

Antibacterial Activity of the Essential Oil of Piper tuberculatum Jacq. Fruits against Multidrug-Resistant Strains: Inhibition of Efflux Pumps and β-Lactamase

 , , ,
, , ,  ,
,  , , , , and
, , , , and 
Abstract
1. Introduction
2. Results
2.1. GC-MS Profile of Piper tuberculatum Essential Oil
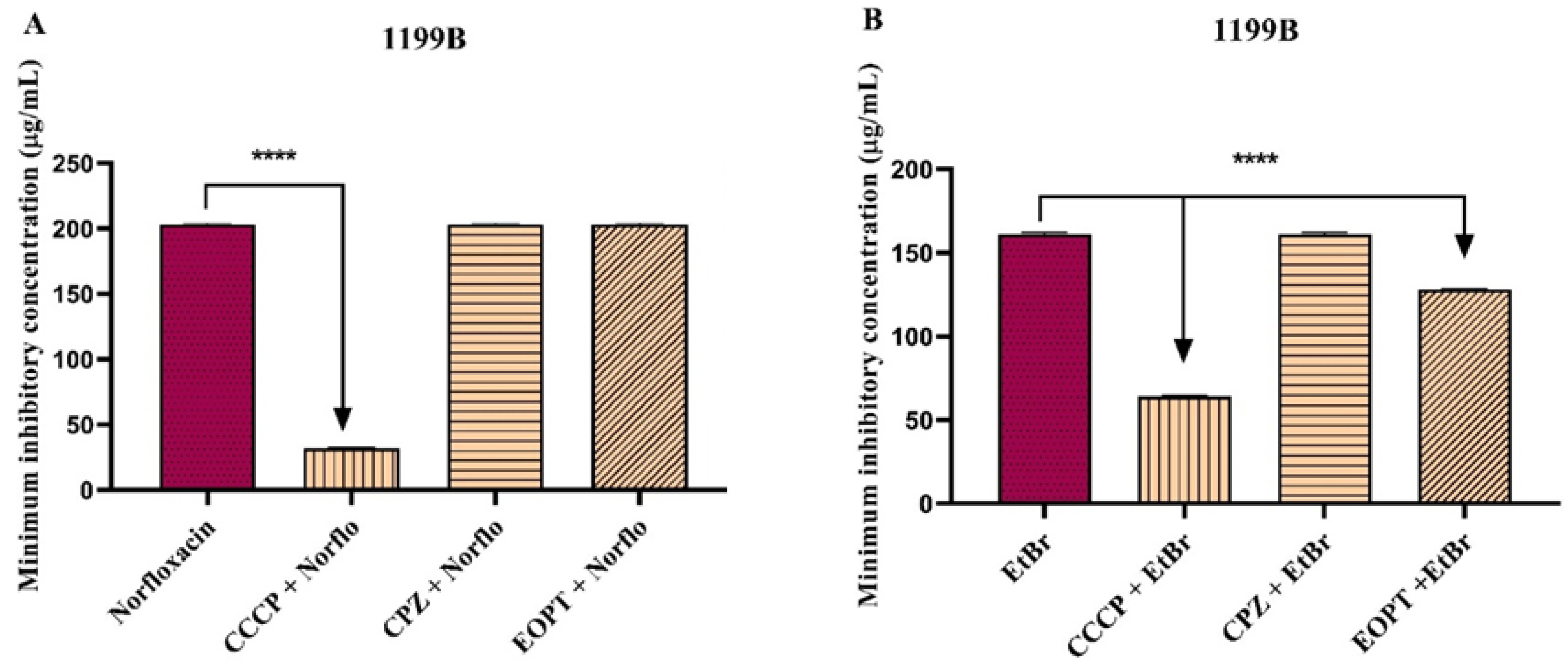
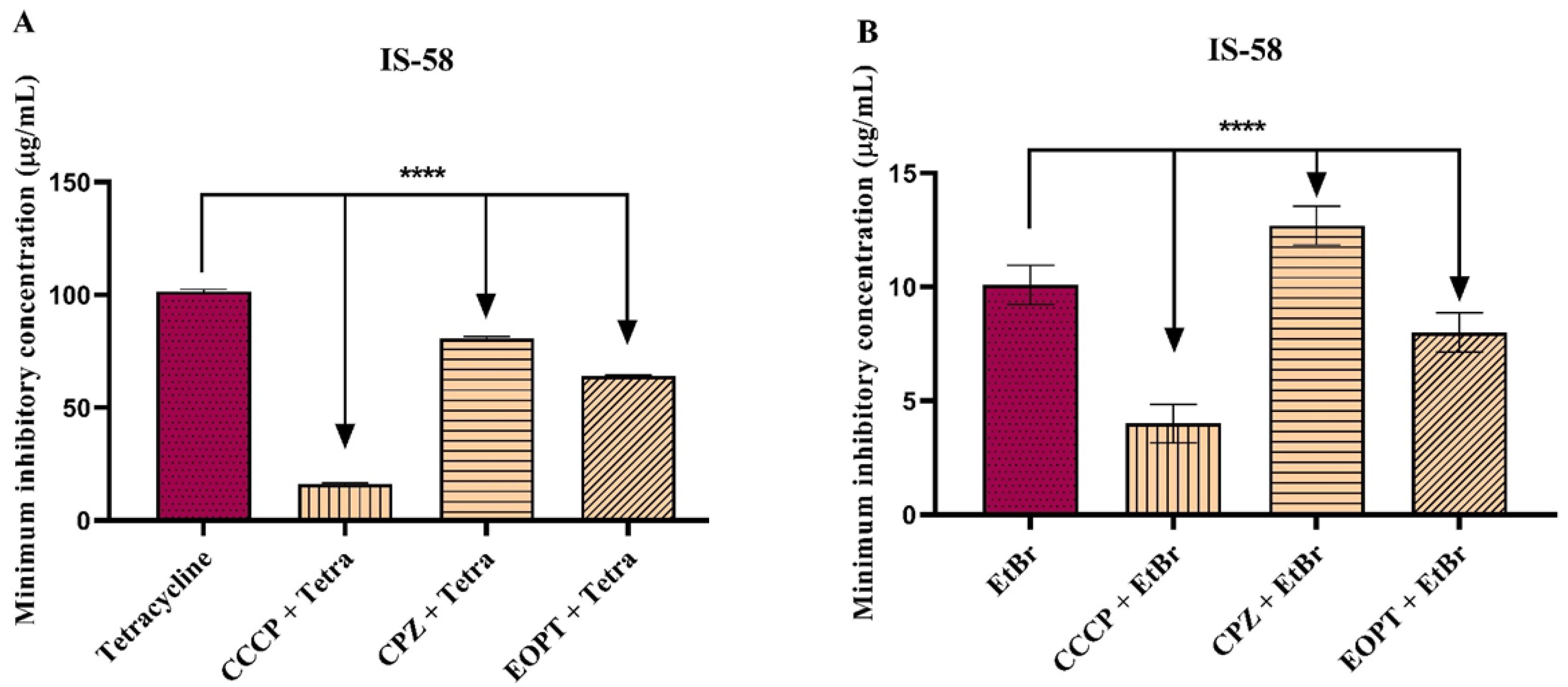
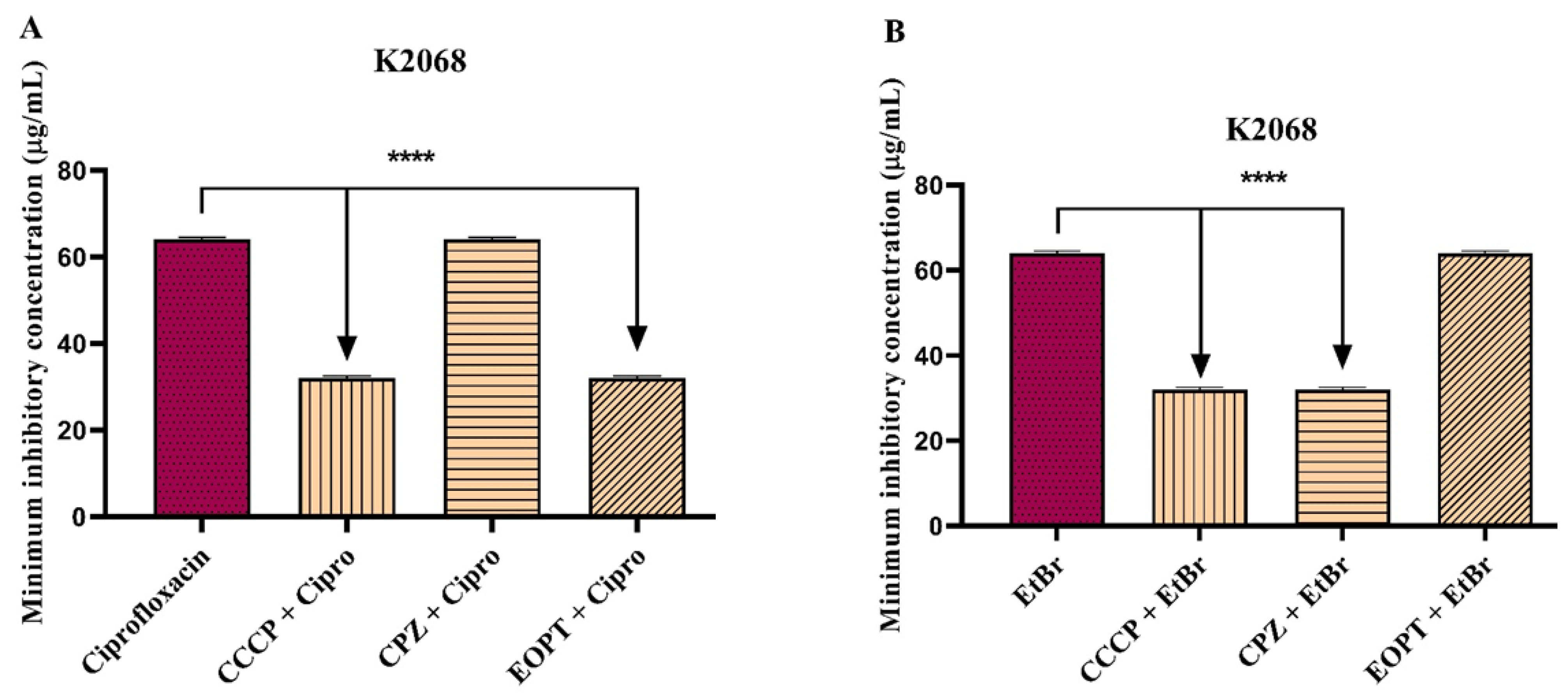
2.2. Antibacterial Activity and Antibiotic Modulation in Efflux Pump-Carrying S. aureus Strains
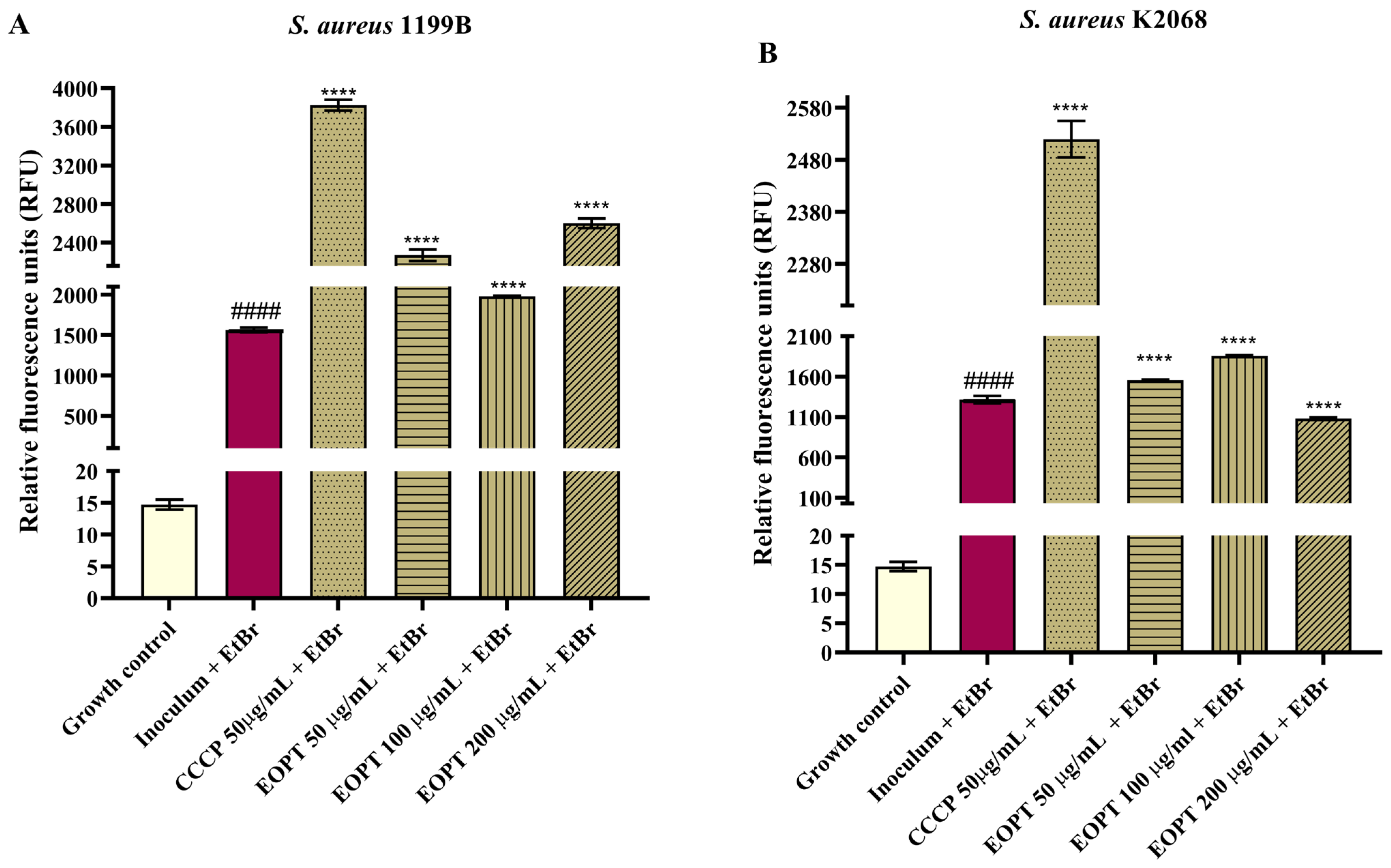
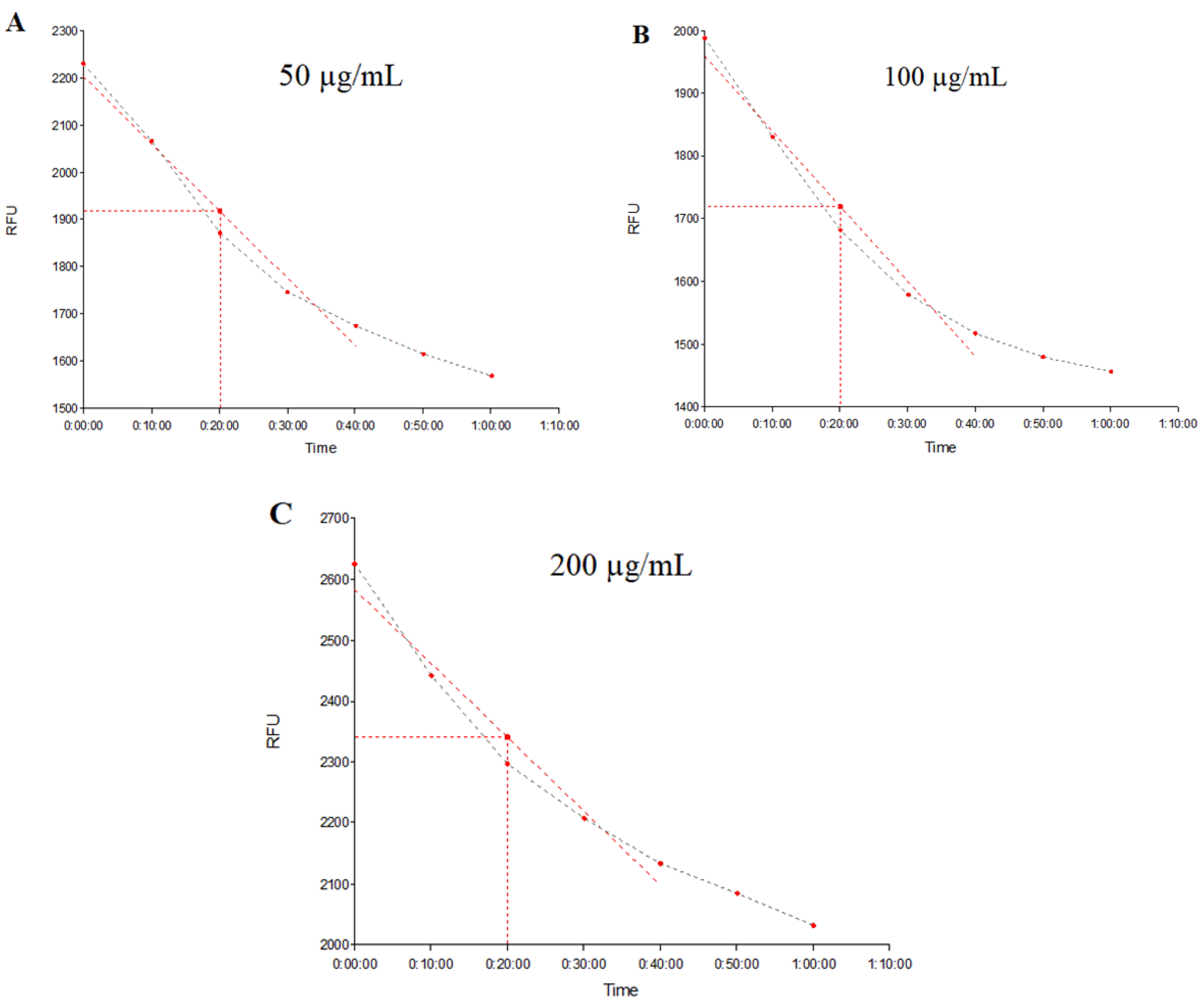
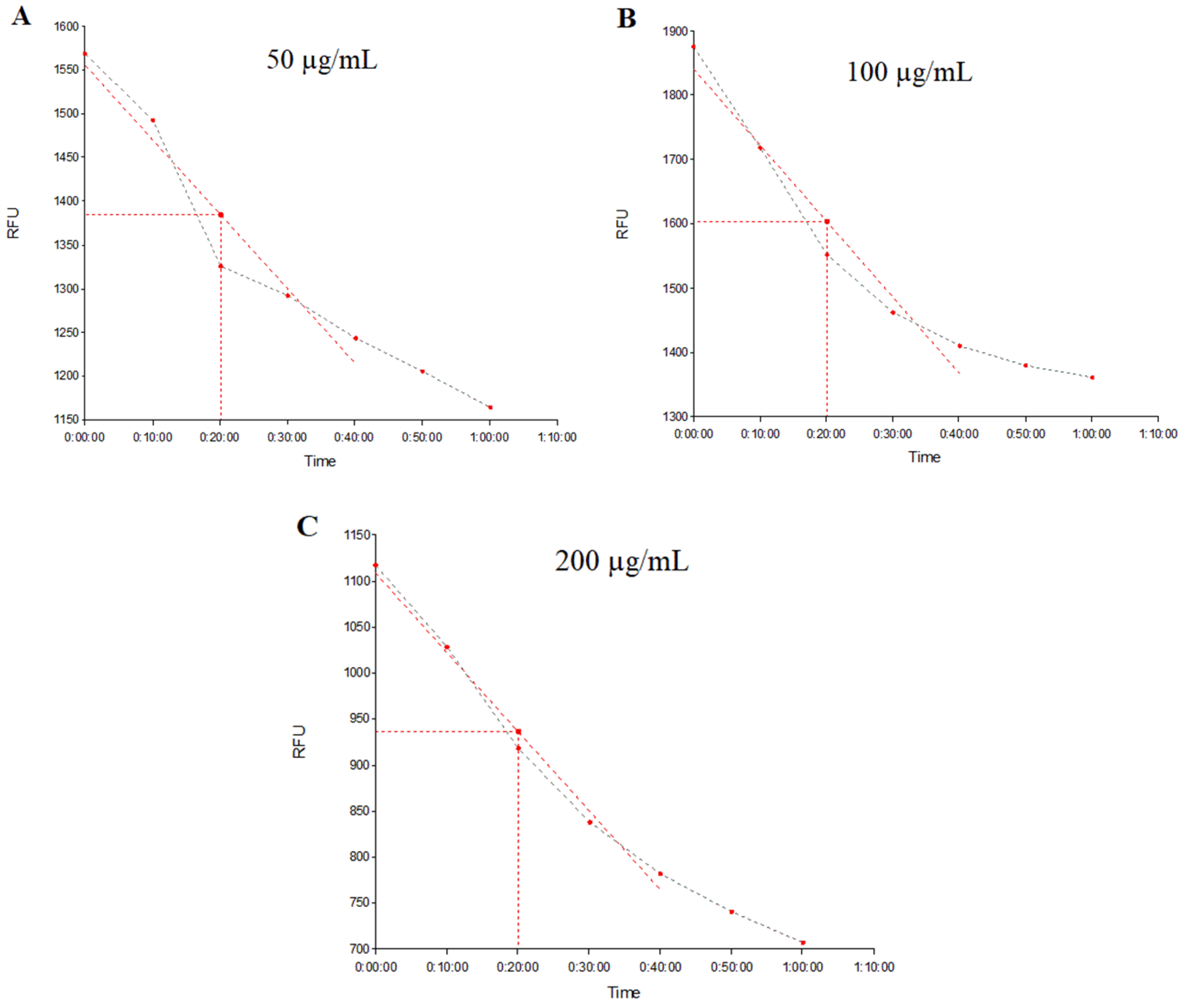
2.3. Evaluation of NorA and MepA Efflux Pump Inhibition by Fluorescence Emission
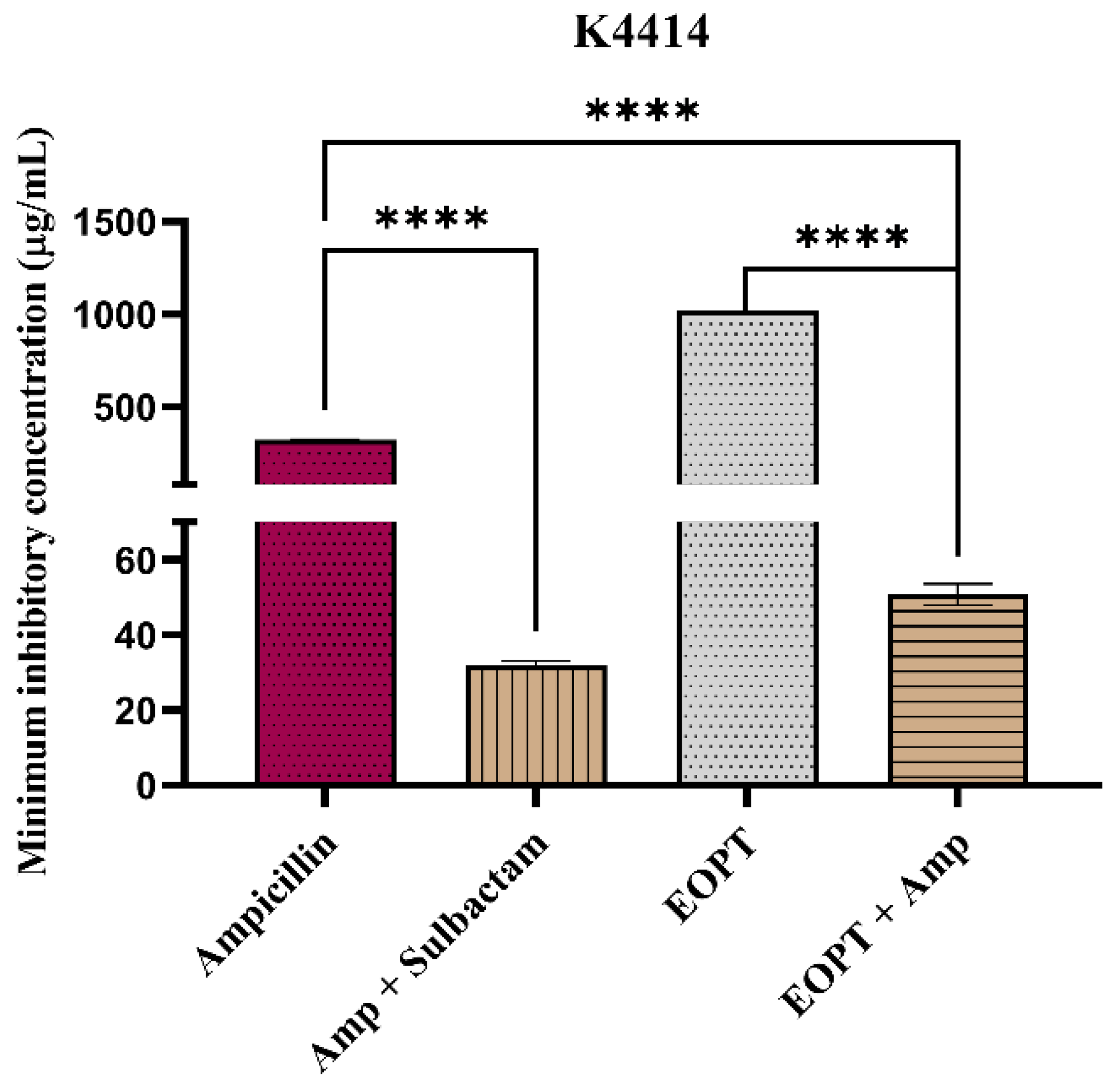
2.4. Evaluation of β-Lactamase Inhibition
3. Discussion
4. Materials and Methods
4.1. Plant Material and Extraction of the Essential Oil
4.2. Determination of the Chemical Profile of the Piper tuberculatum Essential oil via CG-MS
4.3. Bacterial Strains and Culture Media
4.4. Substances
4.5. Minimum Inhibitory Concentration (MIC) Determination
4.6. Evaluation of Efflux Pump Inhibition via Modulation of the MIC of the Antibiotics and EtBr
4.7. Analysis of NorA and MepA Inhibition by Increased EtBr Fluorescence Emission
4.8. Evaluation of β-Lactamase Inhibition
4.9. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Turner, N.A.; Sharma-Kuinkel, B.K.; Maskarinec, S.A.; Eichenberger, E.M.; Shah, P.P.; Carugati, M.; Holland, T.L.; Fowler, V.G. Methicillin-Resistant Staphylococcus aureus: An Overview of Basic and Clinical Research. Nat. Rev. Microbiol. 2019, 17, 203–218. [Google Scholar] [CrossRef] [PubMed]
- Chambers, H.F.; DeLeo, F.R. Waves of Resistance: Staphylococcus aureus in the Antibiotic Era. Nat. Rev. Microbiol. 2009, 7, 629–641. [Google Scholar] [CrossRef] [PubMed]
- Archer, G.L. Staphylococcus aureus: A Well-Armed Pathogen. Clin. Infect. Dis. 1998, 26, 1179–1181. [Google Scholar] [CrossRef]
- El-Tarabily, K.A.; El-Saadony, M.T.; Alagawany, M.; Arif, M.; Batiha, G.E.; Khafaga, A.F.; Elwan, H.A.M.; Elnesr, S.S.; Abd El-Hack, M.E. Using Essential Oils to Overcome Bacterial Biofilm Formation and Their Antimicrobial Resistance. Saudi J. Biol. Sci. 2021, 28, 5145–5156. [Google Scholar] [CrossRef]
- Lucien, M.A.B.; Canarie, M.F.; Kilgore, P.E.; Jean-Denis, G.; Fénélon, N.; Pierre, M.; Cerpa, M.; Joseph, G.A.; Maki, G.; Zervos, M.J.; et al. Antibiotics and Antimicrobial Resistance in the COVID-19 Era: Perspective from Resource-Limited Settings. Int. J. Infect. Dis. 2021, 104, 250–254. [Google Scholar] [CrossRef] [PubMed]
- Nishino, K.; Yamasaki, S.; Nakashima, R.; Zwama, M.; Hayashi-Nishino, M. Function and Inhibitory Mechanisms of Multidrug Efflux Pumps. Front. Microbiol. 2021, 12, 737288. [Google Scholar] [CrossRef] [PubMed]
- Lima, L.M.; da Silva, B.N.M.; Barbosa, G.; Barreiro, E.J. β-Lactam Antibiotics: An Overview from a Medicinal Chemistry Perspective. Eur. J. Med. Chem. 2020, 208, 112829. [Google Scholar] [CrossRef]
