


Efficacy of Carbon Nanodots and Manganese Ferrite (MnFe2O4) Nanoparticles in Stimulating Growth and Antioxidant Activity in Drought-Stressed Maize Inbred Lines

 ,
,
Abstract
1. Introduction
2. Results
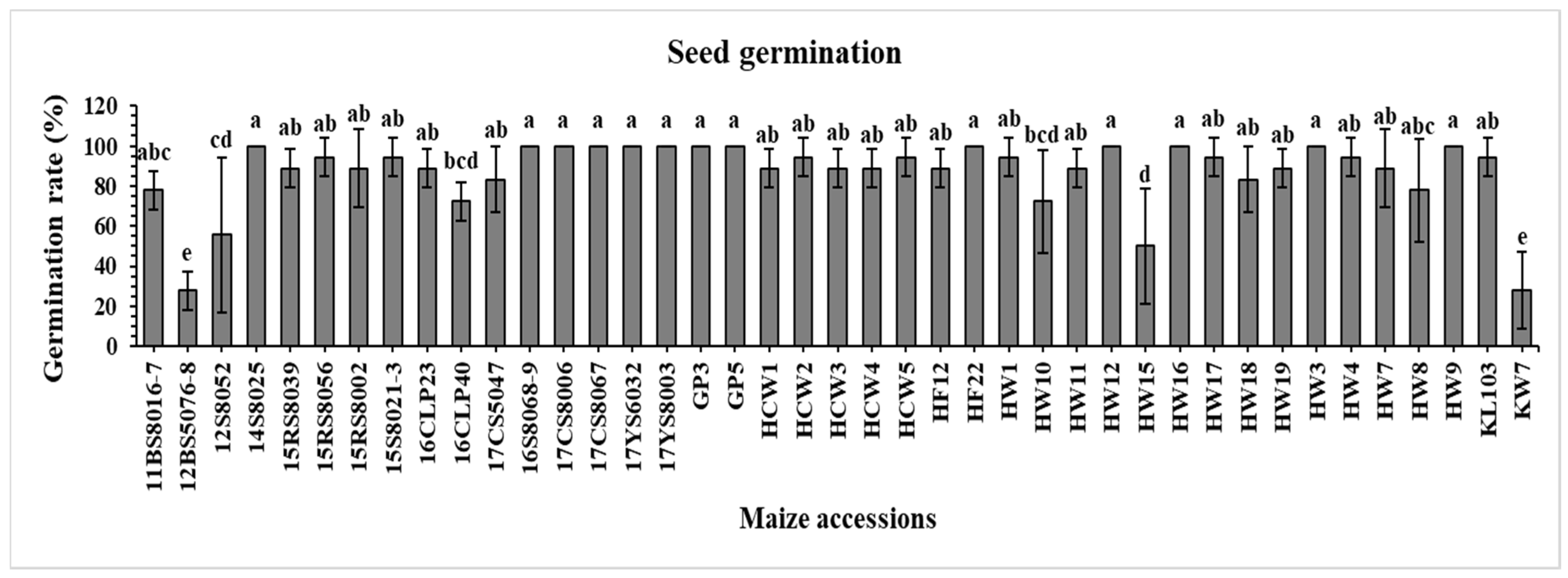
2.1. Seed Germination
2.2. Effects of CND and MnFe2O4 NP on Plant Growth and Morphology under Drought Stress
2.2.1. Leaf Blade Length and Width
2.2.2. Shoot and Root Length
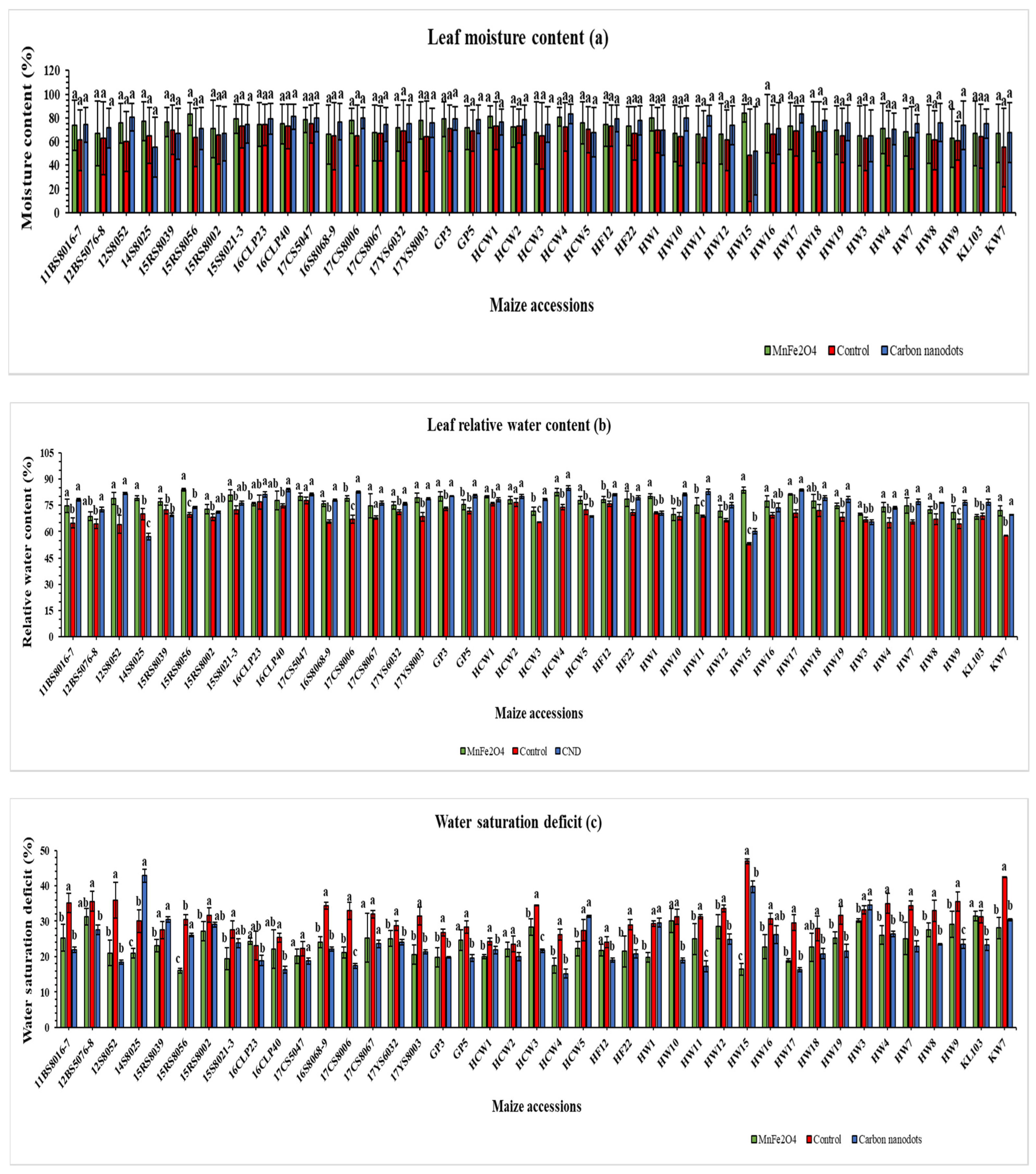
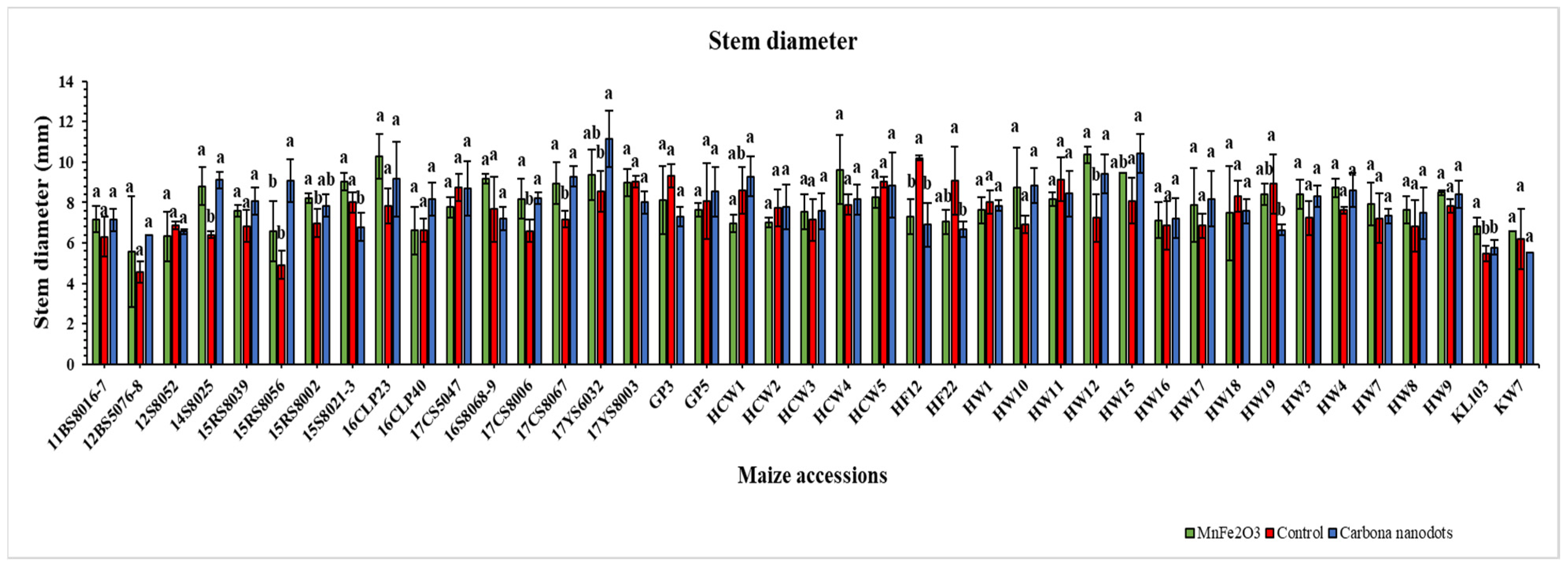
2.2.3. Leaf Water Status and Stem Diameter
2.2.4. Root and Shoot Fresh Weight
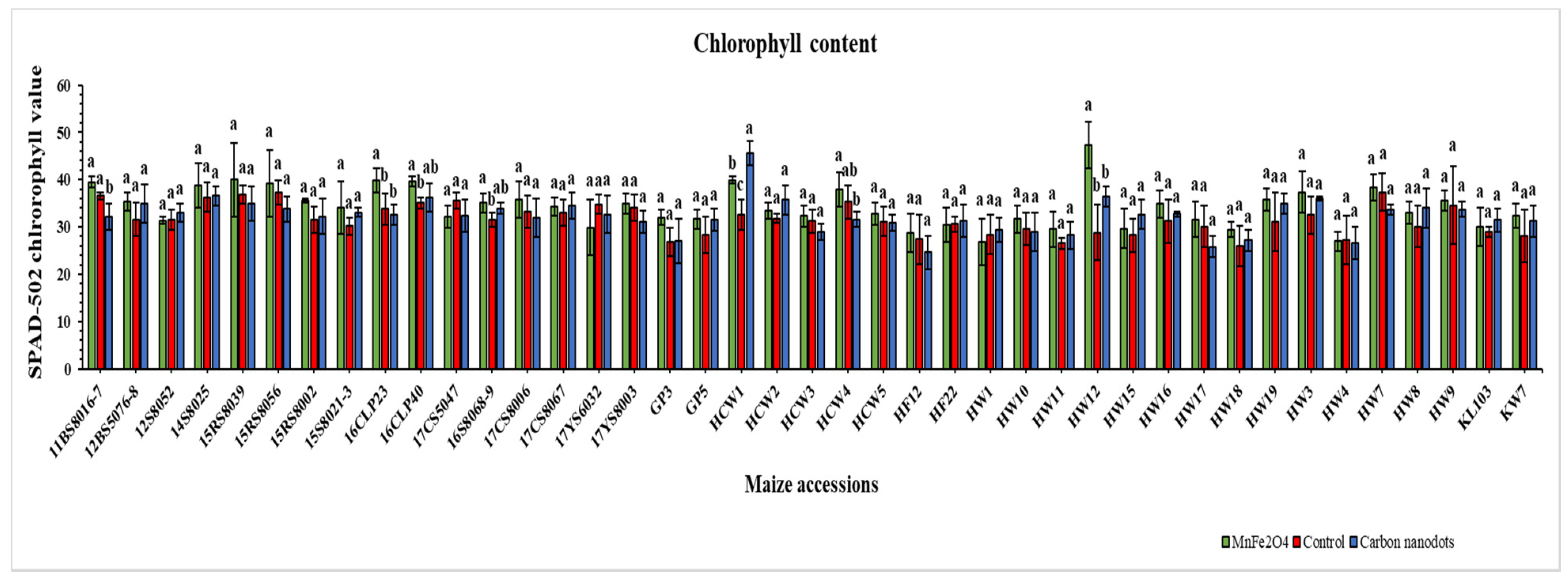
2.3. Chlorophyll Content
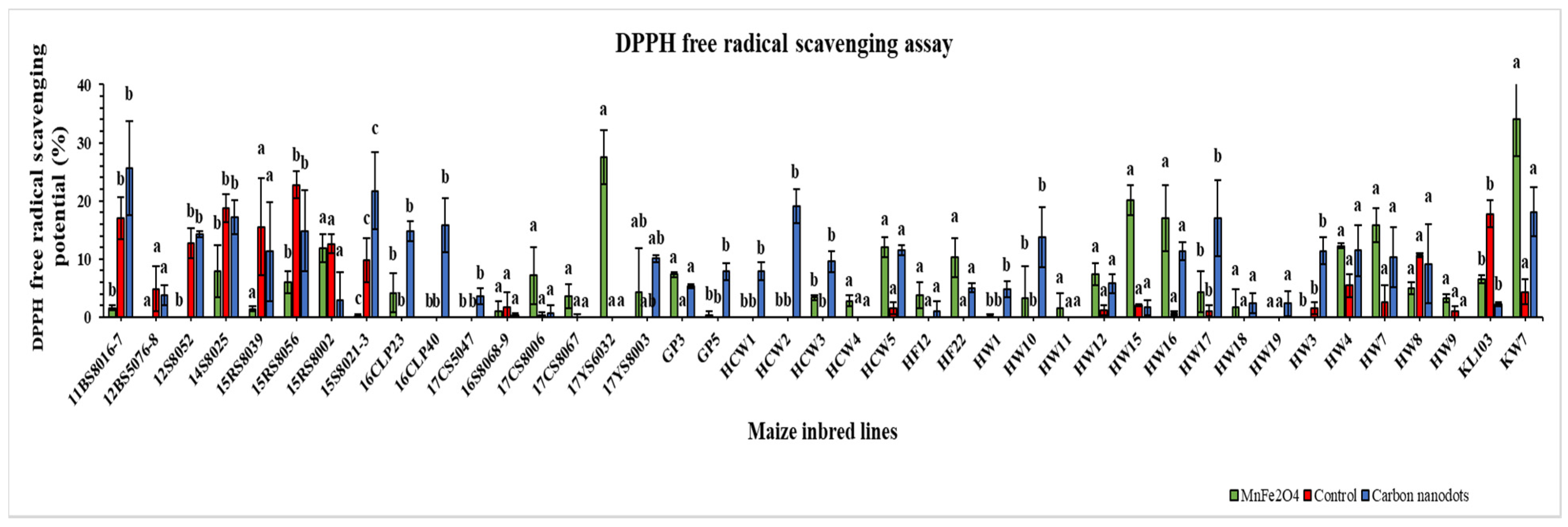
2.4. DPPH Free Radical Scavenging Activity
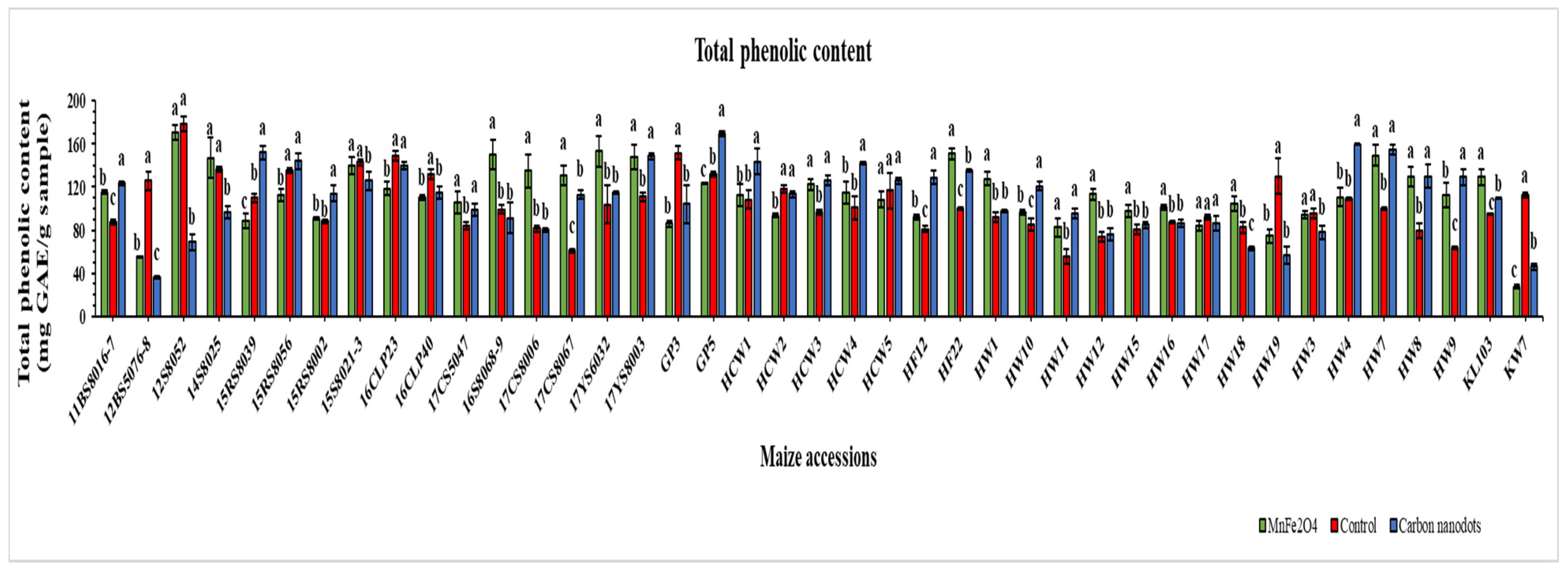
2.5. Total Phenolic Contents (TPC)
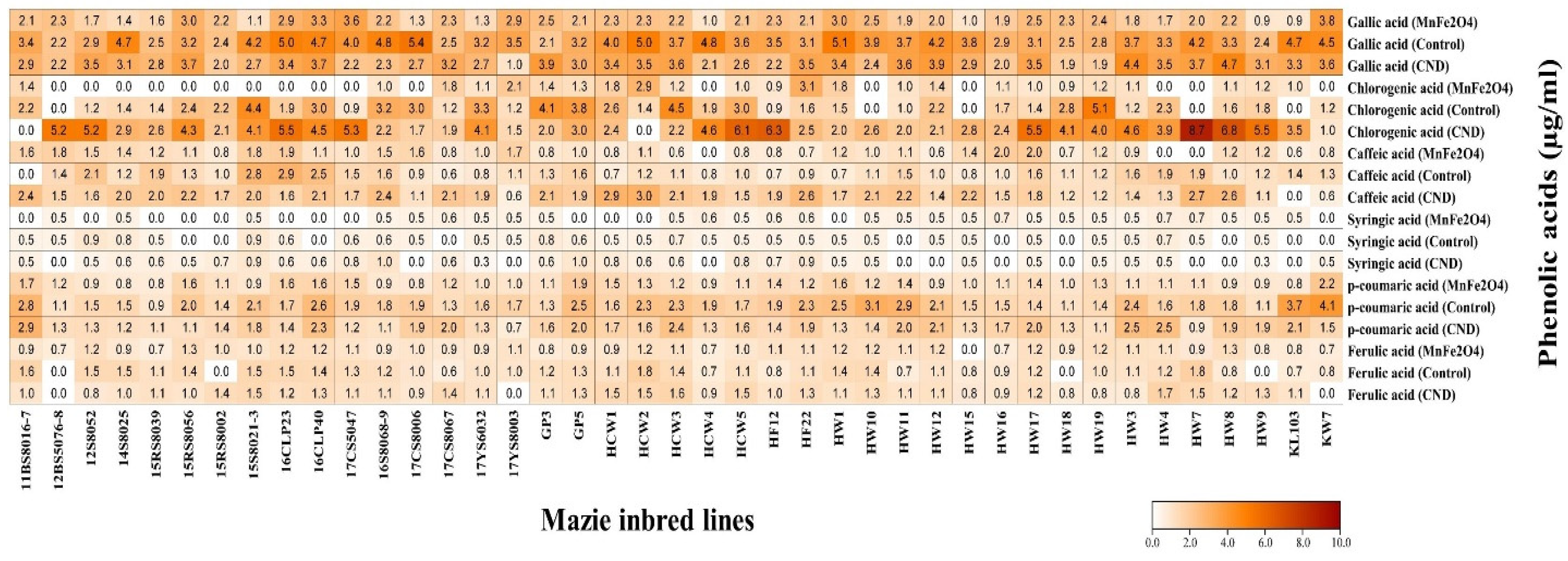
2.6. HPLC-UV Analysis of Phenolic Compounds
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Collection of Experimental Material
5.2. Seedbed Media Characteristics
5.3. Experimental Design and Crop Husbandry
5.4. Data Collection
5.5. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Jia, X.; Mao, K.; Wang, P.; Wang, Y.; Jia, X.; Huo, L.; Sun, X.; Che, R.; Gong, X.; Ma, F. Overexpression of MdATG8i improves water use efficiency in transgenic apple by modulating photosynthesis, osmotic balance, and autophagic activity under moderate water deficit. Hortic. Res. 2021, 8, 81. [Google Scholar] [CrossRef] [PubMed]
- Mansoor, S.; Kour, N.; Manhas, S.; Zahid, S.; Wani, O.A.; Sharma, V.; Wijaya, L.; Alyemeni, M.N.; Alsahli, A.A.; El-Serehy, H.A.; et al. Biochar as a tool for effective management of drought and heavy metal toxicity. Chemosphere 2021, 271, 129458. [Google Scholar] [CrossRef] [PubMed]
- Balbaa, M.G.; Osman, H.T.; Kandil, E.E.; Javed, T.; Lamlom, S.F.; Ali, H.M.; Kalaji, H.M.; Wróbel, J.; Telesiñski, A.; Brysiewicz, A.; et al. Determination of morpho-physiological and yield traits of maize inbred lines (Zea mays L.) under optimal and drought stress conditions. Front. Plant Sci. 2022, 13, 959203. [Google Scholar] [CrossRef] [PubMed]
- Liu, S.; Zenda, T.; Dong, A.; Yang, Y.; Wang, N.; Duan, H. Global transcriptome and weighted gene co-expression network analyses of growth-stage-specific drought stress responses in maize. Front. Genet. 2021, 12, 645443. [Google Scholar] [CrossRef]
- Thirunavukkarasu, N.; Sharma, R.; Singh, N.; Shiriga, K.; Mohan, S.; Mittal, S.; Mittal, S.; Mallikarjuna, M.G.; Rao, A.R.; Dash, P.K.; et al. Genomewide expression and functional interactions of genes under drought stress in maize. Int. J. Genom. 2017, 2017, 14. [Google Scholar] [CrossRef] [PubMed]
- Hu, H.; Xiong, L. Genetic engineering and breeding of drought-resistant crops. Annu. Rev. Plant Biol. 2014, 65, 715–741. [Google Scholar] [CrossRef]
- Li, Z.; Su, X.; Chen, Y.; Fan, X.; He, L.; Guo, J.; Wang, Y.; Yang, Q. Melatonin improves drought resistance in maize seedlings by enhancing the antioxidant system and regulating abscisic acid metabolism to maintain stomatal opening under PEG-induced drought. J. Plant Biol. 2021, 64, 299–312. [Google Scholar] [CrossRef]
- Shemi, R.; Wang, R.; Gheith, E.S.M.S.; Hussain, H.A.; Hussain, S.; Irfan, M.; Cholidah, L.; Zhang, K.; Zhang, S.; Wang, L. Effects of salicylic acid, zinc and glycine betaine on morpho-physiological growth and yield of maize under drought stress. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Anjum, S.A.; Farooq, M.; Wang, L.C.; Xue, L.L.; Wang, S.G.; Wang, L.; Zhang, S.; Chen, M. Gas exchange and chlorophyll synthesis of maize cultivars are enhanced by exogenously-applied glycinebetaine under drought conditions. Plant Soil Environ. 2011, 57, 326–331. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.A. Global synthesis of drought effects on maize and wheat production. PLoS ONE 2016, 11, e0156362. [Google Scholar] [CrossRef]
- Ahmad, A.; Aslam, Z.; Hussain, S.; Bibi, A.; Khaliq, A.; Javed, T.; Alotaibi, S.S.; Kalaji, H.M.; Telesiński, A.; Iwai, C.B.; et al. Rice straw vermicompost enriched with cellulolytic microbes ameliorate the negative effect of drought in wheat through modulating the morpho-physiological attributes. Front. Environ. Sci. 2022, 10, 902999. [Google Scholar] [CrossRef]
- Anjum, S.A.; Ashraf, U.; Tanveer, M.; Khan, I.; Hussain, S.; Shahzad, B.; Zohaib, A.; Abbas, F.; Saleem, M.F.; Ali, I.; et al. Drought induced changes in growth, osmolyte accumulation and antioxidant metabolism of three maize hybrids. Front. Plant Sci. 2017, 8, 69. [Google Scholar] [CrossRef] [PubMed]
- Keenan, T.; Sabate, S.; Gracia, C. The importance of mesophyll conductance in regulating forest ecosystem productivity during drought periods. Glob. Change Biol. 2010, 16, 1019–1034. [Google Scholar] [CrossRef]
- Raja, V.; Qadir, S.U.; Alyemeni, M.N.; Ahmad, P. Impact of drought and heat stress individually and in combination on physio-biochemical parameters, antioxidant responses, and gene expression in Solanum lycopersicum. 3 Biotech 2020, 10, 1–18. [Google Scholar] [CrossRef]
- Anjum, S.A.; Tanveer, M.; Ashraf, U.; Hussain, S.; Shahzad, B.; Khan, I.; Wang, L. Effect of progressive drought stress on growth, leaf gas exchange, and antioxidant production in two maize cultivars. Environ. Sci. Pollut. Res. 2016, 23, 17132–17141. [Google Scholar] [CrossRef] [PubMed]
