
The Endemic Vascular Flora of Sardinia: A Dynamic Checklist with an Overview of Biogeography and Conservation Status

 ,
,  ,
,  ,
,  and
and 
Abstract
1. Introduction
2. Results
3. Discussion
3.1. Quantitative Features
3.2. Taxonomical Analysis
3.3. Life Forms
3.4. Chorological Types
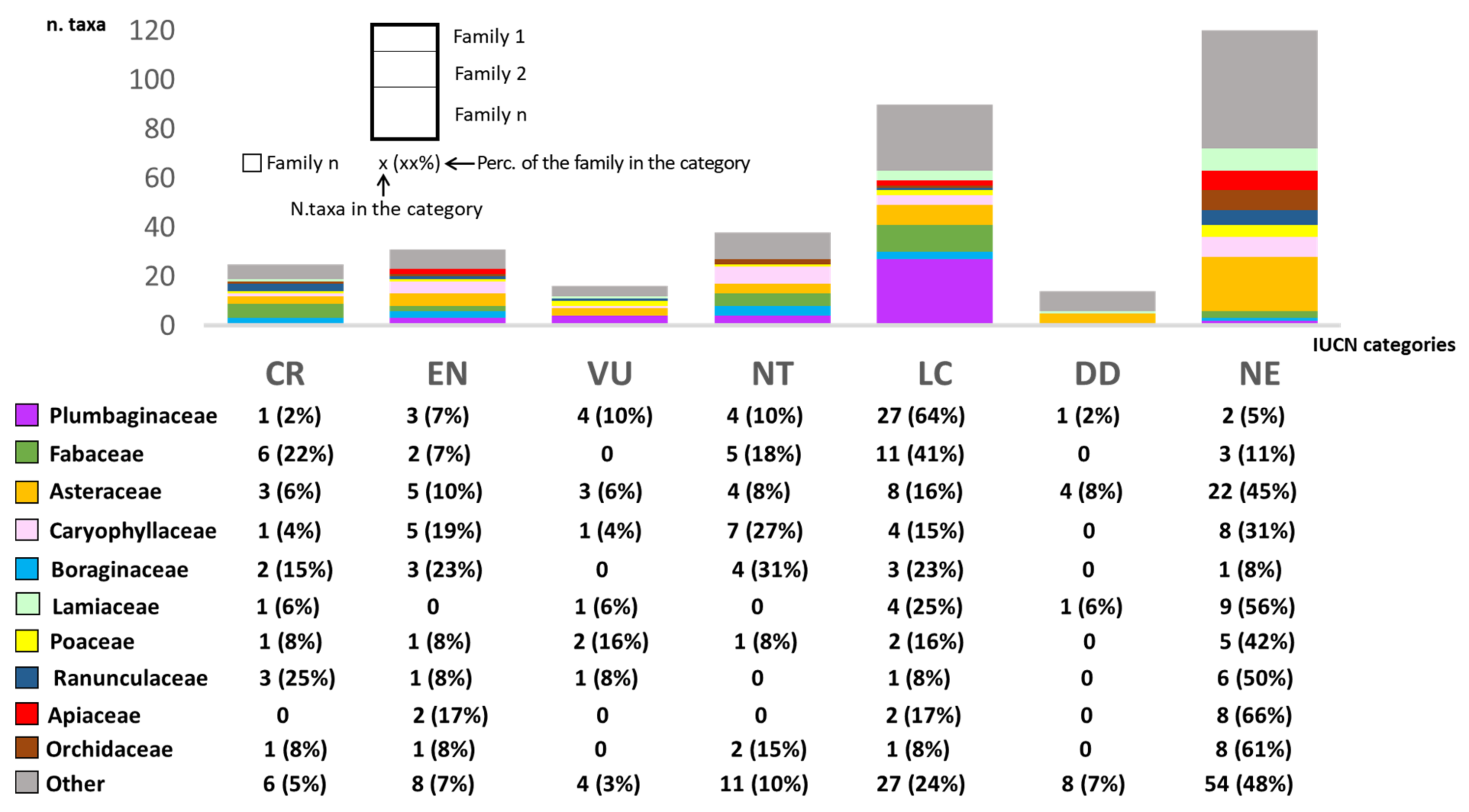
3.5. Conservation Status of the Endemic Sardinian Flora
4. Materials and Methods
4.1. Study Area
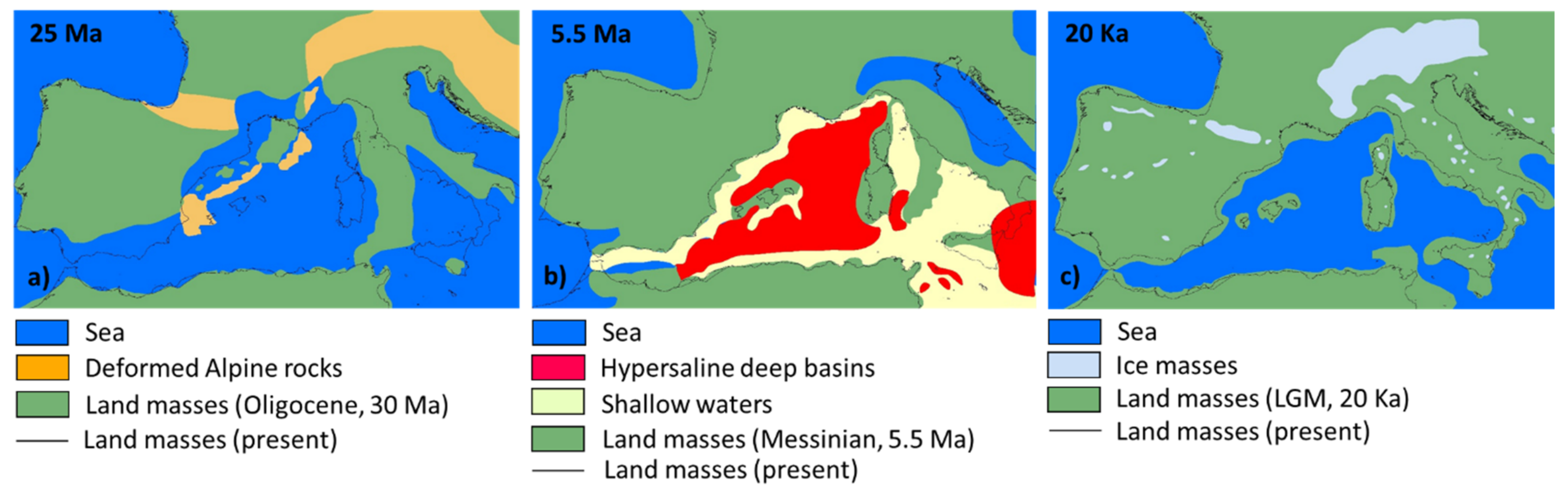
4.2. Origin and Evolution of the Endemic Component of the Sardinian Flora
4.3. Criteria for the Selection of Endemic Taxa
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Thompson, J.D. Plant Evolution in the Mediterranean; Oxford University Press: Oxford, UK, 2005; pp. 1–304. [Google Scholar]
- Favarger, C.; Contandriopoulos, J. Essai sur l’endémisme. Bull. Soc. Bot. Suisse 1961, 77, 383–408. [Google Scholar]
- Contandriopoulos, J.; Favarger, C. Problémes posés par l’endémisme en Méditerranée. Colloq. Int. CNRS 1975, 235, 175–194. [Google Scholar]
- Mansion, G.; Rosenbaum, G.; Schoenenberger, N.; Bacchetta, G.; Rosselló, J.A.; Conti, E. Phylogenetic analysis informed by geological history supports multiple, sequential invasions of the Mediterranean Basin by the angiosperm family Araceae. System. Biol. 2008, 57, 269–285. [Google Scholar] [CrossRef]
- Mansion, G.; Selvi, F.; Guggisberg, A.; Conti, E. Origin of Mediterranean insular endemics in the Boraginales: Integrative evidence from molecular dating and ancestral area reconstruction. J. Biogeogr. 2009, 36, 1282–1296. [Google Scholar] [CrossRef]
- Molins, A.; Bacchetta, G.; Rosato, M.; Rosselló, J.A.; Mayol, M. Molecular phylogeography of Thymus herba-barona (Lamiaceae): Insight into the evolutionary history of the flora of the western Mediterranean islands. Taxon 2011, 60, 1295–1305. [Google Scholar] [CrossRef]
- Carnicero, P.; Schönswetter, P.; Garcia-Jacas, N.; Galbany-Casals, M. Is there a need for accepting paraphyletic taxa? A case study in the Sardinian endemic Cymbalaria muelleri (Plantaginaceae). Bot. J. Linn. Soc. 2019, 191, 325–338. [Google Scholar] [CrossRef]
- López-Alvarado, J.; Mameli, G.; Farris, E.; Susanna, A.; Filigheddu, R.; Garcia-Jacas, N. Islands as a crossroad of evolutionary lineages: A case study of Centaurea sect. Centaurea (Compositae) from Sardinia (Mediterranean Basin). PLoS ONE 2020, 15, e0228776. [Google Scholar] [CrossRef]
- Arrigoni, P.V.; Camarda, I.; Corrias, B.; Diana Corrias, S.; Nardi, E.; Raffaelli, M.; Valsecchi, F. Le piante endemiche della Sardegna. Boll. Soc. Sarda. Sci. Nat. 1976–1992, 1–202, 1–1050. [Google Scholar]
- Arrigoni, P.V. Le piante endemiche della Sardegna. Introduzione. Boll. Soc. Sarda. Sci. Nat. 1976, 16, 259–264. [Google Scholar]
- Bacchetta, G.; Farris, E.; Pontecorvo, C. A new method to set conservation priorities in biodiversity hotspots. Plant Biosyst. 2012, 146, 638–648. [Google Scholar] [CrossRef]
- Thompson, J.D. Plant Evolution in the Mediterranean, Insights for Conservation, 2nd ed.; Oxford University Press: Oxford, UK, 2020; pp. 1–352. [Google Scholar] [CrossRef]
- Bacchetta, G.; Pontecorvo, C. Contribution to the knowledge of the endemic vascular flora of Iglesiente (SW Sardinia-Italy). Candollea 2005, 60, 481–501. [Google Scholar]
- Nikolić, T.; Antonić, O.; Alegro, A.L.; Dobrović, I.; Bogdanović, S.; Liber, Z.; Rešetnik, I. Plant species diversity of Adriatic islands: An introductory survey. Plant Biosyst. 2008, 142, 435–445. [Google Scholar] [CrossRef]