- Lamut, A.; Peterlin Mašič, L.; Kikelj, D.; Tomašič, T. Efflux Pump Inhibitors of Clinically Relevant Multidrug Resistant Bacteria. Med. Res. Rev. 2019, 39, 2460–2504. [Google Scholar] [CrossRef]
- Sharma, A.; Gupta, V.; Pathania, R. Efflux Pump Inhibitors for Bacterial Pathogens: From Bench to Bedside. Indian J. Med. Res. 2019, 149, 129. [Google Scholar] [CrossRef]
- González-Bello, C.; Rodríguez, D.; Pernas, M.; Rodríguez, Á.; Colchón, E. β-Lactamase Inhibitors to Restore the Efficacy of Antibiotics against Superbugs. J. Med. Chem. 2020, 63, 1859–1881. [Google Scholar] [CrossRef]
- Süntar, I. Importance of Ethnopharmacological Studies in Drug Discovery: Role of Medicinal Plants. Phytochem. Rev. 2020, 19, 1199–1209. [Google Scholar] [CrossRef]
- Balunas, M.J.; Kinghorn, A.D. Drug Discovery from Medicinal Plants. Life Sci. 2005, 78, 431–441. [Google Scholar] [CrossRef] [PubMed]
- Santos, M.O.; Ribeiro, D.A.; de Macêdo, D.G.; Macêdo, M.J.F.; Macedo, J.G.F.; de Lacerda, M.N.S.; Macêdo, M.S.; Souza, M.M.A. Medicinal Plants: Versatility and Concordance of Use in the Caatinga Area, Northeastern Brazil. Acad. Bras Cienc 2018, 90, 2767–2779. [Google Scholar] [CrossRef] [PubMed]
- Melo-Batista, A.A.D.; Oliveira, C.R.M.D. Plantas Utilizadas Como Medicinais Em Uma Comunidade Do Semiárido Baiano: Saberes Tradicionais E a Conservação Ambiental. Enciclopédia Biosf. 2014, 10, 74–88. [Google Scholar]
- de Lima, R.S.; Lima, R.A. Levantamento Bibliográfico Do Gênero Piper (Piperaceae) Para Uso Medicinal. Educ -Educ. Soc. Meio Ambiente 2021, 13, 155–170. [Google Scholar]
- da Silva, A.H.; Vieira, R.W.R.; Costa, F.A.F.; Nunes, L.E.; Pinto, D.S.; Vasconcelos, C.J.I.; de Oliveira, C.M.C.; Ramalho, C.R.M. Antimicrobial activity of products obtained from Piper species. Rev. Cuba. Plantas Med. 2016, 21, 168–180. [Google Scholar]
- dos Santos Sales, V.; do Nascimento, E.P.; Monteiro, Á.B.; da Costa, M.H.N.; de Araújo Delmondes, G.; Soares, T.R.C.; Tintino, S.R.; Sobreira, F.R.; de Figuêiredo, D.N.; de Souza Rodrigues, C.K. In Vitro Modulation of the Antibiotic Activity of Essential Oil from Fruits of Piper Tuberculatum Jacq. Rev. Cuba. Plantas Med. 2017, 22, 1–10. [Google Scholar]
- Oliveira, A.K.M.; Pauliquevis, C.F.; Matias, R.; Silva, P.G.; Zanella, D.d.F.P.; Roel, A.R.; Porto, K.R. Efeito Larvicida Do Extrato Etanólico de Piper Umbellatum Sobre o Mosquito Aedes Aegypti. S. Am. J. Basic Educ. Tech. Technol. 2021, 8, 84–101. [Google Scholar]
- Oliveira, F.A.d.S.; Passarini, G.M.; de Medeiros, D.S.S.; Santos, A.P.d.A.; Fialho, S.N.; de Gouveia, A.J.; Latorre, M.; Freitag, E.M.; Medeiros, P.S.D.M.D.; Teles, C.B.G.; et al. Antiplasmodial and Antileishmanial Activities of Compounds from Piper tuberculatum Jacq Fruits. Rev. Soc Bras Med. Trop. 2018, 51, 382–386. [Google Scholar] [CrossRef]
- Dhifi, W.; Bellili, S.; Jazi, S.; Bahloul, N.; Mnif, W. Essential Oils’ Chemical Characterization and Investigation of Some Biological Activities: A Critical Review. Medicines 2016, 3, 25. [Google Scholar] [CrossRef]
