
Transgene Flow: Challenges to the On-Farm Conservation of Maize Landraces in the Brazilian Semi-Arid Region

 ,
,  , , and
, , and
Abstract
1. Introduction
2. Results
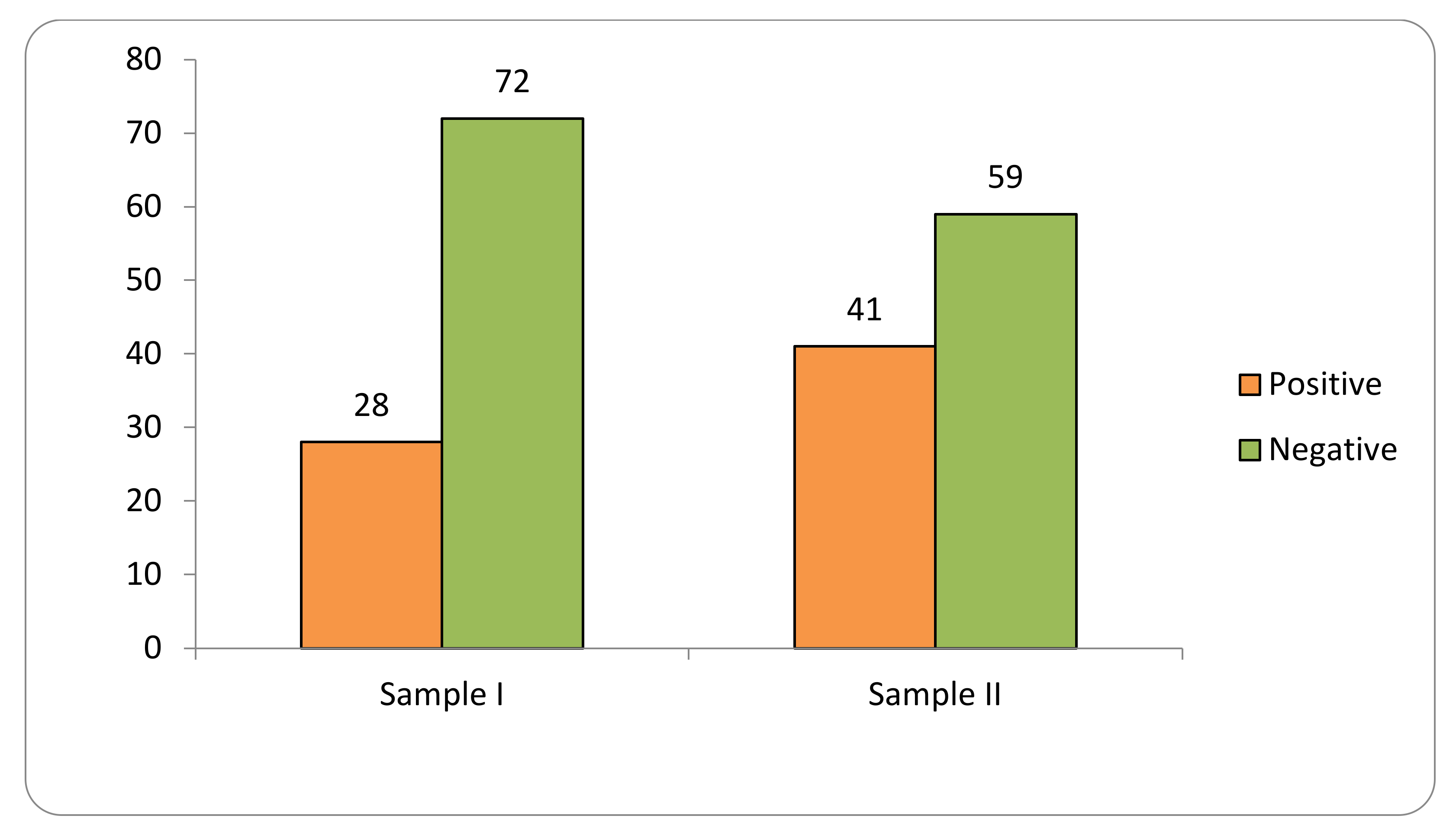
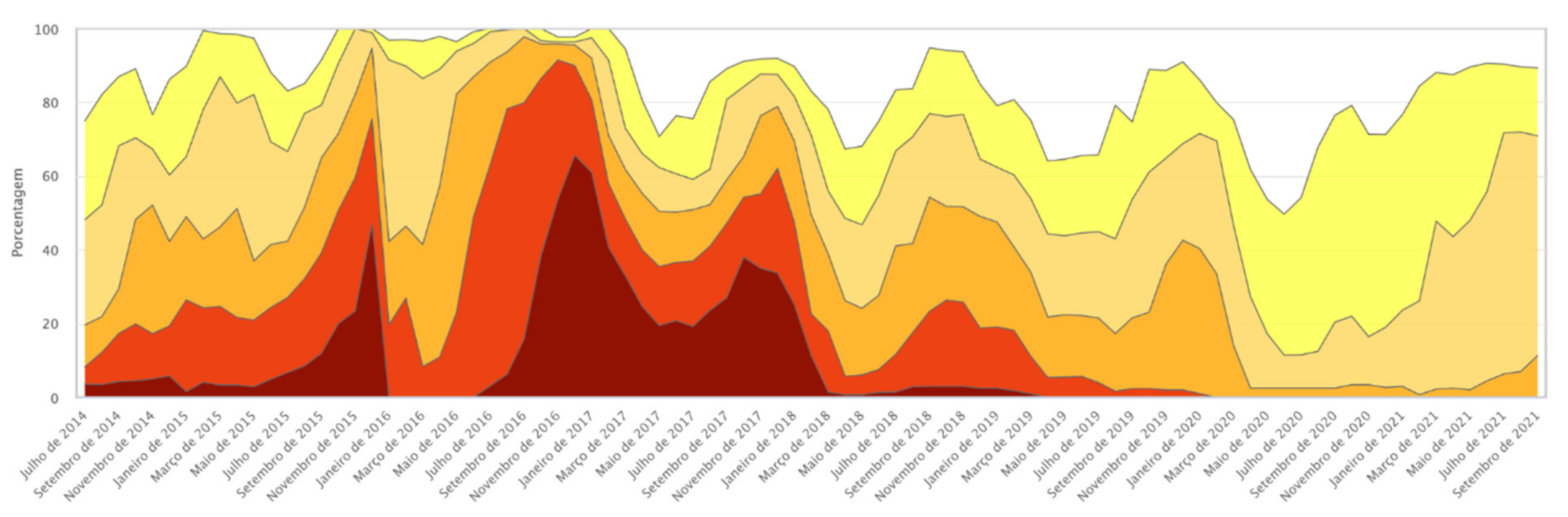
2.1. Transgene Flow Frequency
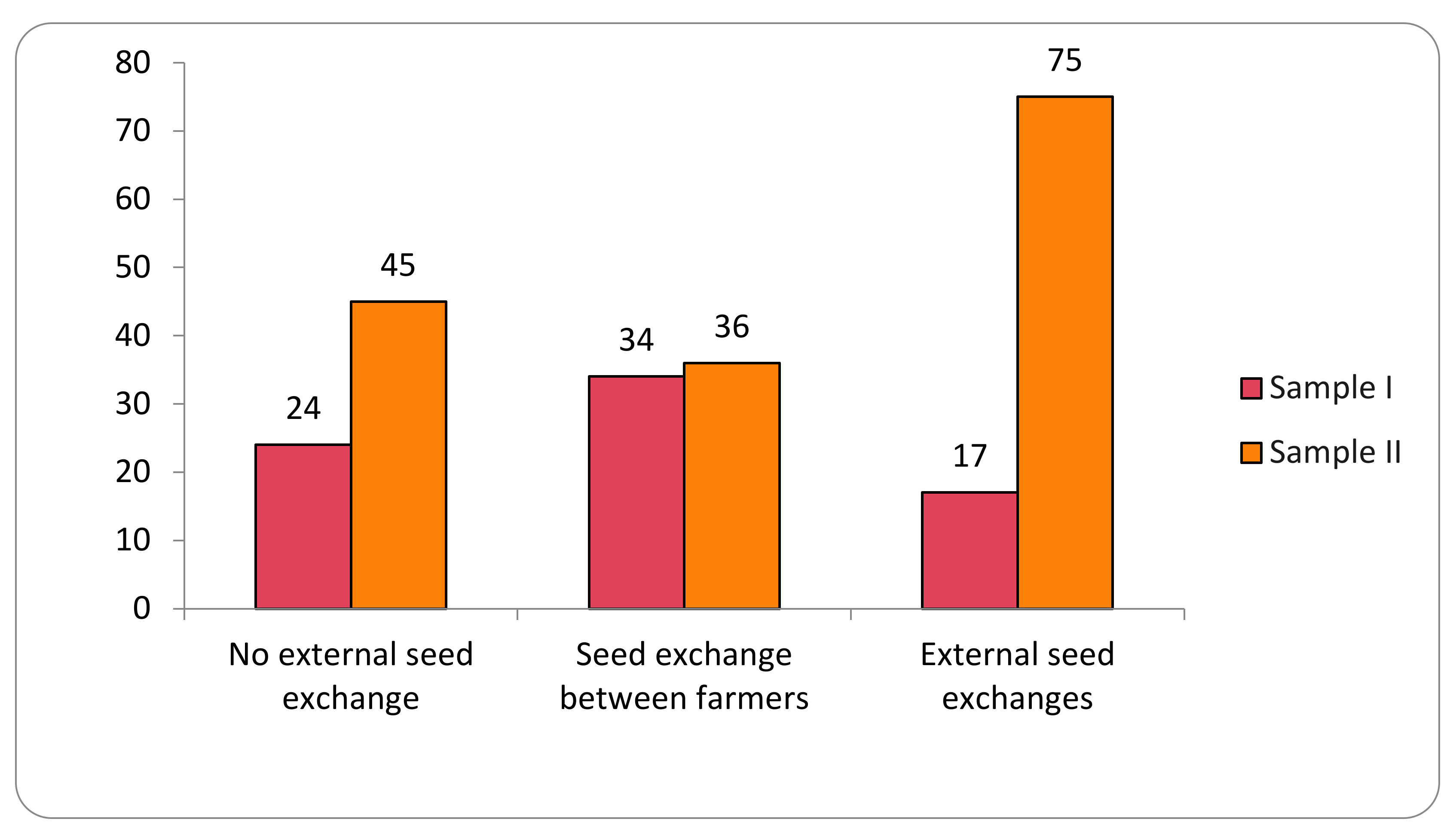
2.2. Transgene Flow According to Seed-Management System
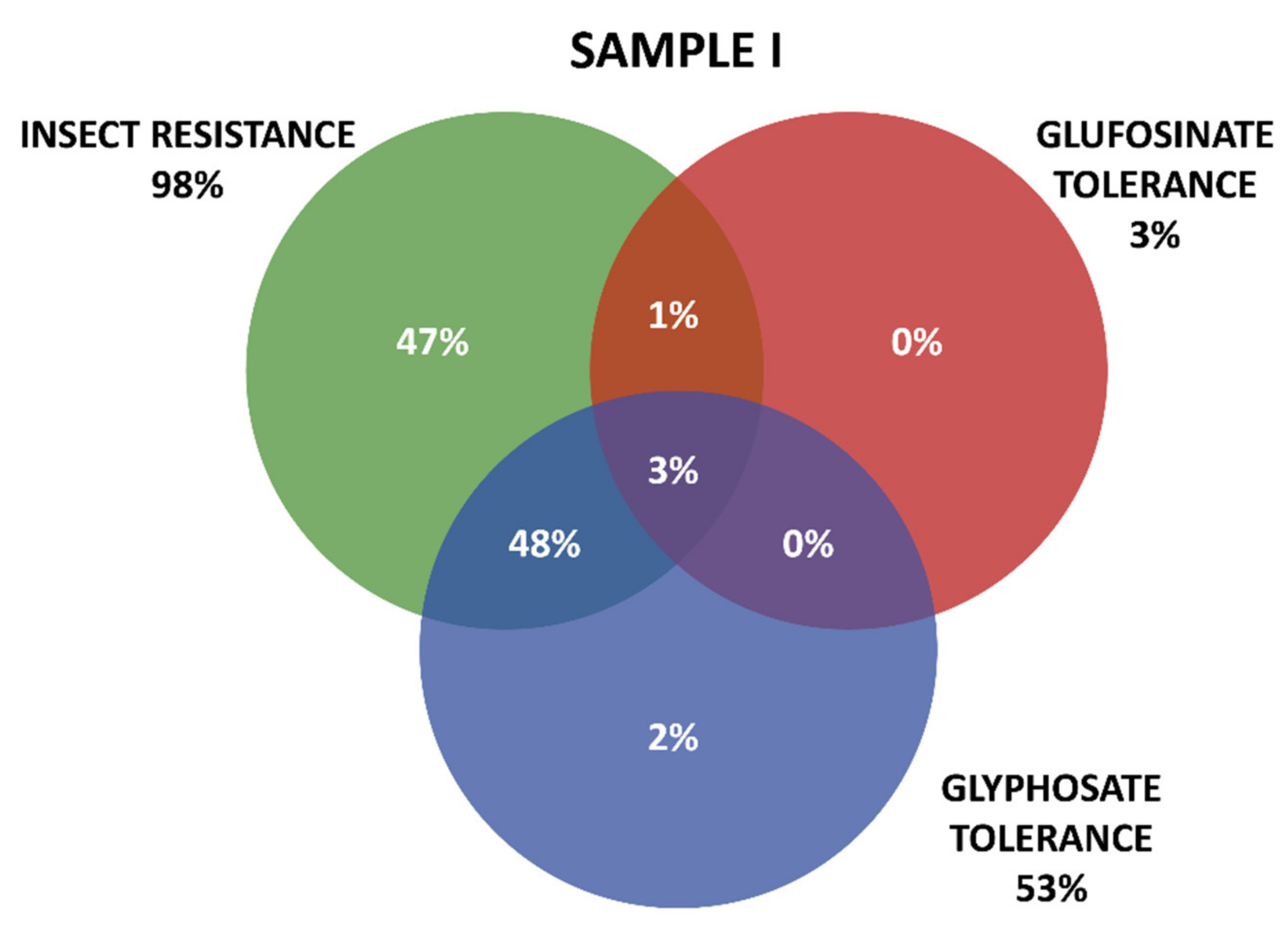
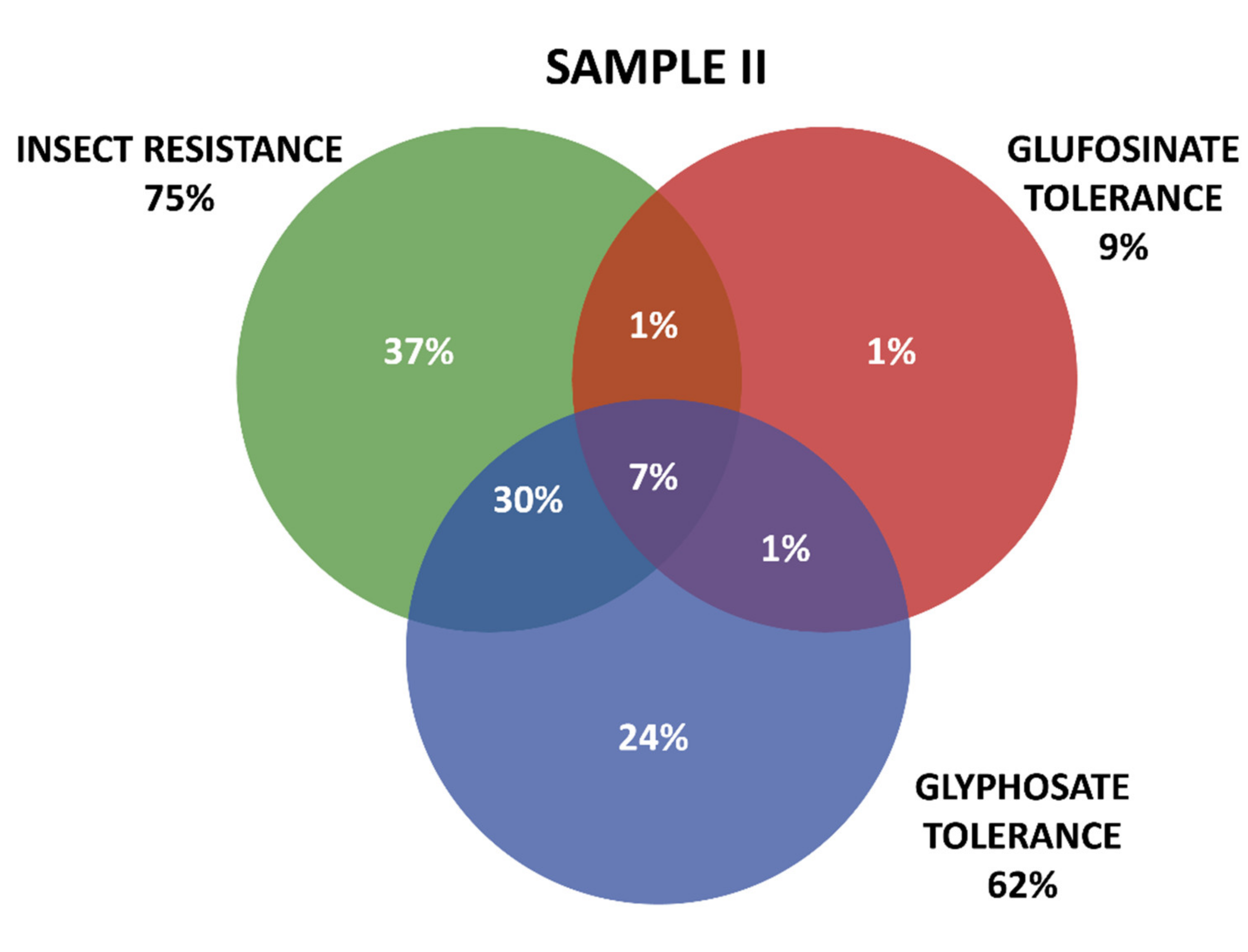
2.3. GM Proteins Detected
Grouping of GM Proteins by Mode of Action (Insect Resistance and Herbicide Tolerance)
3. Discussion
3.1. Unregulated Expansion of GMO Crops and Potential Effects of Transgene Flow into Maize Landraces Conserved by Family Farmers in the SAB
Decreased Drought and Increased Transgene Flow
3.2. Immunochromatographic Tests as a Tool for Local Transgene Flow Monitoring
3.3. Implications of GMO De-Regulation for on-Farm Conservation of Maize Genetic Diversity
4. Materials and Methods
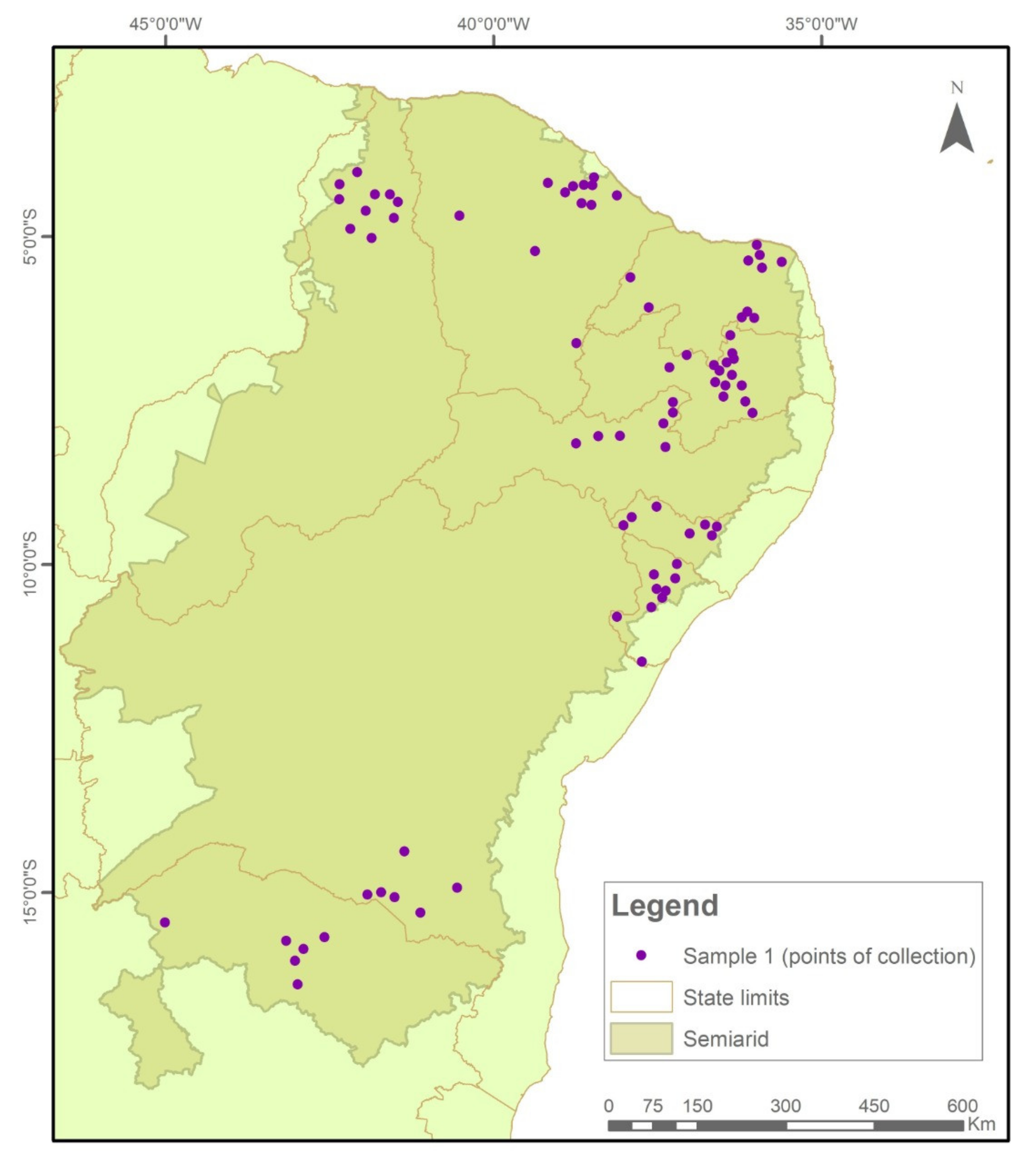
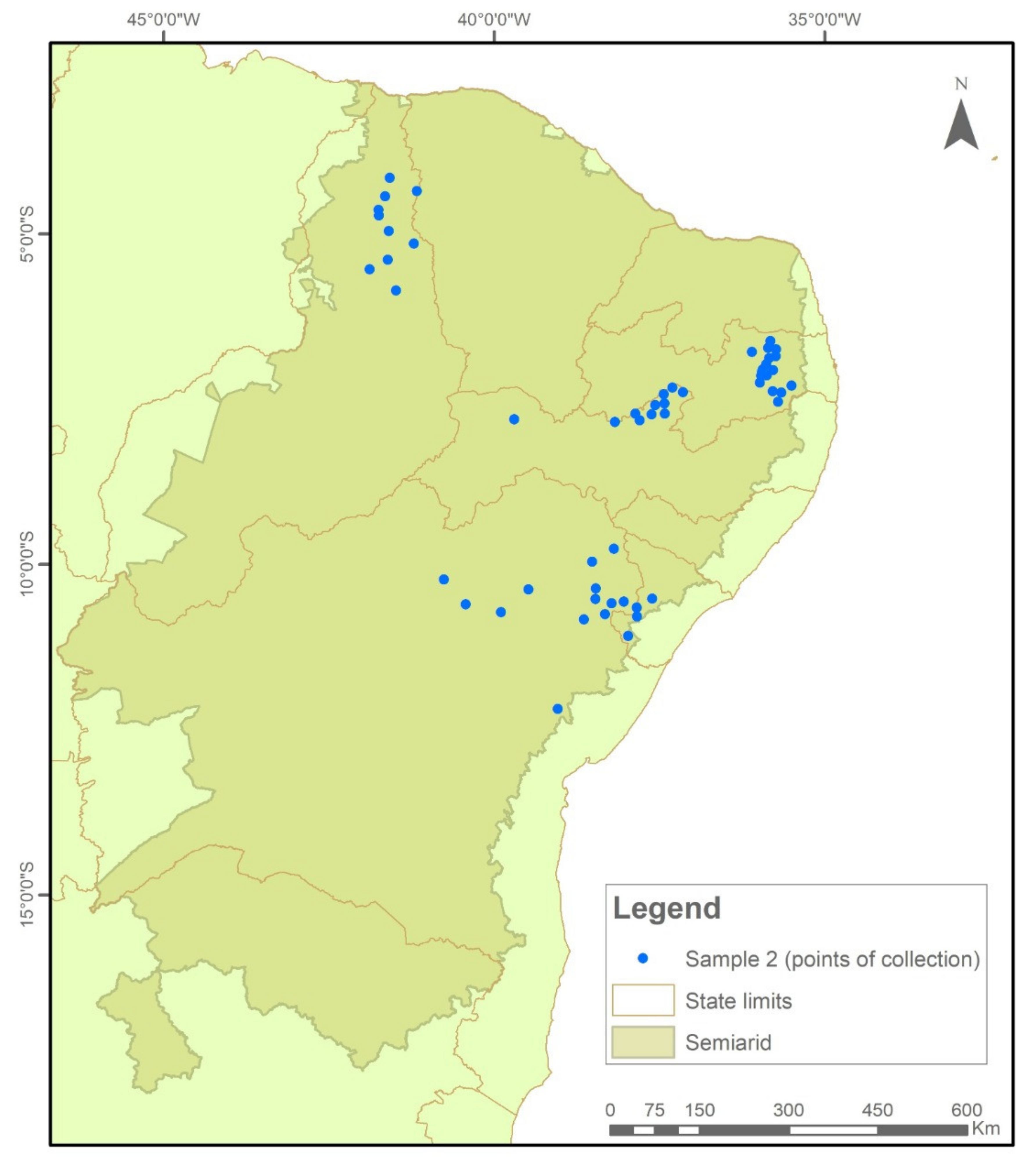
4.1. Study Area and Sample Collection Points
4.2. Collections and Tests Performed
4.3. Data Analysis
- (a)
- Frequency of positive seed lot of farmers’ landraces in immunochromatographic tests carried out in the two collection cycles 2018–2019 and 2020–2021;
- (b)
- Frequency of positive results in seeds according to the seed-management system in the two collection cycles 2018–2019 and 2020–2021;
- (c)
- Frequency of occurrence of the different proteins tested in seeds from varieties positive for transgenic events.
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Ellstrand, N.C. When Transgenes Wander, Should We Worry? Plant Physiol. 2001, 125, 1543–1545. [Google Scholar] [CrossRef] [PubMed]
- Gepts, P.; Papa, R. Possible effects of (trans)gene flow from crops on the genetic diversity from landraces and wild relatives. Environ. Biosaf. Res. 2003, 2, 89–103. [Google Scholar] [CrossRef] [PubMed]
- Altieri, M.A. The Myth of Coexistence: Why Transgenic Crops Are Not Compatible With Agroecologically Based Systems of Production. Bull. Sci. Technol. Soc. 2005, 25, 361–371. [Google Scholar] [CrossRef]
