
Antimicrobial Activity of Diffusible and Volatile Metabolites Emitted by Beauveria bassiana: Chemical Profile of Volatile Organic Compounds (VOCs) Using SPME-GC/MS Analysis




Abstract
1. Introduction
2. Results
2.1. Molecular Identification of the Studied Isolates of Beauveria
2.2. Antagonistic Activity of B. bassiana Isolates
2.3. Antimicrobial Activity of Exo- and Endo-Diffusible Metabolites
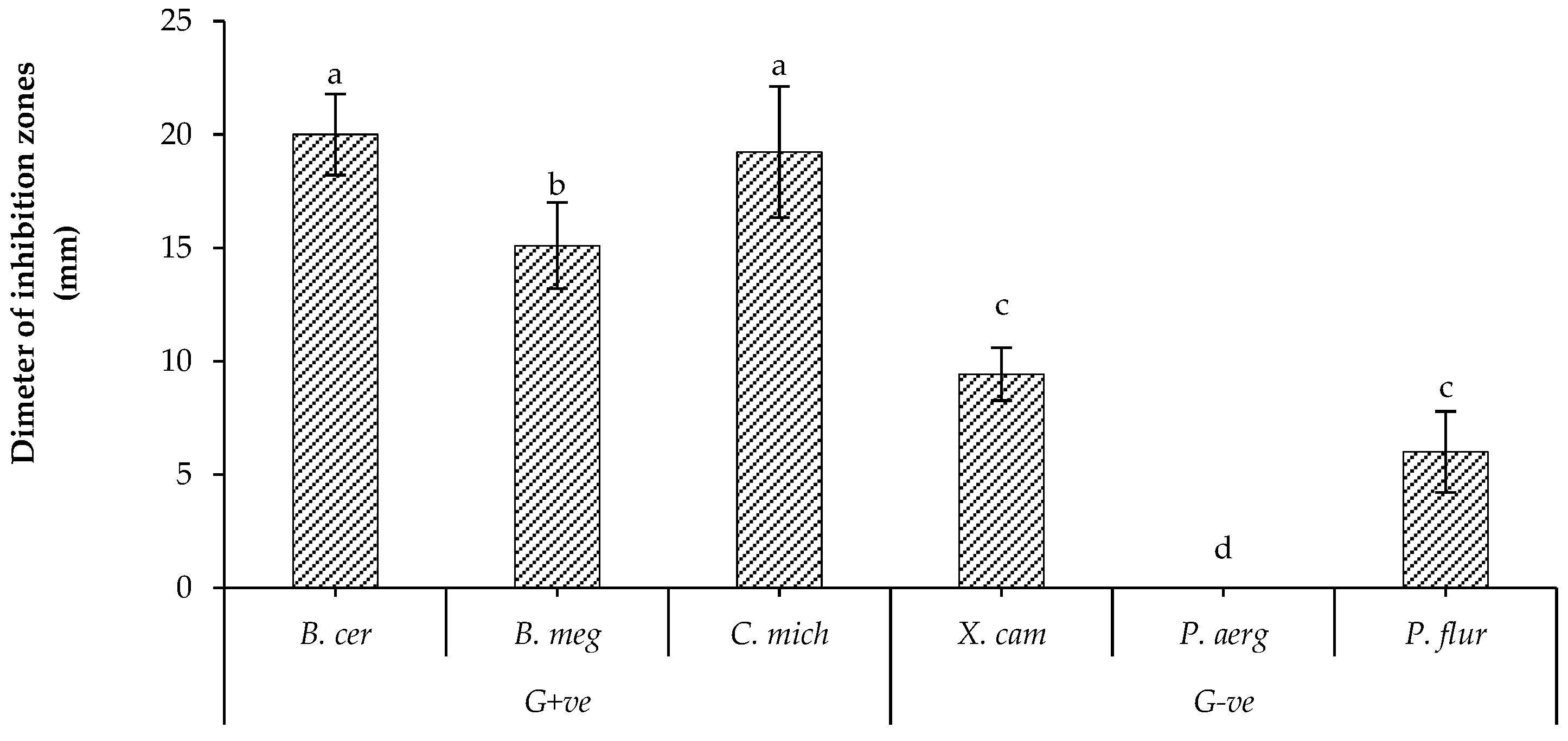
2.4. Antibacterial Activity of Volatiles Metabolites
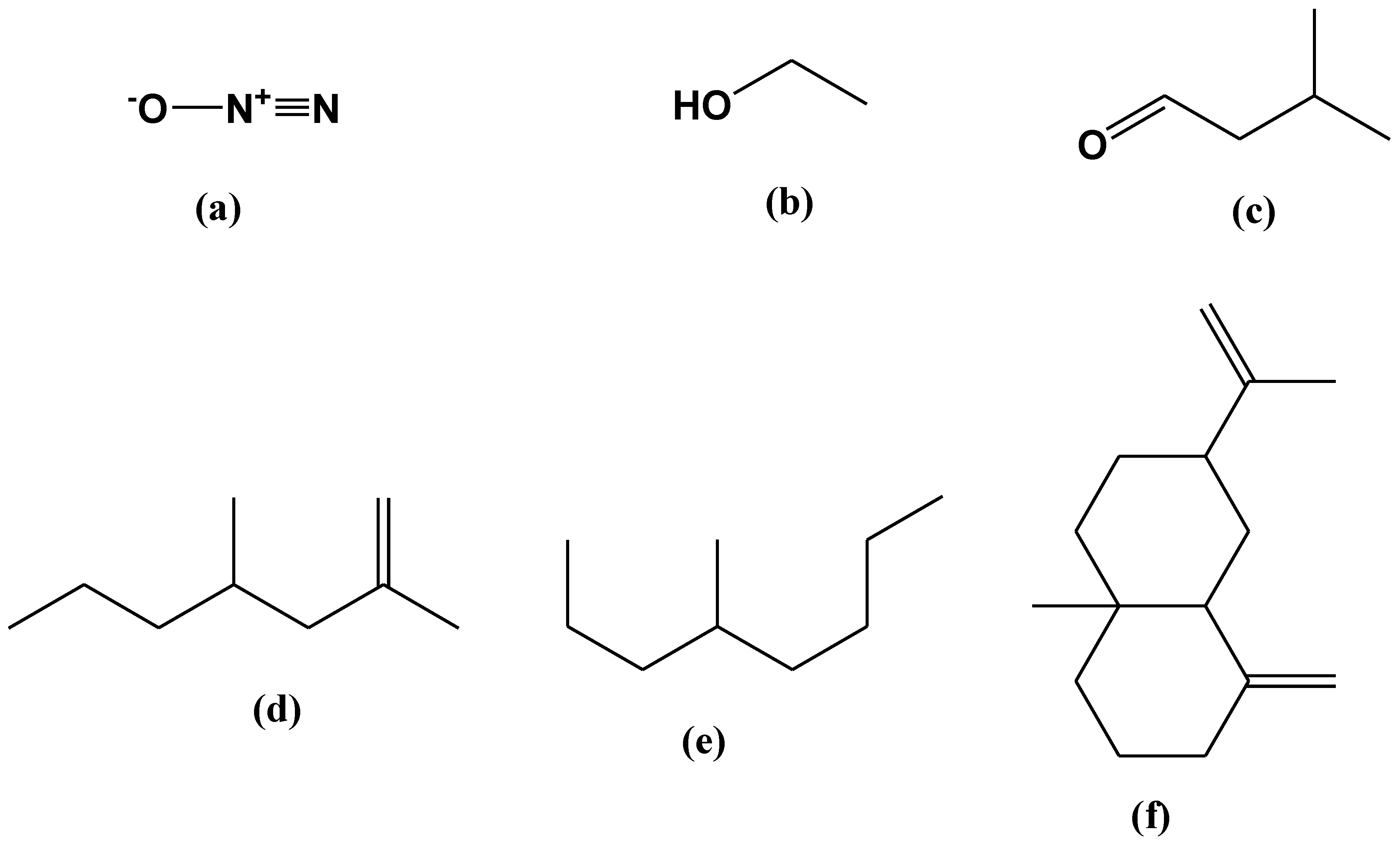
2.5. SPME-GC/MS Analysis of VOCs
3. Discussion
4. Materials and Methods
4.1. Isolation, Culturing and Identification
4.2. Antagonistic Activity
4.3. Extraction of Secondary Metabolites
4.4. Antibacterial Activity of Diffusible Metabolites
4.5. Antibacterial Activity of Volatiles Metabolites
4.6. SPME-GC/MS of VOCs
4.7. Statistical Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Vega, F.E.; Goettel, M.S.; Blackwell, M.; Chandler, D.; Jackson, M.A.; Keller, S.; Koike, M.; Maniania, N.K.; Monz´on, A.; Ownley, B.H.; et al. Fungal entomopathogens: New insights on their ecology. Fungal Ecol. 2009, 2, 149–159. [Google Scholar] [CrossRef]
- Barbarin, A.M.; Jenkins, N.E.; Rajotte, E.G.; Thomas, M.B. A preliminary evaluation of the potential of Beauveria bassiana for bed bug control. J. Invertebrate Pathol. 2012, 111, 82–85. [Google Scholar] [CrossRef] [PubMed]
- Wraight, S.P.; Lopes, R.B.; Faria, M. Chapter 16: Microbial Control of Mite and Insect Pests of Greenhouse Crops. In Microbial Control of Insect and Mite Pests; Academic Press: Yakima, WA, USA, 2017; pp. 237–252. [Google Scholar]
- Yuan, W.; Wang, P.; Zhang, Z.; Li, S. Glycosylation of (–)-maackiain by Beauveria bassiana and Cunninghamella echinulata var. elegans. Biocatal. Biotransform. 2010, 28, 117–121. [Google Scholar] [CrossRef]
- Berestetskiya, A.O.; Ivanovaa, A.N.; Petrovaa, M.O.; Prokof’evab, D.S.; Stepanychevaa, E.A.; Uspanovc, A.M.; Lednev, G.R. Comparative analysis of the biological activity and chromatographic profiles of the extracts of Beauveria bassiana and B. pseudobassiana cultures grown on different nutrient substrates. Microbiology 2018, 87, 200–214. [Google Scholar] [CrossRef]
- Fan, Y.; Zhang, Y.; Yang, X.; Pei, X.; Guo, S.; Pei, Y. Expression of a Beauveria bassiana chitinase (Bbchit1) in Escherichia coli and Pichia pastoris. Protein Expr. Purif. 2007, 56, 93–99. [Google Scholar] [CrossRef]
- Vega, F.E. The use of fungal entomopathogens as endophytes in biological control: A review. Mycologia 2018, 110, 4–30. [Google Scholar] [CrossRef]
- Fabrice, D.H.; Elie, D.A.; Kobi, D.O.; Valerien, Z.A.; Thomas, H.A.; Joëlle, T.; Maurille, E.I.A.T.; Dénis, O.B.; Manuele, T. Toward the efficient use of Beauveria bassiana in integrated cotton insect pest management. J. Cotton Res. 2020, 3, 24. [Google Scholar]