- Cruz De Carvalho, M.H. Drought stress and reactive oxygen species: Production, scavenging and signaling. Plant Signal. Behav. 2008, 3, 156–165. [Google Scholar] [CrossRef]
- Wu, Q.S.; Xia, R.X.; Zou, Y.N. Improved soil structure and citrus growth after inoculation with three arbuscular mycorrhizal fungi under drought stress. Eur. J. Soil Biol. 2008, 44, 122–128. [Google Scholar] [CrossRef]
- Liu, H.; Song, S.; Zhang, H.; Li, Y.; Niu, L.; Zhang, J.; Wang, W. Signaling transduction of ABA, ROS, and Ca2+ in plant stomatal closure in response to drought. Int. J. Mol. Sci. 2022, 23, 14824. [Google Scholar] [CrossRef]
- Kaur, S.; Seem, K.; Duhan, N.; Kumar, S.; Kaundal, R.; Mohapatra, T. Transcriptome and physio-biochemical profiling reveals differential responses of rice cultivars at reproductive-stage drought stress. Int. J. Mol. Sci. 2023, 24, 1002. [Google Scholar] [CrossRef]
- Sun, Y.; Miao, F.; Wang, Y.; Liu, H.; Wang, X.; Wang, H.; Guo, J.; Shao, R.; Yang, Q. L-Arginine alleviates the reduction in photosynthesis and antioxidant activity induced by drought stress in maize seedlings. Antioxidants 2023, 12, 482. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive oxygen species, oxidative damage, and antioxidative defense mechanism in plants under stressful conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef]
- Rabêlo, V.M.; Magalhães, P.C.; Bressanin, L.A.; Carvalho, D.T.; Reis, C.O.D.; Karam, D.; Doriguetto, A.C.; Santos, M.H.D.; Filho, P.R.D.S.S.; Souza, T.C.D. The foliar application of a mixture of semisynthetic chitosan derivatives induces tolerance to water deficit in maize, improving the antioxidant system and increasing photosynthesis and grain yield. Sci. Rep. 2019, 9, 8164. [Google Scholar] [CrossRef]
- Naikoo, M.I.; Dar, M.I.; Raghib, F.; Jaleel, H.; Ahmad, B.; Raina, A.; Khan, F.A.; Naushin, F. Role and regulation of plants phenolics in abiotic stress tolerance: An overview. In Plant Signaling Molecules Role and Regulation under Stressful Environments; Elsevier Inc.: Amsterdam, The Netherlands, 2019; pp. 157–168. [Google Scholar]
- Selmar, D. Potential of salt and drought stress to increase pharmaceutical significant secondary compounds in plants. Landbauforsch. Volkenrode 2008, 58, 139–144. [Google Scholar]
- Robards, K.; Antolovich, M. Analytical chemistry of fruit bioflavonoids. A review. Analyst 1997, 122, 11–34. [Google Scholar] [CrossRef]
- Khan, Z.; Upadhyaya, H. Impact of nanoparticles on abiotic stress responses in plants: An overview. In Nanomaterials in Plants, Algae and Microorganisms: Concepts and Controversies: Volume 2; Elsevier Inc.: Amsterdam, The Netherlands, 2018; pp. 305–322. [Google Scholar]
- Dimkpa, C.O.; Singh, U.; Bindraban, P.S.; Elmer, W.H.; Gardea-torresdey, J.L.; White, J.C. Zinc oxide nanoparticles alleviate drought-induced alterations in sorghum performance, nutrient acquisition, and grain fortification. Sci. Total Environ. 2019, 688, 926–934. [Google Scholar] [CrossRef]
- Martínez-ballesta, M.C.; Chelbi, N.; López-zaplana, A.; Carvajal, M. Discerning the mechanism of the multiwalled carbon nanotubes effect on root cell water and nutrient transport. Plant Physiol. Biochem. 2020, 146, 23–30. [Google Scholar] [CrossRef]
- Ahmadian, K.; Jalilian, J.; Pirzad, A. Nano-fertilizers improved drought tolerance in wheat under deficit irrigation. Agric. Water Manag. 2021, 244, 106544. [Google Scholar] [CrossRef]
- Sarkar, M.M.; Pradhan, N.; Subba, R.; Saha, P.; Roy, S. Sugar-terminated carbon-nanodots stimulate osmolyte accumulation and ROS detoxification for the alleviation of salinity stress in Vigna radiata. Sci. Rep. 2022, 12, 17567. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yao, Y.; Yue, L.; Chen, F.; Cao, X.; Li, J.; Yang, H.; Zhang, N.; Liu, T.; Wang, Z.; et al. Regulation mechanisms of nitrogen-doped carbon dots in enhanced maize photosynthesis under drought stress. ACS Agric. Sci. Technol. 2023, 3, 181–189. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, B.; Man, H.; Chen, L.; Wang, X.; Tu, J.; Guo, Z.; Jin, G.; Lou, J.; Ci, L. Enhanced bioaccumulation efficiency and tolerance for Cd (Ⅱ) in Arabidopsis thaliana by amphoteric nitrogen-doped carbon dots. Ecotoxicol. Environ. Saf. 2020, 190, 110108. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Chen, L.; Nie, X.; Man, H.; Guo, Z.; Wang, X.; Tu, J.; Jin, G.; Ci, L. Impacts of surface chemistry of functional carbon nanodots on the plant growth. Ecotoxicol. Environ. Saf. 2020, 206, 111220. [Google Scholar] [CrossRef]
- Arshad, M.; Asghar, M.; Junaid, M.; Warsi, M.F.; Naveed Rasheed, M.; Hashim, M.; Al-Maghrabi, M.A.; Khan, M.A. Structural and magnetic properties variation of manganese ferrites via Co-Ni substitution. J. Magn. Magn. Mater. 2019, 474, 98–103. [Google Scholar] [CrossRef]
- Salih, S.J.; Mahmood, W.M. Review on magnetic spinel ferrite (MFe2O4) nanoparticles: From synthesis to application. Heliyon 2023, 9, e16601. [Google Scholar] [CrossRef] [PubMed]
- Tang, Z.X.; Sorensen, K.J.; Klabunde, K.J.; Hadjipanayis, G.C. Size-dependent Curie temperature in nanoscale MnFe2O4 particles. Phys. Rev. Lett. 1991, 67, 3602. [Google Scholar] [CrossRef]