- McAfee, K. Corn Culture and Dangerous DNA: Real and Imagined Consequences of Maize Transgene Flow in Oaxaca. J. Lat. Am. Geogr. 2003, 2, 18–42. [Google Scholar] [CrossRef]
- Binimelis, R. Coexistence of Plants and Coexistence of Farmers: Is an Individual Choice Possible? J. Agric. Environ. Ethic 2008, 21, 437–457. [Google Scholar] [CrossRef]
- Levidow, L.; Boschert, K. Coexistence or contradiction? GM crops versus alternative agricultures in Europe. Geoforum 2008, 39, 174–190. [Google Scholar] [CrossRef]
- Heinemann, J.A. Typology of the Effects of (Trans)Gene Flow on the Conservation and Sustainable Use of Genetic Resources; Background study paper; 35; FAO Commission on Genetic Resources for Food and Agriculture Assessments: Rome, Italy, 2007; p. 94. [Google Scholar]
- Vázquez-Barrios, V.; Boege, K.; Sosa-Fuentes, T.G.; Rojas, P.; Wegier, A. Ongoing ecological and evolutionary consequences by the presence of transgenes in a wild cotton population. Sci. Rep. 2021, 11, 1–10. [Google Scholar] [CrossRef]
- FAO. International Treaty on Plant Genetic Resources for Food and Agriculture. 2009. Available online: https://www.fao.org/plant-treaty/en/ (accessed on 28 January 2022).
- De Schutter, O. Seed Policies and the Right to Food: Enhancing Agrobiodiversity, Encouraging Innovation Presented at the 64th Session of the UN General Assembly; Report submitted by the Special Rapporteur on the right to food A/64/170; UN General Assembly: New York, NY, USA, 2009; p. 30. [Google Scholar]
- Agapito-Tenfen, S.; Lopez, F.R.; Mallah, N.; Abou-Slemayne, G.; Trtikova, M.; Nodari, R.; Wickson, F. Transgene flow in Mexican maize revisited: Socio-biological analysis across two contrasting farmer communities and seed management systems. Ecol. Evol. 2017, 7, 9461–9472. [Google Scholar] [CrossRef]
- Quist, D.; Chapela, I.H. Transgenic DNA introgressed into traditional maize landraces in Oaxaca, Mexico. Nature 2001, 414, 541–543. [Google Scholar] [CrossRef]