- Jeanmonod, D. Révision de la section Siphonomorpha Otth du genre Silene, L. en Méditerranée occidentale. II: Le Group Du Silene mollissima. Candollea 1984, 39, 195–259. [Google Scholar]
- Bacchetta, G.; Coppi, A.; Pontecorvo, C.; Selvi, F. Systematics, phylogenetic relationships and conservation of the taxa of Anchusa (Boraginaceae) endemic to Sardinia (Italy). Syst. Biodivers. 2008, 6, 161–174. [Google Scholar] [CrossRef]
- Gentili, R.; Fenu, G.; Mattana, E.; Citterio, S.; De Mattia, F.; Bacchetta, G. Conservation genetics of two island endemic Ribes spp. (Grossulariaceae) of Sardinia: Survival or extinction? Plant Biol. 2015, 17, 1085–1094. [Google Scholar] [CrossRef]
- Laffan, S.W.; Crisp, M.D. Assessing endemism at multiple spatial scales, with an example from the Australian vascular flora. J. Biogeogr. 2003, 30, 511–520. [Google Scholar] [CrossRef]
- Bacchetta, G.; Fenu, G.; Mattana, E. A checklist of the exclusive vascular flora of Sardinia with priority rankings for conservation. An. Jard. Bot. Madr. 2012, 69, 81–89. [Google Scholar] [CrossRef]
- Peruzzi, L.; Conti, F.; Bartolucci, F. An inventory of vascular plants endemic to Italy. Phytotaxa 2014, 168, 1–75. [Google Scholar] [CrossRef]
- Farris, E.; Carta, M.; Circosta, S.; Falchi, S.; Papuga, G.; de Lange, P. The indigenous vascular flora of the forest domain of Anela (Sardinia, Italy). Phytokeys 2018, 113, 97–143. [Google Scholar] [CrossRef]
- Hobohm, C. Endemism in Vascular Plants; Springer Press: Amsterdam, The Netherlands, 2014; pp. 1–348. [Google Scholar] [CrossRef]
- Cañadas, E.M.; Fenu, G.; Peñas, J.; Lorite, J.; Mattana, E.; Bacchetta, G. Hotspots within hotspots: Endemic plant richness, environmental drivers, and implications for conservation. Biol. Conserv. 2014, 170, 282–291. [Google Scholar] [CrossRef]
- Médail, F.; Diadema, K. Glacial refugia influence plant diversity patterns in the Mediterranean Basin. J. Biogeogr. 2009, 36, 1333–1345. [Google Scholar] [CrossRef]
- Fineschi, S.; Cozzolino, S.; Migliaccio, M.; Musacchio, A.; Innocenti, M.; Vendramin, G.G. Sicily represents the Italian reservoir of chloroplast DNA diversity of Quercus ilex L. (Fagaceae). Ann. For. Sci. 2005, 62, 79–84. [Google Scholar] [CrossRef]
- Agrimonti, C.; Bianchi, R.; Bianchi, A.; Ballero, M.; Poli, F.; Marmiroli, N. Understanding biological conservation strategies: A molecular-genetic approach to the case of myrtle (Myrtus communis L.) in two Italian regions: Sardinia and Calabria. Conserv. Genet. 2007, 8, 385–396. [Google Scholar] [CrossRef]
- Fernández-Mazuecos, M.; Jiménez-Mejías, P.; Rotllan-Puig, X.; Vargas, P. Narrow endemics to Mediterranean islands: Moderate genetic diversity but narrow climatic niche of the ancient, critically endangered Naufraga (Apiaceae). Perspect. Plant Ecol. Evol. 2014, 16, 190–202. [Google Scholar] [CrossRef]
- Médail, F. Plant biogeography and vegetation patterns of the Mediterranean islands. Bot. Rev. 2021, in press. [Google Scholar] [CrossRef]
- Fenu, G.; Fois, M.; Cañadas, E.M.; Bacchetta, G. Using endemic-plant distribution, geology and geomorphology in biogeography: The case of Sardinia (Mediterranean Basin). System. Biodivers. 2014, 12, 181–193. [Google Scholar] [CrossRef]
- Fois, M.; Fenu, G.; Cañadas, E.M.; Bacchetta, G. Disentangling the influence of environmental and anthropogenic factors on the distribution of endemic vascular plants in Sardinia. PLoS ONE 2017, 12, e0182539. [Google Scholar] [CrossRef]
- Bacchetta, G.; Iiriti, G.; Pontecorvo, C. Contributo alla conoscenza della flora vascolare endemica della Sardegna. Inform. Bot. Ital. 2005, 37, 306–307. [Google Scholar]
