
Water Shortage Affects Vegetative and Reproductive Stages of Common Bean (Phaseolus vulgaris) Chilean Landraces, Differentially Impacting Grain Yield Components


Abstract
:1. Introduction
2. Results
2.1. Water Stress and Temperature Effect
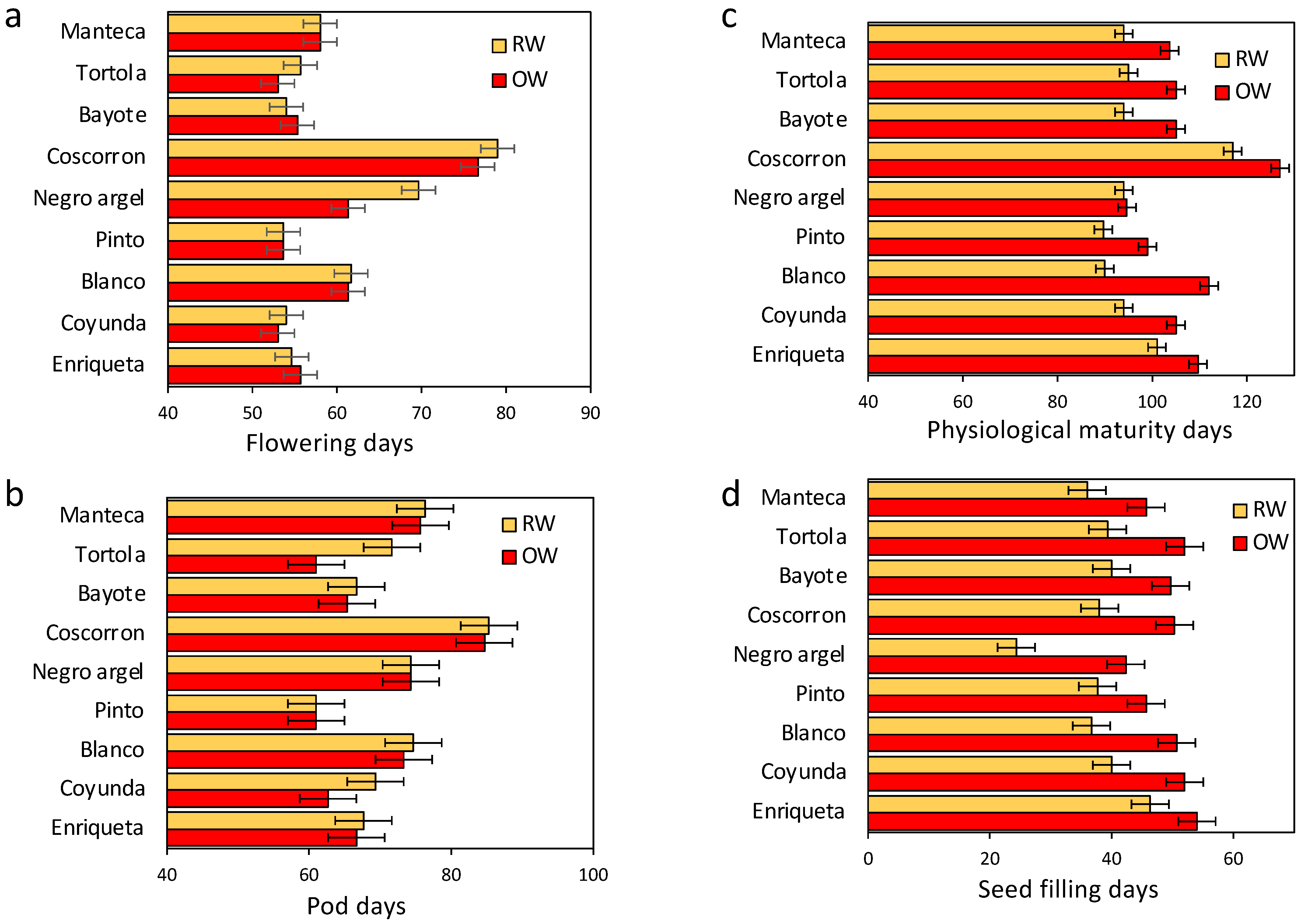
2.2. Phenological Traits
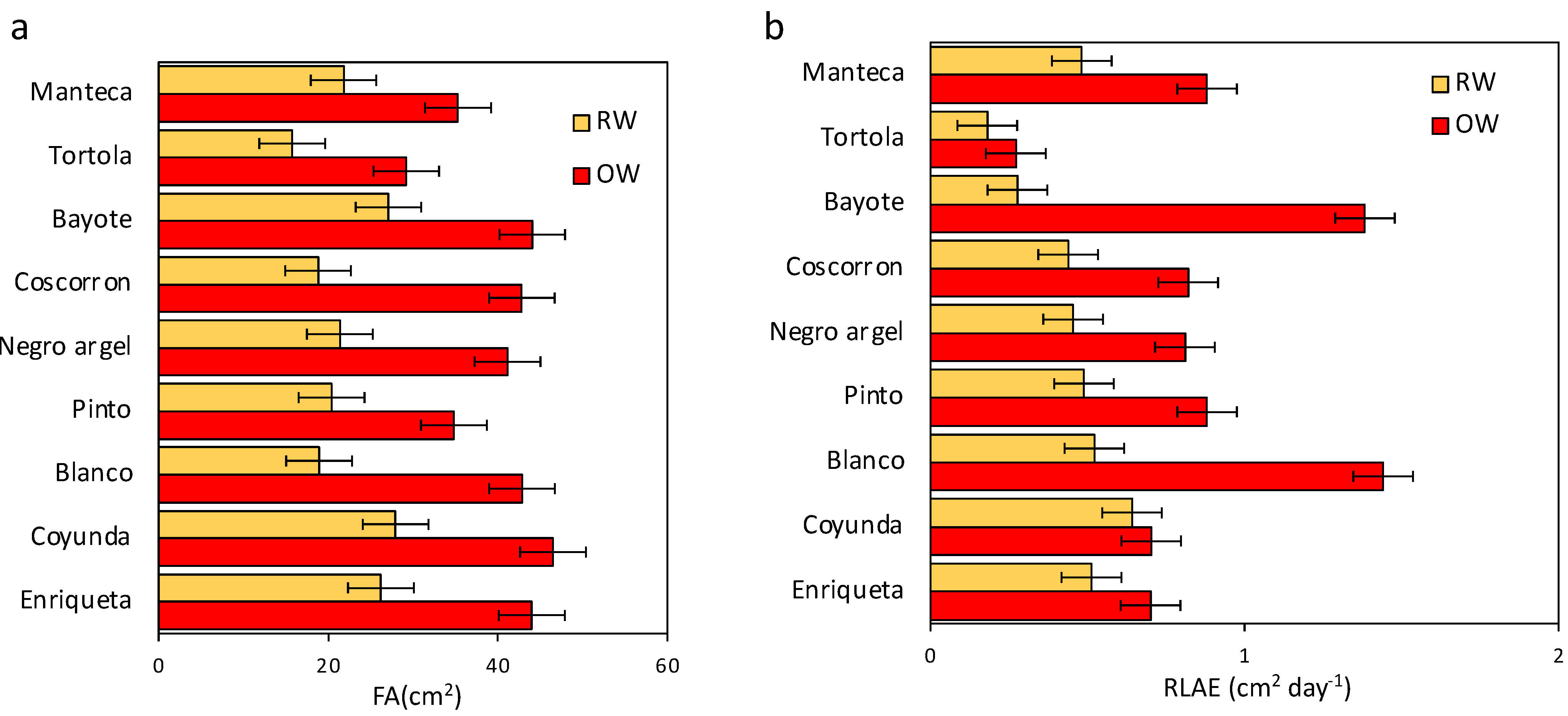
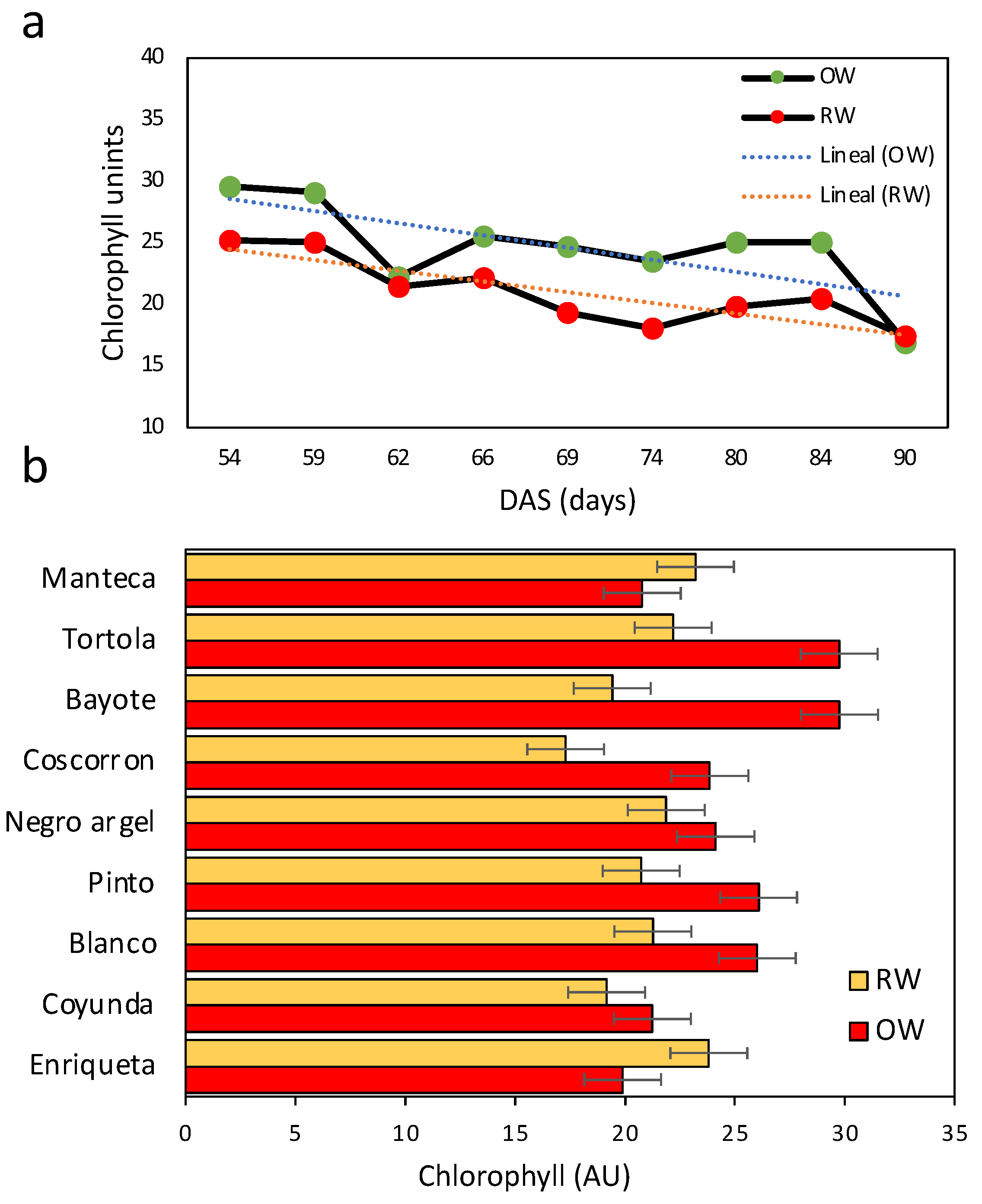
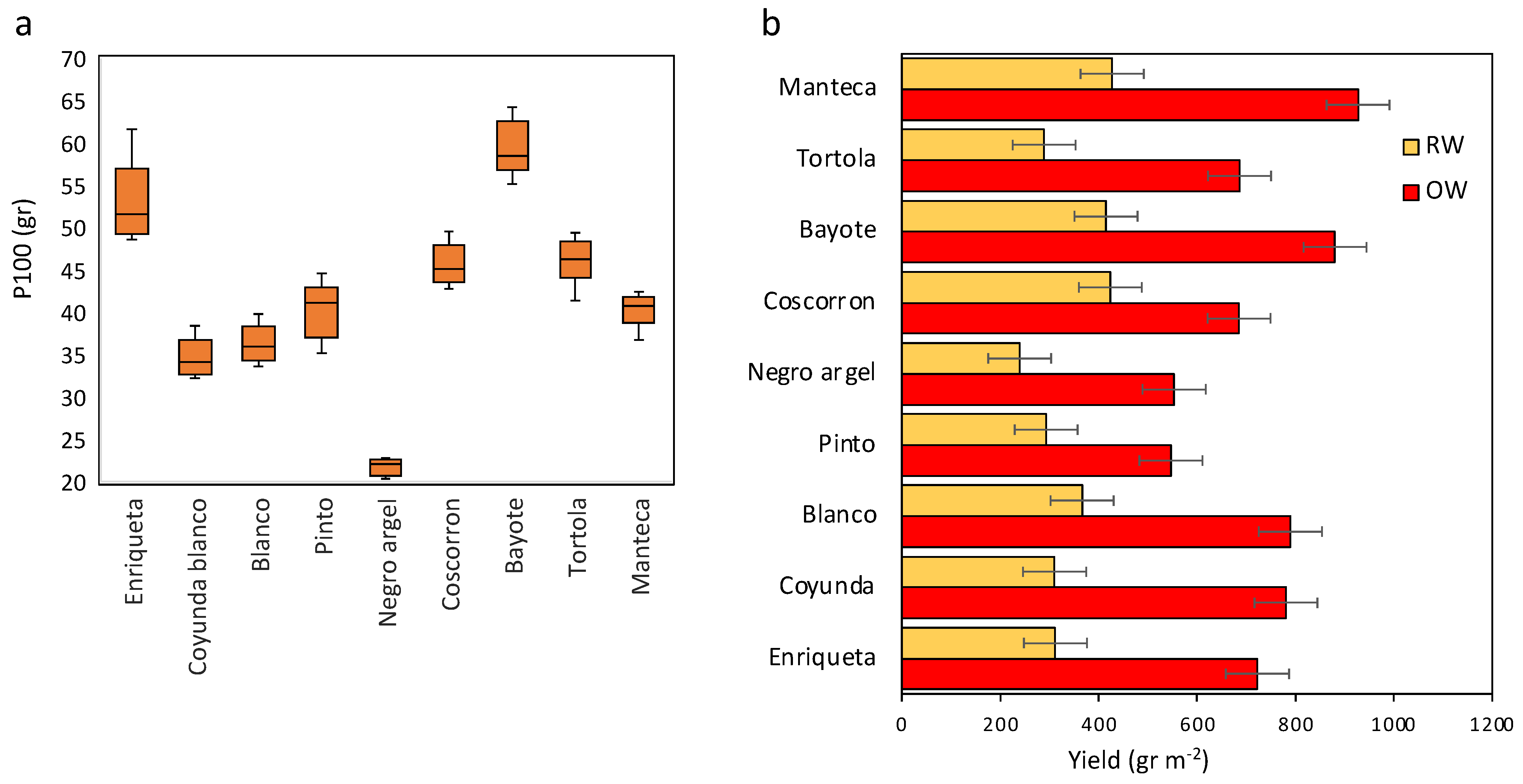
2.3. Growth Traits and Chlorophyll Concentration
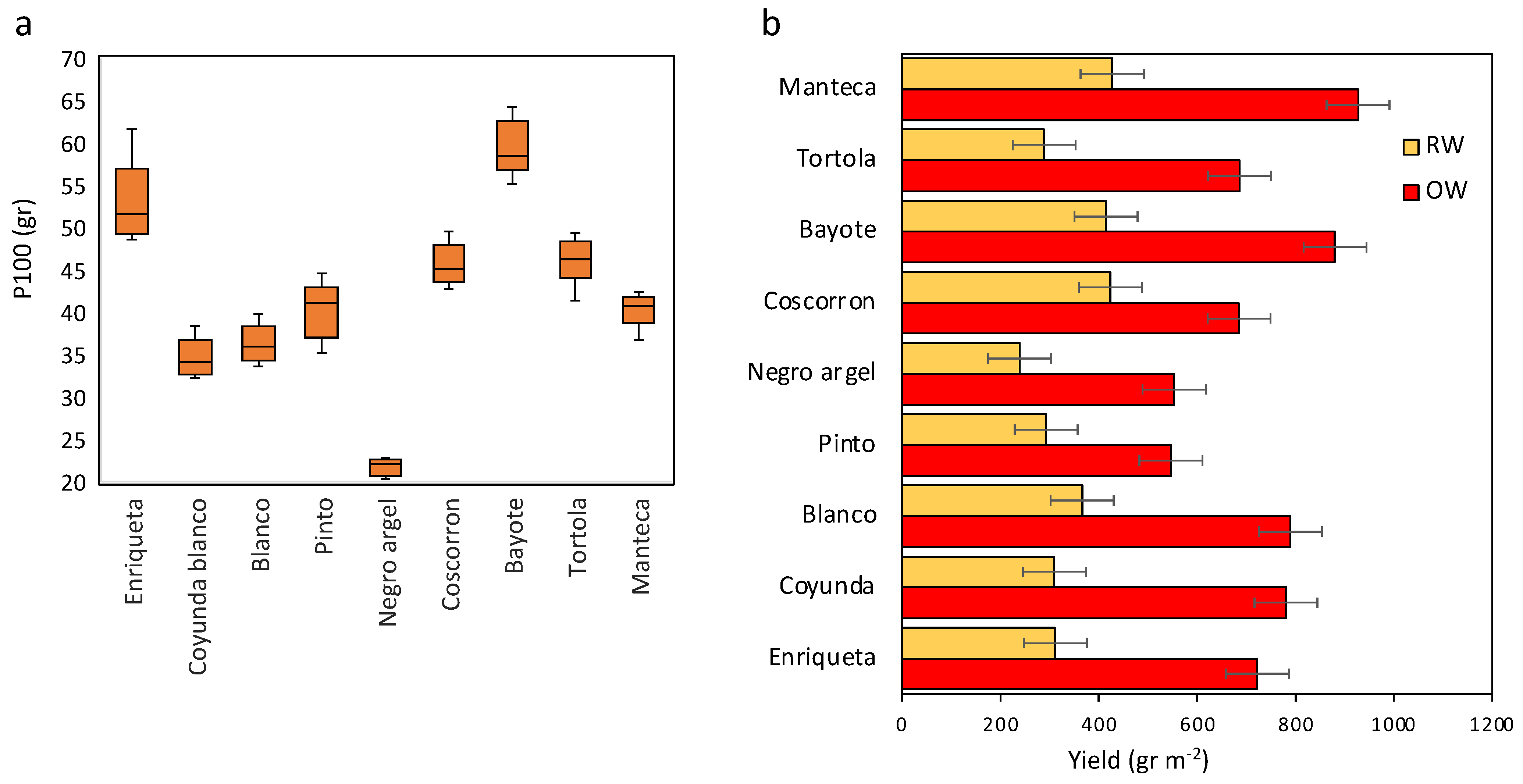
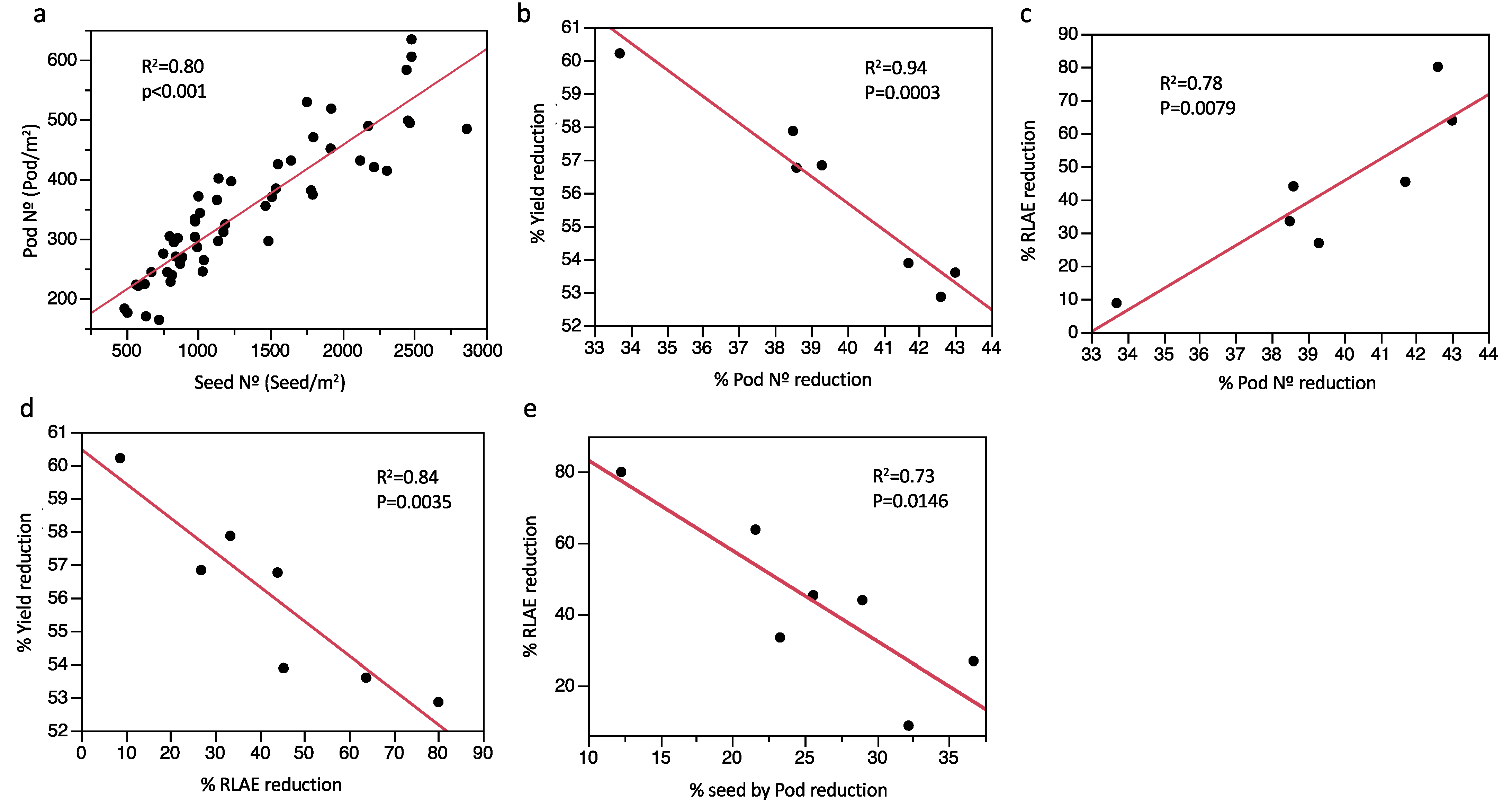
2.4. Yield Traits and % Reduction Rate Relationships based on the Effect of Water Treatment and Drought Tolerance Indices
3. Discussion
4. Materials and Methods
4.1. Plant Material and Experiment
4.2. Soil Water Content
4.3. Data Collection
4.4. Statistical Analysis
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Stagnari, F.; Maggio, A.; Galieni, A.; Pisante, M. Multiple benefits of legumes for agriculture sustainability: An overview. Chem. Biol. Technol. Agric. 2017, 4, 2. [Google Scholar] [CrossRef] [Green Version]