- Galeano, P.; Debat, C.M.; Ruibal, F.; Fraguas, L.F.; Galván, G.A. Cross-fertilization between genetically modified and non-genetically modified maize crops in Uruguay. Environ. Biosaf. Res. 2010, 9, 147–154. [Google Scholar] [CrossRef][Green Version]
- Piñeyro-Nelson, A.; Van Heerwaarden, J.; Perales, H.R.; Serratos-Hernández, J.A.; Rangel, A.; Hufford, M.B.; Gepts, P.; Garay-Arroyo, A.; Rivera-Bustamante, R.; Álvarez-Buylla, E.R. Transgenes in Mexican maize: Molecular evidence and methodological considerations for GMO detection in landrace populations. Mol. Ecol. 2009, 18, 750–761. [Google Scholar] [CrossRef] [PubMed]
- Santa-Maria, M.C.; Lajo-Morgan, G.; Guardia, L. Adventitious presence of transgenic events in the maize supply chain in Peru: A case study. Food Control 2014, 41, 96–101. [Google Scholar] [CrossRef]
- Knispel, A.L.; McLachlan, S.M.; Van Acker, R.C.; Friesen, L.F. Gene Flow and Multiple Herbicide Resistance in Escaped Canola Populations. Weed Sci. 2008, 56, 72–80. [Google Scholar] [CrossRef]
- Schafer, M.G.; Ross, A.A.; Londo, J.; Burdick, C.A.; Lee, E.H.; Travers, S.E.; Van De Water, P.K.; Sagers, C.L. The Establishment of Genetically Engineered Canola Populations in the U.S. PLoS ONE 2011, 6, e25736. [Google Scholar] [CrossRef]
- Ferment, G.; Melgarejo, L.; Fernandes, G.B.; Ferraz, J.M.G. Transgenic Crops–Hazards and Uncertainties: More than 750 Studies Disregarded by the GMOs Regulatory Bodies, 1st ed.; Nead Debate; Nead/MDA: Brasília, Brazil, 2017. [Google Scholar]
- Silva, N.C.d.A.; Costa, F.M.; Vidal, R.; Veasey, E.A. Maíces de Las Tierras Bajas de América Del Sur y Conservación de La Agrobiodiversidad En Brasil y Uruguay, 1st ed.; Atena Editora: Ponta Grossa, Brazil, 2020. [Google Scholar] [CrossRef]
- Kistler, L.; Maezumi, S.Y.; de Souza, J.G.; Przelomska, N.A.S.; Costa, F.M.; Smith, O.; Loiselle, H.; Ramos-Madrigal, J.; Wales, N.; Ribeiro, E.R.; et al. Multiproxy evidence highlights a complex evolutionary legacy of maize in South America. Science 2018, 362, 1309–1313. [Google Scholar] [CrossRef]