- Wang, Y.; Tang, D.; Duan, D.; Wang, Y.; Yu, H. Morphology, molecular characterization, and virulence of Beauveria pseudobassiana isolated from different hosts. J. Inverteb. Pathol. 2020, 72, 107333. [Google Scholar] [CrossRef]
- Sinno, M.; Ranesi, M.; Di Lelio, I.; Iacomino, G.; Becchimanzi, A.; Barra, E.; Molisso, D.; Pennacchio, F.; Digilio, M.C.; Vitale, S.; et al. Selection of endophytic Beauveria bassiana as a dual biocontrol agent of tomato pathogens and pests. Pathogens 2021, 10, 1242. [Google Scholar] [CrossRef]
- Faria, M.R.; Wraight, S.P. Biological control of Bemisia tabaci with fungi. Crop Prot. 2001, 20, 767–778. [Google Scholar] [CrossRef]
- Faria, M.R.; Wraight, S.P. Mycoinsecticides and mycoacaricides: A comprehensive list with worldwide coverage and international classification of formulation types. Biol. Control 2007, 43, 237–256. [Google Scholar] [CrossRef]
- Fargues, J.; Remaudiere, G. Consideration on the specificity of entomopathogenic fungi. Mycopathologia 1977, 62, 31–37. [Google Scholar] [CrossRef]
- Hasaballah, A.I.; Fouda, M.A.; Hassan, M.I.; Omar, G.M. Pathogenicity of Beauveria bassiana and Metarhizium anisopliae on the adult housefly, Musca domestica L. Egypt. Acad. J. Biolog. Sci. A Entomol. 2017, 10, 79–86. [Google Scholar]
- Acharya, N.; Rajotte, E.G.; Jenkins, N.E.; Thomas, M.B. Potential for biocontrol of house flies, Musca domestica, using fungal biopesticides. Biocontrol Sci. Technol. 2015, 25, 513–524. [Google Scholar] [CrossRef]
- Barra-Bucarei, L.; Iglesias, A.F.; González, M.G.; Aguayo, G.S.; Carrasco-Fernández, J.; Castro, J.F.; Campos, J.O. Antifungal activity of Beauveria bassiana Endophyte against Botrytis cinerea in Two Solanaceae Crops. Microorganisms 2020, 8, 65. [Google Scholar] [CrossRef] [PubMed]
- Parine, N.R.; Pathan, A.K.; Sarayu, B.; Nishanth, V.S.; Bobbarala, V. Antibacterial efficacy of secondary metabolites from entomopathogenic fungi Beauveria bassiana. Int. J. Chem. Anal. Sci. 2010, 1, 94–96. [Google Scholar]
- Ownley, B.H.; Bishop, D.G.; Pereira, R.M. Biocontrol of Rhizoctonia damping off of tomato with Beauveria bassiana. Phytopathology 2000, 90, S58. [Google Scholar]
- Tomilova, O.G.; Shaldyaeva, E.M.; Kryukova, N.A.; Pilipova, Y.V.; Schmidt, N.S.; Danilov, V.P.; Kryukov, V.Y.; Glupov, V.V. Entomopathogenic fungi decrease Rhizoctonia disease in potato in field conditions. PeerJ 2020, 8, e9895. [Google Scholar] [CrossRef]
- Della Pepa, T.; Elshafie, H.S.; Capasso, R.; De Feo, V.; Camele, I.; Nazzaro, F.; Scognamiglio, M.R.; Caputo, L. Antimicrobial and phytotoxic activity of Origanum heracleoticum and O. majorana essential oils growing in Cilento (Southern Italy). Molecules 2019, 24, 2576. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Mancini, E.; Camele, I.; Martino, L.D.; De Feo, V. In vivo antifungal activity of two essential oils from Mediterranean plants against postharvest brown rot disease of peach fruit. Ind. Crops Prod. 2015, 66, 11–15. [Google Scholar] [CrossRef]