- Araújo, S.; Beebe, S.; Crespi, M.; Delbreil, B.; González, E.M.; Gruber, V.; Lejeune-Henaut, I.; Link, W.; Monteros, M.J.; Prats, E.; et al. Abiotic Stress Responses in Legumes: Strategies Used to Cope with Environmental Challenges. Crit. Rev. Plant Sci. 2015, 34, 237–280. [Google Scholar] [CrossRef]
- Myers, J.R.; Kmiecik, K. Common Bean: Economic Importance and Relevance to Biological Science Research. In Compendium of Plant Genomes; Springer: Berlin/Heidelberg, Germany, 2017; pp. 1–20. [Google Scholar] [CrossRef]
- Keller, B.; Ariza-Suarez, D.; de La Hoz, J.; Aparicio, J.S.; Portilla-Benavides, A.E.; Buendia, H.F.; Mayor, V.M.; Studer, B.; Raatz, B. Genomic Prediction of Agronomic Traits in Common Bean (Phaseolus vulgaris L.) Under Environmental Stress. Front. Plant Sci. 2020, 11, 1001. [Google Scholar] [CrossRef]
- Aschmann, H. Distribution and Peculiarity of Mediterranean Ecosystems. In Mediterranean Type Ecosystems; di Castri, F., Mooney, H.A., Eds.; Springer: Berlin/Heidelberg, Germany, 1973; Volume 7, pp. 11–19. [Google Scholar] [CrossRef]