- Orsenigo, S.; Montagnani, C.; Fenu, G.; Gargano, D.; Peruzzi, L.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Bovio, M.; et al. Red Listing plants under full national responsibility: Extinction risk and threats in the vascular flora endemic to Italy. Biol. Conserv. 2018, 224, 213–222. [Google Scholar] [CrossRef]
- Orsenigo, S.; Fenu, G.; Gargano, D.; Montagnani, C.; Abeli, T.; Alessandrini, A.; Bacchetta, G.; Bartolucci, F.; Carta, A.; Castello, M.; et al. Red list of threatened vascular plants in Italy. Plant Biosyst. 2021, 155, 310–335. [Google Scholar] [CrossRef]
- Budroni, M.A.; Farris, E.; Zirulia, A.; Pisanu, S.; Filigheddu, R.; Rustici, M. Evidence for age-structured depensation effect in fragmented plant populations: The case of the Mediterranean endemic Anchusa sardoa (Boraginaceae). Ecol. Complex. 2014, 20, 142–150. [Google Scholar] [CrossRef]
- Bacchetta, G.; Fenu, G.; Mattana, E.; Pontecorvo, C. Ecological remarks on Astragalus maritimus and A. verrucosus, two threatened exclusive endemic species of Sardinia. Acta Bot. Gall. 2011, 158, 79–91. [Google Scholar] [CrossRef]
- Pisanu, S.; Farris, E.; Filigheddu, R.; García, M.B. Demographic effects of large, introduced herbivores on a long-lived endemic plant. Plant Ecol. 2012, 213, 1543–1553. [Google Scholar] [CrossRef]
- Farris, E.; Filigheddu, R.; Mameli, G.; Falanga, V.; Vanetti, I.; Rosati, L.; Binelli, G. Is population genetic structure of vascular plants shaped more by ecological or geographic factors? A study case on the Mediterranean endemic Centaurea filiformis (Asteraceae). Plant Biol. 2018, 20, 936–947. [Google Scholar] [CrossRef] [PubMed]
- Bagella, S.; Caria, M.C.; Farris, E.; Filigheddu, R. Spatial-time variability and conservation relevance of plant communities in Mediterranean temporary wet habitats: A case study in Sardinia (Italy). Plant Biosyst. 2009, 143, 435–442. [Google Scholar] [CrossRef]
- Farris, E.; Rosati, L.; Secchi, Z.; Filigheddu, R. Are all pastures eligible for conservation? A phytosociological survey of the Sardinian-Corsican province as a basic tool for the Habitats Directive. Plant Biosyst. 2013, 147, 931–946. [Google Scholar] [CrossRef]
- Fenu, G.; Cogoni, D.; Ulian, T.; Bacchetta, G. The impact of human trampling on a threatened coastal Mediterranean plant: The case of Anchusa littorea Moris (Boraginaceae). Flora 2013, 208, 104–110. [Google Scholar] [CrossRef]
- Farris, E.; Pisanu, S.; Ceccherelli, G.; Filigheddu, R. Human trampling effects on Mediterranean coastal dune plants. Plant Biosyst. 2013, 147, 1043–1051. [Google Scholar] [CrossRef]
- Fois, M.; Fenu, G.; Bacchetta, G. Global analyses underrate part of the story: Finding applicable results for the conservation planning of small Sardinian islets’ flora. Biodivers. Conserv. 2016, 25, 1091–1106. [Google Scholar] [CrossRef]
- Fois, M.; Podda, L.; Médail, F.; Bacchetta, G. Endemic and alien vascular plant diversity in the small Mediterranean islands of Sardinia: Drivers and implications for their conservation. Biol. Conserv. 2020, 244, 108519. [Google Scholar] [CrossRef]
- Fenu, G.; Fois, M.; Cogoni, D.; Porceddu, M.; Pinna, M.S.; Lombraña, A.C.; Nebot, A.; Sulis, E.; Picciau, R.; Santo, A.; et al. The Aichi Biodiversity Target 12 at regional level: An achievable goal? Biodiversity 2015, 16, 120–135. [Google Scholar] [CrossRef]
- Pasta, S.; Perez-Graber, A.; Fazan, L.; de Montmollin, B. The Top 50 Mediterranean Island Plants UPDATE 2017; IUCN/SSC/Mediterranean Plant Specialist Group: Neuchâtel, Switzerland, 2017; pp. 1–141. Available online: https://top50.iucn-mpsg.org/book (accessed on 20 April 2021).
