
Plant-Derived Smoke and Karrikin 1 in Seed Priming and Seed Biotechnology



Abstract
:1. Introduction
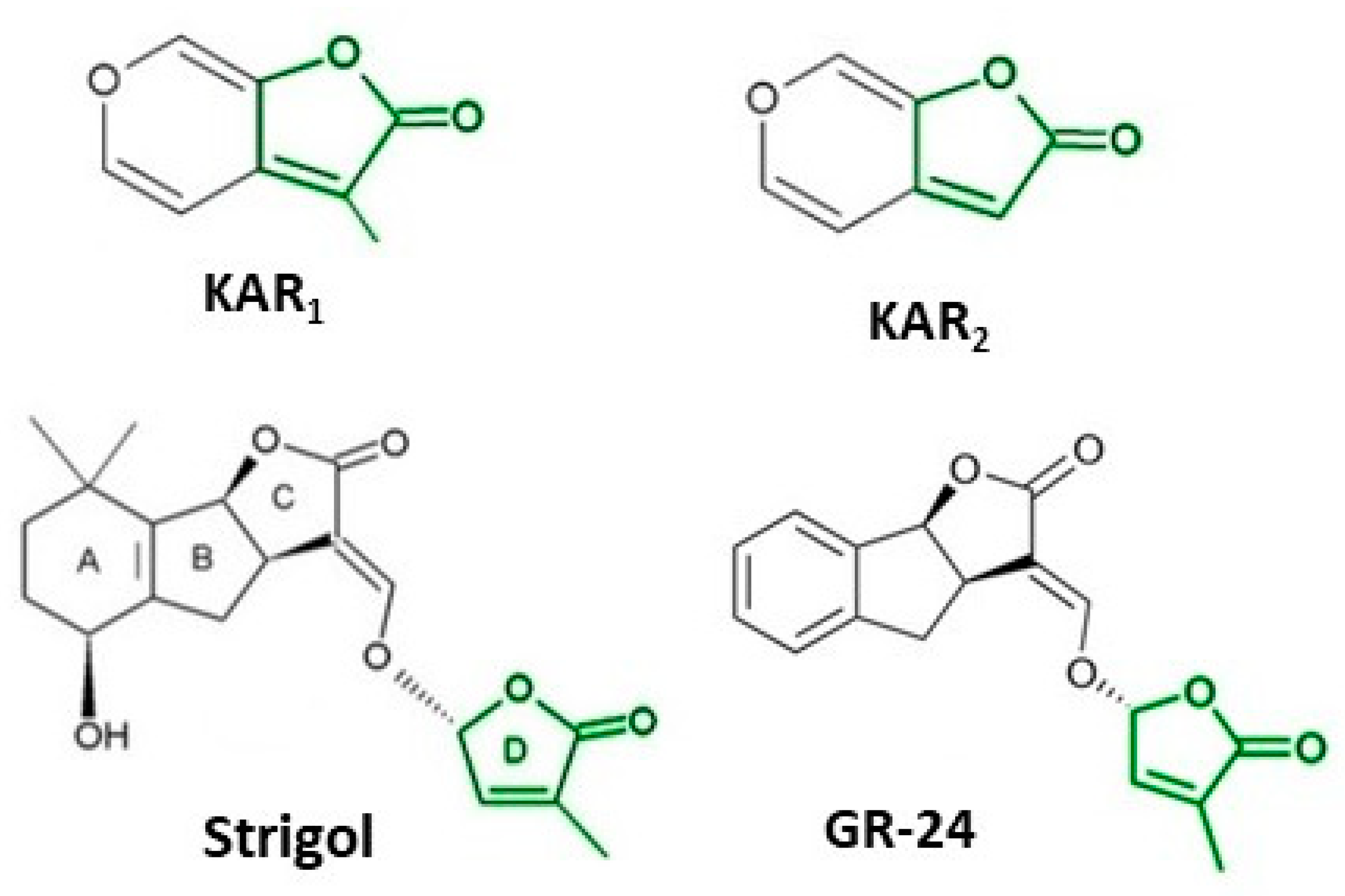
2. The Importance of Discovering the Biological Activity of Smoke and Compounds Isolated from It
3. Seed Priming with SW or KAR1 Solution
4. SW and KAR1 in Seed Biotechnology
5. Mechanism of Karrikin Signalling
6. Summing up Perspectives
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed priming for abiotic stress tolerance: An overview. Acta Physiol. Plant 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Garcia, D.; Zhao, S.; Arif, S.; Zhao, Y.; Ming, L.C.; Huang, D. Seed Priming technology as a key strategy to increase crop plant production under adverse environmental conditions. J. Agri. Horti. Res. 2022, 5, 27–46. [Google Scholar]
- Kępczyński, J. Gas-priming as a novel simple method of seed treatment with ethylene, hydrogen cyanide or nitric oxide. Acta Physiol. Plant 2021, 43, 117. [Google Scholar] [CrossRef]
- Farooq, M.; Usman, M.; Nadeem, F.; ur Rehman, H.; Wahid, A.; Basra, S.M.; Siddique, K.H. Seed priming in field crops: Potential benefits, adoption and challenges. Crop Pasture Sci. 2019, 70, 731–771. [Google Scholar] [CrossRef]
- Kulkarni, M.G.; Light, M.E.; Van Staden, J. Plant-derived smoke: Old technology with possibilities for economic applications in agriculture and horticulture. S. Afr. J. Bot. 2011, 77, 972–979. [Google Scholar] [CrossRef] [Green Version]
- De Lange, J.H.; Boucher, C. Auto ecological studies on Audinia capitata (Bruniaceae). I. Plant-derived smoke as a germination cue. S. Afr. J. Bot. 1990, 56, 700–703. [Google Scholar] [CrossRef] [Green Version]
- Dixon, K.W.; Merritt, D.J.; Flematti, G.R.; Ghisalberti, E.L. Karrikinolide–a phytoreactive compound derived from smoke with applications in horticulture, ecological restoration and agriculture. Acta Hort. 2009, 813, 155–170. [Google Scholar] [CrossRef]
- Nelson, D.C.; Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Regulation of seed germination and seedling growth by chemical signals from burning vegetation. Ann. Rev. Plant Biol. 2012, 63, 107–130. [Google Scholar] [CrossRef] [Green Version]
- Kępczyński, J. Induction of agricultural weed seed germination by smoke and smoke-derived karrikin (KAR 1), with a particular reference to Avena fatua L. Acta Physiol. Plant 2018, 40, 87. [Google Scholar] [CrossRef] [Green Version]
- Kępczyński, J. Progress in utilizing plant-derived smoke water and smoke-derived KAR1 in plants tissue culture. Plant Cell Tiss. Organ Cult. 2020, 140, 271–278. [Google Scholar] [CrossRef]
- Banerjee, A.; Tripathi, D.K.; Roychoudhury, A. The Karrikin ‘calisthenics’: Can compounds derived from smoke help in stress tolerance? Physiol. Plant 2019, 165, 290–302. [Google Scholar] [CrossRef]