- Nadeem, M.A.; Gündoğdu, M.; Ercişli, S.; Karaköy, T.; Saracoğlu, O.; Habyarimana, E.; Lin, X.; Hatipoğlu, R.; Nawaz, M.A.; Sameeullah, M.; et al. Uncovering Phenotypic Diversity and DArTseq Marker Loci Associated with Antioxidant Activity in Common Bean. Genes 2020, 11, 36. [Google Scholar] [CrossRef] [Green Version]
- Giorgi, F.; Lionello, P. Climate change projections for the Mediterranean region. Glob. Planet. Change 2008, 63, 90–104. [Google Scholar] [CrossRef]
- Garreaud, R.D.; Boisier, J.P.; Rondanelli, R.; Montecinos, A.; Sepúlveda, H.H.; Veloso-Aguila, D. The Central Chile Mega Drought (2010–2018): A climate dynamics perspective. Int. J. Clim. 2019, 40, 421–439. [Google Scholar] [CrossRef]
- Asfaw, A.; Blair, M.W.; Struik, P.C. Multienvironment Quantitative Trait Loci Analysis for Photosynthate Acquisition, Accumulation, and Remobilization Traits in Common Bean Under Drought Stress. G3 Genes|Genomes|Genet. 2012, 2, 579–595. [Google Scholar] [CrossRef] [Green Version]
- Diaz, L.M.; Ricaurte, J.; Tovar, E.; Cajiao, C.; Terán, H.; Grajales, M.; Polania, J.; Rao, I.; Beebe, S.; Raatz, B. QTL analyses for tolerance to abiotic stresses in a common bean (Phaseolus vulgaris L.) population. PLoS ONE 2018, 13, e0202342. [Google Scholar] [CrossRef]
