39.0 ± 0.8 o |
| 22 | L. saribus | 1.9 ± 0.4 g,h,i | 0.9 ± 0.0 l,m | 1.8 ± 0.2 f,g,h,i | n.d | n.d | 0.6 ± 0.0 f,g,h,i | 0.4 ± 0.0 f | 0.7 ± 0.0 d,e | 43.3 ± 0.6 m,n,o |
| 23 | Trachycarpus fortunei | 9.2 ± 0.4 b | 14.5 ± 0.5 e,f | 0.7 ± 0.0 h,i,j | 12.9 ± 0.7 a | 1.3 ± 0.2 d,e | 1.5 ± 0.3 d,e | 0.3 ± 0.0 f | 1.2 ± 0.0 c,d,e | 65.3 ± 1.2 i |
| 24 | Washingtonia robusta | 5.0 ± 0.3 d | 15.8 ± 2.1 e | 1.2 ± 0.2 h,i,j | 9.3 ± 0.5 b | 0.9 ± 0.0 e,f | 3.7 ± 0.4 b | 0.1 ± 0.0 f | 0.5 ± 0.0 e | 61.9 ± 2.4 i,j |
| Code | Species | Resveratrol | Piceatannol | Pinocembrin | Genistein | Apigenin | Phloretin | Luteolin | Sakuranetin | Cianidin | Epicatechin (-) |
| m/z precursor ion | 227.07137 | 243.06628 | 255.06628 | 271.0601 | 271.0601 | 275.0914 | 285.04046 | 285.07685 | 287.05501 | 291.08631 | |
| Subfamily Arecoideae | |||||||||||
| Tribe Areceae | |||||||||||
| 1 | Archontophoenix cunninghamiana | - | - | - | - | - | - | + | - | - | + |
| 2 | Chambeyronia macrocarpa | - | - | - | - | - | - | + | - | - | + |
| 3 | Dypsis lutescens | - | - | - | - | - | - | + | - | - | - |
| 4 | Howea belmoreana | - | - | - | - | - | - | - | - | - | + |
| 5A | H. forsteriana | - | - | - | - | - | - | - | - | - | + |
| 5B | H. forsteriana | - | - | - | - | - | - | - | - | - | - |
| Tribe Chamaedoreeae | |||||||||||
| 6 | Chamaedorea microspadix | - | - | - | - | - | - | - | - | - | - |
| 7 | C. oblongata | - | - | - | - | - | - | + | - | - | - |
| Tribe Cocoseae | |||||||||||
| 8 | Butia capitata | - | - | - | - | - | - | - | - | - | + |
| 9 | Cocos nucifera | - | - | - | - | - | - | - | - | - | - |
| 10 | Syagrus romanzofianna | + | + | - | - | - | + | - | + | - | + |
| Subfamily Coryphoideae | |||||||||||
| Tribe Caryoteae | |||||||||||
| 11 | Arenga engleri | - | - | - | - | - | + | - | - | - | + |
| Tribe Phoeniceae | |||||||||||
| 12 | Phoenix canariensis | - | - | - | - | - | - | + | - | - | + |
| - | |||||||||||
| 13A | P. dactylifera var. Deglet Nour | - | - | - | - | - | - | - | - | - | + |
| 13B | P. dactylifera var. Medjool | - | - | - | - | - | - | + | - | - | + |
| 14 | P. reclinata | - | - | - | + | + | - | - | - | - | + |
| Tribe Sabaleae | |||||||||||
| 15 | Sabal bermudana | - | - | - | - | - | - | + | - | - | + |
| 16 | S. minor | - | - | - | - | - | - | + | - | - | + |
| 17 | S. palmetto | - | - | - | - | - | - | + | - | - | + |
| 18 | S. domingensis | - | - | - | + | + | - | - | - | - | + |
| Tribe Trachycarpeae | |||||||||||
| 19A | Chamaerops humilis | - | - | - | - | - | - | - | - | - | + |
| 19B | C. humilis | - | - | - | - | - | - | + | - | - | + |
| 19C | C. humilis | - | - | - | - | - | - | - | - | - | - |
| 19D | C. humilis | - | - | - | - | - | - | - | - | - | - |
| 20 | Livistona chinensis | - | - | - | - | - | - | - | - | - | + |
| 21 | L. fulva | - | + | - | - | - | + | - | - | + | + |
| 22 | L. saribus | - | - | + | + | + | - | - | - | - | + |
| 23 | Trachycarpus fortunei | - | - | - | - | - | - | - | - | - | + |
| 24 | Washingtonia robusta | - | - | - | - | - | - | - | - | - | - |
| Code | Species | Delphinidine | Epigallocatechin (-) | Gallocatechin (-) | Petunidine | Bilobalide | Malvidine | Ferulic Acid Hexoside | Piceid (resveratrol-3-O-beta-D-glucopyranoside) | Apigenin-6-C-glucoside (Isovitexin) | Phloridzin |
| m/z precursor ion | 303.04993 | 305.06668 | 307.08123 | 317.06558 | 325.09289 | 331.08123 | 355.10346 | 391.13874 | 431.09837 | 435.12967 | |