- Soós, V.; Badics, E.; Incze, N.; Balázs, E. Fire-borne life: A brief review of smoke-induced germination. Nat. Prod. Commun. 2019, 14, 1–5. [Google Scholar] [CrossRef]
- Brown, N.A.C.; Van Staden, J. Smoke as a germination cue: A review. Plant Growth Regul. 1997, 22, 115–124. [Google Scholar] [CrossRef]
- Light, M.E.; van Staden, J. The potential of smoke in seed technology. S. Afr. J. Bot. 2004, 70, 97–101. [Google Scholar] [CrossRef] [Green Version]
- Van Staden, J.; Jager, A.K.; Light, M.E.; Burger, B.V. Isolation of the major germination cue from plant-derived smoke. S. Afr. J. B 2004, 70, 654–659. [Google Scholar] [CrossRef] [Green Version]
- Flematti, G.R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. A compound from smoke that promotes seed germination. Science 2004, 305, 977. [Google Scholar] [CrossRef]
- Nelson, D.C.; Riseborough, J.A.; Flematti, G.R.; Stevens, J.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Karrikins discovered in smoke trigger Arabidopsis seed germination by a mechanism requiring gibberellic acid synthesis and light. Plant Physiol. 2009, 149, 863–873. [Google Scholar] [CrossRef] [Green Version]
- Nelson, D.C.; Scaffidi, A.; Dun, E.A.; Waters, M.T.; Flematti, G.R.; Dixon, K.W.; Beveridge, C.A.; Ghisalberti, E.L.; Smith, S.M. F-box protein MAX2 has dual roles in karrikin and strigolactone signaling in Arabidopsis thaliana. Proc. Nat. Acad. Sci. USA 2011, 108, 8897–8902. [Google Scholar] [CrossRef] [Green Version]
- Kępczyński, J.; Cembrowska, D.; Van Staden, J. Releasing primary dormancy in Avena fatua L. caryopses by smoke-derived butanolide. Plant Growth. Regul. 2010, 62, 85–91. [Google Scholar] [CrossRef] [Green Version]
- Goddard-Borger, E.D.; Ghisalberti, E.L.; Stick, R.V. Synthesis of the Germination Stimulant 3-Methyl-2H-furo [2, 3-c] pyran-2-one and Analogous Compounds from Carbohydrates. Eur. J. Org. Chem. 2007, 23, 3925–3934. [Google Scholar] [CrossRef]
- Matsuo, K.; Shindo, M. Efficient synthesis of karrikinolide via Cu (II)-catalyzed lactonization. Tetrahedron 2011, 67, 971–975. [Google Scholar] [CrossRef]
- Sun, K.; Chen, Y.; Wagerle, T.; Linnstaedt, D.; Currie, M.; Chmura, P.; Xu, M. Synthesis of butenolides as seed germination stimulants. Tetrahedron Lett. 2008, 49, 2922–2925. [Google Scholar] [CrossRef]
- Nagase, R.; Katayama, M.; Mura, H.; Matsuo, N.; Tanabe, Y. Synthesis of the seed germination stimulant 3-methyl-2H-furo [2, 3-c] pyran-2-ones utilizing direct and regioselective Ti-crossed aldol addition. Tetrahedron Lett. 2008, 49, 4509–4512. [Google Scholar] [CrossRef]
- Flematti, G.R.; Gavin, R.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. Synthesis of the seed germination stimulant 3-methyl-2H-furo[2, 3-c]pyran-2-one. Tetrahedron Lett. 2005, 46, 5719–5721. [Google Scholar] [CrossRef]