- Beebe, S.E.; Idupulapati, M.R.; Blair, M.W.; Acosta-Gallegos, J.A. Phenotyping common beans for adaptation to drought. Front. Physiol. 2013, 4, 1–20. [Google Scholar] [CrossRef] [Green Version]
- Del Pozo, A.; Brunel-Saldias, N.; Engler, A.; Ortega-Farias, S.; Acevedo-Opazo, C.; Lobos, G.A.; Jara-Rojas, R.; Molina-Montenegro, M.A. Climate Change Impacts and Adaptation Strategies of Agriculture in Mediterranean-Climate Regions (MCRs). Sustainability 2019, 11, 2769. [Google Scholar] [CrossRef] [Green Version]
- Latcham, R.E. La Agricultura Precolombina en Chile y los Países Vecinos; Universidad de Chile: Santiago, Chile, 1936; p. 336. [Google Scholar]
- De Vivar, G.; Sáez-Godoy, L. Crónica y Relación Copiosa y Verdadera de los Reinos de Chile (1558); Colloquium-Verlag: Berlín, Germany, 1979; p. 343. [Google Scholar]
- Tapia, G.; González, M.; Burgos, J.; Vega, M.V.; Méndez, J.; Inostroza, L. Early transcriptional responses in Solanum peruvianum and Solanum lycopersicum account for different acclimation processes during water scarcity events. Sci. Rep. 2021, 11, 1–14. [Google Scholar] [CrossRef]