- Anyanwu, M.U.; Okoye, R.C. Antimicrobial Activity of Nigerian Medicinal Plants. J. Intercult. Ethnopharmacol. 2017, 6, 240–259. [Google Scholar] [CrossRef]
- Keifer, M.C.; Firestone, J. Neurotoxicity of pesticides. J. Agromed. 2007, 12, 17–25. [Google Scholar] [CrossRef]
- Camele, I.; Elshafie, H.S.; Caputo, L.; Sakr, S.H.; De Feo, V. Bacillus mojavensis: Biofilm formation and biochemical investigation of its bioactive metabolites. J. Biol. Res. 2019, 92, 39–45. [Google Scholar] [CrossRef]
- Wang, H.; Peng, H.; Li, W.; Cheng, P.; Gong, M. The toxins of Beauveria bassiana and the strategies to improve their virulence to insects. Front. Microbiol. 2021, 12, 705343. [Google Scholar] [CrossRef] [PubMed]
- Fan, Y.; Liua, X.; Keyhanib, N.O.; Tanga, G.; Peia, Y.; Zhanga, W.; Tonga, S. Regulatory cascade and biological activity of Beauveria bassiana oosporein that limits bacterial growth after host death. Proc. Natl. Acad. Sci. USA 2017, 114, 1578–1586. [Google Scholar] [CrossRef] [PubMed]
- Altschul, S.F.; Madden, T.L.; Schäffer, A.A.; Zhang, J.; Zhang, Z.; Miller, W.; Lipman, D.J. Gapped BLAST and PSI-BLAST: A new generation of protein database search programs. Nucleic Acids Res. 1997, 25, 3389–3402. [Google Scholar] [CrossRef]
- McLafferty, F.W. Wiley Registry of Mass Spectral Data, with NIST Spectral Data CD Rom, 7th ed.; John Wiley & Sons: New York, NY, USA, 1998. [Google Scholar]
- Yang, X.; Ye, W.; Qi, Y.; Ying, Y.; Xia, Z. Overcoming Multidrug Resistance in Bacteria Through Antibiotics Delivery in Surface-Engineered Nano-Cargos: Recent Developments for Future Nano-Antibiotics. Front. Bioeng. Biotechnol. 2021, 9, 696514. [Google Scholar] [CrossRef]
- Soothill, G.; Hu, Y.; Coates, A. Can we prevent antimicrobial resistance by using antimicrobials better? Pathogens 2013, 2, 422–435. [Google Scholar] [CrossRef]
- Zimmermann, G. Review on safety of the entomopathogenic fungi Beauveria bassiana and Beauveria brongniartii. Biocontrol Sci. Technol. 2007, 17, 553–596. [Google Scholar] [CrossRef]
- Chen, W.H.; Liu, M.; Huang, Z.X.; Yang, G.M.; Han, Y.F.; Liang, J.D.; Liang, Z.Q. Beauveria majiangensis, a new entomopathogenic fungus from Guizhou, China. Phytotaxa 2018, 333, 243–250. [Google Scholar] [CrossRef]
- Ownley, B.H.; Pereira, R.M.; Klingeman, W.E.; Quigley, N.B.; Leckie, B.M. Beauveria bassiana, a dual-purpose biological control with activity against insect pests and plant pathogens. Emerg. Concepts Plant Health Manag. 2004, 2004, 255–269. [Google Scholar]
- Suzuki, A.; Kanaoka, M.; Isogai, A.; Tamura, S.; Murakoshi, S.; Ichinoe, M. Bassianolide, a new insecticidal cyclodepsipeptide from Beauveria bassiana and Verticillium lecanii. Tetrahedron Lett. 1977, 18, 2167–2170. [Google Scholar] [CrossRef]
- Wang, S.; Zhao, Z.; Yun-Ting, S.; Zeng, Z.; Zhan, X.; Li, C.; Xie, T. A review of medicinal plant species with elemene in China. Afr. J. Pharm. Pharmacol. 2012, 6, 3032–3040. [Google Scholar] [CrossRef]