| Subfamily Arecoideae | |||||||||||
| Tribe Areceae | |||||||||||
| 1 | Archontophoenix cunninghamiana | - | - | - | - | - | - | + | - | - | - |
| 2 | Chambeyronia macrocarpa | - | - | - | - | - | - | + | - | - | - |
| 3 | Dypsis lutescens | - | - | - | - | - | - | + | - | - | - |
| 4 | Howea belmoreana | - | - | - | - | - | - | - | - | - | - |
| 5A | H. forsteriana | - | - | - | - | - | - | + | - | - | - |
| 5B | H. forsteriana | - | - | - | - | - | - | - | - | - | - |
| Tribe Chamaedoreeae | |||||||||||
| 6 | Chamaedorea microspadix | - | - | - | - | - | - | - | - | - | - |
| 7 | C. oblongata | - | - | - | - | - | - | - | - | - | - |
| Tribe Cocoseae | |||||||||||
| 8 | Butia capitata | - | - | - | - | - | - | - | - | - | - |
| 9 | Cocos nucifera | - | - | - | - | - | - | - | - | - | - |
| 10 | Syagrus romanzofianna | - | - | + | - | - | - | - | - | - | + |
| Subfamily Coryphoideae | |||||||||||
| Tribe Caryoteae | |||||||||||
| 11 | Arenga engleri | - | - | + | - | - | - | - | + | - | + |
| Tribe Phoeniceae | |||||||||||
| 12 | Phoenix canariensis | - | - | - | - | - | - | - | - | - | - |
| 13A | P. dactylifera var. Deglet Nour | - | - | - | - | - | - | - | - | - | - |
| 13B | P. dactylifera var. Medjool | - | - | - | - | - | - | - | - | - | - |
| 14 | P. reclinata | - | - | - | - | + | - | - | - | - | + |
| Tribe Sabaleae | |||||||||||
| 15 | Sabal bermudana | - | - | - | - | - | - | - | - | - | - |
| 16 | S. minor | - | - | - | - | - | - | - | - | - | - |
| 17 | S. palmetto | - | - | - | - | - | - | - | - | - | - |
| 18 | S. domingensis | + | + | + | + | + | - | - | - | + | - |
| Tribe Trachycarpeae | |||||||||||
| 19A | Chamaerops humilis | - | - | - | - | - | - | + | - | - | - |
| 19B | C. humilis | - | - | - | - | - | - | + | - | - | - |
| 19C | C. humilis | - | - | - | - | - | - | - | - | - | - |
| 19D | C. humilis | - | - | - | - | - | - | - | - | - | - |
| 20 | Livistona chinensis | - | - | - | - | - | - | - | - | - | - |
| 21 | L. fulva | + | + | - | + | - | - | - | - | + | - |
| 22 | L. saribus | - | + | + | + | + | + | - | - | - | + |
| 23 | Trachycarpus fortunei | - | + | + | - | - | - | - | - | - | + |
| 24 | Washingtonia robusta | - | + | + | - | - | - | - | - | + | + |
| Code | Species | Quercetin-3-O-rhamnoside (Quercitrin) | Isorhamnetin-3-O-Glucoside | Lithospermic Acid | Procyanidin B1 | Kaempferol-3-O-Rutinoside | Pelargonidine | Eriocitrin | Hesperidin | Delphinidin-3-O-(6-O-p-coumaroyl)-Glucoside | Isohamnetin-3-O-Rutinoside |
| m/z precursor ion | 447.09328 | 479.11840 | 537.10385 | 577.13515 | 593.15119 | 595.16575 | 595.16684 | 609.18249 | 611.13953 | 623.16176 | |
| Subfamily Arecoideae | |||||||||||
| Tribe Areceae | |||||||||||
| 1 | Archontophoenix cunninghamiana | - | - | - | - | - | - | - | - | - | - |
| 2 | Chambeyronia macrocarpa | - | - | - | - | - | - | - | - | - | - |
| 3 | Dypsis lutescens | - | - | - | - | - | - | - | - | - | - |
| 4 | Howea belmoreana | - | - | - | - | - | - | - | - | - | - |
| 5A | H. forsteriana | - | - | - | - | - | - | - | - | - | - |
| 5B | H. forsteriana | - | - | - | - | - | - | - | - | - | - |
| Tribe Chamaedoreeae | |||||||||||
| 6 | Chamaedorea microspadix | - | - | - | - | - | - | - | - | - | - |
| 7 | C. oblongata | - | - | - | - | - | - | - | - | - | - |
| Tribe Cocoseae | |||||||||||
| 8 | Butia capitata | - | - | - | - | - | - | - | - | - | - |