- Aslibeiki, B.; Kameli, P.; Ehsani, M.H. MnFe2O4 bulk, nanoparticles and film: A comparative study of structural and magnetic properties. Ceram. Int. 2016, 42, 12789–12795. [Google Scholar] [CrossRef]
- Islam, K.; Haque, M.; Kumar, A.; Hoq, A.; Hyder, F.; Hoque, S.M. Manganese ferrite nanoparticles (MnFe2O4): Size dependence for hyperthermia and negative/positive contrast enhancement in MRI. Nanomaterials 2020, 10, 2297. [Google Scholar] [CrossRef] [PubMed]
- Reddy, A.V.B.; Yusop, Z.; Jaafar, J.; Reddy, Y.V.M.; Bin Aris, A.; Majid, Z.A.; Talib, J.; Madhavi, G. Recent progress on Fe-based nanoparticles: Synthesis, properties, characterization and environmental applications. J. Environ. Chem. Eng. 2016, 4, 3537–3553. [Google Scholar] [CrossRef]
- Su, Y.; Ashworth, V.; Kim, C.; Adeleye, A.S.; Rolshausen, P.; Roper, C.; White, J.; Jassby, D. Delivery, uptake, fate, and transport of engineered nanoparticles in plants: A critical review and data analysis. Environ. Sci. Nano 2019, 6, 2311–2331. [Google Scholar] [CrossRef]
- Yue, L.; Feng, Y.; Ma, C.; Wang, C.; Chen, F.; Cao, X.; Wang, J.; White, J.C.; Wang, Z.; Xing, B. Molecular mechanisms of early flowering in tomatoes induced by manganese ferrite (MnFe2O4) nanomaterials. ACS Nano 2022, 16, 5636–5646. [Google Scholar] [CrossRef]
- Avellan, A.; Yun, J.; Zhang, Y.; Spielman-Sun, E.; Unrine, J.M.; Thieme, J.; Li, J.; Lombi, E.; Bland, G.; Lowry, G.V. Nanoparticle size and coating chemistry control foliar uptake pathways, translocation, and leaf-to-rhizosphere transport in wheat. ACS Nano 2019, 13, 5291–5305. [Google Scholar] [CrossRef]
- Tombuloglu, H.; Tombuloglu, G.; Slimani, Y.; Ercan, I.; Sozeri, H.; Baykal, A. Impact of manganese ferrite (MnFe2O4) nanoparticles on growth and magnetic character of barley (Hordeum vulgare L.). Environ. Pollut. 2018, 243, 872–881. [Google Scholar] [CrossRef]
- Yang, H.; Wang, C.; Chen, F.; Yue, L.; Cao, X.; Li, J.; Zhao, X.; Wu, F.; Wang, Z.; Xing, B. Foliar carbon dot amendment modulates carbohydrate metabolism, rhizospheric properties and drought tolerance in maize seedling. Sci. Total Environ. 2022, 809, 151105. [Google Scholar] [CrossRef] [PubMed]
- Xu, X.; Wolfe, L.; Diez, J.; Zheng, Y.; Guo, H.; Hu, S. Differential germination strategies of native and introduced populations of the invasive species Plantago virginica. NeoBiota 2019, 43, 101–118. [Google Scholar] [CrossRef]
- Nimbalkar, M.S.; Pawar, N.V.; Pai, S.R.; Dixit, G.B. Synchronized variations in levels of essential amino acids during germination in grain Amaranth. Braz. J. Bot. 2020, 43, 481–491. [Google Scholar] [CrossRef]
- Kakati, J.P.; Fallen, P.; Bridges, W.; Narayanan, S. Characterization of a Soybean (Glycine max L. Merr.) population for germination and seedling root traits under Water Stress. Agronomy 2022, 12, 1944. [Google Scholar] [CrossRef]
- Sun, Y.; Guo, C.; Zhong, Y.; Xing, M.; Li, B.; Yang, Y.; Yuan, X.; Wen, Y.; Shu, C.; Yang, Z.; et al. Relationship between seed germination physiological characteristics and germination percentages of direct-seeded hybrid Indica rice under low-temperature and anaerobic interaction. Seed Sci. Technol. 2022, 50, 241–256. [Google Scholar] [CrossRef]
- Koger, C.H.; Reddy, K.N.; Poston, D.H. Factors affecting seed germination, seedling emergence, and survival of texasweed (Caperonia palustris). Weed Sci. 2004, 52, 989–995. [Google Scholar] [CrossRef]
- Itroutwar, P.D.; Kasivelu, G.; Raguraman, V.; Malaichamy, K.; Sevathapandian, S.K. Effects of biogenic zinc oxide nanoparticles on seed germination and seedling vigor of maize (Zea mays). Biocatal. Agric. Biotechnol. 2020, 29, 101778. [Google Scholar] [CrossRef]
- Itroutwar, P.D.; Govindaraju, K.; Tamilselvan, S.; Kannan, M.; Raja, K.; Subramanian, K.S. Seaweed-based biogenic ZnO nanoparticles for improving agro-morphological characteristics of rice (Oryza sativa L.). J. Plant Growth Regul. 2020, 39, 717–728. [Google Scholar] [CrossRef]
- Li, Y.; Liu, J.; Fu, C.; Khan, M.N.; Hu, J.; Zhao, F.; Wu, H.; Li, Z. CeO2 nanoparticles modulate Cu-Zn superoxide dismutase and lipoxygenase-IV isozyme activities to alleviate membrane oxidative damage to improve rapeseed salt tolerance. Environ. Sci. Nano 2022, 9, 1116–1132. [Google Scholar] [CrossRef]
- Lebedev, S.V.; Korotkova, A.M.; Osipova, E.A. Influence of Fe0 nanoparticles, magnetite Fe3O4 nanoparticles, and iron (II) sulfate (FeSO4) solutions on the content of photosynthetic pigments in Triticum vulgare. Russ. J. Plant Physiol. 2014, 61, 564–569. [Google Scholar] [CrossRef]
- Ghimire, B.K.; Seong, E.S.; Nguyen, T.X.; Yoo, J.H.; Yu, C.Y.; Kim, S.H.; Chung, I.M. Assessment of morphological and phytochemical attributes in triploid and hexaploid plants of the bioenergy crop Miscanthus × giganteus. Ind. Crops Prod. 2016, 89, 231–243. [Google Scholar] [CrossRef]
- Kim, Y.; Chung, Y.S.; Lee, E.; Tripathi, P.; Heo, S.; Kim, K.H. Root response to drought stress in rice (Oryza sativa L.). Int. J. Mol. Sci. 2020, 21, 1513. [Google Scholar] [CrossRef] [PubMed]
- Wang, C.; Yang, H.; Chen, F.; Yue, L.; Wang, Z.; Xing, B. Nitrogen-doped carbon dots increased light conversion and electron supply to improve the corn photosystem and yield. Environ. Sci. Technol. 2021, 55, 12317–12325. [Google Scholar] [CrossRef]