- Sidhu, M.S.; Heir, E.; Sørum, H.; Holck, A. Genetic Linkage between Resistance to Quaternary Ammonium Compounds and β-Lactam Antibiotics in Food-Related Staphylococcus spp. Microb. Drug Resist. 2001, 7, 363–371. [Google Scholar] [CrossRef] [PubMed]
- Sidhu, M.S.; Heir, E.; Leegaard, T.; Wiger, K.; Holck, A. Frequency of Disinfectant Resistance Genes and Genetic Linkage with -Lactamase Transposon Tn552 among Clinical Staphylococci. Antimicrob. Agents Chemother. 2002, 46, 2797–2803. [Google Scholar] [CrossRef] [PubMed]
- Salehi, B.; Zakaria, Z.A.; Gyawali, R.; Ibrahim, S.A.; Rajkovic, J.; Shinwari, Z.K.; Khan, T.; Sharifi-Rad, J.; Ozleyen, A.; Turkdonmez, E.; et al. Piper Species: A Comprehensive Review on Their Phytochemistry, Biological Activities and Applications. Molecules 2019, 24, 1364. [Google Scholar] [CrossRef]
- Thin, D.B.; Chinh, H.V.; Luong, N.X.; Hoi, T.M.; Dai, D.N.; Ogunwande, I.A. Chemical Analysis of Essential Oils of Piper laosanum and Piper acre (Piperaceae) from Vietnam. J. Essent. Oil Bear. Plants 2018, 21, 181–188. [Google Scholar] [CrossRef]
- Dos Santos Sales, V.; Monteiro, Á.B.; Delmondes, G.d.A.; do Nascimento, E.P.; Sobreira Dantas Nóbrega de Figuêiredo, F.R.; de Souza Rodrigues, C.K.; Evangelista de Lacerda, J.F.; Fernandes, C.N.; Barbosa, M.d.O.; Brasil, A.X.; et al. Antiparasitic Activity and Essential Oil Chemical Analysis of the Piper tuberculatum Jacq Fruit. Iran J. Pharm. Res. 2018, 17, 268–275. [Google Scholar] [PubMed]
- Monteiro, M. Antibacterial Activity of the Piper Aduncum Oil and Dillapiole, Its Main Constituent, Against Multidrug-Resistant Strains. Bol. Lat. Caribe Plantas Med. Aromat 2014, 13, 517–526. [Google Scholar]
- Leal, A.L.A.B.; Bezerra, C.F.; Confortin, C.; da Silva, L.E.; Marinho, E.M.; Marinho, M.M.; Vasconcelos, M.A.; da Silva, T.G.; Marinho, E.S.; Teixeira, A.M.R.; et al. Chemical Composition and Potentiating Action of Norfloxacin Mediated by the Essential Oil of Piper Caldense C.D.C. against Staphylococcus aureus Strains Overexpressing Efflux Pump Genes. Arch. Microbiol. 2021, 203, 4727–4736. [Google Scholar] [CrossRef]
- Carneiro, J.N.P.; da Cruz, R.P.; Campina, F.F.; Costa, M.d.S.; dos Santos, A.T.L.; Sales, D.L.; Bezerra, C.F.; da Silva, L.E.; de Araujo, J.P.; do Amaral, W.; et al. GC/MS Analysis and Antimicrobial Activity of the Piper mikanianum (Kunth) Steud. Essential Oil. Food Chem. Toxicol. 2020, 135, 110987. [Google Scholar] [CrossRef]
- Mahizan, N.A.; Yang, S.-K.; Moo, C.-L.; Song, A.A.; Chong, C.-M.; Chong, C.-W.; Abushelaibi, A.; Lim, S.-H.E.; Lai, K.-S. Terpene Derivatives as a Potential Agent against Antimicrobial Resistance (AMR) Pathogens. Molecules 2019, 24, 2631. [Google Scholar] [CrossRef]
- Salehi, B.; Upadhyay, S.; Erdogan Orhan, I.; Kumar Jugran, A.; Jayaweera, S.L.D.; Dias, D.A.; Sharopov, F.; Taheri, Y.; Martins, N.; Baghalpour, N.; et al. Therapeutic Potential of α- and β-Pinene: A Miracle Gift of Nature. Biomolecules 2019, 9, 738. [Google Scholar] [CrossRef]