| 9 | Cocos nucifera | - | - | - | - | - | - | - | - | - | - |
| 10 | Syagrus romanzofianna | - | - | - | + | - | - | - | + | - | - |
| Subfamily Coryphoideae | |||||||||||
| Tribe Caryoteae | |||||||||||
| 11 | Arenga engleri | + | - | - | + | - | - | - | + | - | - |
| Tribe Phoeniceae | |||||||||||
| 12 | Phoenix canariensis | - | - | - | + | - | - | - | - | - | - |
| 13A | P. dactylifera var. Deglet Nour | - | - | - | - | - | - | - | - | - | - |
| 13B | P. dactylifera var. Medjool | - | - | - | - | - | - | - | - | - | - |
| 14 | P. reclinata | - | - | - | - | + | + | - | - | - | - |
| Tribe Sabaleae | |||||||||||
| 15 | Sabal bermudana | - | - | + | - | - | - | - | - | - | - |
| 16 | S. minor | - | - | - | - | - | - | - | - | - | - |
| 17 | S. palmetto | - | - | - | - | - | - | - | - | - | - |
| 18 | S. domingensis | - | - | - | + | - | - | - | - | - | - |
| Tribe Trachycarpeae | |||||||||||
| 19A | Chamaerops humilis | - | - | - | - | - | - | - | - | - | - |
| 19B | C. humilis | - | - | - | - | - | - | - | - | - | - |
| 19C | C. humilis | - | - | - | - | - | - | - | - | - | - |
| 19D | C. humilis | - | - | - | - | - | - | - | - | - | - |
| 20 | Livistona chinensis | - | - | + | - | + | - | - | - | - | - |
| 21 | L. fulva | + | + | - | + | + | + | + | + | + | - |
| 22 | L. saribus | + | - | - | + | + | + | - | - | - | - |
| 23 | Trachycarpus fortunei | + | - | - | + | - | + | + | + | - | + |
| 24 | Washingtonia robusta | + | - | + | + | + | - | - | + | - | + |
| Sample Code | Species | Common Name | Sample Location | Geographical Coordinates | Collection Date |
|---|---|---|---|---|---|
| Subfamily Arecoideae | |||||
| Tribe Areceae | |||||
| 1 | Archontophoenix cunninghamiana H. Wendl. & Drude | Bangalow palm | Botanic gardens of wood Rui Vieira, Portugal | 32.662316, −16.894604 | 2020 |
| 2 | Chambeyronia macrocarpa (Brongn.) Vieill. Ex Becc. | Red leaf palm | Botanic gardens of wood Rui Vieira, Portugal | 32.662316, −16.894604 | 2020 |
| 3 | Dypsis lutescens (H. Wendl.) Beentje & J.Dransf. (H. Wendl.) Beentje & J. Dransf. (Syn. Chrysalidocarpus lutescens H. Wendl.) | Yellow cane palm Yellow areca palm | Botanic gardens of wood Rui Vieira, Portugal | 32.662316, −16.894604 | 2020 |
| 4 | Howea belmoreana (C. Moore & F. Muell.) Becc. | Curly palm | Botanic gardens of wood Rui Vieira, Portugal | 32.662316, −16.894604 | 2020 |
| 5A | Howea forsteriana Becc. | Kentia palm | Botanic gardens of wood Rui Vieira, Portugal | 32.662316, −16.894604 | 2020 |
| 5B | H. forsteriana Bec | Kentia palm | Botanischer Garten Berlin−Dahlem 3550, Germany | 52.456684, 13.304710 | 2020 |
| Tribe Chamaedoreeae | |||||
| 6 | Chamaedorea microspadix Burret | Hardy bamboo palm | Palm Society, San Leandro, California | 37.727389, −122.180107 | 2021 |
| 7 | Chamaedorea oblongata Mart. VU bk. | Caquib, palmilla, chate | Botanic garden of the University, Bulgaria | 42.697102, 23.334565 | 2018 |
| Tribe Cocoseae | |||||
| 8 | Butia capitata (Mart.) Becc. | Butià, Jelly palm | Botanic gardens of wood Rui Vieira, Portugal | 32.662316, −16.894604 | 2020 |
| 9 | Cocos nucifera L. | Coconut palm | Malaga, Spain (Purchased) | 2021 | |
| 10 | Syagrus romanzoffiana (Cham.) Glassman | Queen palm | University of Almería gardens, Spain | 36.829694, −2.404185 | 2021 |
| Subfamily Coryphoideae | |||||
| Tribe Caryoteae | |||||