- Allioni, C. Fasciculus stirpium Sardiniae in Diocesi Calari lectarum a M.A. Plazza, Chirurgo taurinensi, quos in usum botanicorum recenset. C.A. Misc. Phil.-Math. Soc. Priv. Taur. 1759, 1, 88–103. [Google Scholar]
- Moris, G.G. Flora Sardoa; Ex Regio Typographeo: Taurini, Italy, 1837–1859; Volume 1–3. [Google Scholar]
- Barbey, W. Florae Sardoae Compendium; Georges Bridel Editeur: Lausanne, Switzerland, 1884. [Google Scholar]
- Martelli, U. Monocotyledones Sardoae; Tip. Niccolai: Firenze, Italy, 1896; Volume 1–2. [Google Scholar]
- Martelli, U. Monocotyledones Sardoae, 3rd ed.; Casciano, R.S., Ed.; Stab. Tip. Cappelli: Naples, Italy, 1904. [Google Scholar]
- Terracciano, A. La Flora Sardoa di M.A. Plazza da Villafranca redatta coi suoi manoscritti. Mem. R. Acc. Sc. Torino 1914, 64, 1–54. [Google Scholar]
- Terracciano, A. La Flora Sardoa di M.A. Plazza da Villafranca redatta coi suoi manoscritti. Mem. R. Acc. Sc. Torino 1914, 65, 1–53. [Google Scholar]
- Terracciano, A. La Flora Sardoa di M.A. Plazza da Villafranca redatta coi suoi manoscritti. Mem. R. Acc. Sc. Torino 1930, 67, 1–78. [Google Scholar]
- Arrigoni, P.V. Flora dell’Isola di Sardegna (Flora of the island of Sardinia); Delfino: Sassari, Italy, 2006–2015; Volume 1–6. [Google Scholar]
- Viciani, D.; Ferretti, G.; Bernardini, A.; Raffaelli, M.; Lastrucci, L. An annotated checklist of the vascular flora of the Arezzo administrative province (Tuscany, Italy). Plant Biosyst. 2020, 155, 971–982. [Google Scholar] [CrossRef]
- Beierkuhnlein, C.; Walentowitz, A.; Welss, W. FloCan—A Revised Checklist for the Flora of the Canary Islands. Diversity 2021, 13, 480. [Google Scholar] [CrossRef]
- Duminil, J.; Di Michele, M. Plant species delimitation: A comparison of morphological and molecular markers. Plant Biosyst. 2009, 143, 528–542. [Google Scholar] [CrossRef]
- Arrigoni, P.V.; Diana, S. Karyology, chorology and bioecology of the genus Limonium (Plumbaginaceae) in Sardinia. Plant Biosyst. 1999, 133, 63–71. [Google Scholar] [CrossRef]
- Bacchetta, G.; Brullo, S.; Terrasi, M.C. A new species of Hypochaeris L. (Asteraceae, Cichorieae) from Sardinia. Willdenowia 2003, 33, 71–78. [Google Scholar] [CrossRef]
- Lussu, M.; Marignani, M.; Lai, R.; Loi, M.C.; Cogoni, A.; Cortis, P. A Synopsis of Sardinian Studies: Why Is it Important to Work on Island Orchids? Plants 2020, 9, 853. [Google Scholar] [CrossRef]
- Bedini, G.; Garbari, F.; Peruzzi, L. Karyological knowledge of Italian vascular flora as inferred by the analysis of Chrobase.it. Plant Biosyst. 2012, 145, 889–899. [Google Scholar] [CrossRef]
- Romeiras, M.M.; Carine, M.; Duarte, M.C.; Catarino, S.; Dias, F.S.; Borda-de-Água, L. Bayesian methods to analyze historical collections in time and space: A case study using Cabo Verde Endemic Flora. Front. Plant Sci. 2020, 11, 278. [Google Scholar] [CrossRef] [PubMed]
- Médail, F. The specific vulnerability of plant biodiversity and vegetation on Mediterranean islands in the face of global change. Reg. Environ. Change 2017, 17, 1775–1790. [Google Scholar] [CrossRef]
- Zhu, H. Biogeographical evidences help revealing the origin of Hainan Island. PLoS ONE 2016, 11, e0151941. [Google Scholar] [CrossRef] [PubMed]
- Meyer, J.-Y. Threat of invasive alien plants to native flora and forest vegetation of Eastern Polynesia. Pac. Sci. 2004, 58, 357–375. [Google Scholar] [CrossRef]
- Médail, F.; Quezel, P. Hot-Spots analysis for conservation of plant biodiversity in the Mediterranean Basin. Ann. Mo. Bot. Gard. 1997, 84, 112–127. [Google Scholar] [CrossRef]
- Médail, F.; Quezel, P. Biodiversity Hot-spots in the Mediterranean Basin: Setting global conservation priorities. Conserv. Biol. 1999, 13, 1510–1513. [Google Scholar] [CrossRef]
- Brullo, C.; Brullo, S.; Giusso del Galdo, G. Considerations on the endemic flora of Sicily. In Proceedings of the Second Botanical Conference in Menorca “Islands and Plants: Preservation and understanding of flora on Mediterranean islands”, Menorca, Spain, 26–30 April 2011; Cardona Pons, E., Estaun Clarisó, I., Comas Casademont, M., Fraga i Arguimbau, P., Eds.; Menorquí d’Estudis, Consell Insular de Menorca: Maó, Spain, 2013; pp. 177–199. [Google Scholar]
- Sáez, L.; Fraga, P.; López-Alvarado, J. The Flora of the Balearic Islands. In Proceedings of the Second Botanical Conference in Menorca “Islands and Plants: Preservation and Understanding of Flora on Mediterranean Islands”, Menorca, Spain, 26–30 April 2011; Cardona Pons, E., Estaun Clarisó, I., Comas Casademont, M., Fraga i Arguimbau, P., Eds.; Menorquí d’Estudis, Consell Insular de Menorca: Maó, Spain, 2013; pp. 91–103. [Google Scholar]
- Jeanmonod, D.; Naciri, Y.; Schlüssel, A.; Gamisans, J. Floristic analyses of the Corsican flora: Biogeographical origin and endemism. Candollea 2015, 70, 21–41. [Google Scholar] [CrossRef]
- Cho, M.-S.; Yang, J.Y.; Yang, T.-J.; Kim, S.-C. Evolutionary comparison of the chloroplast genome in the woody Sonchus alliance (Asteraceae) on the Canary Islands. Genes 2019, 10, 217. [Google Scholar] [CrossRef]
- Dolcher, T.; Pignatti, S. Un’ipotesi sull’evoluzione dei Limonium del bacino del Mediterraneo. Plant Biosyst. 1971, 105, 95–107. [Google Scholar] [CrossRef]
- Koutroumpa, K.; Warren, B.H.; Theodoridis, S.; Coiro, M.; Romeiras, M.M.; Jiménez, A.; Conti, E. Geo-climatic changes and apomixis as major drivers of diversification in the Mediterranean sea lavenders (Limonium Mill.). Front. Plant Sci. 2021, 11, 2127. [Google Scholar] [CrossRef]
- Bacchetta, G.; Brullo, S.; Velari, T.C.; Chiapella, L.F.; Kosovel, V. Analysis of the Genista ephedroides group (Fabaceae) based on karyological, molecular and morphological data. Caryologia 2012, 65, 47–61. [Google Scholar] [CrossRef]