- Farré-Armengol, G.; Filella, I.; Llusià, J.; Peñuelas, J. β-Ocimene, a Key Floral and Foliar Volatile Involved in Multiple Interactions between Plants and Other Organisms. Molecules 2017, 22, 1148. [Google Scholar] [CrossRef] [PubMed]
- Milenković, A.N.; Stanojević, J.S.; Troter, D.Z.; Pejčić, M.G.; Stojanović-Radić, Z.Z.; Cvetković, D.J.; Stanojević, L.P. Chemical composition, antimicrobial and antioxidant activities of essential oils isolated from black (Piper nigrum L.) and cubeb pepper (Piper cubeba L.) fruits from the Serbian market. J. Essent. Oil Res. 2023, 35, 262–273. [Google Scholar] [CrossRef]
- Oyedeji, O.A.; Adeniyi, B.A.; Ajayi, O.; König, W.A. Essential Oil Composition of Piper guineense and Its Antimicrobial Activity. Another Chemotype from Nigeria. Phytother. Res. Int. J. Devoted Pharmacol. Toxicol. Eval. Nat. Prod. Deriv. 2005, 19, 362–364. [Google Scholar]
- Braga, A.L.; da Cruz, R.P.; Carneiro, J.N.P.; dos Santos, A.T.L.; Sales, D.L.; Bezerra, C.F.; Fonseca, V.J.A.; Rocha, J.E.; de Freitas, T.S.; Campina, F.F.; et al. Piper regnellii (Miq.) C. DC.: Chemical Composition, Antimicrobial Effects, and Modulation of Antimicrobial Resistance. S. Afr. J. Bot. 2021, 142, 495–501. [Google Scholar] [CrossRef]
- Singh, S.; Kapoor, I.P.S.; Singh, G.; Schuff, C.; De Lampasona, M.P.; Catalan, C.A.N. Chemistry, Antioxidant and Antimicrobial Potentials of White Pepper (Piper nigrum L.) Essential Oil and Oleoresins. Proc. Natl. Acad. Sci. USA India Sect. B Biol. Sci. 2013, 83, 357–366. [Google Scholar] [CrossRef]
- Dorman, H.; Deans, S.G. Antimicrobial Agents from Plants: Antibacterial Activity of Plant Volatile Oils. J. Appl. Microbiol. 2000, 88, 308–316. [Google Scholar] [CrossRef]
- De, A.; Morandim-Giannetti, A.; Pin, A.R.; Ama, N.; Pietro, S.; Cesar De Oliveira, H.; Soares Mendes-Giannini, M.J.; Alecio, A.C.; Kato, M.J.; De Oliveira, J.E.; et al. Composition and Antifungal Activity against Candida albicans, Candida aarapsilosis, Candida krusei and Cryptococcus Neoformans of Essential Oils from Leaves of Piper and Peperomia Species. J. Med. Plants Res. 2010, 4, 1810–1814. [Google Scholar] [CrossRef]
- Gomes Pinto, E.; Jacobi, A.; Lana, D.; Lima, R.A. Estudo Fitoquímico Do Extrato Etanólico De Piper Tuberculatum JACQ. Sobre Cepas De Escherichia coli in Vitro Phytochemical Study of The Ethanolic Extract of Piper tuberculatum JACQ. on Strains of Escherichia coli in Vitro. S. Am. J. Basic Educ. Tech. Technol. 2016, 3. [Google Scholar]
- Restrepo Osorio, J.; Colmenares Dulcey, A.J.; Mora, L.E.; Sánchez Andica, R. Extraction, Chemical Composition and Antimicrobial Activity of The Essential Oils of Pipilongo (Piper tuberculatum) Using Supercritical Carbon Dioxide. Rev. Cienc. 2015, 17, 45–56. [Google Scholar] [CrossRef]
