
In Vitro Technology in Plant Conservation: Relevance to Biocultural Diversity



Abstract
1. Introduction
2. The Loss of Plant Diversity: Threats, Drivers, and Magnitude
Biocultural Diversity and Plant Conservation
3. Plant Conservation and Social Dimension of Technoscience
3.1. Studies on the Conservation of Plants: Limitations and Opportunities
3.2. Indigenous Plants
3.3. Social Dimension, Technoscience and Plant Relationships: Concerns
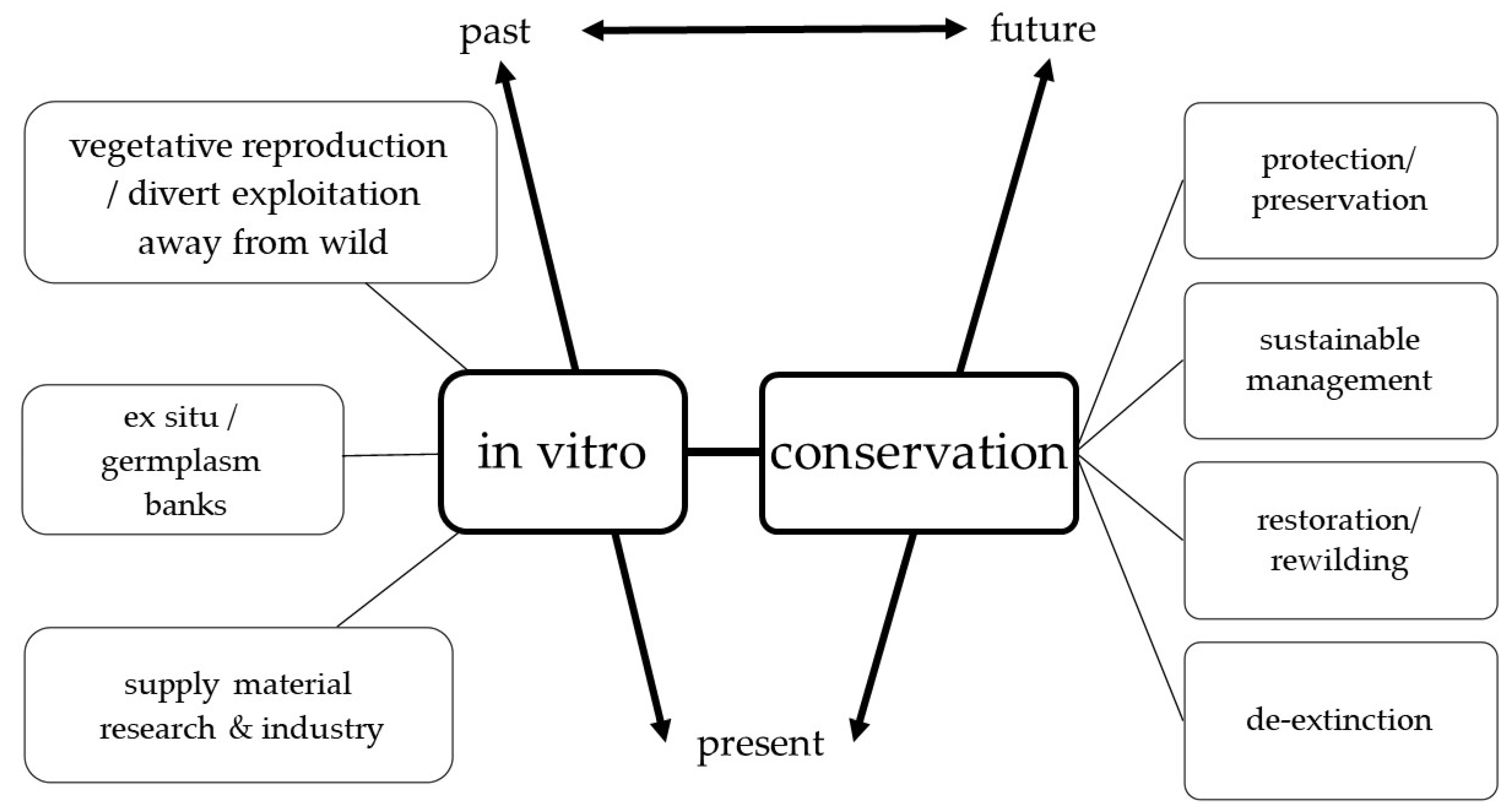
4. The Nexus of In Vitro Technology and Plant Conservation
4.1. Micropropagation and Cryopreservation Methods
4.2. Advantages of In Vitro Technology
- In vitro methods facilitate tapping into the abilities of plant tissue to reproduce vegetatively from limited starting material, thus reducing the need to harvest whole plants or numerous plants from the wild, preventing the depletion of vulnerable populations in their natural habitats. Plant multiplication by in vitro technology is achieved by proliferation of the apical or axillary meristems, which consist of rapidly growing cells, are generally genetically consistent, relatively virus-free, and bear greater capacity for multiplication compared to non-meristematic tissues. Alternately, plants can be propagated by regeneration, in which individual plant cells express their inherent capacity, referred to as “totipotency”, to divide and differentiate to form complete plants. Both modes of plant propagation have specific advantages. While multiplication using pre-existing meristems is known to produce genetically identical clones that can be used to enrich a specific population, the plants produced by regeneration may exhibit genetic variations that can be exploited to create genetically diverse plant populations. Thus, in vitro technology allows regeneration of fully functional specimens from small amounts of tissue, saving time and money and reducing the need to harvest numerous specimens. The mass production of rare or useful plants via micropropagation diverts from overexploitation of wild specimens for commercial use [131,169].