- Inostroza, L.; Espinoza, S.; Barahona, V.; Gerding, M.; Humphries, A.; del Pozo, A.; Ovalle, C. Phenotypic Diversity and Productivity of Medicago sativa Subspecies from Drought-Prone Environments in Mediterranean Type Climates. Plants 2021, 10, 862. [Google Scholar] [CrossRef]
- Baginsky, C.; Ramos, L. Situación de las legumbres en Chile: Una mirada agronómica. Rev. Chil. Nutr. 2018, 45, 21–31. [Google Scholar] [CrossRef] [Green Version]
- Bascur, G.; Tay, J. Colecta, Caracterización y Utilización de la Variabilida Genética en Germoplasma Chileno de Poroto (Phaseolus vulgaris L.). Agric. Técnica 2005, 65, 135–146. [Google Scholar] [CrossRef]
- Paredes, O.M.; Gepts, P. Extensive introgression of Middle American germplasm into Chilean common bean cultivars. Genet. Resour. Crop Evol. 1995, 42, 29–41. [Google Scholar] [CrossRef]
- Blair, M.W.; Díaz S, J.M.; Duque, M.C.; Hidalgo, R. Evidencia Molecular de Diferenciación Genética e Introgresión Entre Razas de Fríjol Común del Acervo Andino. Acta Agronómica 2007, 56, 165–170. Available online: http://www.scielo.org.co/scielo.php?script=sci_arttext&pid=S0120-28122007000400002 (accessed on 21 September 2021).