- Costa, F.M.; Silva, N.C.D.A.; Vidal, R.; Clement, C.R.; Alves, R.P.; Bianchini, P.C.; Haverroth, M.; Freitas, F.D.O.; Veasey, E.A. Entrelaçado, a rare maize race conserved in Southwestern Amazonia. Genet. Resour. Crop Evol. 2021, 68, 51–58. [Google Scholar] [CrossRef]
- Carvalho, V.P.; Ruas, C.F.; Ferreira, J.M.; Moreira, R.M.; Ruas, P.M. Genetic diversity among maize (Zea mays L.) landraces assessed by RAPD markers. Genet. Mol. Biol. 2004, 27, 228–236. [Google Scholar] [CrossRef]
- Silva, N.C.D.A.; Vidal, R.; Costa, F.M.; Veasey, E.A. Clasificación de Las Razas de Maíz de Brasil y Uruguay: Enfoque Metodológico y Principales Resultados. In Maíces de las Tierras Bajas de América del Sur y Conservación de la Agrobiodiversidad en Brasil y Uruguay; Atena Editora: Ponta Grossa, Brazil, 2020; pp. 87–109. [Google Scholar] [CrossRef]
- Costa, F.M.; Silva, N.C.D.A.; Vidal, R.; Veasey, E.A. Micro-Centros de Diversidade Genética Do Milho Nas Terras Baixas Da América Do Sul. In Milhos das Terras Baixas da América do Sul e Conservação da Agrobiodiversidade no Brasil e no Uruguai; Atena Editora: Ponta Grossa, Brazil, 2020; pp. 109–123. [Google Scholar]
- Silva, N.C.D.A.; Vidal, R.; Ogliari, J.B. New popcorn races in a diversity microcenter of Zea mays L. in the Far West of Santa Catarina, Southern Brazil. Genet. Resour. Crop Evol. 2017, 64, 1191–1204. [Google Scholar] [CrossRef]
- Costa, F.M.; Silva, N.C.D.A.; Ogliari, J.B. Maize diversity in southern Brazil: Indication of a microcenter of Zea mays L. Genet. Resour. Crop Evol. 2017, 64, 681–700. [Google Scholar] [CrossRef]
- Silva, N.C.D.A.; Costa, F.M.; Vidal, R.; Veasey, E.A. Microcentros de Diversidad Genética Del Maíz En Las Tierras Bajas de América Del Sur. In Maíces de las Tierras Bajas de América del Sur y Conservación de la Agrobiodiversidad en Brasil y Uruguay; Atena Editora: Ponta Grossa, Brazil, 2020; pp. 110–124. [Google Scholar] [CrossRef]
- Fiore, M.C.; Raimondo, F.M.; Mercati, F.; DiGangi, I.; Sunseri, F.; Scialabba, A. Preserving Biodiversity in Marginal Rural Areas: Assessment of Morphological and Genetic Variability of a Sicilian Common Bean Germplasm Collection. Plants 2020, 9, 989. [Google Scholar] [CrossRef]