- El Kichaoui, A.; Elnabris, K.; Shafie, A.; Fayyad, N.; Arafa, M.; El Hindi, M. Development of Beauveria bassiana based bio-fungicide against Fusarium wilt pathogens for Capsicum annuum, a promising approach toward vital biocontrol industry in Gaza strip. IUG J. Nat. Stud. 2017, 25, 183–190. [Google Scholar]
- Vining, L.C.; Kelleher, W.J.; Schwarting, A.E. Oosporein production by a strain of Beauveria bassiana originally identified as Amanita muscaria. Can. J. Microbiol. 1962, 8, 931–933. [Google Scholar] [CrossRef]
- Hamill, R.L.; Higgens, C.E.; Boaz, H.E.; Gorman, M. The structure of beauvericin, a new depsipeptide antibiotic toxic to Artemia salina. Tetrahedron Lett. 1969, 10, 4255–4258. [Google Scholar] [CrossRef]
- Logrieco, A.; Moretti, A.; Castella, G.; Kostecki, M.; Golinski, P.; Ritieni, A.; Chelkowski, J. Beauvericin production by Fusarium species. Appl. Environ. Microbiol. 1998, 64, 3084–3088. [Google Scholar] [CrossRef]
- Nagaoka, T.; Nakata, K.; Kouno, K. Antifungal activity of oosporein from an antagonistic fungus against Phytophthora infestans. Z. Naturforschung C 2004, 59, 302–304. [Google Scholar] [CrossRef][Green Version]
- Wang, Q.; Xu, L. Beauvericin, a bioactive compound produced by fungi: A short review. Molecules 2012, 17, 2367–2377. [Google Scholar] [CrossRef]
- Manning, R.O.; Wyatt, R.D. Comparative toxicity of Chaetomium contaminated corn and various chemical forms of oosporein in broiler chicks. Poult. Sci. 1984, 63, 251–259. [Google Scholar] [CrossRef]
- Chiron, N.; Michelot, D. Mushrooms odors, chemistry and role in the biotic interactions—A review. Cryptogr. Mycol. 2005, 26, 299–365. [Google Scholar]
- Effmert, U.; Kalderás, J.; Warnke, R.; Piechulla, B. Volatile Mediated Interactions Between Bacteria and Fungi in the Soil. J. Chem. Ecol. 2012, 38, 665–703. [Google Scholar] [CrossRef] [PubMed]
- Mannaa, M.; Kim, K.D. Biocontrol Activity of Volatile-Producing Bacillus megaterium and Pseudomonas protegens Against Aspergillus and Penicillium spp. Predominant in Stored Rice Grains: Study II. Mycobiology 2018, 27, 52–63. [Google Scholar] [CrossRef] [PubMed]
- Saravanakumar, K.; Chelliah, R.; Hu, X.; Oh, D.-H.; Kathiresan, K.; Wang, M.-H. Antioxidant, Anti-Lung Cancer, and Anti-Bacterial Activities of Toxicodendron vernicifluum. Biomolecules 2019, 9, 127. [Google Scholar] [CrossRef] [PubMed]
- Epsky, N.D.; Heath, R.R.; Dueben, B.D.; Lauzon, C.R.; Proveaux, A.T.; Maccollom, G.B. Attraction of 3-methyl-l-butanol and ammonia identified from Enterobacter agglomerans to Anastrepha suspensa. J. Chem. Ecol. 1998, 24, 1867–1880. [Google Scholar] [CrossRef]
- Salih, L.M.; Eid, F.A.; Elhaw, M.H.; Hamed, A. In vitro cytotoxic, antioxidant, antimicrobial activity and volatile constituents of Coccoloba peltata Schott cultivated in Egypt. Egypt. J. Chem. 2021, 64, 7157–7163. [Google Scholar]