- Wang, H.; Zhang, M.; Song, Y.; Li, H.; Huang, H.; Shao, M.; Liu, Y.; Kang, Z. Carbon dots promote the growth and photosynthesis of mung bean sprouts. Carbon N. Y. 2018, 136, 94–102. [Google Scholar] [CrossRef]
- Su, L.X.; Ma, X.L.; Zhao, K.K.; Shen, C.L.; Lou, Q.; Yin, D.M.; Shan, C.X. Carbon nanodots for enhancing the stress resistance of peanut plants. ACS Omega 2018, 3, 17770–17777. [Google Scholar] [CrossRef]
- Cantu, J.M.; Ye, Y.; Hernandez-Viezcas, J.A.; Zuverza-Mena, N.; White, J.C.; Gardea-Torresdey, J.L. Tomato fruit nutritional quality is altered by the foliar application of various metal oxide nanomaterials. Nanomaterials 2022, 12, 2349. [Google Scholar] [CrossRef]
- Nykiel, M.; Gietler, M.; Fidler, J.; Graska, J.; Rybarczyk-Płońska, A.; Prabucka, B.; Muszyńska, E.; Bocianowski, J.; Labudda, M. Differential water deficit in leaves is a principal factor modifying barley response to drought stress. Int. J. Mol. Sci. 2022, 23, 15240. [Google Scholar] [CrossRef]
- Chen, Q.; Cao, X.; Nie, X.; Li, Y.; Liang, T.; Ci, L. Alleviation role of functional carbon nanodots for tomato growth and soil environment under drought stress. J. Hazard. Mater. 2022, 423, 127260. [Google Scholar] [CrossRef]
- Osman, H.S.; Gowayed, S.M.; Elbagory, M.; Omara, A.E.D.; Abd El-Monem, A.M.; Abd El-Razek, U.A.; Hafez, E.M. Interactive impacts of beneficial microbes and Si-Zn nanocomposite on growth and productivity of soybean subjected to water deficit under salt-affected soil conditions. Plants 2021, 10, 1396. [Google Scholar] [CrossRef]
- Abdelkader, M.; Geioushy, R.A.; Fouad, O.A.; Khaled, A.G.A.; Liudmila, P.V. Investigation the activities of photosynthetic pigments, antioxidant enzymes and inducing genotoxicity of cucumber seedling exposed to copper oxides nanoparticles stress. Sci. Hortic. 2022, 305, 111364. [Google Scholar] [CrossRef]
- Zhang, M.; Jin, Z.Q.; Zhao, J.; Zhang, G.; Wu, F. Physiological and biochemical responses to drought stress in cultivated and Tibetan wild barley. Plant Growth Regul. 2015, 75, 567–574. [Google Scholar] [CrossRef]
- Islam, M.R.; Kamal, M.M.; Hossain, M.F.; Hossain, J.; Azam, M.G.; Akhter, M.M.; Hasan, M.K.; Al-Ashkar, I.; Almutairi, K.F.; Sabagh, A.E.L.; et al. Drought tolerance in mung bean is associated with the genotypic divergence, regulation of proline, photosynthetic pigment and water relation. Phyton-Int. J. Exp. Bot. 2023, 92, 955–981. [Google Scholar] [CrossRef]
- Velasco, E.A.P.; Galindo, R.B.; Aguilar, L.A.V.; Fuentes, J.A.G.; Urbina, B.A.P.; Morales, S.A.L.; Valdés, S.S. Effects of the morphology, surface modification and application methods of ZnO-NPs on the growth and biomass of tomato plants. Molecules 2020, 25, 1282. [Google Scholar] [CrossRef]
- Mazhar, M.W.; Ishtiaq, M.; Maqbool, M.; Akram, R.; Shahid, A.; Shokralla, S.; Al-Ghobari, H.; Alataway, A.; Dewidar, A.Z.; El-Sabrout, A.M.; et al. Seed priming with iron oxide nanoparticles raises biomass production and agronomic profile of water-stressed flax plants. Agronomy 2022, 12, 982. [Google Scholar] [CrossRef]
- López-Vargas, E.R.; González-García, Y.; Pérez-Álvarez, M.; Cadenas-Pliego, G.; González-Morales, S.; Benavides-Mendoza, A.; Cabrera, R.I.; Juárez-Maldonado, A. Seed priming with carbon nanomaterials to modify the germination, growth, and antioxidant status of tomato seedlings. Agronomy 2020, 10, 639. [Google Scholar] [CrossRef]
- Chen, Q.; Cao, X.; Li, Y.; Sun, Q.; Dai, L.; Li, J.; Guo, Z.; Zhang, L.; Ci, L. Functional carbon nanodots improve soil quality and tomato tolerance in saline-alkali soils. Sci. Total Environ. 2022, 830, 154817. [Google Scholar] [CrossRef]
- Jhanji, S.; Sekhon, N.K. Chlorophyll meter (SPAD-502), an effective and reliable tool for estimating chlorophyll content-How? Natl. Acad. Sci. Lett. 2011, 34, 407–412. [Google Scholar]
- Swift, T.A.; Fagan, D.; Benito-Alifonso, D.; Hill, S.A.; Yallop, M.L.; Oliver, T.A.A.; Lawson, T.; Galan, M.C.; Whitney, H.M. Photosynthesis and crop productivity are enhanced by glucose-functionalised carbon dots. New Phytol. 2021, 229, 783–790. [Google Scholar] [CrossRef]
- Khatri, K.; Rathore, M.S. Plant nanobionics and its applications for developing plants with improved photosynthetic capacity. In Photosynthesis—From Its Evolution to Future Improvements in Photosynthetic Efficiency Using Nanomaterials; IntechOpen: London, UK, 2018; pp. 95–112. [Google Scholar]
- Ilyas, M.Z.; Lee, J.K.; Ali, M.W.; Tariq, S.; Nadeem, M. Relative assessment of biochemical constituents and antioxidant potential of fermented wheat grains using Bacillus subtilis KCTC 13241. Fermentation 2022, 8, 113. [Google Scholar] [CrossRef]
- Li, Y.; Li, W.; Yang, X.; Kang, Y.; Zhang, H.; Liu, Y.; Lei, B. Salvia miltiorrhiza-derived carbon dots as scavengers of reactive oxygen species for reducing oxidative damage of plants. ACS Appl. Nano Mater. 2021, 4, 113–120. [Google Scholar] [CrossRef]
- Zahedi, S.M.; Moharrami, F.; Sarikhani, S.; Padervand, M. Selenium and silica nanostructure-based recovery of strawberry plants subjected to drought stress. Sci. Rep. 2020, 10, 17672. [Google Scholar] [CrossRef] [PubMed]