- Bellon, M.R.; Hodson, D.; Hellin, J. Assessing the vulnerability of traditional maize seed systems in Mexico to climate change. Proc. Natl. Acad. Sci. USA 2011, 108, 13432–13437. [Google Scholar] [CrossRef]
- Galluzzi, G.; Seyoum, A.; Halewood, M.; Noriega, I.L.; Welch, E.W. The Role of Genetic Resources in Breeding for Climate Change: The Case of Public Breeding Programmes in Eighteen Developing Countries. Plants 2020, 9, 1129. [Google Scholar] [CrossRef]
- Maronhas, M.E.S.; Pinilla, N.N.; Lira, J.L. Caminos Del Semiárido Brasileño Para La Gestión de La Agrobiodiversidad. Biodiversidad LA. 28 August 2020. Available online: https://www.biodiversidadla.org/Documentos/Caminos-del-Semiarido-brasileno-para-la-gestion-de-la-agrobiodiversidad (accessed on 28 January 2022).
- Fischer, K. Why new crop technology is not scale-neutral—A critique of the expectations for a crop-based African Green Revolution. Res. Policy 2016, 45, 1185–1194. [Google Scholar] [CrossRef]
- CTNBio. Resumo Geral de Plantas Geneticamente Modificadas Aprovadas Para Comercialização. In Ministério da Ciência, Tecnologia, Inovações e Comunicação: Brasília, Brazil; 29 September 2021. Available online: https://www.gov.br/agricultura/pt-br/assuntos/insumos-agropecuarios/insumos-pecuarios/alimentacao-animal/TabeladePlantasAprovadasparaComercializao.pdf (accessed on 28 January 2022).
- ISAAA. Global Status of Commercialized Biotech/GM Crops in 2017: Biotech Crop Adoption Surges as Economic Benefits Accumate in 22 Years; ISAAA Brief, 53; ISAAA: Ithaca, NY, USA, 2017; p. 2. [Google Scholar]
- Pereira Filho, I.A.; Borghi, E. Sementes de Milho: Nova Safra, Novas Cultivares e Continua a Dominância Dos Transgênicos; Documentos, 251; Embrapa: Sete Lagoas, Brazil, 2020; p. 60. [Google Scholar]
- CTNBio. Liberações Comerciais-Milho. In Ministério da Ciência, Tecnologia, Inovações e Comunicação: Brasília, Brazil 2020. Available online: http://ctnbio.mctic.gov.br/liberacao-comercial#/liberacao-comercial/consultar-processo (accessed on 28 January 2022).