- Bacchetta, G.; Brullo, S.; Chiapella, L.F.; Velari, T.C.; Fenu, G.; Del Galdo, G.G. Taxonomic remarks on Genista salzmannii group (Fabaceae) in Sardinia and Corsica. Phytotaxa 2020, 449, 31–51. [Google Scholar] [CrossRef]
- Bacchetta, G.; Brullo, S.; Casti, M.; Giusso del Galdo, G.P. Taxonomic revision of the Dianthus sylvestris group (Caryophyllaceae) in central-southern Italy, Sicily and Sardinia. Nord. J. Bot. 2010, 28, 137–173. [Google Scholar] [CrossRef]
- Terlević, A.; Rešetnik, I. Inventory of the historical Dianthus sylvestris herbarium materials from Herbarium Croaticum and Herbarium Ivo and Marija Horvat. Glas. Hrvat. Bot. Druš. 2020, 8, 8–14. [Google Scholar] [CrossRef]
- Nebot, A.; Cogoni, D.; Fenu, G.; Bacchetta, G. Breeding system and inbreeding depression in a translocated population of the endangered plant Dianthus morisianus (Caryophyllaceae). Flora 2020, 262, 151488. [Google Scholar] [CrossRef]
- Valente, L.M.; Savolainen, V.; Vargas, P. Unparalleled rates of species diversification in Europe. Proc. R. Soc. B 2010, 277, 1489–1496. [Google Scholar] [CrossRef]
- Naciri, Y.; Du Pasquier, P.E.; Lundberg, M.; Jeanmonod, D.; Oxelman, B. A phylogenetic circumscription of Silene sect. Siphonomorpha (Caryophyllaceae) in the Mediterranean Basin. Taxon 2017, 66, 91–108. [Google Scholar] [CrossRef]
- Murru, V.; Grillo, O.; Santo, A.; Ucchesu, M.; Piazza, C.; Gaio, A.; Carta, A.; Bacchetta, G. Seed morpho-colorimetric analysis on some Tyrrhenian species of the Silene mollissima aggregate (Caryophyllaceae). Flora 2019, 258, 151445. [Google Scholar] [CrossRef]
- Selvi, F.; Coppi, A.; Bigazzi, M. Karyotype variation, evolution and phylogeny in Borago (Boraginaceae), with emphasis on subgenus Buglossites in the Corso-Sardinian system. Ann. Bot. 2006, 98, 857–868. [Google Scholar] [CrossRef]
- Farris, E.; Orrù, M.; Ucchesu, M.; Amadori, A.; Porceddu, M.; Bacchetta, G. Morpho-Colorimetric characterization of the Sardinian endemic taxa of the genus Anchusa L. by seed image analysis. Plants 2020, 9, 1321. [Google Scholar] [CrossRef]
- Callmander, M.W.; Phillipson, P.B.; Schatz, G.E.; Andriambololonera, S.; Rabarimanarivo, M.; Rakotonirina, N.; Raharimampionona, J.; Chatelain, C.; Gautier, L.; Lowry, P.P. The endemic and non-endemic vascular flora of Madagascar updated. Plant Ecol. Evol. 2011, 144, 121–125. [Google Scholar] [CrossRef]
- Gentili, R.; Fenu, G.; Labra, M.; Bruni, I.; Mattana, E.; Bacchetta, G. Lamyropsis genus in the Mediterranean area: Phylogenetic position of L. microcephala (Asteraceae: Cardueae). Plant Biosyst. 2015, 149, 944–948. [Google Scholar] [CrossRef]
- Gentili, R.; Fenu, G.; Porceddu, M.; Bruni, I.; Citterio, S.; Bacchetta, G. Genetic variability of the first-generation of Ribes sardoum, a threatened relic plant requiring translocation measures. Plant Biosyst. 2019, 153, 1–4. [Google Scholar] [CrossRef]
- Bocchieri, E.; Iiriti, G. Biodiversità vegetale in sistemi microinsulari della Sicilia e della Sardegna. Biogeographia 2011, 30, 177–196. [Google Scholar] [CrossRef]
- Pisanu, S.; Mameli, G.; Farris, E.; Binelli, G.; Filigheddu, R. A natural homoploid hybrid between Centaurea horrida and Centaurea filiformis (Asteraceae) as revealed by morphological and genetic traits. Folia Geobot. 2011, 46, 69–86. [Google Scholar] [CrossRef]
- Pignatti, S. Plant geographical and morphological evidences in the evolution of the mediterranean flora (with particular reference to the Italian representatives). Webbia 1979, 34, 243–255. [Google Scholar] [CrossRef]
- Sciandrello, S.; Guarino, R.; Minissale, P.; Spampinato, G. The endemic vascular flora of Peloritani Mountains (NE Sicily): Plant functional traits and phytogeographical relationships in the most isolated and fragmentary micro-plate of the Alpine orogeny. Plant Biosyst. 2015, 149, 838–854. [Google Scholar] [CrossRef]
- De Castro, O.; Brullo, S.; Colombo, P.; Jury, S.; De Luca, P.; Di Maio, A. Phylogenetic and biogeographical inferences for Pancratium (Amaryllidaceae), with an emphasis on the Mediterranean species based on plastid sequence data. Bot. J. Linn. Soc. 2012, 170, 12–28. [Google Scholar] [CrossRef][Green Version]
- Meloni, M.; Dettori, C.A.; Reid, A.; Bacchetta, G.; Hugot, L.; Conti, E. High genetic diversity and presence of genetic structure characterise the endemics Ruta corsica and Ruta lamarmorae (Rutaceae). Caryologia 2020, 73, 11–26. [Google Scholar] [CrossRef]
- Conti, F.; Bartolucci, F.; Bacchetta, G.; Pennesi, R.; Lakušić, D.; Niketić, M. A taxonomic revision of the Siler montanum group (Apiaceae) in Italy and the Balkan Peninsula. Willdenowia 2021, 51, 321–347. [Google Scholar] [CrossRef]
- Bocchieri, E.; Satta, V. Flora and vegetal landscape on the island of Figarolo (NE Sardinia). Lagascalia 1999, 21, 17–46. [Google Scholar]
- Maccioni, A.; Canopoli, L.; Cubeddu, V.; Cucca, E.; Dessena, S.; Morittu, S.; Filigheddu, R.; Padedda, B.M.; Farris, E. Gradients of salinity and plant community richness and diversity in two different Mediterranean coastal ecosystems in NW Sardinia. Biodivers. Data J. 2021, 9, e71247. [Google Scholar] [CrossRef] [PubMed]
- Podda, L.; Meloni, F.; Fenu, G.; Iiriti, G.; Bacchetta, G. The vascular flora of the Marine Protected Area of “Capo Carbonara” (SE-Sardinia). Fl. Medit. 2021, 31, 415–449. [Google Scholar] [CrossRef]
- Fois, M.; Fenu, G.; Bacchetta, G. Identifying and assessing the efficiency of different networks of a fine-scale hierarchy of biodiversity hotspots. Plant Ecol. Divers. 2018, 11, 121–132. [Google Scholar] [CrossRef]
- Fenu, G.; Mattana, E.; Congiu, A.; Bacchetta, G. The endemic vascular flora of Supramontes (Sardinia), a priority plant conservation area. Candollea 2010, 65, 347–358. [Google Scholar] [CrossRef]