- Paredes, C.; Becerra, V.; Tay, U.; Blair, M.W.; Bascur, G.B.G. Selection of a Representative Core Collection from the Chilean Common Bean Germplasm. Chil. J. Agric. Res. 2010, 70, 3–15. [Google Scholar] [CrossRef] [Green Version]
- Ramírez-Vallejo, P.; Kelly, J.D. Traits related to drought resistance in common bean. Euphytica 1998, 99, 127–136. [Google Scholar] [CrossRef]
- Assefa, T.; Beebe, S.E.; Rao, I.M.; Cuasquer, J.B.; Duque, M.C.; Rivera, M.; Battisti, A.; Lucchin, M. Pod harvest index as a selection criterion to improve drought resistance in white pea bean. Field Crop. Res. 2013, 148, 24–33. [Google Scholar] [CrossRef]
- Polania, J.; Rao, I.M.; Cajiao, C.; Rivera, M.; Raatz, B.; Beebe, S. Physiological traits associated with drought resistance in Andean and Mesoamerican genotypes of common bean (Phaseolus vulgaris L.). Euphytica 2016, 210, 17–29. [Google Scholar] [CrossRef]
- Terán, H.; Singh, S.P. Comparison of Sources and Lines Selected for Drought Resistance in Common Bean. Crop Sci. 2002, 42, 64–70. [Google Scholar] [CrossRef]
- Beebe, S.E.; Rao, I.M.; Cajiao, C.; Grajales, M. Selection for Drought Resistance in Common Bean Also Improves Yield in Phosphorus Limited and Favorable Environments. Crop Sci. 2008, 48, 582–592. [Google Scholar] [CrossRef]
- Beebe, S.E.; Rao, I.M.; Blair, M.W. Breeding for Abiotic Stress Tolerance in Common Bean: Present and Future Challenges. 2009. Available online: https://www.researchgate.net/publication/228612601 (accessed on 21 September 2021).
- Hageman, A.N.; Urban, M.O.; van Volkenburgh, E. Sensitivity of leaflet growth rate to drought predicts yield in common bean (Phaseolus vulgaris). Funct. Plant Biol. 2020, 47, 792–802. [Google Scholar] [CrossRef] [PubMed]
- Hageman, A.; van Volkenburgh, E. Sink Strength Maintenance Underlies Drought Tolerance in Common Bean. Plants 2021, 10, 489. [Google Scholar] [CrossRef] [PubMed]
- Farooq, M.; Wahid, A.; Kobayashi, N.; Fujita, D.; Basra, S.M.A. Plant drought stress: Effects, mechanisms and management. Agron. Sustain. Dev. 2009, 29, 185–212. [Google Scholar] [CrossRef] [Green Version]
- Rosales, M.A.; Ocampo, E.; Rodríguez-Valentín, R.; Olvera-Carrillo, Y.; Acosta-Gallegos, J.; Covarrubias, A.A. Physiological analysis of common bean (Phaseolus vulgaris L.) cultivars uncovers characteristics related to terminal drought resistance. Plant Physiol. Biochem. 2012, 56, 24–34. [Google Scholar] [CrossRef]
- Daryanto, S.; Wang, L.; Jacinthe, P.-A. Global Synthesis of Drought Effects on Food Legume Production. PLoS ONE 2015, 10, e0127401. [Google Scholar] [CrossRef] [Green Version]
- Gallegos, J.A.A.; Shibata, J.K. Effect of water stress on growth and yield of indeterminate dry-bean (Phaseolus vulgaris) cultivars. Field Crop. Res. 1989, 20, 81–93. [Google Scholar] [CrossRef]
- Graham, P.; Ranalli, P. Common bean (Phaseolus vulgaris L.). Field Crop. Res. 1997, 53, 131–146. [Google Scholar] [CrossRef]
- Turner, N.C.; Wright, G.C.; Siddique, K.H.M. Adaptation of grain legumes (pulses) to water-limited environments. Adv. Agron. 2001, 71, 193–231. [Google Scholar] [CrossRef]
- Inostroza, L.; Acuña, H. Water use efficiency and associated physiological traits of nine naturalized white clover populations in Chile. Plant Breed. 2010, 129, 700–706. [Google Scholar] [CrossRef]
- Fahad, S.; Bajwa, A.; Nazir, U.; Anjum, S.A.; Farooq, A.; Zohaib, A.; Sadia, S.; Nasim, W.; Adkins, S.; Saud, S.; et al. Crop Production under Drought and Heat Stress: Plant Responses and Management Options. Front. Plant Sci. 2017, 8, 1147. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sehgal, A.; Sita, K.; Siddique, K.H.M.; Kumar, R.; Bhogireddy, S.; Varshney, R.K.; HanumanthaRao, B.; Nair, R.M.; Prasad, P.V.V.; Nayyar, H. Drought or/and Heat-Stress Effects on Seed Filling in Food Crops: Impacts on Functional Biochemistry, Seed Yields, and Nutritional Quality. Front. Plant Sci. 2018, 9, 1705. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beebe, S. Common Bean Breeding in the Tropics. Plant Breed. Rev. 2012, 36, 357–426. [Google Scholar] [CrossRef]