| 11 | Arenga engleri Becc. | Taiwan sugar palm | University Botanic Garden of Valencia, Spain | 39.475663, −0.386351 | 2021 |
| Tribe Phoeniceae | |||||
| 12 | Phoenix canariensis Chabaud | Canary Island date palm | Botanic gardens of wood Rui Vieira, Portugal | 32.662316, −16.894604 | 2020 |
| 13A | Phoenix dactylifera L. var. Deglet Nour | Date palm | Algeria (Purchased) | 2021 | |
| 13B | P. dactylifera L. var. Medjool | Date palm | Spain (Purchased) | 2021 | |
| 14 | Phoenix reclinata Jacq. | Wild date palm | University Botanic Garden of Valencia, Spain | 39.475663, −0.386351 | 2021 |
| Tribe Sabaleae | |||||
| 15 | Sabal bermudana L. H. Bailey ex Knuth | Bermuda palmetto | University Botanic gardens, Bulgaria | 42.697102, 23.334565 | 2018 |
| 18 | Sabal domingensis Becc. | Hispaniola palmetto | University Botanic Garden of Valencia, Spain | 39.475663, −0.386351 | 2021 |
| 16 | Sabal minor (Jacq.) Pers. | Dwarf palmetto | University Botanic gardens, Bulgaria | 42.697102, 23.334565 | 2019 |
| 17 | Sabal palmetto (Walt.) Lodd. | Cabbage palm | Florida, Miami, Coral Gables, USA | 25.294750, −76.188889 | 2021 |
| Tribe Trachycarpeae | |||||
| 19A | Chamaerops humilis L. | Mediterranean fan palm | Bulgaria, University Botanic gardens | 42.697102, 23.334565 | 2020 |
| 19B | C. humilis L. | Mediterranean fan palm | Bulgaria, University Botanic gardens | 42.697102, 23.334565 | 2020 |
| 19C | C. humilis L. | Mediterranean fan palm | Pernambuco, Brazil | −969.12055107, −36.6077802 | 2021 |
| 19D | C. humilis L. | Mediterranean fan palm | El Toyo, Almería, Spain | 36.836508, −2.326255 | 2021 |
| 20 | Livistona chinensis (Jacq.) R.Br. ex Mart. sf. | Chinese fan palm | Bulgaria, University Botanic gardens | 42.697102, 23.334565 | 2020 |
| 21 | Livistona fulva Rodd | Blackdown fan palm | University Botanic Garden of Valencia, Spain | 39.475663, −0.386351 | 2020 |
| 22 | Livistona saribus (Lour.) Merr. ex A.Chev. | Taraw palm | University Botanic Garden of Valencia, Spain | 39.475663, −0.386351 | 2022 |
| 23 | Trachycarpus fortunei (Hook.) H.Wendl | Windmill palm | University Botanic Garden of Valencia, Spain | 39.475663, −0.386351 | 2022 |
| 24 | Washingtonia robusta H. Wendl. | Washington fan palm | University Botanic Garden of Valencia, Spain | 39.475663, −0.386351 | 2021 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rincón-Cervera, M.Á.; Lahlou, A.; Chileh-Chelh, T.; Lyashenko, S.; López-Ruiz, R.; Guil-Guerrero, J.L. Arecaceae Seeds Constitute a Healthy Source of Fatty Acids and Phenolic Compounds. Plants 2023, 12, 226. https://doi.org/10.3390/plants12020226
Rincón-Cervera MÁ, Lahlou A, Chileh-Chelh T, Lyashenko S, López-Ruiz R, Guil-Guerrero JL. Arecaceae Seeds Constitute a Healthy Source of Fatty Acids and Phenolic Compounds. Plants. 2023; 12(2):226. https://doi.org/10.3390/plants12020226
Chicago/Turabian StyleRincón-Cervera, Miguel Ángel, Abdallah Lahlou, Tarik Chileh-Chelh, Svetlana Lyashenko, Rosalía López-Ruiz, and José Luis Guil-Guerrero. 2023. "Arecaceae Seeds Constitute a Healthy Source of Fatty Acids and Phenolic Compounds" Plants 12, no. 2: 226. https://doi.org/10.3390/plants12020226
APA StyleRincón-Cervera, M. Á., Lahlou, A., Chileh-Chelh, T., Lyashenko, S., López-Ruiz, R., & Guil-Guerrero, J. L. (2023). Arecaceae Seeds Constitute a Healthy Source of Fatty Acids and Phenolic Compounds. Plants, 12(2), 226. https://doi.org/10.3390/plants12020226