- Camarda, I.; Brundu, G.; Carta, L.; Vacca, G.; Brunu, A. Plants and grazing: An evaluation of the effects on Sardinian endemic plants conservation. Flora Medit. 2015, 25, 263–275. [Google Scholar] [CrossRef]
- Fenu, G.; Cogoni, D.; Navarro, F.; Concas, E.; Bacchetta, G. The importance of the Cisto-Lavanduletalia coastal habitat on population persistence of the narrow endemic Dianthus morisianus (Caryophyllaceae). Plant Species Biol. 2017, 32, 156–168. [Google Scholar] [CrossRef]
- Bagella, S.; Filigheddu, R.; Benesperi, R.; Giordani, P.; Minuto, L.; Viciani, D.; Caria, M.C.; Pisanu, S.; Casazza, G. Thorn, spine and prickle patterns in the Italian flora. Plant Biosyst. 2019, 153, 118–133. [Google Scholar] [CrossRef]
- Adamson, K.R.; Hughes, P.D.; Woodward, J.C. Pleistocene glaciation of the Mediterranean mountains. Quat. Newsl. 2013, 131, 2–15. [Google Scholar]
- Mazza, P.P.; Lovari, S.; Masini, F.; Masseti, M.; Rustioni, M. A multidisciplinary approach to the analysis of multifactorial land mammal colonization of islands. BioScience 2013, 63, 939–951. [Google Scholar] [CrossRef]
- Carta, A.; Bedini, G.; Guidi, T.; Foggi, B. Tuscan Archipelago flora: From genesis to conservation. In Proceedings of the Second Botanical Conference in Menorca “Islands and Plants: Preservation and understanding of flora on Mediterranean islands”, Menorca, Spain, 26–30 April 2011; Cardona Pons, E., Estaun Clarisó, I., Comas Casademont, M., Fraga i Arguimbau, P., Eds.; Menorquí d’Estudis, Consell Insular de Menorca: Maó, Spain, 2013; pp. 157–166. [Google Scholar]
- Bacchetta, G.; Fenu, G.; Guarino, R.; Mandis, G.; Mattana, E.; Nieddu, G.; Scudu, C. Floristic traits and biogeographic characterization of the Gennargentu massif (Sardinia). Candollea 2013, 68, 209–220. [Google Scholar] [CrossRef]
- Bacchetta, G.; Iiriti, G.; Mossa, L. La flora endemica del Sarrabus-Gerrei: Un patrimonio da tutelare e gestire. In Analisi e Sistemi di Gestione del Territorio (Sarrabus-Gerrei); Artizzu, D., Ed.; Comune di Sinnai: Sinnai, Italy, 2005; pp. 105–112. [Google Scholar]
- Calvia, G.; Ruggero, A. The vascular flora of Mount Limbara (northern Sardinia): From a troubled past towards an uncertain future. Fl. Medit. 2020, 30, 293–313. [Google Scholar] [CrossRef]
- Arrigoni, P.V. Rapporti floristici tra l’Arcipelago Toscano e le terre vicine. Lav. Soc. Ital. Biogeogr. 1975, 5, 55–65. [Google Scholar] [CrossRef]
- Arrigoni, P.V. Aspetti corologici della flora sarda. Biogeographia 1983, 8, 65. [Google Scholar] [CrossRef]
- Dapporto, L.; Palagi, E.; Turillazzi, S. Cuticular Hydrocarbons of Polistes dominulus as a Biogeographic Tool: A Study of Populations from the Tuscan Archipelago and Surrounding Areas. J. Chem. Ecol. 2004, 30, 2139–2151. [Google Scholar] [CrossRef]
- Schmitt, T.; Fritz, U.; Delfino, M.; Ulrich, W.; Habel, J.C. Biogeography of Italy revisited: Genetic lineages confirm major phylogeographic patterns and a pre-Pleistocene origin of its biota. Front. Zool. 2021, 18, 34. [Google Scholar] [CrossRef]
- Contandriopoulos, J. Recherches sur la flore endémique de la Corse et sur ses origines. Ann. Fac. Sci. Marseille 1962, 32, 1–354. [Google Scholar]
- Médail, F.; Verlaque, R. Ecological characteristics and rarity of endemic plants from southeast France and Corsica: Implications for biodiversity conservation. Biol. Conserv. 1997, 80, 269–281. [Google Scholar] [CrossRef]
- Hamel, T.; Boulemtafes, A. Decouverte d’une endemique tyrrhenienne Soleirolia soleirolii (Urticaceae) en Algerie (Afrique du Nord). Fl. Medit. 2017, 27, 185–193. [Google Scholar] [CrossRef]
- Bobo-Pinilla, J.; de León, S.B.B.; Colomar, J.S.; Fenu, G.; Bacchetta, G.; de Giles, J.P.; Martínez-Ortega, M.M. Phylogeography of Arenaria balearica L. (Caryophyllaceae): Evolutionary history of a disjunct endemic from the Western Mediterranean continental islands. PeerJ 2016, 4, e2618. [Google Scholar] [CrossRef] [PubMed]
- Bocquet, G. The Messinian model: A new outlook for the floristics and systematics of the Mediterranean area. Candollea 1978, 33, 269–287. [Google Scholar]
- Rosselló, J.A.; Cosín, R.; Bacchetta, G.; Brullo, S.; Mayol, M. Nuclear and chloroplast DNA variation in Cephalaria squamiflora (Dipsacaceae), a disjunct Mediterranean species. Taxon 2009, 58, 1242–1253. [Google Scholar] [CrossRef]
- Fenu, G.; Mattana, E.; Bacchetta, G. Distribution, status and conservation of a Critically Endangered, extremely narrow endemic: Lamyropsis microcephala (Asteraceae) in Sardinia. Oryx 2011, 45, 180–186. [Google Scholar] [CrossRef]
- Garrido, J.L.; Fenu, G.; Mattana, E.; Bacchetta, G. Spatial genetic structure of Aquilegia taxa endemic to the island of Sardinia. Ann. Bot. 2012, 109, 953–964. [Google Scholar] [CrossRef]
- Cogoni, D.; Sulis, E.; Bacchetta, G.; Fenu, G. The unpredictable fate of the single population of a threatened narrow endemic Mediterranean plant. Biodivers. Conserv. 2019, 28, 1799–1813. [Google Scholar] [CrossRef]
- Murru, V.; Farris, E.; Santo, A.; Grillo, O.; Piazza, C.; Gaio, A.; Bacchetta, G.; Thompson, J.D. Niche Differentiation at Multiple Spatial Scales on Large and Small Mediterranean Islands for the Endemic Silene velutina Pourr. ex Loisel. (Caryophyllaceae) Plants 2021, 10, 2298. [Google Scholar] [CrossRef]
- Porceddu, M.; Fenu, G.; Bacchetta, G. New findings on seed ecology of Ribes sardoum: Can it provide a new opportunity to prevent the extinction of a threatened plant species? Syst. Biodivers. 2017, 15, 480–488. [Google Scholar] [CrossRef]