- In vitro grown specimens of threatened or rare plants can also help populate ex situ collections in botanical gardens and other research institutions that can be later re-introduced in natural habitats [92,170,171]. This allows for the short and long-term storage of germplasm, which protects it from current threats, and also facilitates the selection of genotypes for future use [172].
4.3. Limitations of In Vitro Technologies
4.4. Challenges and Opportunities at the Interface of Plant Conservation, In Vitro Technology, and Indigenous Plants
5. Final Considerations about In Vitro Technology in Biocultural Conservation
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Barnhill-Dilling, S.K.; Delborne, J.A. Whose Intentions? What Consequences? Interrogating “Intended Consequences” for Conservation with Environmental Biotechnology. Conserv. Sci. Pract. 2021, 3, e406. [Google Scholar] [CrossRef]
- Pretty, J.; Adams, B.; Berkes, F.; Ferreira, S.; Dudley, N.; Hunn, E.; Maffi, L.; Milton, K.; Rapport, D.; Robbins, P.; et al. The Intersections of Biological Diversity and Cultural Diversity: Towards Integration. Conserv. Soc. 2009, 7, 100–112. [Google Scholar]
- Dunn, L.; Burney, L. Ethnobotany, the Science of Survival: A Declaration from Kaua’i. Econ. Bot. 2007, 61, 1–2. [Google Scholar] [CrossRef]
- Isbell, F.; Calcagno, V.; Hector, A.; Connolly, J.; Harpole, W.S.; Reich, P.B.; Scherer-Lorenzen, M.; Schmid, B.; Tilman, D.; van Ruijen, J.; et al. High Plant Diversity Is Needed to Maintain Ecosystem Services. Nature 2011, 477, 199–202. [Google Scholar] [CrossRef] [PubMed]
- Heywood, V.H. Plant Conservation in the Anthropocene–Challenges and Future Prospects. Plant Divers. 2017, 39, 314–330. [Google Scholar] [CrossRef] [PubMed]
- Amos, R. Reassessing the Role of Plants in Society. Int. J. Law Context 2017, 13, 295–315. [Google Scholar] [CrossRef]
- Cunsolo, A.; Ellis, N.R. Ecological Grief as a Mental Health Response to Climate Change-Related Loss. Nat. Clim. Chang. 2018, 8, 275–281. [Google Scholar] [CrossRef]
- Benn, J. What Is Biodiversity? United Nations Environment Programme, 2.; World Conservation Monitoring Centre: New York, NY, USA, 2010; Available online: https://www.unesco.pl/fileadmin/user_upload/pdf/BIODIVERSITY_FACTSHEET.pdf (accessed on 14 September 2021).