- Golembiovska, O.; Tsurkan, A.; Vynogradov, B. Components of Prunella vulgaris L. Grown in Ukraine. J. Pharmacogn. Phytochem. 2014, 2, 140–146. [Google Scholar]
- Bernotienë, G.; Nivinskienë, O.; Butkienë, R.; Mockutë, D. Chemical composition of essential oils of hops (Humulus lupulus L.) growing wild in Aukštaitija. Chemija 2004, 15, 31–36. [Google Scholar]
- Fischedick, J.T.; Hazekamp, A.; Erkelens, T.; Choi, Y.H.; Verpoorte, R. Metabolic fingerprinting of Cannabis sativa L., cannabinoids and terpenoids for chemotaxonomic and drug standardization purposes. Phytochemistry 2010, 71, 2058–2073. [Google Scholar] [CrossRef]
- Sieniawska, E.; Sawicki, R.; Golus, J.; Swatko-Ossor, M.; Ginalska, G.; Skalicka-Wozniak, K. Nigella damascena L. essential Oil—A valuable source of β-Elemene for antimicrobial testing. Molecules 2018, 23, 256. [Google Scholar] [CrossRef]
- Mendanha, S.A.; Alonso, A. Effects of terpenes on fluidity and lipid extraction in phospholipid membranes. Biophys. Chem. 2015, 198, 45–54. [Google Scholar] [CrossRef]
- Barrero, A.F.; Quilez del Moral, J.F.; Lara, A.; Herrador, M.M. Antimicrobial activity of sesquiterpenes from the essential oil of Juniperus thurifera Wood. Planta Med. 2005, 71, 67–71. [Google Scholar] [CrossRef]
- Drage, S.; Mitter, B.; Tröls, C.; Muchugi, A.; Jamnadass, R.H.; Sessitsch, A.; Hadacek, F. Antimicrobial drimane sesquiterpenes and their effect on endophyte communities in the medical tree Warburgiau gandensis. Front. Microbiol. 2014, 5, 13. [Google Scholar] [CrossRef]
- Monga, A.; Sharma, A. Chapter 9—Natural products encompassing antituberculosis activities. Stud. Nat. Prod. Chem. 2020, 64, 263–301. [Google Scholar]
- Zhai, B.; Zhang, N.; Han, X.; Li, Q.; Zhang, M.; Chen, X.; Li, G.; Zhang, R.; Chen, P.; Wang, W.; et al. Molecular targets of β-elemene, a herbal extract used in traditional Chinese medicine, and its potential role in cancer therapy: A review. Biomed. Pharmacoth. 2019, 14, 108812. [Google Scholar] [CrossRef] [PubMed]
- Dong, T.; Yan, Y.; Chai, H.; Chen, S.; Xiong, X.; Sun, D.; Yu, Y.; Deng, L.; Cheng, F. Pyruvate kinase M2 affects liver cancer cell behavior through up-regulation of HIF-1α and Bcl-xL in culture. Biomed. Pharmacother. 2015, 69, 277–284. [Google Scholar] [CrossRef] [PubMed]
- van Niekerk, G.; Engelbrecht, A.M. Role of PKM2 in directing the metabolic fate of glucose in cancer: A potential therapeutic target. Cell. Oncol. 2018, 41, 343–351. [Google Scholar] [CrossRef]
- Sun, X.; Peng, Y.; Zhao, J.; Xie, Z.; Lei, X.; Tang, G. Discovery and development of tumor glycolysis rate-limiting enzyme inhibitors. Bioorg. Chem. 2021, 112, 104891. [Google Scholar] [CrossRef]
- Pan, Y.; Wang, W.; Huang, S.; Ni, W.; Wei, Z.; Cao, Y.; Yu, S.; Jia, Q.; Wu, Y.; Chai, C.; et al. Beta-elemene inhibits breast cancer metastasis through blocking pyruvate kinase M2 dimerization and nuclear translocation. J. Cell. Mol. Med. 2019, 23, 6846–6858. [Google Scholar] [CrossRef]