- Zenner de Polania, I. Transgenic Bt Maize in South-and Central America: The Pros and Cons. Rev. Colomb. Cienc. Hortícolas 2021, 15, 1–26. [Google Scholar] [CrossRef]
- Lohn, A.F.; Trtikova, M.; Chapela, I.; Berg, J.V.D.; Du Plessis, H.; Hilbeck, A. Transgene behavior in Zea mays L. crosses across different genetic backgrounds: Segregation patterns, cry1Ab transgene expression, insecticidal protein concentration and bioactivity against insect pests. PLoS ONE 2020, 15, e0238523. [Google Scholar] [CrossRef] [PubMed]
- Duncan, B.; Leyva-Guerrero, E.; Werk, T.; Stojšin, D.; Baltazar, B.M.; García-Lara, S.; Zavala-López, M.; De La Fuente-Martínez, J.M.; Meng, C. Assessment of potential impacts associated with gene flow from transgenic hybrids to Mexican maize landraces. Transgenic Res. 2019, 28, 509–523. [Google Scholar] [CrossRef]
- Kohli, A. Transgene integration, organization and interaction in plants. Plant Mol. Biol. 2003, 52, 247–258. [Google Scholar] [CrossRef]
- Kumar, S.; Fladung, M. Gene stability in transgenic aspen (Populus). II. Molecular characterization of variable expression of transgene in wild and hybrid aspen. Planta 2001, 213, 731–740. [Google Scholar] [CrossRef]
- Latham, J.R.; Wilson, A.K.; Steinbrecher, R.A. The Mutational Consequences of Plant Transformation. J. Biomed. Biotechnol. 2006, 2006, 025376. [Google Scholar] [CrossRef]
- Yin, Z.; Plader, W.; Malepszy, S. Transgene inheritance in plants. J. Appl. Genet. 2004, 45, 127–144. [Google Scholar]
- Zanatta, C.B.; Benevenuto, R.F.; Nodari, R.O.; Agapito-Tenfen, S.Z. Stacked genetically modified soybean harboring herbicide resistance and insecticide rCry1Ac shows strong defense and redox homeostasis disturbance after glyphosate-based herbicide application. Environ. Sci. Eur. 2020, 32, 104. [Google Scholar] [CrossRef]
- Louette, D.; Smale, M. Farmers’ seed selection practices and traditional maize varieties in Cuzalapa, Mexico. Euphytica 2000, 113, 25–41. [Google Scholar] [CrossRef]
- Louette, D.; Charrier, A.; Berthaud, J. In Situ conservation of maize in Mexico: Genetic diversity and Maize seed management in a traditional community. Econ. Bot. 1997, 51, 20–38. [Google Scholar] [CrossRef]
- Bellon, M.; Brush, S.B. Keepers of maize in Chiapas, Mexico. Econ. Bot. 1994, 48, 196–209. [Google Scholar] [CrossRef]
- Louette, D. Traditional Management of Seed and Genetic Diversity: What Is a Landrace? In Genes in the field on-farm conservation of crop diversity; International Plant Genetic Resources Institute: Rome, Italy; International Development Research Centre: Ottawa, ON, Canada; Lewis Publishers: Boca Raton, FL, USA, 2000; pp. 109–142. [Google Scholar]
- Hugo, P.R.; Brush, S.B.; Qualset, C.O. Dynamic Management of Maize Landraces in Central Mexico. Econ. Bot. 2003, 57, 21–34. [Google Scholar] [CrossRef]
- Guzzon, F.; Rios, L.W.A.; Cepeda, G.M.C.; Polo, M.C.; Cabrera, A.C.; Figueroa, J.M.; Hoyos, A.E.M.; Calvo, T.W.J.; Molnar, T.L.; León, L.A.N.; et al. Conservation and Use of Latin American Maize Diversity: Pillar of Nutrition Security and Cultural Heritage of Humanity. Agronomy 2021, 11, 172. [Google Scholar] [CrossRef]
- Melgarejo, M.E.D.; Estigarribia, P.E.P.; Iehisa, J.M.; Arrua, J.M.M.; Martínez, C.C.C.; Arrua, A.A. Contamination of corn grain for human consumption with transgenic sequences in Paraguay. Food Sci. Technol. 2020, 40, 537–542. [Google Scholar] [CrossRef]
- Bellon, M. The ethnoecology of maize variety management: A case study from Mexico. Hum. Ecol. 1991, 19, 389–418. [Google Scholar] [CrossRef]
- Brasil Decreto, N. 680, de 24 de Abril de 2003; Presidência da República, Casa Civil, Subchefia para Assuntos Jurídicos: Brasília, Brazil, 2003. Available online: https://www.planalto.gov.br/ccivil_03/decreto/2003/d4680.htm (accessed on 28 January 2022).
- Lacey, H. Seeds and the Knowledge They Embody. Peace Rev. 2000, 12, 563–569. [Google Scholar] [CrossRef]
- Hilbeck, A.; Lebrecht, T.; Vogel, R.; Heinemann, J.A.; Binimelis, R. Farmer’s choice of seeds in four EU countries under different levels of GM crop adoption. Environ. Sci. Eur. 2013, 25, 12. [Google Scholar] [CrossRef]
- Capellesso, A.J.; Cazella, A.A.; Filho, A.L.S.; Farley, J.; Martins, D.A. Economic and environmental impacts of production intensification in agriculture: Comparing transgenic, conventional, and agroecological maize crops. Agroecol. Sustain. Food Syst. 2016, 40, 215–236. [Google Scholar] [CrossRef]
- Hilbeck, A.; Binimelis, R.; Defarge, N.; Steinbrecher, R.; Székács, A.; Wickson, F.; Antoniou, M.; Bereano, P.L.; Clark, E.A.; Hansen, M.; et al. No scientific consensus on GMO safety. Environ. Sci. Eur. 2015, 27, 1. [Google Scholar] [CrossRef]
- Fatoretto, J.C.; Michel, A.P.; Silva-Filho, M.; Silva, N. Adaptive Potential of Fall Armyworm (Lepidoptera: Noctuidae) Limits Bt Trait Durability in Brazil. J. Integr. Pest Manag. 2017, 8. [Google Scholar] [CrossRef]
- Hilbeck, A.; Schmidt, J.E.U. Another View on Bt Proteins–How Specific Are They and What Else Might They Do? Biopestic. Int. 2006, 2, 1–50. [Google Scholar]
- Curis, M.C.; Bertolaccini, M.A.T.I. Influência de presas criadas sobre maíces Bt sobre parámetros biológicos de Eriopis connexa (Coleoptera: Coccinellidae). Rev. Ciênc. Agrár. 2013, 36, 174–181. [Google Scholar]
- Tibbett, M.; Fraser, T.D.; Duddigan, S. Identifying potential threats to soil biodiversity. PeerJ 2020, 8, e9271. [Google Scholar] [CrossRef]
- Aksoy, E.; Louwagie, G.; Gardi, C.; Gregor, M.; Schröder, C.; Löhnertz, M. Assessing soil biodiversity potentials in Europe. Sci. Total Environ. 2017, 589, 236–249. [Google Scholar] [CrossRef]
- Eker, S.; Ozturk, L.; Yazici, A.; Erenoglu, B.; Romheld, V.; Cakmak, I. Foliar-Applied Glyphosate Substantially Reduced Uptake and Transport of Iron and Manganese in Sunflower (Helianthus annuus L.) Plants. J. Agric. Food Chem. 2006, 54, 10019–10025. [Google Scholar] [CrossRef]
- Cakmak, I.; Yazici, A.; Tutus, Y.; Ozturk, L. Glyphosate reduced seed and leaf concentrations of calcium, manganese, magnesium, and iron in non-glyphosate resistant soybean. Eur. J. Agron. 2009, 31, 114–119. [Google Scholar] [CrossRef]
- Carvalho, F.P.; França, A.C.; Franco, M.H.R.; Avelar, M.; Moreira, S.D.; Alecrim, A.O.; Dos Santos, J.B. Sensibilidade de plantas de café micorrizadas à herbicidas. Rev. Bras. de Herbic. 2014, 13, 134. [Google Scholar] [CrossRef]
- De Almeida, V.E.S.; Friedrich, K.; Tygel, A.F.; Melgarejo, L.; Carneiro, F.F. Use of genetically modified crops and pesticides in Brazil: Growing hazards. Ciência Saúde Coletiva 2017, 22, 3333–3339. [Google Scholar] [CrossRef] [PubMed]