- Rikabad, M.M.; Pourakbar, L.; Moghaddam, S.S.; Popović-Djordjević, J. Agrobiological, chemical and antioxidant properties of saffron (Crocus sativus L.) exposed to TiO2 nanoparticles and ultraviolet-B stress. Ind. Crops Prod. 2019, 137, 137–143. [Google Scholar] [CrossRef]
- Pinedo-Guerrero, Z.H.; Cadenas-Pliego, G.; Ortega-Ortiz, H.; González-Morales, S.; Benavides-Mendoza, A.; Valdés-Reyna, J.; Juárez-Maldonado, A. Form of silica improves yield, fruit quality and antioxidant defense system of tomato plants under salt stress. Agriculture 2020, 10, 367. [Google Scholar] [CrossRef]
- Páramo, L.; Feregrino-Pérez, A.A.; Vega-González, M.; Escobar-Alarcón, L.; Esquivel, K. Medicago sativa L. plant response against possible eustressors (Fe, Ag, Cu)-TiO2: Evaluation of physiological parameters, total phenol content, and flavonoid quantification. Plants 2023, 12, 659. [Google Scholar] [CrossRef]
- Abdelkader, M.A.; El-Gabry, Y.A.E.G.; Sayed, A.N.; Shahin, M.G.; Darwish, H.A.; Aboukota, M.E.; Hashem, F.A.E.; Abd-Elrahman, S.H. Evaluation of physio-biochemical criteria in maize inbred lines and their F1 hybrids grown under water-deficit conditions. Ann. Agric. Sci. 2022, 67, 220–231. [Google Scholar] [CrossRef]
- Makhafola, T.J.; Elgorashi, E.E.; McGaw, L.J.; Verschaeve, L.; Eloff, J.N. The correlation between antimutagenic activity and total phenolic content of extracts of 31 plant species with high antioxidant activity. BMC Complement. Altern. Med. 2016, 16, 490. [Google Scholar] [CrossRef]
- Migut, D.; Jańczak-pieniążek, M.; Piechowiak, T.; Buczek, J.; Balawejder, M. Physiological response of maize plants (Zea mays L.) to the use of the potassium quercetin derivative. Int. J. Mol. Sci. 2021, 22, 7384. [Google Scholar] [CrossRef]
- Ali, S.; Gill, R.A.; Ulhassan, Z.; Zhang, N.; Hussain, S.; Zhang, K.; Huang, Q.; Sagir, M.; Tahir, M.B.; Gill, M.B.; et al. Exogenously applied melatonin enhanced the tolerance of Brassica napus against cobalt toxicity by modulating antioxidant defense, osmotic adjustment, and expression of stress response genes. Ecotoxicol. Environ. Saf. 2023, 252, 114624. [Google Scholar] [CrossRef]
- Ghasemi, A.; Farzaneh, S.; Moharramnejad, S.; Sharifi, R.S.; Youesf, A.F.; Telesinski, A.; Kalaji, H.M.; Mojski, J. Impact of 24-epibrassinolide, spermine, and silicon on plant growth, antioxidant defense systems, and osmolyte accumulation of maize under water stress. Sci. Rep. 2022, 12, 14648. [Google Scholar] [CrossRef]
- Parveen, A.; Liu, W.; Hussain, S.; Asghar, J.; Perveen, S.; Xiong, Y. Silicon priming regulates morpho-physiological growth and oxidative metabolism in maize under drought stress. Plants 2019, 8, 431. [Google Scholar] [CrossRef] [PubMed]
- Lorigooini, Z.; Jamshidi-Kia, F.; Hosseini, Z. Analysis of aromatic acids (phenolic acids and hydroxycinnamic acids). In Recent Advances in Natural Products Analysis; Elsevier Inc.: Amsterdam, The Netherlands, 2020; pp. 199–219. [Google Scholar]
- Shahidi, F.; Naczk, M. Food phenolics: Sources, chemistry, effects, applications. In Trends in Food Science & Technology; Elsevier Science Ltd.: London, UK, 1996; Volume 7, p. 243. [Google Scholar]
- Kolo, Z.; Majola, A.; Phillips, K.; Ali, A.E.E.; Sharp, R.E.; Ludidi, N. Water deficit-induced changes in phenolic acid content in maize leaves is associated with altered expression of cinnamate 4-hydroxylase and p-coumaric acid 3-hydroxylase. Plants 2023, 12, 101. [Google Scholar] [CrossRef]
- Rayee, R.; Xuan, T.D.; Tran, H.D.; Fakoori, N.A.; Khanh, T.D.; Dat, T.D. Responses of flavonoids, phenolics, and antioxidant activity in rice seedlings between japonica and indica subtypes to chilling stress. Int. Lett. Nat. Sci. 2020, 77, 41–50. [Google Scholar] [CrossRef]
- Sarker, U.; Oba, S. Augmentation of leaf color parameters, pigments, vitamins, phenolic acids, flavonoids and antioxidant activity in selected Amaranthus tricolor under salinity stress. Sci. Rep. 2018, 8, 12349. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Oba, S. Drought stress enhances nutritional and bioactive compounds, phenolic acids and antioxidant capacity of Amaranthus leafy vegetable. BMC Plant Biol. 2018, 18, 258. [Google Scholar] [CrossRef]
- Stagnari, F.; Galieni, A.; D’Egidio, S.; Falcinelli, B.; Pagnani, G.; Pace, R.; Pisante, M.; Benincasa, P. Effects of sprouting and salt stress on polyphenol composition and antiradical activity of einkorn, emmer and durum wheat. Ital. J. Agron. 2017, 12, 293–301. [Google Scholar] [CrossRef]
- Peuke, A.D.; Schraml, C.; Hartung, W.; Rennenberg, H. Identification of drought-sensitive beech ecotypes by physiological parameters. New Phytol. 2002, 154, 373–387. [Google Scholar] [CrossRef]
- Schonfeld, M.A.; Johnson, R.C.; Carver, B.F.; Mornhinweg, D.W. Water relations in winter wheat as drought resistance indicators. Crop. Sci. 1988, 28, 526–531. [Google Scholar] [CrossRef]
- Ruas, K.F.; Baroni, D.F.; de Souza, G.A.R.; Bernado, W.D.P.; Paixão, J.S.; dos Santos, G.M.; Filho, J.A.M.; de Abreu, D.P.; de Sousa, E.F.; Rakocevic, M.; et al. A Carica papaya L. genotype with low leaf chlorophyll concentration copes successfully with soil water stress in the field. Sci. Hortic. 2022, 293, 110722. [Google Scholar] [CrossRef]