- Flematti, G.R.; Waters, M.T.; Scaffidi, A.; Merritt, D.J.; Ghisalberti, E.L.; Dixon, K.W.; Smith, S.M. Karrikin and cyanohydrin smoke signals provide clues to new endogenous plant signaling compounds. Mol. Plant. 2013, 6, 29–37. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Keeley, J.E.; Fotheringham, C.J. Trace gas emissions in smoke-induced germination. Science 1997, 276, 1248–1250. [Google Scholar] [CrossRef] [Green Version]
- Flematti, G.R.; Goddard-Borger, E.D.; Merritt, D.J.; Ghisalberti, E.L.; Dixon, K.W.; Trengove, R.D. Preparation of 2 H-furo [2,3-c] pyran-2-one derivatives and evaluation of their germination-promoting activity. J. Agr. Food Chem. 2007, 55, 2189–2194. [Google Scholar] [CrossRef]
- Light, M.E.; Burger, B.V.; Staerk, D.; Kohout, L.; van Staden, J. Butenolides from plant-derived smoke: Natural plant-growth regulators with antagonistic actions on seed germination. J. Nat. Prod. 2010, 73, 267–269. [Google Scholar] [CrossRef] [Green Version]
- Flematti, G.R.; Merritt, D.J.; Piggott, M.J.; Trengove, R.D.; Smith, S.M.; Dixon, K.W.; Ghisalberti, E.L. Burning vegetation produces cyanohydrins that liberate cyanide and stimulate seed germination. Nat. Commun. 2011, 2, 360. [Google Scholar] [CrossRef] [Green Version]
- Kamran, M.; Khan, A.L.; Ali, L.; Hussain, J.; Waqas, M.; Al-Harrasi, A.; Imran, Q.M.; Kim, Y.H.; Kang, S.M.; Yun, B.W.; et al. Hydroquinone: A novel bioactive compopund from plant-derived smoke can cue seed germination of lettuce. Front. Chem. 2017, 5, 30. [Google Scholar] [CrossRef] [Green Version]
- Burger, B.V.; Pošta, M.; Light, M.E.; Kulkarni, M.G.; Viviers, M.Z.; Van Staden, J. More butenolides from plant-derived smoke with germination inhibitory activity against karrikinolide. S. Afr. J. Bot. 2018, 115, 256–263. [Google Scholar] [CrossRef]
- Light, M.E.; Gardner, M.J.; Jager, A.K.; van Staden, J. Dual regulation of seed germination by smoke solutions. Plant Growth Regul. 2002, 37, 135–141. [Google Scholar] [CrossRef]
- Khatoon, A.; Rehman, S.F.; Aslam, M.M.; Jamil, M.; Komatsu, S. Plant-derived smoke affects biochemical mechanism on plant growth and seed germination. Inter. J. Mol. Sci. 2020, 21, 7760. [Google Scholar] [CrossRef]
- Moyo, M.; Amoo, S.O.; Van Staden, J. Seed priming with smoke water and karrikin improves germination and seedling vigor of Brassica napus under varying environmental conditions. Plant Growth Regul. 2022, 97, 315–326. [Google Scholar] [CrossRef]
- Demir, I.; Ozden, E.; Yıldırım, K.C.; Sahin, O.; Van Staden, J. Priming with smoke-derived karrikinolide enhances germination and transplant quality of immature and mature pepper seed lots. S. Afr. J. Bot. 2018, 115, 264–268. [Google Scholar] [CrossRef]