- Palacios, Z.G.F.; Delgado, G.E.; Moreno, M.C.; Consuelo Rojas, M.J.K. In Vitro Antifungal Activity of Crude Extracts of Piper tuberculatum. Rev. Peru Biol. 2009, 16, 209–214. [Google Scholar]
- Rezende Lara Júnior, C.; Lopes de Oliveira, G.; Caroline Ferreira Mota, B.; de Fátima Gonçalves Fernandes, M.; Figueiredo, L.S.; Martins, E.R.; de Lima Moreira, D.; Auxiliadora Coelho Kaplan, M. Antimicrobial Activity of Essential Oil of Piper aduncum L. (Piperaceae). J. Med. Plants Res. 2012, 6, 3800–3805. [Google Scholar] [CrossRef]
- Rahman, M.M.; Haque, M. Antioxidant, Antibacterial and Antifungal Properties of Black Pepper Essential Oil (Piper nigrum Linn.) and Molecular Docking and Pharmacokinetic Studies of Its’ Major Component. Artic. Orient. J. Chem. 2022, 38, 1554–1560. [Google Scholar] [CrossRef]
- Bezerra, J.W.A.; Rodrigues, F.C.; da Cruz, R.P.; da Silva, L.E.; do Amaral, W.; Rebelo, R.A.; Begnini, I.M.; Bezerra, C.F.; Iriti, M.; Varoni, E.M.; et al. Antibiotic Potential and Chemical Composition of the Essential Oil of Piper Caldense C. DC. (Piperaceae). Appl. Sci. 2020, 10, 631. [Google Scholar] [CrossRef]
- Noriega, P.; Ballesteros, J.; De la Cruz, A.; Veloz, T. Chemical Composition and Preliminary Antimicrobial Activity of the Hydroxylated Sesquiterpenes in the Essential Oil from Piper barbatum Kunth Leaves. Plants 2020, 9, 211. [Google Scholar] [CrossRef] [PubMed]
- Chouhan, S.; Sharma, K.; Guleria, S. Antimicrobial Activity of Some Essential Oils—Present Status and Future Perspectives. Medicines 2017, 4, 58. [Google Scholar] [CrossRef] [PubMed]
- Chitsaz, M.; Brown, M.H. The Role Played by Drug Efflux Pumps in Bacterial Multidrug Resistance. Essays Biochem. 2017, 61, 127–139. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Venter, H.; Ma, S. Efflux Pump Inhibitors: A Novel Approach to Combat Efflux-Mediated Drug Resistance in Bacteria. Curr. Drug Targets 2015, 17, 702–719. [Google Scholar] [CrossRef] [PubMed]
- Shriram, V.; Khare, T.; Bhagwat, R.; Shukla, R.; Kumar, V. Inhibiting Bacterial Drug Efflux Pumps via Phyto-Therapeutics to Combat Threatening Antimicrobial Resistance. Front. Microbiol. 2018, 9, 2990. [Google Scholar] [CrossRef]
- Wright, G.D. Bacterial Resistance to Antibiotics: Enzymatic Degradation and Modification. Adv. Drug Deliv. Rev. 2005, 57, 1451–1470. [Google Scholar] [CrossRef]
- Michiels, J.E.; Van den Bergh, B.; Verstraeten, N.; Michiels, J. Molecular Mechanisms and Clinical Implications of Bacterial Persistence. Drug Resist. Updat. 2016, 29, 76–89. [Google Scholar] [CrossRef]
- Abreu, A.C.; McBain, A.J.; Simões, M. Plants as Sources of New Antimicrobials and Resistance-Modifying Agents. Nat. Prod. Rep. 2012, 29, 1007. [Google Scholar] [CrossRef] [PubMed]
- Gutierrez, A.; Stokes, J.M.; Matic, I. Our Evolving Understanding of the Mechanism of Quinolones. Antibiotics 2018, 7, 32. [Google Scholar] [CrossRef] [PubMed]
- Ayaz, M.; Ullah, F.; Sadiq, A.; Ullah, F.; Ovais, M.; Ahmed, J.; Devkota, H.P. Synergistic Interactions of Phytochemicals with Antimicrobial Agents: Potential Strategy to Counteract Drug Resistance. Chem. Biol. Interact. 2019, 308, 294–303. [Google Scholar] [CrossRef] [PubMed]