- Choi, J.S.; Lee, J.H.; Park, H.J.; Kim, H.G.; Young, H.S.; Mun, S.I. Screening for antioxidant activity of plants and marine algae and its active principles from Prunus davidiana. Korean J. Pharmacogn. 1993, 24, 299–303. [Google Scholar]
- Sreelatha, S.; Padma, P.R. Antioxidant activity and total phenolic content of Moringa oleifera leaves in two stages of maturity. Plant Foods Hum. Nutr. 2009, 64, 303–311. [Google Scholar] [CrossRef] [PubMed]
- Sarker, U.; Hossain, M.N.; Oba, S.; Ercisli, S.; Marc, R.A.; Golokhvast, K.S. Salinity stress ameliorates pigments, minerals, polyphenolic profiles, and antiradical capacity in Lalshak. Antioxidants 2023, 12, 173. [Google Scholar] [CrossRef] [PubMed]
- Chen, C.; Chen, H.; Zhang, Y.; Thomas, H.R.; Frank, M.H.; He, Y.; Xia, R. TBtools: An integrative toolkit developed for interactive analyses of big biological data. Mol. Plant 2020, 13, 1194–1202. [Google Scholar] [CrossRef] [PubMed]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Maize Accessions | Leaf Blade Length (cm) | Leaf Width (cm) | ||||
|---|---|---|---|---|---|---|
| MnFe2O4 | Control | CND | MnFe2O4 | Control | CND | |
| 11BS8016-7 | 42.4 ± 1.4 b | 36.4 ± 2.1 c | 48.6 ± 2.8 a | 2.3 ± 0.3 a | 2.2 ± 0.4 a | 2.8 ± 0.7 a |
| 12BS5076-8 | 36.9 ± 6.1 a | 29.5 ± 1.9 b | 32.7 ± 0.0 ab | 2.0 ± 0.3 b | 1.8 ± 0.2 b | 2.3 ± 0.0 a |
| 12S8052 | 40.7 ± 2.6 b | 36.3 ± 1.2 b | 47.8 ± 5.2 a | 2.1 ± 0.3 b | 2.4 ± 0.4 ab | 2.8 ± 0.2 a |
| 14S8025 | 40.1 ± 3.9 a | 38.1 ± 1.7 a | 36.4 ± 0.9 a | 3.0 ± 0.3 a | 3.1 ± 0.4 a | 2.3 ± 0.4 b |
| 15RS8039 | 32.9 ± 2.6 ab | 27.9 ± 3.5 b | 35.2 ± 2.9 a | 2.2 ± 0.4 a | 2.4 ± 0.3 a | 2.4 ± 0.6 a |
| 15RS8056 | 29.3 ± 2.5 b | 31.7 ± 2.1 b | 43.2 ± 3.9 a | 2.3 ± 0.3 a | 2.0 ± 0.3 a | 2.1 ± 0.3 a |
| 15RS8002 | 38.9 ± 2.7 a | 32.6 ± 3.0 ab | 31.4 ± 4.1 b | 3.0 ± 0.3 a | 2.0 ± 0.5 a | 2.4 ± 0.8 a |
| 15S8021-3 | 38.7 ± 2.8 a | 38.7 ± 1.7 a | 38.9 ± 2.1 a | 2.4 ± 0.4 a | 2.3 ± 0.5 a | 2.1 ± 0.7 a |
| 16CLP23 | 32.7 ± 0.8 b | 43.9 ± 4.2 a | 48.2 ± 2.3 a | 2.3 ± 0.6 a | 2.6 ± 0.5 a | 3.0 ± 0.2 a |
| 16CLP40 | 34.2 ± 3.1 b | 39.8 ± 2.4 ab | 44.3 ± 3.6 a | 3.4 ± 0.5 a | 2.9 ± 0.4 a | 3.3 ± 0.6 a |
| 17CS5047 | 33.4 ± 2.2 b | 44.7 ± 2.2 a | 45.7 ± 3.3 a | 2.9 ± 0.4 a | 3.0 ± 0.7 a | 2.8 ± 0.9 a |
| 16S8068-9 | 34.4 ± 2.6 b | 41.2 ± 1.8 a | 32.1 ± 1.8 b | 3.1 ± 0.4 a | 3.2 ± 0.4 a | 3.2 ± 0.4 a |
| 17CS8006 | 42.0 ± 3.3 a | 42.2 ± 3.3 a | 36.6 ± 1.9 a | 3.9 ± 0.3 a | 3.7 ± 0.4 a | 2.8 ± 0.6 b |
| 17CS8067 | 39.5 ± 2.8 a | 29.8 ± 2.2 b | 41.4 ± 3.0 a | 2.2 ± 0.4 a | 2.6 ± 0.3 a | 2.3 ± 0.4 a |
| 17YS6032 | 42.0 ± 3.1 a | 43.4 ± 2.0 a | 45.7 ± 2.8 a | 3.2 ± 0.4 a | 2.9 ± 0.4 a | 3.5 ± 0.5 a |
| 17YS8003 | 33.2 ± 2.5 ab | 35.1 ± 2.9 a | 28.5 ± 3.4 b | 2.9 ± 0.3 a | 3.1 ± 0.4 a | 2.7 ± 0.6 a |
| GP3 | 46.4 ± 1.9 a | 43.0 ± 3.2 a | 46.5 ± 3.7 a | 3.0 ± 0.5 a | 2.5 ± 0.6 a | 2.4 ± 0.6 a |
| GP5 | 49.6 ± 7.9 a | 43.8 ± 3.9 a | 54.0 ± 2.4 a | 3.4 ± 0.5 a | 2.9 ± 0.8 a | 3.0 ± 0.8 a |
| HCW1 | 38.0 ± 5.4 a | 33.4 ± 2.0 a | 35.3 ± 3.5 a | 3.0 ± 0.5 a | 2.0 ± 0.5 b | 3.0 ± 0.3 a |
| HCW2 | 40.5 ± 3.9 a | 37.6 ± 2.4 a | 44.3 ± 4.3 a | 2.7 ± 0.4 a | 2.7 ± 0.7 a | 3.2 ± 0.9 a |
| HCW3 | 35.7 ± 2.8 a | 43.9 ± 3.9 b | 45.8 ± 2.4 b | 2.5 ± 0.4 b | 2.3 ± 0.7 b | 3.4 ± 0.2 a |
| HCW4 | 42.2 ± 4.3 a | 39.5 ± 3.2 a | 46.0 ± 4.2 a | 2.9 ± 0.4 a | 2.9 ± 0.6 a | 3.3 ± 0.6 a |
| HCW5 | 35.3 ± 5.8 a | 37.8 ± 2.4 a | 38.8 ± 1.2 a | 3.9 ± 0.4 a | 3.1 ± 0.6 a | 2.9 ± 0.8 a |
| HF12 | 47.2 ± 4.8 a | 48.3 ± 2.7 a | 47.1 ± 4.4 a | 3.2 ± 0.5 a | 2.2 ± 0.2 a | 2.8 ± 0.8 a |
| HF22 | 40.3 ± 2.0 ab | 36.9 ± 2.2 b | 45.7 ± 5.5 a | 3.4 ± 0.5 a | 2.5 ± 0.4 b | 3.3 ± 0.4 ab |
| HW1 | 41.4 ± 3.3 a | 42.4 ± 1.5 a | 41.2 ± 4.4 a | 3.3 ± 0.7 a | 2.6 ± 0.7 a | 3.1 ± 0.3 a |
| HW10 | 48.7 ± 5.4 a | 43.5 ± 2.0 a | 49.8 ± 1.5 a | 2.8 ± 0.6 a | 2.6 ± 0.2 a | 3.6 ± 0.7 a |
| HW11 | 38.8 ± 4.3 ab | 43.7 ± 3.7 a | 36.1 ± 2.1 b | 2.6 ± 0.3 a | 2.9 ± 0.7 a | 3.4 ± 0.6 a |
| HW12 | 48.2 ± 2.8 a | 45.6 ± 4.0 a | 49.0 ± 4.2 a | 2.9 ± 0.3 a | 2.8 ± 0.4 a | 3.1 ± 0.4 a |
| HW15 | 48.8 ± 0.0 a | 46.5 ± 1.9 a | 39.9 ± 3.3 b | 3.2 ± 0.0 a | 2.7 ± 0.5 a | 3.0 ± 0.8 a |
| HW16 | 36.5 ± 1.9 b | 36.7 ± 2.2 b | 43.5 ± 4.3 a | 2.3 ± 0.6 a | 2.0 ± 0.4 a | 2.0 ± 0.4 a |
| HW17 | 36.5 ± 1.3 b | 44.3 ± 3.4 a | 40.2 ± 1.4 ab | 1.9 ± 0.8 a | 2.9 ± 0.9 a | 2.5 ± 0.9 a |
| HW18 | 35.9 ± 1.4 b | 44.9 ± 1.1 a | 34.5 ± 3.2 b | 2.7 ± 0.2 a | 3.3 ± 0.5 a | 3.1 ± 0.5 a |
| HW19 | 41.4 ± 2.2 a | 41.3 ± 2.2 a | 35.3 ± 3.4 b | 3.6 ± 0.5 ab | 3.1 ± 0.2 b | 3.9 ± 0.4 a |
| HW3 | 42.5 ± 2.8 a | 41.6 ± 3.7 a | 44.9 ± 3.8 a | 3.8 ± 0.7 a | 1.6 ± 0.3 b | 3.2 ± 0.4 a |
| HW4 | 42.2 ± 6.0 a | 49.3 ± 2.8 a | 46.5 ± 3.2 a | 2.2 ± 0.4 b | 2.1 ± 0.4 b | 3.1 ± 0.5 a |
| HW7 | 42.8 ± 1.3 a | 44.0 ± 3.2 a | 39.2 ± 2.4 a | 2.7 ± 0.6 a | 2.8 ± 0.4 a | 3.0 ± 0.8 a |
| HW8 | 32.5 ± 2.7 b | 28.4 ± 1.6 b | 37.8 ± 3.2 a | 2.7 ± 1.0 a | 2.1 ± 0.5 a | 3.2 ± 0.5 a |
| HW9 | 38.7 ± 2.4 a | 33.5 ± 3.4 a | 36.3 ± 3.4 a | 2.9 ± 0.4 a | 2.5 ± 0.3 a | 2.4 ± 0.4 a |
| KL103 | 43.0 ± 3.7 a | 33.5 ± 2.8 b | 36.6 ± 4.5 ab | 2.5 ± 0.6 a | 2.5 ± 0.4 a | 2.2 ± 0.4 a |
| KW7 | 36.8 ± 0.0 a | 37.1 ± 2.1 a | 34.3 ± 0.0 b | 2.4 ± 0.0 a | 1.9 ± 0.4 b | 2.7 ± 0.0 a |
| Maize Accessions | Shoot Length (cm) | Root Length (cm) | ||||
|---|---|---|---|---|---|---|
| MnFe2O4 | Control | CND | MnFe2O4 | Control | CND | |