- Lanna, A.C.; Silva, R.A.; Ferraresi, T.M.; Mendonça, J.A.; Coelho, G.R.C.; Moreira, A.; Valdisser, P.A.M.R.; Brondani, C.; Vianello, R.P. Physiological characterization of common bean (Phaseolus vulgaris L.) under abiotic stresses for breeding purposes. Environ. Sci. Pollut. Res. 2018, 25, 31149–31164. [Google Scholar] [CrossRef]
- Tapia, G.; Méndez, J.; Inostroza, L. Different combinations of morpho-physiological traits are responsible for tolerance to drought in wild tomatoes Solanum chilense and Solanum peruvianum. Plant Biol. 2015, 18, 406–416. [Google Scholar] [CrossRef]
- Teran, J.C.B.M.Y.; Konzen, E.R.; Medina, V.; Palkovic, A.; Ariani, A.; Tsai, S.M.; Gilbert, M.; Gepts, P. Root and shoot variation in relation to potential intermittent drought adaptation of Mesoamerican wild common bean (Phaseolus vulgaris L.). Ann. Bot. 2019, 124, 917–932. [Google Scholar] [CrossRef] [Green Version]
- Viviana Becerra, V.; Mario Paredes, C.; Carmen Rojo, M.; Díaz, L.M.; Blair, M.W. Microsatellite Marker Characterization of Chilean Common Bean (Phaseolus vulgaris L.) Germplasm. Crop Sci. 2010, 50, 1932–1941. [Google Scholar] [CrossRef]
- White, J.; Laing, D. Photoperiod response of flowering in diverse genotypes of common bean (Phaseolus vulgaris). Field Crop. Res. 1989, 22, 113–128. [Google Scholar] [CrossRef]
- White, J.W.; Singh, S.P.; Pino, C.; Buddenhagen, I. Effects of seed size and photoperiod response on crop growth and yield of common bean. Field Crop. Res. 1992, 28, 295–307. [Google Scholar] [CrossRef]
- Gutierrez, L.; van Wuytswinkel, O.; Castelain, M.; Bellini, C. Combined networks regulating seed maturation. Trends Plant Sci. 2007, 12, 294–300. [Google Scholar] [CrossRef]
- Bennett, E.J.; Roberts, J.A.; Wagstaff, C. The role of the pod in seed development: Strategies for manipulating yield. New Phytol. 2011, 190, 838–853. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kyparissis, A.; Drilias, P.; Manetas, Y. Seasonal fluctuations in photoprotective (xanthophyll cycle) and photoselective (chlorophylls) capacity in eight Mediterranean plant species belonging to two different growth forms. Funct. Plant Biol. 2000, 27, 265–272. [Google Scholar] [CrossRef]
- Munné-Bosch, S.; Alegre, L. Changes in carotenoids, tocopherols and diterpenes during drought and recovery, and the biological significance of chlorophyll loss in Rosmarinus officinalis plants. Planta 2000, 210, 925–931. [Google Scholar] [CrossRef] [PubMed]
- Baccari, S.; Elloumi, O.; Chaari-Rkhis, A.; Fenollosa, E.; Morales, M.; Drira, N.; Ben Abdallah, F.; Fki, L.; Munné-Bosch, S. Linking Leaf Water Potential, Photosynthesis and Chlorophyll Loss with Mechanisms of Photo- and Antioxidant Protection in Juvenile Olive Trees Subjected to Severe Drought. Front. Plant Sci. 2020, 11, 614144. [Google Scholar] [CrossRef] [PubMed]
- Darkwa, K.; Ambachew, D.; Mohammed, H.; Asfaw, A.; Blair, M.W. Evaluation of common bean (Phaseolus vulgaris L.) genotypes for drought stress adaptation in Ethiopia. Crop J. 2016, 4, 367–376. [Google Scholar] [CrossRef] [Green Version]
- McLaughlin, J.; Boyer, J.S. Sugar-responsive Gene Expression, Invertase Activity, and Senescence in Aborting Maize Ovaries at Low Water Potentials. Ann. Bot. 2004, 94, 675–689. [Google Scholar] [CrossRef]
- Barnabás, B.; Jäger, K.; Fehér, A. The effect of drought and heat stress on reproductive processes in cereals. Plant Cell Environ. 2008, 31, 11–38. [Google Scholar] [CrossRef]
- Hedhly, A.; Hormaza, J.I.; Herrero, M. Global warming and sexual plant reproduction. Trends Plant Sci. 2009, 14, 30–36. [Google Scholar] [CrossRef]
- Patrick, J.W.; Stoddard, F.L. Physiology of flowering and grain filling in faba bean. Field Crop. Res. 2010, 115, 234–242. [Google Scholar] [CrossRef]
- Ochatt, S.J. Agroecological impact of an in vitro biotechnology approach of embryo development and seed filling in legumes. Agron. Sustain. Dev. 2015, 35, 535–552. [Google Scholar] [CrossRef] [Green Version]
- Turc, O.; Tardieu, F. Drought affects abortion of reproductive organs by exacerbating developmentally driven processes via expansive growth and hydraulics. J. Exp. Bot. 2018, 69, 3245–3254. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Banan, D.; van Volkenburgh, E. Growth response of common bean (Phaseolus vulgaris L.) lines to water deficit. Bean Improv. Coop. Annu. Rep. 2012, 55, 1–2. [Google Scholar]