- Masondo, N.A.; Kulkarni, M.G.; Finnie, J.F.; Van Staden, J. Influence of biostimulants-seed-priming on Ceratotheca triloba germination and seedling growth under low temperatures, low osmotic potential and salinity stress. Ecotoxicol. Environ. Saf. 2018, 147, 43–48. [Google Scholar] [CrossRef] [PubMed]
- Sardar, R.; Ahmed, S.; Yasin, N.A. Seed priming with karrikinolide improves growth and physiochemical features of Coriandrum sativum under cadmium stress. Environ. Adv. 2021, 5, 100082. [Google Scholar] [CrossRef]
- Mavi, K.; Light, M.E.; Demir, I.; Van Staden, J.; Yasar, F. Positive effect of smoke-derived butenolide priming on melon seedling emergence and growth. N. Z. J. Crop Hort. Sci. 2010, 38, 147–155. [Google Scholar] [CrossRef]
- Akeel, A.; Khan, M.M.A.; Jaleel, H.; Uddin, M. Smoke-saturated Water and Karrikinolide Modulate Germination, Growth, Photosynthesis and Nutritional Values of Carrot (Daucus carota L.). J. Plant Growth Regul. 2019, 38, 1387–1401. [Google Scholar] [CrossRef]
- Ghebrehiwot, H.M.; Kulkarni, M.G.; Kirkman, K.P.; Van Staden, J. Smoke-water and a smoke-isolated butenolide improve germination and seedling vigour of Eragrostis Tef (Zucc.) Trotter under high temperature and low osmotic potential. J. Agron. Crop Sci. 2008, 194, 270–277. [Google Scholar] [CrossRef]
- Jain, N.; Van Staden, J. The potential of the smoke-derived compound 3-methyl-2H-furo[2, 3-c]pyran-2-one as a priming agent for tomato seeds. Seed Sci. Res. 2007, 17, 175–181. [Google Scholar] [CrossRef]
- Jamil, M.; Kanwal, M.A.; Aslam, M.M.; Khan, S.; Tu, J.; Réhman, S.U. Effect of plant-derived smoke priming on physiological and biochemical characteristics of rice under salt stress condition. Austr. J. Crop Sci. 2014, 8, 159–170. [Google Scholar]
- Abdollahi, M.R.; Mehrshad, B.; Moosavi, S.S. Effect of method of seed treatment with plant derived smoke solutions on germination and seedling growth of milk thistle (Silybum marianum L.). Seed Sci. Technol. 2011, 39, 225–229. [Google Scholar] [CrossRef]
- Commander, L.E.; Merritt, D.J.; Rokich, D.P.; Flematti, G.R.; Dixon, K.W. Seed germination of Solanum spp. (Solanaceae) for use in rehabilitation and commercial industries. Aust. J. Bot. 2008, 56, 333–341. [Google Scholar] [CrossRef]
- Baxter, B.J.M.; Van Staden, J. Plant-derived smoke: An effective seed pre-treatment. Plant Growth Regul. 1994, 14, 279–282. [Google Scholar] [CrossRef]
- Aslam, M.M.; Jamil, M.; Khatoon, A.; Hendawy, S.E.; Al-Suhaibani, N.A.; Malook, I.; Rehman, S.U. Physiological and biochemical responses of maize (Zea mays L.) to plant derived smoke solution. Pak. J. Bot. 2017, 49, 435–443. Available online: https://w.w.w.researchgate.net/publications/315808314 (accessed on 31 May 2023).