- Benbrook, C.M. Trends in glyphosate herbicide use in the United States and globally. Environ. Sci. Eur. 2016, 28, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Binimelis, R.; Pengue, W.; Monterroso, I. “Transgenic treadmill”: Responses to the emergence and spread of glyphosate-resistant johnsongrass in Argentina. Geoforum 2009, 40, 623–633. [Google Scholar] [CrossRef]
- Pengue, W.A. Transgenic Crops in Argentina: The Ecological and Social Debt. Bull. Sci. Technol. Soc. 2005, 25, 314–322. [Google Scholar] [CrossRef]
- Melgarejo, L. Biosafety Regulations and Practices and Consequences in Brazil: Who Wants to Hide the Problems? In Brazil in the Anthropocene: Conflicts between Predatory Development and Environmental Policies; Issberner, L.-R., Léna, P., Eds.; Routledge: New York, NY, USA, 2018; pp. 143–157. [Google Scholar]
- Souza, M.M.O.; Gurgel, A.d.M.; Fernandes, G.B.; Melgarejo, L.; Bittencourt, N.A.; Friedrich, K. Agrotóxicos e Transgênicos: Retrocessos Socioambientais e Avanços Conservadores No Governo Bolsonaro. Rev. Anpege 2020, 16, 319–352. [Google Scholar] [CrossRef]
- Petry, C.; Calliari, M.R.T.; Melgarejo, L.; Fernandes, G.B.; Bittencourt, N.A.; Souza, M.M.O.; Mulinari, J.; Reichert Júnior, R.; Francisco, W. Food in Security, Agrochemicals, New Biotechnologies and Democracy. In Agroecology: Insights, Experiences and Perspectives; Mossi, A.J., Petry, C., Reichert, F.W., Eds.; Agriculture issues and policies; Nova Science Publishers: Hauppauge, NY, USA, 2020; pp. 253–288. [Google Scholar]
- Lipton, C.R.; Dautlick, J.X.; Grothaus, G.D.; Hunst, P.L.; Magin, K.M.; Mihaliak, C.A.; Rubio, F.M.; Stave, J.W. Guidelines for the Validation and Use of Immunoassays for Determination of Introduced Proteins in Biotechnology Enhanced Crops and Derived Food Ingredients. Food Agric. Immunol. 2000, 12, 153–164. [Google Scholar] [CrossRef]
- Stave, J.W. Detection of new or modified proteins in novel foods derived from GMO–future needs. Food Control 1999, 10, 367–374. [Google Scholar] [CrossRef]
- Curado, F.F.; Santos, A.D.S.; Fagundes, R.d.C.; Silva, A.C.d.L.; Bianchini, P.C. Manejo Comunitário Da Agrobiodiversidade: Produção Agroecológica de Sementes de Variedades Crioulas Por Agricultores Familiares; Documentos, 237; Embrapa: Aracaju, Brazil, 2020; p. 54. [Google Scholar]
- Dos Santos, A.D.; Curado, F.F.; Silva, A.C.d.L.; Bianchini, P.C.; Fagundes, R.d.C. Manual de Instalação de Ensaios Participativos Para Comparação de Variedades Crioulas; Documentos, 239; Embrapa: Aracaju, Brazil, 2020; p. 37. [Google Scholar]
- Ferment, G.; Zanoni, M.; Brack, P.; Kageyama, P.; Nodari, R.O. Coexistência: O Caso Do Milho: Proposta de Revisão Da Resolução Normativa N 4 Da CTNBio; NEAD debate; MDA: Brasília, Brazil, 2009. [Google Scholar]
- Fernandes, G.B. Seminário Sobre Proteção Da Agrobiodiversidade e Direito Dos Agricultores: Propostas Para Enfrentar a Contaminação Transgênica Do Milho: Atas, Discussões e Encaminhamentos; Fernandes, G.B., Ferment, G., Avanci, J., Eds.; NEAD debate; Ministério do Desenvolvimento Agrário: Brasília, Brazil, 2010. [Google Scholar]
- Fernandes, G.B.; Marinho, W. O Caminho Da Liberalização Dos Transgênicos No Brasil. Agroecología 2017, 12, 19–28. [Google Scholar]
- CTNBio. Normative Resolution No. 04 of August 16th, 2007. In Ministério da Ciência, Tecnologia, Inovações e Comunicação: Brasília, Brazil; 2007. Available online: http://ctnbio.mctic.gov.br/resolucoes-normativas/-/asset_publisher/OgW431Rs9dQ6/content/resolucao-normativa-n%C2%BA-4-de-16-de-agosto-de-2007;jsessionid=E1A3F6C2EBB3AE8C1C5621F81DB416B4.columba#:~:text=Estabelecer%20as%20dist%C3%A2ncias%20m%C3%ADnimas%20de,sistemas%20de%20produ%C3%A7%C3%A3o%20no%20campo (accessed on 28 January 2022).
- Hofmann, F.; Otto, M.; Wosniok, W. Maize pollen deposition in relation to distance from the nearest pollen source under common cultivation-results of 10 years of monitoring (2001 to 2010). Environ. Sci. Eur. 2014, 26, 24. [Google Scholar] [CrossRef]
- Dyer, G.A.; Serratos-Hernández, J.A.; Perales, H.R.; Gepts, P.; Piñeyro-Nelson, A.; Chávez, A.; Salinas-Arreortua, N.; Yúnez-Naude, A.; Taylor, J.E.; Alvarez-Buylla, E.R. Dispersal of Transgenes through Maize Seed Systems in Mexico. PLoS ONE 2009, 4, e5734. [Google Scholar] [CrossRef]
- Iversen, M.; Grønsberg, I.M.; Berg, J.V.D.; Fischer, K.; Aheto, D.W.; Bøhn, T. Detection of transgenes in local maize varieties of small-scale farmers in Eastern cape, South Africa. PLoS ONE 2014, 9, e116147. [Google Scholar] [CrossRef] [PubMed]
- Díaz, L.; Galindo, I. Detección e Identificación de Eventos Asociados a Organismos Vivos Modificados En Semillas de Maíz (Zea Mays L.) En Venezuela Empleando Métodos de Inmunoensayo y Análisis Por PCR. Rev. Fac. Agron. UCV 2014, 40, 37–49. [Google Scholar]
- Kawashima, S.; Nozaki, H.; Hamazaki, T.; Sakata, S.; Hama, T.; Matsuo, K.; Nagasawa, A. Environmental effects on long-range outcrossing rates in maize. Agric. Ecosyst. Environ. 2011, 142, 410–418. [Google Scholar] [CrossRef]
- Munarini, A.; Nerling, D.; Coelho, C.M.M.; Nodari, R.O. Maize landraces management to avoid transgenic contamination, decreases yield and seed quality. Bragantia 2021, 80, 3921. [Google Scholar] [CrossRef]
- CTNBio. Resolução Normativa No 9, de 2 de Dezembro de 2011-REVOGADA PELA RN 24; Ministério da Ciência, Tecnologia, Inovações e Comunicação: Brasília, Brazil, 2011; p. 20. Available online: http://ctnbio.mctic.gov.br/resolucoes-normativas/-/asset_publisher/OgW431Rs9dQ6/content/resolucao-normativa-n%C2%BA-9-de-2-de-dezembro-de-2011-revogada-pela-rn-24;jsessionid=E1A3F6C2EBB3AE8C1C5621F81DB416B4.columba#:~:text=O%20objetivo%20do%20monitoramento%20p%C3%B3s,com%20sua%20aplica%C3%A7%C3%A3o%20de%20uso (accessed on 28 January 2022).
- CTNBio. Resolução Normativa No 24, de 07 de Janeiro de 2020-REVOGADA PELA RN 32; Ministério da Ciência, Tecnologia, Inovações e Comunicação: Brasília, Brazil, 2020; p. 26. Available online: http://ctnbio.mctic.gov.br/resolucoes-normativas/-/asset_publisher/OgW431Rs9dQ6/content/resolucao-normativa-n%C2%BA-24-de-07-de-janeiro-de-2020-revogada-pela-rn-32?redirect=http%3A%2F%2Fctnbio.mctic.gov.br%2Fresolucoes-normativas%3Fp_p_id%3D101_INSTANCE_OgW431Rs9dQ6%26p_p_lifecycle%3D0%26p_p_state%3Dnormal%26p_p_mode%3Dview%26p_p_col_id%3Dcolumn-2%26p_p_col_count%3D3 (accessed on 28 January 2022).
- CTNBio. Resolução Normativa No 32, de 15 de Junho de 2021; Ministério da Ciência, Tecnologia, Inovações e Comunicação: Brasília, Brazil, 2021. Available online: https://www.in.gov.br/en/web/dou/-/resolucao-normativa-n-32-de-15-de-junho-de-2021-326241632#:~:text=Disp%C3%B5e%20sobre%20as%20normas%20para,de%20origem%20vegetal%20e%20animal.&text=%C2%A7%203%C2%B0%20N%C3%A3o%20se,tenha%20sido%20por%20ela%20aprovado (accessed on 28 January 2022).