- Porceddu, M.; Santo, A.; Orru, M.; Meloni, F.; Ucchesu, M.; Picciau, R.; Sarigu, M.; Cuena-Lombraña, A.; Podda, L.; Sau, S.; et al. Seed conservation actions for the preservation of plant diversity: The case of the Sardinian Germplasm Bank (BG-SAR). Plant Sociol. 2017, 54, 111–117. [Google Scholar]
- Fenu, G.; Bacchetta, G.; Christodoulou, C.S.; Cogoni, D.; Fournaraki, C.; Giusso del Galdo, G.; Gotsiou, P.; Kyratzis, A.; Piazza, C.; Vicens, M.; et al. A common approach to the conservation of threatened island vascular plants: First results in the Mediterranean basin. Diversity 2020, 12, 157. [Google Scholar] [CrossRef]
- Médail, F.; Baumel, A. Using phylogeography to define conservation priorities: The case of narrow endemic plants in the Med-578 iterranean Basin hotspot. Biol. Conserv. 2018, 224, 258–266. [Google Scholar] [CrossRef]
- Bobo-Pinilla, J.; Salmerón-Sánchez, E.; Mendoza-Fernández, A.J.; Mota, J.F.; Peñas, J. Conservation and phylogeography of plants: From the Mediterranean to the rest of the world. Diversity 2022, 14, 78. [Google Scholar] [CrossRef]
- Gauthier, P.; Debussche, M.; Thompson, J.D. Regional priority setting for rare species based on a method combining three criteria. Biol. Conserv. 2010, 143, 1501–1509. [Google Scholar] [CrossRef]
- Carmignani, L.; Oggiano, G.; Funedda, A.; Conti, P.; Pasci, S. The geological map of Sardinia (Italy) at 1: 250,000 scale. J. Maps 2016, 12, 826–835. [Google Scholar] [CrossRef]
- Bacchetta, G. Flora vascolare del Sulcis (Sardegna sud-occidentale, Italia). Guineana 2006, 12, 1–369. [Google Scholar]
- Bacchetta, G.; Bagella, S.; Biondi, E.; Farris, E.; Filigheddu, R.; Mossa, L. Vegetazione forestale e serie di vegetazione della Sardegna (con rappresentazione cartografica alla scala 1:350.000). Fitosociologia 2009, 46, 3–82. [Google Scholar]
- Canu, S.; Rosati, L.; Fiori, M.; Motroni, A.; Filigheddu, R.; Farris, E. Bioclimate map of Sardinia (Italy). J. Maps 2015, 11, 711–718. [Google Scholar] [CrossRef]
- Danin, A.; Buldrini, F.; Bandini Mazzanti, M.; Bosi, G.; Caria, M.C.; Dandria, D.; Lanfranco, E.; Mifsud, S.; Bagella, S. Diversification of Portulaca oleracea L. complex in the Italian peninsula and adjacent islands. Bot. Lett. 2016, 163, 261–272. [Google Scholar] [CrossRef]
- Rosenbaum, G.; Lister, G.S.; Duboz, C. Reconstruction of the tectonic evolution of the western Mediterranean since the Oligocene. J. Virtual Explor. 2002, 8, 107–130. [Google Scholar] [CrossRef]
- Favre, E.; François, L.; Fluteau, F.; Cheddadi, R.; Thévenod, L.; Suc, J.P. Messinian vegetation maps of the Mediterranean region using models and interpolated pollen data. Geobios 2007, 40, 433–443. [Google Scholar] [CrossRef]
- Hsü, K.J.; Montadert, L.; Bernoulli, D.; Cita, M.B.; Erickson, A.; Garrison, R.E.; Kidd, R.B.; Melieres, F.; Muller, C.; Wright, R. History of Mediterranean salinity crisis. Nature 1977, 267, 399–403. [Google Scholar] [CrossRef]
- Cherchi, A.; Montadert, L. Oligo-Miocene rift of Sardinia and the history of the Western Mediterranean Basin. Nature 1982, 298, 736–739. [Google Scholar] [CrossRef]
- Fiz-Palacios, O.; Vargas, P.; Vila, R.; Papadopulos, A.S.; Aldasoro, J.J. The uneven phylogeny and biogeography of Erodium (Geraniaceae): Radiations in the Mediterranean and recent recurrent intercontinental colonization. Ann. Bot. 2010, 106, 871–884. [Google Scholar] [CrossRef]
- De Castro, O.; Innangi, M.; Di Maio, A.; Menale, B.; Bacchetta, G.; Pires, M.; Noble, V.; Gestri, G.; Conti, F.; Peruzzi, L. Disentangling phylogenetic relationships in a hotspot of diversity: The butterworts (Pinguicula L., Lentibulariaceae) endemic to Italy. PLoS ONE 2016, 11, e0167610. [Google Scholar] [CrossRef] [PubMed]
- Míguez, M.; Gehrke, B.; Maguilla, E.; Jiménez-Mejías, P.; Martín-Bravo, S. Carex sect. Rhynchocystis (Cyperaceae): A Miocene subtropical relict in the Western Palaearctic showing a dispersal-derived Rand Flora pattern. J. Biogeogr. 2017, 44, 2211–2224. [Google Scholar] [CrossRef]
- Mossa, L.; Bacchetta, G.; Brullo, S. Quercus ichnusae (Fagaceae), a new species from Sardinia. Isr. J. Plant Sci. 1999, 47, 199–207. [Google Scholar] [CrossRef]
- Fineschi, S.; Taurchini, D.; Grossoni, P.; Petit, R.J.; Vendramin, G.G. Chloroplast DNA variation of white oaks in Italy. For. Ecol. Manag. 2002, 156, 103–114. [Google Scholar] [CrossRef]
- Petit, R.J.; Brewer, S.; Bordács, S.; Burg, K.; Cheddadi, R.; Coart, E.; Coart, E.; Cottrell, J.; Csaikl, U.M.; van Damh, B.; et al. Identification of refugia and post-glacial colonisation routes of European white oaks based on chloroplast DNA and fossil pollen evidence. For. Ecol. Manag. 2002, 156, 49–74. [Google Scholar] [CrossRef]
- Castroviejo, S. Flora Iberica; Real Jardín Botánico-CSIC: Madrid, Spain, 1986–2019; Volume 1–18, 21. [Google Scholar]
- Giardina, G.; Raimondo, F.M.; Spadaro, V. A catalogue of plants growing in Sicily. Bocconea 2007, 20, 5–582. [Google Scholar]
- Véla, E.; Pavon, D. The vascular flora of Algerian and Tunisian small islands: If not biodiversity hotspots, at least biodiversity hotchpotchs. Biodivers. J. 2012, 3, 343–362. [Google Scholar]
- Jeanmonod, D.; Gamisans, J. Flora Corsica; Édisud: Aix-en-Provence, France, 2007. [Google Scholar]
- Pignatti, S.; Guarino, R.; La Rosa, M. Flora d’Italia, 2nd ed.; Edagricole: Bologna, Italy, 2017–2019; Volume 1–4. [Google Scholar]
- Bartolucci, F.; Domina, G.; Argenti, C.; Bacchetta, G.; Ballelli, S.; Banfi, E.; Barberis, D.; Barberis, G.; Bertolli, A.; Bolpagni, R.; et al. Notulae to the Italian native vascular flora. Ital. Bot. 2021, 12, 85–103. [Google Scholar] [CrossRef]
- Euro+Med. Euro+Med PlantBase—The Information Resource for Euro-Mediterranean Plant Diversity. 2006–2020. Available online: http://ww2.bgbm.org/EuroPlusMed/ (accessed on 16 June 2020).