- Van Staden, J.; Sparg, S.G.; Kulkarni, M.G.; Light, M.E. Post-germination effects of the smoke-derived compound 3-methyl-2H-furo [2,3-c] pyran-2-one, and its potential as a preconditioning agent. Field Crops Res. 2006, 98, 98–105. [Google Scholar] [CrossRef]
- Abu, Y.; Romo, J.T.; Bai, Y.; Coulman, B. Priming seeds in aqueous smoke solution to improve seed germination and biomass production of perennial forage species. Can. J. Plant Sci. 2016, 96, 551–563. [Google Scholar] [CrossRef]
- Keeley, J.E. Smoke-induced flowering in the fire-lily Cyrtanthus ventricosus. S. Afr. J. Bot. 1993, 59, 638. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, M.G.; Ascough, G.D.; Van Staden, J. Improved flowering of a South African Watsonia with smoke treatment. S. Afr. J. Bot. 2007, 73, 298. [Google Scholar] [CrossRef]
- Papenfus, H.B.; Kumari, A.; Kulkarni, M.G.; Finnie, J.F.; Van Staden, J. Smoke-water enhances in vitro pollen germination and tube elongation of three species of Amaryllidaceae. S. Afr. J. Bot. 2014, 90, 87–92. [Google Scholar] [CrossRef] [Green Version]
- Kumari, A.; Papenfus, H.B.; Kulkrni, M.G.; Pošta, M.; van Staden, J. Efect of smoke derivatives on in vitro pollen germination and pollen tube elongation of species from diferent plant families. Plant Biol. 2015, 17, 825–830. [Google Scholar] [CrossRef] [PubMed]
- Ma, G.-H.; Bunn, E.; Dixon, K.; Flematti, G. Comparative enhancement of germination and vigor in seed and somatic embryos by the smoke chemical 3-methyl-2H-furo [2,3-C] pyran-2-one in Baloskion tetraphyllum (Restionaceae). Vitr. Cell Dev. Biol. 2006, 42, 305–308. [Google Scholar] [CrossRef]
- Senaratna, T.; Dixon, K.; Bunn, E.; Touchell, D. Smoke-saturated water promotes somatic embryogenesis in geranium. Plant Growth Regul. 1999, 28, 95–99. [Google Scholar] [CrossRef]
- Malabadi, R.B.; Nataraja, K. Smoke-saturated water infuences somatic embryogenesis using vegetative shoot apices of mature trees of Pinus wallichiana AB Jacks. J Plant Sci. 2007, 2, 45–53. [Google Scholar] [CrossRef] [Green Version]
- Ghazanfari, P.; Abdollahi, M.R.; Moieni, A.; Moosavi, S.S. Efect of plant-derived smoke extract on in vitro plantlet regeneration from rapeseed (Brassica napus L. cv. Topas) microspore-derived embryos. Int. J. Plant Prod. 2012, 6, 1735–6814. [Google Scholar] [CrossRef]
- Abdollahi, M.R.; Ghazanfari, P.; Corral-Martínez, P.; Moieni, A.; Seguí-Simarro, J.M. Enhancing secondary embryogenesis in Brassica napus by selecting hypocotyl- derived embryos and using plant-derived smoke extract in culture medium. Plant Cell Tissue Organ Cult. 2012, 110, 307–315. [Google Scholar] [CrossRef]
- Papenfus, H.B.; Naidoo, D.; Pošta, M.; Finnie, J.F.; Van Staden, J. The efects of smoke derivatives on in vitro seed germination and development of the leopard orchid Ansellia africana. Plant Biol. 2016, 18, 289–294. [Google Scholar] [CrossRef]
- Malabadi, R.B.; da Silva, J.A.T.; Mulgund, G.S. Smoke-saturated water influences in vitro seed germination of Vanda parviflora Lindl. Seed Sci. Biotechnol. 2008, 2, 65–69. [Google Scholar]
- Malabadi, R.B.; Vijaykumar, S.; da Silva, J.A.T.; Mulgund, G.S.; Nataraja, K. In vitro seed germination of an epiphytic orchid Xenikophyton smeeanum (Reichb. f.) by using smoke-saturated-water as a natural growth promoter. Int. J. Biol. Technol. 2011, 2, 35–40. [Google Scholar]