- CTNBio. Resolução Normativa No 15, de 13 de Fevereiro de 2015-REVOGADA PELA RN 24; Ministério da Ciência, Tecnologia, Inovações e Comunicação: Brasília, Brazil, 2015; 26p. Available online: http://ctnbio.mctic.gov.br/resolucoes-normativas/-/asset_publisher/OgW431Rs9dQ6/content/resolucao-n%C2%BA-20-de-23-de-marco-de-2018-revogada-pela-rn-24?redirect=http%3A%2F%2Fctnbio.mctic.gov.br%2Fresolucoes-normativas%3Fp_p_id%3D101_INSTANCE_OgW431Rs9dQ6%26p_p_lifecycle%3D0%26p_p_state%3Dnormal%26p_p_mode%3Dview%26p_p_col_id%3Dcolumn-2%26p_p_col_count%3D3 (accessed on 28 January 2022).
- Reis, J.N.P. A insustentável distribuição da terra no semiárido brasileiro. Cad. Ciências Sociais Apl. 2019, 16, 21–38. Available online: https://repositorio.ufc.br/bitstream/riufc/51276/1/2019_art_jnpreis.pdf (accessed on 28 January 2022). [CrossRef]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Mode of Action | Tested Protein | Sample I | Sample II |
|---|---|---|---|
| Total positive collections | 161 | 213 | |
| IR | VIP3A, Bt-Cry1A, Bt-Cry3Bb, Bt-Cry1F, Bt-Cry2Ab, mCry3A, Cry1Ac | 158 | 159 |
| GAT | PAT | 5 | 19 |
| GT | CP4 EPSPS | 84 | 131 |
| Mode of Action | Tested Protein | Total Positive Samples | |
| IR | VIP3A, Bt-Cry1A, Bt-Cry3Bb, Bt-Cry1F, Bt-Cry2Ab, mCry3A, Cry1Ac | 317 | |
| GAT | PAT | 24 | |
| GT | CP4 EPSPS | 215 | |
| Category | Percentile | Description | Potential Impacts |
|---|---|---|---|
| S0 | 30% | Weak Drought | Entering drought: short-term heat wave decreasing planting, crop growth, or grazing. Coming out of drought: some prolonged water deficits, pastures or crops not fully recovered. |
| S1 | 20% | Moderate Drought | Some damage to crops, pastures, streams, reservoirs, or wells with low levels, some developing or imminent water shortages; voluntary restrictions on water use requested. |
| S2 | 10% | Severe drought | Potential crop or pasture losses; regular water shortages; water restrictions imposed. |
| S3 | 5% | Extreme drought | Large crop/pasture losses; widespread water scarcity or restrictions |
| S4 | 2% | Exceptional Drought | Exceptional and widespread crop/pasture losses; scarcity of water in reservoirs, streams, and water wells, creating emergency situations. |
| Company | Product | Event (Trait) | Detection Limit (%) |
|---|---|---|---|
| 2018/2019 | |||
| Romer Labs | AgraStrip© Triple Trait Bulk Grain | CP4 EPSPS | 0.1 |
| Bt-Cry1A (Bt-Cry1Ab, Bt-Cry1Ac, Bt-Cry1A.105), Bt-Cry3Bb | 9.5 | ||
| AgraStrip© Cry1F Bulk Grain (water extract) | Bt-Cry1F | 0.9 | |
| AgraStrip© Cry2Ab Bulk Grain | Bt-Cry2Ab | 0.9 | |
| AgraStrip© VIP Bulk Grain | VIP3A | 0.33 | |
| AgraStrip© LL Bulk Grain | PAT | 0.9 | |
| 2020/2021 | |||
| Romer Labs | AgraStrip© PMI Bulk Grain | PMI (Vip3A, mCry3A) | 0.5 |
| AgraStrip© RUR-HS Bulk Grain | CP4 EPSPS | 0.125 | |
| AgraStrip© Cry1Ac Bulk Grain | Cry1Ac | 0.9 | |
| AgraStrip© LL Bulk Grain | PAT | 0.9 | |
| Year | Unique Identifier | Event (Trait) | Protein |
|---|---|---|---|
| 2018 | MON-89Ø34-3 x DAS-Ø15Ø7-1 x MON-ØØ6Ø3-6 x SYN-IR162-4 x DAS-4Ø278-9 | MON 89034 x TC1507 x MIR162 x NK603 x DAS-40278-9 | Cry1A105, Cry2Ab2, Cry1F and Vip3Aa20, PAT, CP4 epsps and AAD-1 |
| SYN-ØØØ98-3 | MZIR098 (food/feed) | mCry3A/eCry3.1Ab | |
| 2019 | MON-87427-7 x MON-89Ø34-3 x DAS-Ø15Ø7-1 x MON-87411-9 x DAS-59122-7 x DAS-4Ø278-9 | MON-87427-7 x MON-89034-3 x DAS-01507-1 x MON-87411-9 x DAS-59122-7 x DAS-40278-9 | CP4 epsps (aroA:CP4), Cry2Ab2, Cry1A.105, Cry1F, PAT, Cry34Ab1, Cry35Ab1, Cry3Bb |
| MON-87427-7 x MON87419-8 x MON-ØØ6Ø3-6 | MON 87427 × MON 87419 × NK603 | CP4 EPSPS, DMO and PAT | |
| MON-87427-7 x MON-89Ø34-3 x SYN-IR162-4 x MON-ØØ6Ø3-6 | MON 87427 x MON 89034 x MIR162 x NK603 (and subcombinations) | CP4 EPSPS, Cry1 A.1 05, Cry2Ab2 and Vip3Aa | |
| 2020 | MON-ØØ6Ø3-6 x ACS-ZMØØ3-2 X DAS-4Ø278-9 | NK603 x T25 x DAS-40278 | CP4 EPSPS, PAT, AAD1 |
| MON-89Ø34-3 x DAS-Ø15Ø7-1 x MON-ØØ6Ø3-6 x SYN-IR162-4 x DAS-4Ø278-9 | MON-89034-3 x DAS-01507-1 x SYN-IR162-4 x MON-00630-6 x DAS40278-9 (and subcombinations) | Cry1A.105 e Cry2Ab2, Cry1F, PAT, VIP3Aa20, CP4 EPSPS, AAD-1 | |
| MON-95379-3 | MON 95379 | Cry1Da_7, Cry1B.868, Cry1Be, Cry1Ca, Cry1Ab | |
| 2021 | DP-ØØ4114-3 | DP4114-3 | Cry1F, Cry34Ab1 and Cry35Ab1, PAT |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Fernandes, G.B.; Silva, A.C.d.L.; Maronhas, M.E.S.; Santos, A.d.S.d.; Lima, P.H.C. Transgene Flow: Challenges to the On-Farm Conservation of Maize Landraces in the Brazilian Semi-Arid Region. Plants 2022, 11, 603. https://doi.org/10.3390/plants11050603
Fernandes GB, Silva ACdL, Maronhas MES, Santos AdSd, Lima PHC. Transgene Flow: Challenges to the On-Farm Conservation of Maize Landraces in the Brazilian Semi-Arid Region. Plants. 2022; 11(5):603. https://doi.org/10.3390/plants11050603
Chicago/Turabian StyleFernandes, Gabriel Bianconi, Ana Cláudia de Lima Silva, Maitê Edite Sousa Maronhas, Amaury da Silva dos Santos, and Paola Hernandez Cortez Lima. 2022. "Transgene Flow: Challenges to the On-Farm Conservation of Maize Landraces in the Brazilian Semi-Arid Region" Plants 11, no. 5: 603. https://doi.org/10.3390/plants11050603
APA StyleFernandes, G. B., Silva, A. C. d. L., Maronhas, M. E. S., Santos, A. d. S. d., & Lima, P. H. C. (2022). Transgene Flow: Challenges to the On-Farm Conservation of Maize Landraces in the Brazilian Semi-Arid Region. Plants, 11(5), 603. https://doi.org/10.3390/plants11050603