| 11BS8016-7 | 52.9 ± 0.2 b | 52.4 ± 0.7 b | 57.0 ± 2.3 a | 18.4 ± 1.5 ab | 15.8 ± 1.6 b | 21.9 ± 4.2 a |
| 12BS5076-8 | 48.2 ± 14.1 a | 32.0 ± 2.5 b | 34.1 ± 0.0 b | 23.7 ± 1.1 a | 8.6 ± 0.8 c | 16.2 ± 0.0 b |
| 12S8052 | 54.5 ± 0.9 b | 58.2 ± 1.6 b | 65.1 ± 4.8 a | 20.8 ± 3.3 b | 22.4 ± 0.4 ab | 28.1 ± 6.9 a |
| 14S8025 | 52.7 ± 3.0 a | 45.1 ± 2.9 b | 52.3 ± 2.7 a | 18.4 ± 2.3 a | 20.0 ± 4.1 a | 20.6 ± 2.0 a |
| 15RS8039 | 50.5 ± 3.7 b | 41.7 ± 1.7 c | 55.8 ± 1.0 a | 30.9 ± 6.4 a | 28.0 ± 4.6 a | 24.0 ± 3.9 a |
| 15RS8056 | 40.5 ± 1.1 b | 39.8 ± 2.7 b | 53.7 ± 1.6 a | 17.6 ± 2.4 b | 22.4 ± 1.2 b | 36.9 ± 9.4 a |
| 15RS8002 | 48.0 ± 9.1 a | 43.3 ± 1.3 a | 54.6 ± 4.4 a | 24.7 ± 1.7 a | 25.1 ± 3.3 a | 24.5 ± 3.2 a |
| 15S8021-3 | 54.0 ± 3.6 a | 42.6 ± 2.6 b | 55.3 ± 0.3 a | 33.4 ± 4.9 a | 22.2 ± 3.5 b | 18.9 ± 1.6 b |
| 16CLP23 | 51.2 ± 1.6 a | 59.0 ± 0.3 a | 56.2 ± 7.3 a | 32.2 ± 3.8 ab | 30.6 ± 2.3 b | 37.8 ± 2.9 a |
| 16CLP40 | 55.6 ± 5.5 a | 55.2 ± 3.4 a | 58.9 ± 2.2 a | 37.0 ± 3.0 a | 30.6 ± 1.9 b | 37.8 ± 3.1 a |
| 17CS5047 | 52.1 ± 2.1 a | 53.8 ± 2.6 a | 48.2 ± 10.8 a | 17.6 ± 3.1 a | 21.1 ± 3.4 a | 23.6 ± 4.6 a |
| 16S8068-9 | 54.1 ± 2.1 a | 46.9 ± 5.9 a | 48.4 ± 5.4 a | 29.2 ± 1.7 b | 36.9 ± 2.6 a | 40.9 ± 3.7 a |
| 17CS8006 | 44.0 ± 1.2 a | 55.0 ± 1.8 a | 55.7 ± 10.4 a | 30.1 ± 11.0 a | 22.9 ± 7.8 a | 24.7 ± 6.1 a |
| 17CS8067 | 55.1 ± 1.6 a | 44.6 ± 3.3 b | 57.6 ± 2.2 a | 24.9 ± 3.9 b | 22.4 ± 1.0 b | 32.5 ± 2.8 a |
| 17YS6032 | 56.7 ± 2.7 a | 52.0 ± 2.9 a | 56.7 ± 6.2 a | 24.3 ± 2.1 ab | 30.4 ± 0.9 a | 20.7 ± 5.3 b |
| 17YS8003 | 41.4 ± 3.4 a | 47.7 ± 10.5 a | 42.9 ± 6.6 a | 22.1 ± 2.5 a | 14.5 ± 2.9 a | 19.6 ± 5.1 a |
| GP3 | 61.8 ± 1.7 a | 55.8 ± 3.6 a | 58.3 ± 5.5 a | 16.8 ± 1.0 b | 27.4 ± 3.9 a | 26.5 ± 3.9 a |
| GP5 | 60.8 ± 1.8 a | 61.5 ± 2.2 a | 65.1 ± 2.4 a | 21.1 ± 1.9 a | 20.3 ± 3.4 a | 21.3 ± 5.8 a |
| HCW1 | 55.8 ± 5.6 a | 47.7 ± 5.6 a | 51.7 ± 3.3 a | 34.1 ± 0.3 a | 25.3 ± 6.3 a | 24.4 ± 6.8 a |
| HCW2 | 53.7 ± 1.2 a | 50.4 ± 2.1 a | 51.9 ± 6.6 a | 32.6 ± 1.8 ab | 26.0 ± 8.7 b | 45.2 ± 7.8 a |
| HCW3 | 54.4 ± 4.1 a | 55.8 ± 2.9 a | 62.2 ± 5.8 a | 24.8 ± 6.1 a | 24.3 ± 5.3 a | 30.1 ± 2.2 a |
| HCW4 | 52.8 ± 0.8 a | 55.4 ± 3.5 a | 55.5 ± 2.6 a | 33.4 ± 9.8 a | 23.4 ± 4.3 a | 34.9 ± 4.8 a |
| HCW5 | 46.0 ± 0.7 a | 54.6 ± 4.0 b | 55.6 ± 1.9 b | 25.9 ± 2.4 a | 24.1 ± 6.1 a | 20.4 ± 1.0 a |
| HF12 | 71.5 ± 0.6 a | 61.5 ± 2.7 c | 67.5 ± 2.0 b | 23.0 ± 3.2 a | 17.1 ± 1.1 b | 23.1 ± 3.2 a |
| HF22 | 55.6 ± 1.8 a | 55.2 ± 2.7 a | 57.9 ± 4.8 a | 23.7 ± 5.8 a | 25.8 ± 3.7 a | 20.7 ± 7.7 a |
| HW1 | 54.3 ± 0.6 ab | 52.5 ± 2.1 b | 58.9 ± 4.2 a | 31.3 ± 15.8 a | 20.2 ± 2.2 a | 15.1 ± 3.7 a |
| HW10 | 59.5 ± 0.9 b | 56.2 ± 1.0 c | 63.9 ± 2.4 a | 21.7 ± 4.3 a | 27.2 ± 7.9 a | 29.8 ± 6.5 a |
| HW11 | 59.6 ± 2.1 a | 60.4 ± 2.7 a | 56.6 ± 3.8 a | 26.8 ± 4.2 a | 23.7 ± 5.6 a | 22.9 ± 6.4 a |
| HW12 | 55.8 ± 2.4 a | 50.8 ± 14.6 a | 58.4 ± 2.7 a | 34.7 ± 2.6 a | 23.2 ± 5.8 b | 21.4 ± 3.5 b |
| HW15 | 57.4 ± 0.0 a | 51.8 ± 6.0 a | 57.7 ± 2.9 a | 31.6 ± 0.0 a | 20.3 ± 4.3 b | 15.5 ± 2.2 b |
| HW16 | 41.9 ± 1.6 ab | 40.9 ± 3.4 b | 48.5 ± 4.5 a | 25.3 ± 1.6 a | 30.9 ± 7.7 a | 25.6 ± 3.5 a |
| HW17 | 53.9 ± 5.7 a | 54.5 ± 6.1 a | 61.4 ± 1.9 a | 25.1 ± 4.7 a | 17.6 ± 1.7 a | 22.7 ± 4.2 a |
| HW18 | 50.3 ± 10.8 a | 56.4 ± 3.1 a | 60.6 ± 1.6 a | 22.1 ± 3.3 a | 24.1 ± 2.2 a | 18.2 ± 3.1 a |
| HW19 | 43.8 ± 6.7 a | 50.7 ± 6.2 a | 51.4 ± 1.0 a | 19.7 ± 2.9 a | 26.8 ± 2.6 a | 21.9 ± 5.6 a |
| HW3 | 66.8 ± 2.9 a | 52.6 ± 4.0 b | 56.7 ± 5.3 b | 22.9 ± 2.0 a | 17.9 ± 2.7 a | 21.8 ± 5.4 a |
| HW4 | 53.1 ± 1.3 ab | 50.4 ± 6.5 b | 59.9 ± 3.4 a | 25.8 ± 8.7 a | 20.4 ± 3.2 a | 23.2 ± 6.2 a |
| HW7 | 52.1 ± 1.8 a | 45.8 ± 5.2 a | 47.8 ± 2.9 a | 23.0 ± 2.6 a | 27.4 ± 8.2 a | 26.8 ± 9.3 a |
| HW8 | 44.2 ± 3.1 a | 50.4 ± 3.9 a | 48.8 ± 3.4 a | 21.4 ± 6.3 a | 15.9 ± 1.3 a | 14.3 ± 3.0 a |
| HW9 | 55.8 ± 2.6 a | 52.4 ± 3.9 a | 51.7 ± 9.4 a | 28.2 ± 4.6 ab | 18.5 ± 3.2 b | 37.6 ± 7.9 a |
| KL103 | 51.2 ± 2.8 a | 40.1 ± 1.4 a | 45.1 ± 10.8 a | 14.7 ± 1.9 b | 18.2 ± 2.9 ab | 28.3 ± 9.9 a |
| KW7 | 53.4 ± 0.0 a | 54.5 ± 4.1 a | 47.6 ± 0.0 b | 6.5 ± 0.0 b | 19.5 ± 1.8 b | 22.0 ± 0.0 a |
| Maize Accessions | Root Fresh Weight (g) | Shoot Fresh Weight (g) | ||||
|---|---|---|---|---|---|---|
| MnFe2O4 | Control | CND | MnFe2O4 | Control | CND | |
| 11BS8016-7 | 2.0 ± 0.5 b | 3.0 ± 0.5 a | 3.8 ± 0.3 a | 7.8 ± 0.5 a | 8.5 ± 0.8 a | 8.9 ± 0.6 a |
| 12BS5076-8 | 1.5 ± 0.4 b | 1.6 ± 0.5 b | 6.1 ± 0.0 a | 6.6 ± 1.3 a | 4.7 ± 0.5 b | 7.1 ± 0.0 a |
| 12S8052 | 2.9 ± 0.6 ab | 2.3 ± 0.3 b | 3.4 ± 0.2 a | 6.2 ± 0.3 b | 8.6 ± 0.5 a | 8.7 ± 0.8 a |
| 14S8025 | 4.2 ± 0.4 a | 4.5 ± 0.4 a | 3.1 ± 0.3 b | 9.9 ± 0.4 a | 9.1 ± 0.4 ab | 8.3 ± 0.7 b |
| 15RS8039 | 4.3 ± 0.5 a | 4.7 ± 0.6 a | 4.0 ± 0.4 a | 10.3 ± 1.8 a | 7.9 ± 0.3 b | 7.0 ± 0.5 b |
| 15RS8056 | 2.2 ± 0.6 b | 1.1 ± 0.2 c | 4.3 ± 0.4 a | 5.4 ± 0.7 c | 7.1 ± 0.2 b | 8.6 ± 0.4 a |
| 15RS8002 | 3.8 ± 0.9 b | 4.3 ± 0.4 b | 6.0 ± 0.3 a | 11.2 ± 1.4 a | 10.0 ± 0.3 a | 11.3 ± 0.5 a |
| 15S8021-3 | 3.2 ± 0.3 a | 2.1 ± 0.4 b | 2.2 ± 0.3 b | 9.3 ± 0.8 a | 7.6 ± 0.3 b | 9.3 ± 0.6 a |
| 16CLP23 | 4.9 ± 0.6 a | 5.5 ± 0.9 a | 3.9 ± 0.8 a | 10.6 ± 1.1 a | 7.9 ± 0.4 b | 8.6 ± 1.3 b |
| 16CLP40 | 3.4 ± 0.4 b | 3.1 ± 0.4 b | 5.1 ± 0.6 a | 9.5 ± 0.9 a | 7.5 ± 0.3 b | 8.8 ± 0.9 ab |