- Leite, A.M.; Lima, E.d.O.; de Souza, E.L.; Diniz, M.d.F.F.M.; Trajano, V.N.; de Medeiros, I.A. Inhibitory Effect of Beta-Pinene, Alpha-Pinene and Eugenol on the Growth of Potential Infectious Endocarditis Causing Gram-Positive Bacteria. Rev. Bras. Ciências Farm. 2007, 43, 121–126. [Google Scholar] [CrossRef]
- de Araújo, A.C.J.; Freitas, P.R.; dos Santos Barbosa, C.R.; Muniz, D.F.; de Almeida, R.S.; Alencar de Menezes, I.R.; Ribeiro-Filho, J.; Tintino, S.R.; Coutinho, H.D.M. In Vitro and In Silico Inhibition of Staphylococcus Aureus Efflux Pump NorA by α-Pinene and Limonene. Curr. Microbiol. 2021, 78, 3388–3393. [Google Scholar] [CrossRef]
- Agreles, M.A.A.; Cavalcanti, I.D.L.; Cavalcanti, I.M.F. The Role of Essential Oils in the Inhibition of Efflux Pumps and Reversion of Bacterial Resistance to Antimicrobials. Curr Microbiol. 2021, 78, 3609–3619. [Google Scholar] [CrossRef]
- Oliveira-Tintino, C.D.d.M.; Muniz, D.F.; dos Santos Barbosa, C.R.; Silva Pereira, R.L.; Begnini, I.M.; Rebelo, R.A.; da Silva, L.E.; Mireski, S.L.; Nasato, M.C.; Lacowicz Krautler, M.I.; et al. NorA, Tet(K), MepA, and MsrA Efflux Pumps in Staphylococcus aureus, Their Inhibitors and 1,8-Naphthyridine Sulfonamides. Curr. Pharm. Des. 2022, 29, 323–355. [Google Scholar] [CrossRef]
- Blair, J.M.A.; Piddock, L.J.V. How to Measure Export via Bacterial Multidrug Resistance Efflux Pumps. mBio 2016, 7, e00840-16. [Google Scholar] [CrossRef]
- Schindler, B.D.; Jacinto, P.; Kaatz, G.W. Inhibition of Drug Efflux Pumps in Staphylococcus Aureus: Current Status of Potentiating Existing Antibiotics. Future Microbiol. 2013, 8, 491–507. [Google Scholar] [CrossRef]
- Costa, S.; Falcão, C.; Viveiros, M.; MacHado, D.; Martins, M.; Melo-Cristino, J.; Amaral, L.; Couto, I. Exploring the Contribution of Efflux on the Resistance to Fluoroquinolones in Clinical Isolates of Staphylococcus aureus. BMC Microbiol. 2011, 11, 241. [Google Scholar] [CrossRef]
- El-Baky, R.M.A.; Sandle, T.; John, J.; Abuo-Rahma, G.E.D.A.; Hetta, H.F. A Novel Mechanism of Action of Ketoconazole: Inhibition of the Nora Efflux Pump System and Biofilm Formation in Multidrug-Resistant Staphylococcus aureus. Infect. Drug Resist. 2019, 12, 1703–1718. [Google Scholar] [CrossRef]
- Freitas, P.R.; de Araújo, A.C.J.; Barbosa, C.R.; Muniz, D.F.; Tintino, S.R.; Ribeiro-Filho, J.; Siqueira Júnior, J.P.; Filho, J.M.B.; de Sousa, G.R.; Coutinho, H.D.M. Inhibition of Efflux Pumps by Monoterpene (α-Pinene) and Impact on Staphylococcus aureus Resistance to Tetracycline and Erythromycin. Curr. Drug Metab. 2021, 22, 123–126. [Google Scholar] [CrossRef] [PubMed]
- Cappiello, F.; Loffredo, M.R.; Del Plato, C.; Cammarone, S.; Casciaro, B.; Quaglio, D.; Mangoni, M.L.; Botta, B.; Ghirga, F. The Revaluation of Plant-Derived Terpenes to Fight Antibiotic-Resistant Infections. Antibiotics 2020, 9, 325. [Google Scholar] [CrossRef] [PubMed]