- Glass, N.L.; Donaldson, G.C. Development of primer sets designed for use with the PCR to amplify conserved genes from filamentous ascomycetes. Appl. Environ. Microbiol. 1995, 61, 1323–1330. [Google Scholar] [CrossRef]
- White, T.J.; Bruns, T.; Lee, S.; Taylor, J.W. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols: A Guide to Methods and Applications; Innis, M.A., Gelfand, D.H., Sninsky, J.J., White, T.J., Eds.; Academic Press: New York, NY, USA, 1990; pp. 315–322. [Google Scholar]
- Johnson, D.M.; White, R.L.; Pereira, R.M.; Geden, C.J. Beauveria bassiana culturing and harvesting for bioassays with house flies. J. Insect Sci. 2020, 20, 14. [Google Scholar] [CrossRef] [PubMed]
- Elshafie, H.S.; Racioppi, R.; Bufo, S.A.; Camele, I. In vitro study of biological activity of four strains of Burkholderia gladioli pv. agaricicola and identification of their bioactive metabolites using GC–MS. Saudia J. Biol. Sci. 2017, 24, 295–301. [Google Scholar]
- Elshafie, H.S.; Viggiani, L.; Mostafa, M.S.; El-Hashash, M.A.; Bufo, S.A.; Camele, I. Biological activity and chemical identification of ornithine lipid produced by Burkholderia gladioli pv. agaricicola ICMP 11096 using LC-MS and NMR analyses. J. Biol. Res. 2017, 90, 96–103. [Google Scholar]
- Sofo, A.; Elshafie, H.S.; Scopa, A.; Mang, S.M.; Camele, I. Impact of airborne zinc pollution on the antimicrobial activity of olive oil and the microbial metabolic profiles of Zn-contaminated soils in an Italian olive orchard. J. Trace Elem. Med. Biol. 2018, 49, 276–284. [Google Scholar] [CrossRef]
- Wan, M.G.; Li, G.Q.; Zhang, J.B.; Jiang, D.H.; Huang, H.C. Effect of volatile substances of Streptomyces platensis F-1 on control of plant fungal diseases. Biol. Control 2008, 46, 552–559. [Google Scholar] [CrossRef]
- Elshafie, H.S.; Bufo, S.A.; Racioppi, R.; Camele, I. Biochemical Characterization of Volatile Secondary Metabolites Produced by Burkholderia gladioli pv. agaricicola. Int. J. Drug Discov. 2013, 5, 181–184. [Google Scholar]
- Elshafie, H.S.; Camele, I.; Racioppi, R.; Scrano, L.; Iacobellis, N.S.; Bufo, S.A. In vitro antifungal activity of Burkholderia gladioli pv. agaricicola against some Phytopathogenic fungi. Int. J. Mol. Sci. 2012, 13, 16291–16302. [Google Scholar] [PubMed]



{kind=link}
{kind=link}
| Tested Bacteria | Diameter of Inhibition Zones (mm) | |||
|---|---|---|---|---|
| Exo-ME 16 mg/mL | Endo-ME 20 mg/mL | Tetracycline 1600 µg/mL | ||
| G+ve | B. cereus | 8.5 ± 1.0 ab | 0.0 ± 0.0 c | 20.8 ± 1.1 b |
| B. megaterium | 10.0 ± 1.9 ab | 4.0 ± 1.7 b | 25.9 ± 2.3 ab | |
| C. michiganensis | 12.5 ± 2.2 a | 0.0 ± 0.0 c | 39.5 ± 2.5 a | |
| G−ve | X. campestris | 9.5 ± 2.5 ab | 9.0 ± 1.9 a | 23.5 ± 1.7 ab |
| P. aeruginosa | 0.0 ± 0.0 c | 6.5 ± 2.8 ab | 10.6 ± 0.7 c | |
| P. fluorescens | 6.5 ± 1.5 b | 4.5 ± 1.7 b | 12.3 ± 0.9 c | |