| 17CS5047 | 3.1 ± 0.5 a | 3.8 ± 0.8 a | 2.8 ± 0.6 a | 6.1 ± 0.7 b | 7.4 ± 0.8 a | 5.9 ± 0.1 b |
| 16S8068-9 | 3.3 ± 0.5 b | 2.2 ± 0.4 c | 4.7 ± 0.6 a | 11.4 ± 1.9 a | 10.6 ± 0.5 a | 9.2 ± 1.0 a |
| 17CS8006 | 2.4 ± 0.3 b | 2.8 ± 0.3 b | 4.1 ± 0.9 a | 9.2 ± 0.3 ab | 10.0 ± 0.9 a | 8.3 ± 0.9 b |
| 17CS8067 | 3.0 ± 1.0 a | 2.4 ± 0.7 a | 3.4 ± 1.2 a | 8.0 ± 0.5 b | 12.1 ± 0.3 a | 7.5 ± 0.8 b |
| 17YS6032 | 5.8 ± 0.5 a | 5.7 ± 0.5 a | 5.8 ± 0.4 a | 12.2 ± 0.9 a | 11.1 ± 0.5 ab | 10.1 ± 0.6 b |
| 17YS8003 | 3.6 ± 0.4 b | 2.8 ± 0.3 b | 6.8 ± 0.6 a | 8.9 ± 1.2 a | 5.9 ± 0.2 b | 7.5 ± 1.0 ab |
| GP3 | 4.3 ± 0.3 b | 3.8 ± 0.4 b | 5.8 ± 0.7 a | 10.5 ± 0.7 a | 10.7 ± 0.5 a | 10.1 ± 0.6 a |
| GP5 | 4.1 ± 0.4 b | 5.0 ± 0.5 ab | 5.9 ± 0.8 a | 9.4 ± 1.0 a | 7.1 ± 0.8 b | 9.1 ± 0.2 a |
| HCW1 | 3.6 ± 0.3 b | 3.8 ± 0.6 b | 5.1 ± 0.6 a | 9.8 ± 1.3 a | 8.3 ± 0.1 a | 9.0 ± 0.8 a |
| HCW2 | 2.8 ± 0.7 a | 3.8 ± 0.7 a | 4.0 ± 0.8 a | 10.3 ± 0.7 a | 9.3 ± 0.7 ab | 8.4 ± 0.3 b |
| HCW3 | 3.2 ± 0.3 c | 5.3 ± 0.4 b | 7.0 ± 0.4 a | 10.0 ± 1.0 a | 9.2 ± 0.6 a | 9.2 ± 0.6 a |
| HCW4 | 3.9 ± 0.2 b | 3.6 ± 0.3 b | 6.0 ± 0.9 a | 10.5 ± 1.2 a | 9.1 ± 0.4 ab | 8.2 ± 1.0 b |
| HCW5 | 4.1 ± 0.5 b | 5.3 ± 0.8 a | 2.6 ± 0.4 c | 6.0 ± 0.3 b | 8.5 ± 0.5 a | 8.4 ± 0.3 a |
| HF12 | 3.1 ± 0.7 a | 3.8 ± 0.7 a | 3.7 ± 0.7 a | 8.6 ± 0.7 c | 11.6 ± 0.3 a | 10.0 ± 0.7 b |
| HF22 | 3.2 ± 0.2 c | 4.3 ± 0.4 b | 5.4 ± 0.3 a | 9.9 ± 0.6 a | 9.9 ± 0.6 a | 9.0 ± 0.2 a |
| HW1 | 4.7 ± 0.5 a | 2.6 ± 0.3 b | 2.3 ± 0.6 b | 11.4 ± 1.0 a | 8.6 ± 0.2 b | 8.9 ± 0.8 b |
| HW10 | 4.1 ± 0.5 b | 6.1 ± 0.4 a | 2.4 ± 0.6 c | 7.1 ± 0.7 b | 7.2 ± 0.4 b | 10.6 ± 0.4 a |
| HW11 | 5.2 ± 0.3 a | 5.0 ± 0.5 a | 4.6 ± 0.4 a | 7.2 ± 1.1 b | 11.7 ± 0.4 a | 7.1 ± 0.2 b |
| HW12 | 4.2 ± 0.4 a | 4.3 ± 0.7 a | 5.1 ± 0.3 a | 11.9 ± 0.4 a | 12.3 ± 0.6 a | 8.2 ± 0.6 b |
| HW15 | 4.1 ± 0.0 ab | 3.4 ± 0.6 b | 4.4 ± 0.5 a | 12.5 ± 0.0 a | 7.9 ± 0.3 b | 8.1 ± 0.5 b |
| HW16 | 3.7 ± 0.2 a | 5.3 ± 0.3 a | 4.0 ± 1.4 a | 7.5 ± 0.5 b | 7.6 ± 0.5 b | 9.0 ± 0.2 a |
| HW17 | 4.5 ± 0.4 a | 4.3 ± 0.5 a | 5.0 ± 0.7 a | 9.2 ± 0.6 b | 10.1 ± 0.3 a | 8.4 ± 0.2 b |
| HW18 | 3.8 ± 0.4 ab | 3.9 ± 0.5 a | 2.8 ± 0.6 b | 6.3 ± 0.6 b | 9.8 ± 1.0 a | 10.4 ± 0.5 a |
| HW19 | 3.0 ± 0.3 b | 5.8 ± 0.5 a | 3.4 ± 0.9 b | 8.3 ± 1.7 a | 8.1 ± 0.4 a | 6.8 ± 0.5 a |
| HW3 | 4.4 ± 0.8 a | 5.3 ± 0.6 a | 4.8 ± 0.9 a | 9.2 ± 0.6 a | 8.8 ± 0.4 a | 9.0 ± 0.6 a |
| HW4 | 4.2 ± 0.4 b | 6.0 ± 0.3 a | 4.9 ± 0.4 b | 9.4 ± 0.9 a | 7.7 ± 0.5 b | 9.3 ± 0.3 a |
| HW7 | 5.0 ± 0.5 a | 4.0 ± 0.9 a | 5.3 ± 0.6 a | 10.9 ± 0.7 a | 6.8 ± 0.2 b | 7.0 ± 0.2 b |
| HW8 | 4.7 ± 0.3 a | 2.3 ± 0.5 b | 5.2 ± 0.4 a | 8.0 ± 0.2 a | 7.0 ± 0.3 b | 8.4 ± 0.2 a |
| HW9 | 4.6 ± 0.4 a | 2.3 ± 0.3 b | 5.1 ± 0.8 a | 10.5 ± 1.3 a | 8.5 ± 0.7 b | 8.0 ± 0.5 b |
| KL103 | 3.1 ± 0.3 a | 2.4 ± 0.3 a | 2.2 ± 0.7 a | 7.0 ± 0.7 a | 4.1 ± 0.3 b | 4.1 ± 0.2 b |
| KW7 | 2.2 ± 0.0 b | 3.6 ± 0.3 a | 2.4 ± 0.0 b | 7.9 ± 0.0 a | 5.0 ± 0.6 b | 8.3 ± 0.0 a |
| Characteristics | CND | MnFe2O4 NP |
|---|---|---|
| Product name | Carbon nanodots-deep UV fluorescent | Manganese ferrite nanoparticles |
| Purity | 98.5% | 98.95% |
| Average particle size | 1.6–1.8 nm | 55 nm |
| Shape of particle | - | Spherical |
| Physical state | Clear liquid | Powder |
| Molecular weight | 12.011 g/mol | - |
| Concentration | >250 mg/mL | - |
| UV-Vis | <190 and 270 nm | - |
| Emission peak | λem. = 302 and 420 nm by λexc. at 179 nm | - |
| Photoluminescence quantum yield | 11.3% | - |
| pH value | 6.7–7 | - |
| Seedbed Characteristics | Proportions |
|---|---|
| Bulk density | 0.15–0.25 Mg m−3 |
| pH (1:5, v/v) | 5.5–7.0 |
| Electrical conductivity | 0.65 ± dS m−1 |
| NO3—N | 200–350 mg L−1 |
| NH4+-N | below 150 mg L−1 |
| Cation exchange capacity | 35–55 cmol+ L−1 |
| Available phosphorus (P2O5) | 200–350 mg L−1 |
| Raw material and mixing ratio (%) | Zeolite 4, perlite 7, vermiculite 6, coco peat 68, peat moss 14.73, fertilizers 0.201, wetting agent 0.064, pH adjusting agent 0.005 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ilyas, M.Z.; Park, H.; Baek, Y.S.; Sa, K.J.; Kim, M.J.; Lee, J.K. Efficacy of Carbon Nanodots and Manganese Ferrite (MnFe2O4) Nanoparticles in Stimulating Growth and Antioxidant Activity in Drought-Stressed Maize Inbred Lines. Plants 2023, 12, 2922. https://doi.org/10.3390/plants12162922
Ilyas MZ, Park H, Baek YS, Sa KJ, Kim MJ, Lee JK. Efficacy of Carbon Nanodots and Manganese Ferrite (MnFe2O4) Nanoparticles in Stimulating Growth and Antioxidant Activity in Drought-Stressed Maize Inbred Lines. Plants. 2023; 12(16):2922. https://doi.org/10.3390/plants12162922
Chicago/Turabian StyleIlyas, Muhammad Zahaib, Hyeon Park, Young Sun Baek, Kyu Jin Sa, Myong Jo Kim, and Ju Kyong Lee. 2023. "Efficacy of Carbon Nanodots and Manganese Ferrite (MnFe2O4) Nanoparticles in Stimulating Growth and Antioxidant Activity in Drought-Stressed Maize Inbred Lines" Plants 12, no. 16: 2922. https://doi.org/10.3390/plants12162922
APA StyleIlyas, M. Z., Park, H., Baek, Y. S., Sa, K. J., Kim, M. J., & Lee, J. K. (2023). Efficacy of Carbon Nanodots and Manganese Ferrite (MnFe2O4) Nanoparticles in Stimulating Growth and Antioxidant Activity in Drought-Stressed Maize Inbred Lines. Plants, 12(16), 2922. https://doi.org/10.3390/plants12162922