- Scherf, J.R.; Barbosa dos Santos, C.R.; Sampaio de Freitas, T.; Rocha, J.E.; Macêdo, N.S.; Mascarenhas Lima, J.N.; Melo Coutinho, H.D.; Bezerra da Cunha, F.A. Effect of Terpinolene against the Resistant Staphylococcus aureus Strain, Carrier of the Efflux Pump QacC and β-Lactamase Gene, and Its Toxicity in the Drosophila Melanogaster Model. Microb. Pathog. 2020, 149, 104528. [Google Scholar] [CrossRef] [PubMed]
- De Angelis, G.; Del Giacomo, P.; Posteraro, B.; Sanguinetti, M.; Tumbarello, M. Molecular Mechanisms, Epidemiology, and Clinical Importance of β-Lactam Resistance in Enterobacteriaceae. Int. J. Mol. Sci. 2020, 21, 5090. [Google Scholar] [CrossRef]
- Papp-Wallace, K.M.; Mack, A.R.; Taracila, M.A.; Bonomo, R.A. Resistance to Novel β-Lactam–β-Lactamase Inhibitor Combinations. Infect. Dis. Clin. N. Am. 2020, 34, 773–819. [Google Scholar] [CrossRef] [PubMed]
- Tooke, C.L.; Hinchliffe, P.; Bragginton, E.C.; Colenso, C.K.; Hirvonen, V.H.A.; Takebayashi, Y.; Spencer, J. β-Lactamases and β-Lactamase Inhibitors in the 21st Century. J. Mol. Biol. 2019, 431, 3472–3500. [Google Scholar] [CrossRef]
- Adams, R. Identification of Essential Oil Components by Gas Chromatography/Quadrupole Mass Spectroscopy. Carol. Stream 2005, 16, 65–120. [Google Scholar]
- Javadpour, M.M.; Juban, M.M.; Lo, W.C.J.; Bishop, S.M.; Alberty, J.B.; Cowell, S.M.; Becker, C.L.; McLaughlin, M.L. De Novo Antimicrobial Peptides with Low Mammalian Cell Toxicity. J. Med. Chem. 1996, 39, 3107–3113. [Google Scholar] [CrossRef]
- Humphries, R.; Bobenchik, A.M.; Hindler, J.A.; Schuetz, A.N. Overview of Changes to the Clinical and Laboratory Standards Institute Performance Standards for Antimicrobial Susceptibility Testing, M100, 31st Edition. J. Clin. Microbiol. 2021, 59, e0021321. [Google Scholar] [CrossRef]
- Coutinho, H.D.M.; Costa, J.G.M.; Lima, E.O.; Falcão-Silva, V.S.; Siqueira, J.P. Enhancement of the Antibiotic Activity against a Multiresistant Escherichia coli by Mentha arvensis L. and Chlorpromazine. Chemotherapy 2008, 54, 328–330. [Google Scholar] [CrossRef] [PubMed]
- Oliveira-Tintino, C.D.d.M.; Tintino, S.R.; Muniz, D.F.; Rodrigues dos Santos Barbosa, C.; Pereira, R.L.S.; Begnini, I.M.; Rebelo, R.A.; da Silva, L.E.; Mireski, S.L.; Nasato, M.C.; et al. Chemical Synthesis, Molecular Docking and MepA Efflux Pump Inhibitory Effect by 1,8-Naphthyridines Sulfonamides. Eur. J. Pharm. Sci. 2021, 160, 105753. [Google Scholar] [CrossRef] [PubMed]
- Pal, S.; Misra, A.; Banerjee, S.; Dam, B. Adaptation of Ethidium Bromide Fluorescence Assay to Monitor Activity of Efflux Pumps in Bacterial Pure Cultures or Mixed Population from Environmental Samples. J. King Saud Univ. Sci. 2020, 32, 939–945. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | % | RT | IR 1 | IR 2 | |
|---|---|---|---|---|---|
| 1 | α-pinene | 28.38 | 3.99 | 1072 | 1071 |
| 2 | Camphene | 0.23 | 4.15 | 1092 | 1090 |
| 3 | β-pinene | 31.51 | 4.47 | 1117 | 1118 |