| Tested Bacteria | Bacterial Growth Inhibition (%) | |||
|---|---|---|---|---|
| GVC | AQS | Tetracycline 1600 µg/mL | ||
| G+ve | B. cereus | 35.0 ± 5.8 c | 60.0 ± 5.8 c | 19.5 ± 1.4 b |
| B. megaterium | 77.5 ± 2.9 a | 92.0 ± 3.5 a | 23.8 ± 2.1 ab | |
| C. michiganensis | 55.0 ± 5.8 b | 77.5 ± 2.9 b | 38.5 ± 2.4 a | |
| G−ve | X. campestris | 27.5 ± 2.9 d | 77.5 ± 8.7 b | 24.3 ± 1.3 ab |
| P. aeruginosa | 37.5 ± 2.9 c | 87.5 ± 2.9 a | 11.2 ± 0.8 c | |
| P. fluorescens | 52.5 ± 2.9 b | 72.5 ± 2.9 b | 13.1 ± 0.7c | |
| Compound a | R.T. b (min) | R.A. c (%) | M.Wt d (g/mol) | Formula | CAS No. | Probability e (%) | Identification f |
|---|---|---|---|---|---|---|---|
| Carbon dioxide | 1.11 | 2.75 ± 0.03 | 44.01 | CO2 | 000124-38-9 | 80 | NIST, IR |
| Benzaldehyde, 2-nitro-, diaminomethylidenhydrazone | 1.21 | 1.24 ± 0.03 | 207.19 | C8H9N5O2 | 102632-31-5 | 43 | NIST, IR |
| Nitrous oxide | 1.48 | 27.57 ± 0.04 | 44.013 | N2O | 010024-97-2 | 65 | NIST, IR |
| Ethanol | 1.57 | 4.69 ± 0.03 | 46.07 | C2H5OH | 000064-17-5 | 90 | NIST, IR |
| Silanol, trimethyl- | 1.68 | 0.66 ± 0.05 | 90.2 | C3H10OSi | 001066-40-6 | 74 | NIST, IR |
| Acetone | 1.77 | 0.72 ± 0.06 | 58.08 | C3H6O | 000067-64-1 | 79 | NIST, IR |
| Formamide, N-methylthio | 2.03 | 0.61 ± 0.06 | 75.14 | C2H5NS | 018952-41-5 | 63 | NIST, IR |
| Butanal, 3-methyl- | 2.71 | 1.32 ± 0.05 | 86.13 | C5H10O | 000590-86-3 | 81 | NIST, IR |
| Butanal, 2-methyl- | 2.83 | 0.44 ± 0.04 | 86.13 | C5H10O | 000096-17-3 | 90 | NIST, IR |
| 1-Butanol, 3-methyl- | 3.88 | 3.72 ± 0.03 | 88.15 | C5H12O | 000123-51-3 | 83 | NIST, IR |
| Arsenous acid, tris(trimethylsilyl) ester | 4.98 | 1.68 ± 0.02 | 342.49 | C9H27AsO3Si3 | 055429-29-3 | 70 | NIST, IR |
| 2,4-Dimethyl-1-heptene | 5.37 | 0.63 ± 0.03 | 126.24 | C9H18 | 019549-87-2 | 90 | NIST, IR |
| Heptane, 2,3-dimethyl- | 5.56 | 0.76 ± 0.01 | 128.25 | C9H20 | 003074-71-3 | 87 | NIST, IR |
| Octane, 4-methyl- | 5.66 | 1.99 ± 0.03 | 128.25 | C9H20 | 002216-34-4 | 93 | NIST, IR |
| Octane, 2,3,6,7-tetramethyl- | 7.30 | 0.19 ± 0.02 | 170.33 | C12H26 | 052670-34-5 | 63 | NIST, IR |
| Oxalic acid, 2-ethylhexyl nonyl ester | 7.33 | 0.21 ± 0.03 | 328.5 | C19H36O4 | 1000309-39-2 | 74 | NIST, IR |
| Dodecane, 2,6,11-trimethyl- | 7.70 | 0.49 ± 0.03 | 212.41 | C15H32 | 031295-56-4 | 69 | NIST, IR |
| Decane, 3,6-dimethyl- | 7.76 | 5.47 ± 0.03 | 170.33 | C12H26 | 017312-53-7 | 72 | NIST, IR |
| Heptane, 2,4-dimethyl- | 7.81 | 2.37 ± 0.02 | 128.25 | C9H20 | 002213-23-2 | 79 | NIST, IR |
| 2-Undecene, 4-methyl- | 7.97 | 1.02 ± 0.03 | 168.32 | C12H24 | 091695-32-8 | 63 | NIST, IR |
| Oxalic acid, isohexyl neopentyl ester | 8.09 | 0.54 ± 0.04 | 244.33 | C13H24O4 | 1000309-73-0 | 64 | NIST, IR |
| Decane, 3,7-dimethyl- | 8.15 | 2.37 ± 0.01 | 170.33 | C12H26 | 017312-54-8 | 87 | NIST, IR |
| Sulfurous acid, hexyl 2-pentyl ester | 8.27 | 0.65 ± 0.04 | 236.37 | C11H24O3S | 1000309-15-6 | 69 | NIST, IR |