- Staden, J.; Carmi, A. The effects of decapitation on the distribution of cytokinins and growth of Phaseolus vulgaris plants. Physiol. Plant. 1982, 55, 39–44. [Google Scholar] [CrossRef]
- Tenhaken, R. Cell wall remodeling under abiotic stress. Front. Plant Sci. 2015, 5, 771. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, F.; Andersen, M.N.; Jensen, C.R. Loss of pod set caused by drought stress is associated with water status and ABA content of reproductive structures in soybean. Funct. Plant Biol. 2003, 30, 271–280. [Google Scholar] [CrossRef]
- Clauw, P.; Coppens, F.; Korte, A.; Herman, D.; Slabbinck, B.; Dhondt, S.; van Daele, T.; de Milde, L.; Vermeersch, M.; Maleux, K.; et al. Leaf Growth Response to Mild Drought: Natural Variation in Arabidopsis Sheds Light on Trait Architecture. Plant Cell 2016, 28, 2417–2434. [Google Scholar] [CrossRef] [Green Version]
- Westgate, M.E.; Grant, D.T. Effect of Water Deficits on Seed Development in Soybean. Plant Physiol. 1989, 91, 975–979. [Google Scholar] [CrossRef] [Green Version]
- Acosta-Díaz, E.; Acosta-Gallegos, J.A.; Trejo-López, C.; Padilla-Ramírez, J.S.; Amador-Ramírez, M.D. Adaptation traits in dry bean cultivars grown under drought stress. Agric. Técnica México 2009, 35, 419–428. [Google Scholar]
- Stolpe, N. Descripciones de los Principales Suelos de la VIII Región de Chile; Universidad de Concepción, Departamento de Suelos y Recursos Naturales: Chillán, Chile, 2006; Volume 1. [Google Scholar]
- Cabello, R.; Monneveux, P.; de Mendiburu, F.; Bonierbale, M. Comparison of yield based drought tolerance indices in improved varieties, genetic stocks and landraces of potato (Solanum tuberosum L.). Euphytica 2013, 193, 147–156. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Landrace | Pod N° (Pod/m2) | Seed N° (Seed/m2) | Seed by Pod | PHI (%) | ||||
|---|---|---|---|---|---|---|---|---|
| OW | RW | OW | RW | OW | RW | OW | RW | |
| Enriqueta | 340.4 | 206.7 | 1457.3 | 553.0 | 4.2 | 2.7 | 67.5 | 64.0 |
| Coyunda | 451.3 | 299.2 | 2136.6 | 945.4 | 4.8 | 3.2 | 74.2 | 65.6 |
| Blanco | 572.1 | 325.8 | 2227.7 | 987.7 | 3.9 | 3.0 | 69.5 | 66.3 |
| Pinto | 352.5 | 254.6 | 1281.8 | 767.7 | 3.7 | 3.0 | 79.4 | 77.4 |
| Negro argel | 465.8 | 286.3 | 2518.4 | 1108.7 | 5.4 | 3.8 | 78.8 | 76.5 |
| Coscorron | 437.9 | 290.8 | 1492.5 | 948.4 | 3.4 | 3.3 | 74.4 | 81.2 |
| Bayote | 353.3 | 202.9 | 1479.3 | 707.1 | 4.2 | 3.6 | 72.2 | 68.5 |
| Tortola | 390.4 | 240.0 | 1429.4 | 663.9 | 3.6 | 2.8 | 75.7 | 71.5 |
| Manteca | 520.8 | 303.8 | 2353.9 | 1037.9 | 4.6 | 3.4 | 79.5 | 75.9 |
| p ≤ 0.05 | * | ns | * | * | ns | * | ** | *** |
| LSD | 114.2 | - | 600.4 | 294.0 | - | 1.0 | 4.6 | 3.4 |
| Mean | 431.6 a | 267.8 b | 1819.7 a | 857.8 b | 4.2 a | 3.2 b | 74.6 a | 71.9 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tapia, G.; Méndez, J.; Inostroza, L.; Lozano, C. Water Shortage Affects Vegetative and Reproductive Stages of Common Bean (Phaseolus vulgaris) Chilean Landraces, Differentially Impacting Grain Yield Components. Plants 2022, 11, 749. https://doi.org/10.3390/plants11060749
Tapia G, Méndez J, Inostroza L, Lozano C. Water Shortage Affects Vegetative and Reproductive Stages of Common Bean (Phaseolus vulgaris) Chilean Landraces, Differentially Impacting Grain Yield Components. Plants. 2022; 11(6):749. https://doi.org/10.3390/plants11060749
Chicago/Turabian StyleTapia, Gerardo, José Méndez, Luis Inostroza, and Camila Lozano. 2022. "Water Shortage Affects Vegetative and Reproductive Stages of Common Bean (Phaseolus vulgaris) Chilean Landraces, Differentially Impacting Grain Yield Components" Plants 11, no. 6: 749. https://doi.org/10.3390/plants11060749
APA StyleTapia, G., Méndez, J., Inostroza, L., & Lozano, C. (2022). Water Shortage Affects Vegetative and Reproductive Stages of Common Bean (Phaseolus vulgaris) Chilean Landraces, Differentially Impacting Grain Yield Components. Plants, 11(6), 749. https://doi.org/10.3390/plants11060749