| 4 | α-Phellandrene | 0.23 | 4.75 | 1137 | 1140 |
| 5 | β-terpinyl acetate | 6.52 | 5.03 | 1157 | 1168 |
| 6 | β-cis-ocimene | 20.22 | 5.17 | 1166 | 1173 |
| 7 | cis-sabinene hydrate | 0.26 | 5.39 | 1182 | 1191 |
| 8 | Linalool | 0.63 | 5.65 | 1200 | 1202 |
| 9 | Terpinen-4-ol | 0.39 | 6.46 | 1242 | 1231 |
| 10 | α-Terpineol | 0.77 | 6.55 | 1246 | 1257 |
| 11 | α-Copaene | 1.06 | 8.26 | 1350 | 1351 |
| 12 | trans-Caryophyllene | 6.11 | 8.67 | 1381 | 1392 |
| 13 | α-Humulene | 0.41 | 8.98 | 1408 | 1413 |
| 14 | Alloaromadendrene | 0.18 | 9.06 | 1417 | 1423 |
| 15 | α-Amorphene | 0.14 | 9.14 | 1426 | 1437 |
| 16 | Germacrene-D | 1.08 | 9.23 | 1438 | 1444 |
| 17 | byciclogermancrene | 0.82 | 9.40 | 1457 | 1464 |
| 18 | δ-Cadinene | 0.14 | 9.59 | 1480 | 1494 |
| 19 | Nerolidol | 0.25 | 9.88 | 1511 | 1514 |
| 20 | α-Muurolol | 0.15 | 10.27 | 1545 | 1561 |
| 21 | Caryophyllene oxide | 0.11 | 10.40 | 1556 | 1565 |
| TOTAL | 99.59 | ||||
| Monoterpenes | 89.14 | ||||
| Oxygenated monoterpenes | 8.57 | ||||
| Sesquiterpenes | 10.45 | ||||
| Oxygenated sesquiterpenes | 0.51 | ||||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
da Silva, L.Y.S.; Paulo, C.L.R.; Moura, T.F.; Alves, D.S.; Pessoa, R.T.; Araújo, I.M.; de Morais Oliveira-Tintino, C.D.; Tintino, S.R.; Nonato, C.d.F.A.; da Costa, J.G.M.; et al. Antibacterial Activity of the Essential Oil of Piper tuberculatum Jacq. Fruits against Multidrug-Resistant Strains: Inhibition of Efflux Pumps and β-Lactamase. Plants 2023, 12, 2377. https://doi.org/10.3390/plants12122377
da Silva LYS, Paulo CLR, Moura TF, Alves DS, Pessoa RT, Araújo IM, de Morais Oliveira-Tintino CD, Tintino SR, Nonato CdFA, da Costa JGM, et al. Antibacterial Activity of the Essential Oil of Piper tuberculatum Jacq. Fruits against Multidrug-Resistant Strains: Inhibition of Efflux Pumps and β-Lactamase. Plants. 2023; 12(12):2377. https://doi.org/10.3390/plants12122377
Chicago/Turabian Styleda Silva, Lucas Yure Santos, Cicera Laura Roque Paulo, Talysson Felismino Moura, Daniel Sampaio Alves, Renata Torres Pessoa, Isaac Moura Araújo, Cícera Datiane de Morais Oliveira-Tintino, Saulo Relison Tintino, Carla de Fatima Alves Nonato, José Galberto Martins da Costa, and et al. 2023. "Antibacterial Activity of the Essential Oil of Piper tuberculatum Jacq. Fruits against Multidrug-Resistant Strains: Inhibition of Efflux Pumps and β-Lactamase" Plants 12, no. 12: 2377. https://doi.org/10.3390/plants12122377
APA Styleda Silva, L. Y. S., Paulo, C. L. R., Moura, T. F., Alves, D. S., Pessoa, R. T., Araújo, I. M., de Morais Oliveira-Tintino, C. D., Tintino, S. R., Nonato, C. d. F. A., da Costa, J. G. M., Ribeiro-Filho, J., Coutinho, H. D. M., Kowalska, G., Mitura, P., Bar, M., Kowalski, R., & Menezes, I. R. A. d. (2023). Antibacterial Activity of the Essential Oil of Piper tuberculatum Jacq. Fruits against Multidrug-Resistant Strains: Inhibition of Efflux Pumps and β-Lactamase. Plants, 12(12), 2377. https://doi.org/10.3390/plants12122377