- Plants of the World Online POWO. Facilitated by the Royal Botanic Gardens, Kew. Available online: http://www.plantsoftheworldonline.org/2020 (accessed on 11 February 2021).
- Martellos, S.; Bartolucci, F.; Conti, F.; Galasso, G.; Moro, A.; Pennesi, R.; Peruzzi, L.; Pittao, E.; Nimis, P.L. FlorItaly—The portal to the Flora of Italy. PhytoKeys 2020, 156, 55–71. [Google Scholar] [CrossRef] [PubMed]
- ANTHOS: Sistema de Información sobre las Plantas de España. Available online: www.programanthos.org (accessed on 21 March 2021).
- Inventaire National du Patrimoine Naturel—INPN. Available online: https://inpn.mnhn.fr/ (accessed on 10 April 2021).
- Bartolucci, F.; Peruzzi, L.; Galasso, G.; Albano, A.; Alessandrini, A.; Ardenghi, N.M.G.; Astuti, G.; Bacchetta, G.; Ballelli, S.; Banfi, E.; et al. An updated checklist of the vascular flora native to Italy. Plant Biosyst. 2018, 152, 179–303. [Google Scholar] [CrossRef]
- Bacchetta, G.; Casti, M.; Mossa, L. New ecological and distributive data on the rupestrian flora of Sardinia. J. Bot. Soc. Bot. Fr. 2007, 38, 73–83. [Google Scholar]
- Bocchieri, E.; Iiriti, G. Contribution to knowledge of the endemic vascular flora of the capes and promontories of Sardinia (Italy). Bocconea 2009, 23, 353. [Google Scholar]
- IUCN. The IUCN Red List of Threatened Species. Version 2021-3. 2021. Available online: https://www.iucnredlist.org (accessed on 20 November 2021).
- Rossi, G.; Montagnani, C.; Abeli, T.; Gargano, D.; Peruzzi, L.; Fenu, G.; Wagensommer, R.P.; Ravera, S.; Cogoni, A.; Aleffi, M.; et al. Are Red Lists really useful for plant conservation? The New Red List of the Italian Flora in the perspective of national conservation policies. Plant Biosyst. 2014, 148, 187–190. [Google Scholar] [CrossRef]
- Fenu, G.; Bacchetta, G.; Giacanelli, V.; Gargano, D.; Montagnani, C.; Orsenigo, S.; Cogoni, D.; Rossi, G.; Conti, F.; Santangelo, A.; et al. Conserving plant diversity in Europe: Outcomes, criticisms and perspectives of the Habitats Directive application in Italy. Biodivers. Conserv. 2017, 26, 309–328. [Google Scholar] [CrossRef]
- Fois, M.; Bacchetta, G.; Cogoni, D.; Fenu, G. Current and future effectiveness of the Natura 2000 network for protecting plant species in Sardinia: A nice and complex strategy in its raw state? J. Environ. Plan. Manag. 2018, 61, 332–347. [Google Scholar] [CrossRef]
- Fois, M.; Bacchetta, G.; Caria, M.C.; Cogoni, D.; Farris, E.; Fenu, G.; Manca, M.; Pinna, M.S.; Pisanu, S.; Rivieccio, G.; et al. Proposals for improvement of Annex I of Directive 92/43/EEC: Sardinia. Plant Sociol. 2021, 58, 65–76. [Google Scholar] [CrossRef]
- Cuena-Lombraña, A.; Fois, M.; Cogoni, A.; Bacchetta, G. Where we Come from and where to Go: Six Decades of Botanical Studies in the Mediterranean Wetlands, with Sardinia (Italy) as a Case Study. Wetlands 2021, 41, 69. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Family | No. Taxa | No. inquirenda | Most Representative Genera (No. Taxa) |
|---|---|---|---|
| Asteraceae | 49 (20%) | 19 | Hieracium (7), Taraxacum (6) |
| Plumbaginaceae | 42 (75%) | 12 | Limonium (38), Armeria (4) |
| Fabaceae | 27 (12%) | 0 | Genista (18), Astragalus (7) |
| Caryophyllaceae | 26 (21%) | 1 | Silene (9), Dianthus (9) |
| Lamiaceae | 16 (22%) | 3 | Stachys (3), Micromeria (2), Mentha (2), Teucrium (2) |
| Boraginaceae | 13 (28%) | 2 | Anchusa (7), Borago (2) |
| Orchidaceae | 13 (20%) | 4 | Ophrys (9), Orchis (2), Serapias (2) |
| Apiaceae | 12 (13%) | 2 | Seseli (2), Siler (2) |
| Poaceae | 12 (6%) | 2 | Sesleria (3), Festuca (2), Trisetaria (2) |
| Ranunculaceae | 12 (19%) | 1 | Ranunculus (5), Aquilegia (4) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fois, M.; Farris, E.; Calvia, G.; Campus, G.; Fenu, G.; Porceddu, M.; Bacchetta, G. The Endemic Vascular Flora of Sardinia: A Dynamic Checklist with an Overview of Biogeography and Conservation Status. Plants 2022, 11, 601. https://doi.org/10.3390/plants11050601
Fois M, Farris E, Calvia G, Campus G, Fenu G, Porceddu M, Bacchetta G. The Endemic Vascular Flora of Sardinia: A Dynamic Checklist with an Overview of Biogeography and Conservation Status. Plants. 2022; 11(5):601. https://doi.org/10.3390/plants11050601
Chicago/Turabian StyleFois, Mauro, Emmanuele Farris, Giacomo Calvia, Giuliano Campus, Giuseppe Fenu, Marco Porceddu, and Gianluigi Bacchetta. 2022. "The Endemic Vascular Flora of Sardinia: A Dynamic Checklist with an Overview of Biogeography and Conservation Status" Plants 11, no. 5: 601. https://doi.org/10.3390/plants11050601
APA StyleFois, M., Farris, E., Calvia, G., Campus, G., Fenu, G., Porceddu, M., & Bacchetta, G. (2022). The Endemic Vascular Flora of Sardinia: A Dynamic Checklist with an Overview of Biogeography and Conservation Status. Plants, 11(5), 601. https://doi.org/10.3390/plants11050601