- Espinosa-Leal, C.A.; Garcia-Lara, S. In vitro germination and initial seedling development of krantz aloe by smoke-saturated water and seed imbibition. Hort Technol. 2020, 30, 619–623. [Google Scholar] [CrossRef]
- Monthony, A.S.; Baethke, K.; Erland, L.A.; Murch, S.J. Tools for conservation of Balsamorhiza deltoidea and Balsamorhiza sagittata: Karrikin and thidiazuron-induced growth. Vitr. Cell. Develop. Biol.-Plant. 2020, 56, 398–406. [Google Scholar] [CrossRef]
- Waters, M.T.; Nelson, D.C. Karrikin perception and signalling. New Phytol. 2023, 237, 1525–1541. [Google Scholar] [CrossRef]
- Morffy, N.; Faure, L.; Nelson, D. Smoke and hormone mirrors:action and evolution of Karrikin and strigolactone signaling. Trends Genet. 2016, 3, 176–188. [Google Scholar] [CrossRef] [PubMed]
- Bürger, M.; Mashiguchi, K.; Lee, H.J.; Nakano, M.; Takemoto, K.; Seto, Y.; Yamaguchi, S.; Chory, J. Structural. basis of Karrikin and non-natural strigolactone perception in Physcomitrella patens. Cell Rep. 2019, 26, 855–865. [Google Scholar] [CrossRef] [Green Version]
- Blázquez, M.A.; Nelson, D.C.; Weijers, D. Evolution of plant hormone response pathways. Annu. Rev. Plant. Biol. 2020, 71, 327–353. [Google Scholar] [CrossRef] [Green Version]
- Kępczyński, J. Induction of dormancy release in agricultural weed seeds by plant-derived smoke and smoke-derived Karrikin 1(KAR1). A relationship with plant hormones. In Strigolactones, Karrikins and Alkamides in Plants; Soumya, M., Aftab, T., Eds.; Publisher Taylor & Francis: Boca Raton, FL, USA, 2023; pp. 225–240. [Google Scholar]




{kind=link}
{kind=link}
{kind=link}
| Substrates | Structure | References |
|---|---|---|
| D-xylose |  | [20,21] |
| Ethyl-4-methyl-2-oxo-2,5-dihydro-furan-3-carboxylate |  | [22] |
| Furfurylmethanol |  | [23] |
| Pyromeconic acid |  | [24] |
| Compound | Plant Species | References | |
|---|---|---|---|
| Nitrogen dioxide | Emmenanthe penduliflora | ↑ | [26] |
| KAR1 (3-methyl-2H-furo [2, 3-c]pyran-2-one) | Lactuca sativa | ↑ | [15,16] |
| KAR2, KAR3 | Arabidopsis thaliana | ↑ | [17] |
| KAR4 | Lactuca sativa | ↑ | [27] |
| 3,4,5-trimethylfuran-2(5H)-one (trimethylbutenolide; TMB) | Lactuca sativa | ↓ | [28] |
| Glyceronitrile | Angiozanthos manglesi | ↑ | [29] |
| Hydroquinone | Lactuca sativa | ↑ | [30] |
| 5,5-dimethylfuran-2(5H) -one | Lactuca sativa | ↓ | [31] |
| (5RS) -5-ethylfuran-2(5H) -one | Lactuca sativa | ↓ | [31] |
| Plant Species | SW, Dilution KAR1, M or μg/L−1 | Methods of Application | Beneficial Effect on: | References |
|---|---|---|---|---|
| Brassica napus L. | 1:1000 10−8 | imbibition at 25 °C for 12 h, blotting dry | final percentage, time of germination, at heat stress at 40 °C, CAT activity after 7 d | [34] |
| Brassica napus L. | 1:250 10−7 | imbibition at 25 °C for 12 h, blotting dry | growth of seedlings at 25 °C after 7 d | [34] |
| Capsicum annuum L. | 10−7 | imbibition at 25 °C, for 40 h, rinsing, drying at room temp. for 24 h | germination of immature seeds at 25 °C after 10 d, seedling emergence from immature seeds, fresh weight of seedlings from immature and mature seeds at 23 °C after 20 d | [35] |
| Ceratotheca triloba (Bernh.) Hook. | 1:500 10−6 | imbibition at 25 °C, for 48 h | germination, vigor, seedling growth, at 10/15 °C, 10 °C after 15 d, germination under osmotic stress at 25 °C after 25 d | [36] |