| Decane, 2,3,5-trimethyl- | 9.56 | 0.68 ± 0.05 | 184.36 | C13H28 | 062238-11-3 | 80 | NIST, IR |
| Dodecane, 2,6,10-trimethyl- | 9.62 | 0.14 ± 0.03 | 212.41 | C15H32 | 003891-98-3 | 72 | NIST, IR |
| Hexadecane | 9.66 | 0.18 ± 0.04 | 226.44 | C16H34 | 000544-76-3 | 78 | NIST, IR |
| Dodecane, 2,6,10-trimethyl- | 9.72 | 0.30 ± 0.01 | 212.41 | C15H32 | 003891-98-3 | 64 | NIST, IR |
| Heptadecane | 9.89 | 0.40 ± 0.03 | 240.5 | C17H36 | 000629-78-7 | 72 | NIST, IR |
| 7-Chloro-2,3-dihydro-3-(4-N,N-dimethylaminobenzylidene)-5-phenyl-1H-1,4-benzodiazepin-2-one | 10.30 | 0.37 ± 0.01 | 401.9 | C24H20ClN3O | 055056-35-4 | 46 | NIST, IR |
| 3,6-Dioxa-2,4,5,7-tetrasilaoctane,2,2,4,4,5,5,7,7-octamethyl- | 10.39 | 0.20 ± 0.02 | 294.68 | C10H30O2Si4 | 004342-25-0 | 65 | NIST, IR |
| β-elemene g | 10.46 | 6.98 ± 0.03 | 204.35 | C15H24 | 000515-13-9 | 91 | NIST, IR |
| 3-Hydroxybromoazepam, bis(trimethylsilyl)- deriv. | 11.19 | 0.40 ± 0.04 | 476.5 | C20H26BrN3O2Si2 | 1000079-50-7 | 72 | NIST, IR |
| 2-Amino-2-oxo-acetic acid,N-[3,4-dimethylphenyl]-, ethyl ester | 11.42 | 0.22 ± 0.03 | 221.25 | C12H15NO3 | 024451-17-0 | 77 | NIST, IR |
| Diethyl Phthalate | 11.84 | 0.65 ± 0.01 | 222.24 | C12H14O4 | 000084-66-2 | 90 | NIST, IR |
| Bacteria Name | Collection Number | Gram Type |
|---|---|---|
| B. cereus Frankland and Frankland | UniB12421 | G+ve |
| B. megaterium de Bary | UniB12421 | |
| C. michiganensis (Smith) Davis | UniB3718 | |
| X. campestris (Pammel) Dowson | UniB7718 | G−ve |
| P. aeruginosa (Schröter) Migula | UniB02421 | |
| P. fluorescens (Flügge) Migula | UniB05421 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Camele, I.; Sadeek, S.A.; Racioppi, R.; Elshafie, H.S. Antimicrobial Activity of Diffusible and Volatile Metabolites Emitted by Beauveria bassiana: Chemical Profile of Volatile Organic Compounds (VOCs) Using SPME-GC/MS Analysis. Plants 2023, 12, 2854. https://doi.org/10.3390/plants12152854
Camele I, Sadeek SA, Racioppi R, Elshafie HS. Antimicrobial Activity of Diffusible and Volatile Metabolites Emitted by Beauveria bassiana: Chemical Profile of Volatile Organic Compounds (VOCs) Using SPME-GC/MS Analysis. Plants. 2023; 12(15):2854. https://doi.org/10.3390/plants12152854
Chicago/Turabian StyleCamele, Ippolito, Sadeek A. Sadeek, Rocco Racioppi, and Hazem S. Elshafie. 2023. "Antimicrobial Activity of Diffusible and Volatile Metabolites Emitted by Beauveria bassiana: Chemical Profile of Volatile Organic Compounds (VOCs) Using SPME-GC/MS Analysis" Plants 12, no. 15: 2854. https://doi.org/10.3390/plants12152854
APA StyleCamele, I., Sadeek, S. A., Racioppi, R., & Elshafie, H. S. (2023). Antimicrobial Activity of Diffusible and Volatile Metabolites Emitted by Beauveria bassiana: Chemical Profile of Volatile Organic Compounds (VOCs) Using SPME-GC/MS Analysis. Plants, 12(15), 2854. https://doi.org/10.3390/plants12152854