| Coriandrum sativum L. | 10−6 | priming in solution at room temperature for 15 h | seed germination, seedling growth, chlorophyll a, b, carotenoids and proline contents, membrane stability, leaf osmotic potential, photosynthesis rate, the activity of SOD, POD and CAT at 25 °C after 15 d under cadmium stress | [37] |
| Cucumis melo L. | 10−7 | imbibition at 25 °C for 21 h, rinsing, surface drying | seedling emergence at 20° and 25 °C, fresh and dry weight of seedling after 24 d | [38] |
| Daucus carota L. | 51.6 μg/L 1.5 μg/L | soaking for 12 h, rinsing, drying | germination in soil, seedling growth at environmental conditions, photosynthesis, ascorbic acid content after 120 d | [39] |
| Eragrostis tef (Zucc.) Trotter | 1:500 10−8 | imbibition for 48 h at 25 °C | vigor at 20°to 40 °C, seedling length and vigor index, germination at 25–40 °C, at 25 °C under osmotic stress | [40] |
| Lycopersicon esculentum Mill. | 10−7 | soaking at 23 °C for 24 h, drying up to the initial weight | rate of germination, vigor seedling in water at 10°–35 °C, in salt solutions or PEG solutions at 23 °C after seven days | [41] |
| Oryza sativa L. | 1:1000 1:500 | primed in solutens for 24 h, air drying at room temperature | seed germination up to 3 d under salt stress simulated by NaCl at 30 °C, chlorophyll, carotenoids, K+, Ca + contents | [42] |
| Silybum marianum L. Gaertn. Solanum centrale J.M.Black, S.dioicum W.Fitzg, S. orbiculatum Dunal ex Poir | 1:250 1:10 0.67μM | soaking 1 h at room temperature, drying, soaking for 24 h, rinsing | speed of germination, vigor, at 25 °C index seedling length after 14 d germination at 26/13 °C, 33/18 °C on water agar, vigor index seedling length after 18 d | [43] [44] |
| Themeda triandra Forssk. | SW | soaking 1 h at room temperature, drying, and stored at 25 for 3 to 21 days | germination at 25 °C | [45] |
| Zea mays L. | 1:500 | soaking 6–18 h | seedling growth at 28 °C after 8 d, chlorophyll and carotenoids contents | [46] |
| Zea mays L. | 1:500 10−7 | soaking 1 h, surface drying | seedling growth in the soil after 30 d in greenhouse conditions | [47] |
| Process | Plant Species | SW | KAR1 | References |
|---|---|---|---|---|
| Somatic embryogenesis Efficiency | Baloskion tetraphyllum Pelargonium hortorum Pinus wallichiana | ND + + | + ND ND | [53] [54] [55] |
| Somatic embryo germination | Baloskion tetraphyllum Pinus wallichiana | ND + | + ND | [53] [55] |
| Conversion to plantlets | Baloskion tetraphyllum Brassica napus Pinus wallichiana | ND + + | + ND ND | [53] [56] [55] |
| Secondary embryogenesis | Brassica napus | + | ND | [57] |
| Seed germination | Ansellia africana Baloskion tetraphyllum Vanda parviflora Xenikophyton smeeanum | + ND + + | − + ND ND | [58] [53] [59] [60] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kępczyński, J.; Kępczyńska, E. Plant-Derived Smoke and Karrikin 1 in Seed Priming and Seed Biotechnology. Plants 2023, 12, 2378. https://doi.org/10.3390/plants12122378
Kępczyński J, Kępczyńska E. Plant-Derived Smoke and Karrikin 1 in Seed Priming and Seed Biotechnology. Plants. 2023; 12(12):2378. https://doi.org/10.3390/plants12122378
Chicago/Turabian StyleKępczyński, Jan, and Ewa Kępczyńska. 2023. "Plant-Derived Smoke and Karrikin 1 in Seed Priming and Seed Biotechnology" Plants 12, no. 12: 2378. https://doi.org/10.3390/plants12122378
APA StyleKępczyński, J., & Kępczyńska, E. (2023). Plant-Derived Smoke and Karrikin 1 in Seed Priming and Seed Biotechnology. Plants, 12(12), 2378. https://doi.org/10.3390/plants12122378