- Cardinale, B.J.; Duffy, J.E.; Gonzalez, A.; Hooper, D.U.; Perrings, C.; Venail, P.; Narwani, A.; Mace, G.M.; Tilman, D.; Wardle, D.A. Biodiversity Loss and Its Impact on Humanity. Nature 2012, 486, 59–67. [Google Scholar] [CrossRef]
- Mace, G.M. The Ecology of Natural Capital Accounting. Oxf. Rev. Econ. Policy 2019, 35, 54–67. [Google Scholar] [CrossRef]
- Pimm, S.L.; Alibhai, S.; Bergl, R.; Dehgan, A.; Giri, C.; Jewell, Z.; Joppa, L.; Kays, R.; Loarie, S. Emerging Technologies to Conserve Biodiversity. Trends Ecol. Evol. 2015, 30, 685–696. [Google Scholar] [CrossRef]
- Harfoot, M.B.; Tittensor, D.P.; Knight, S.; Arnell, A.P.; Blyth, S.; Brooks, S.; Burgess, N.D. Present and Future Biodiversity Risks from Fossil Fuel Exploitation. Conserv. Lett. 2018, 11, e12448. [Google Scholar] [CrossRef]
- Bradshaw, C.J.; Ehrlich, P.R.; Beattie, A.; Ceballos, G.; Crist, E.; Diamond, J.; Dirzo, R.; Ehrlich, A.; Harte, M.E.; Pyke, G.; et al. Underestimating the Challenges of Avoiding a Ghastly Future. Front. Conserv. Sci. 2021, 1, 9. [Google Scholar] [CrossRef]
- Isbell, F.; Adler, P.R.; Eisenhauer, N.; Fornara, D.; Kimmel, K.; Kremen, C.; Letourneau, D.K.; Liebman, M.; Polley, H.W.; Quijas, S. Benefits of Increasing Plant Diversity in Sustainable Agroecosystems. J. Ecol. 2017, 105, 871–879. [Google Scholar] [CrossRef]
- Zhang, Y.; Lü, X.; Isbell, F.; Stevens, C.; Han, X.; He, N.; Zhang, G.; Yu, Q.; Huang, J.; Han, X. Rapid Plant Species Loss at High Rates and at Low Frequency of N Addition in Temperate Steppe. Glob. Chang. Biol. 2014, 20, 3520–3529. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, G.; Ehrlich, P.R.; Barnosky, A.D.; Garcia, A.; Pringle, R.; Palmer, T. Accelerated Modern Human–Induced Species Losses: Entering the Sixth Mass Extinction. Sci. Adv. 2015, 1, e1400253. [Google Scholar] [CrossRef] [PubMed]
- De Vos, J.; Joppa, L.; Gittleman, J.L.; Stephens, P.R.; Pimm, S.L. Estimating the Normal Background Rate of Species Extinction. Conserv. Biol. 2015, 29, 452–462. [Google Scholar] [CrossRef] [PubMed]
- Ceballos, G.; Ehrlich, P.R.; Raven, P.H. Vertebrates on the Brink as Indicators of Biological Annihilation and the Sixth Mass Extinction. Proc. Natl. Acad. Sci. USA 2020, 117, 13596–13602. [Google Scholar] [CrossRef]
- Hawkins, B. Plants for Life: Medicinal Plant Conservation and Botanic Gardens; BGCI: Richmond, UK, 2007. [Google Scholar]
- Pandit, R.; Pörtner, H.O.; Scholes, R.J.; Agard, J.; Archer, E.; Arneth, A.; Bai, X.; Barnes, D.; Burrows, M.; Chan, L.; et al. Scientific Outcome of the IPBES-IPCC Co-Sponsored Workshop on Biodiversity and Climate Change; IPBES Secretariat—UWA School of Agriculture: Bonn, Germany, 2021; p. 256. [Google Scholar]
- Schmidt, B.; Gemeinholzer, B.; Treolar, A. Open Data in Global Environmental Research: The Belmont Forum’s Open Data Survey. PLoS ONE 2016, 11, e0146695. [Google Scholar] [CrossRef]
- Hanspach, J.; Haider, L.J.; Oteros-Rozas, E.; Olafsson, A.S.; Gulsrud, N.M.; Raymond, C.M.; Torralba, M.; Martín-López, B.; Bieling, C.; Garcia-Martin, M. Biocultural Approaches to Sustainability: A Systematic Review of the Scientific Literature. People Nat. 2020, 2, 643–659. [Google Scholar] [CrossRef]
- Elands, B.; Vierikko, K.; Andersson, E.; Fischer, L.; Goncalves, P.; Haase, D.; Kowarik, I.; Luz, A.; Niemelä, J.; Santos-Reis, M. Biocultural Diversity: A Novel Concept to Assess Human-Nature Interrelations, Nature Conservation and Stewardship in Cities. Urban For. Urban Green. 2019, 40, 29–34. [Google Scholar] [CrossRef]
- Hemstrom, K.; Palmer, H. On participatory research, knowledge integration and societal transformation. In Anatomy of a 21st-Century Sustainability Project: The Untold Stories; Dymitrow, M., Ingelhag, K., Eds.; Mistra Urban Futures; Chalmers University of Technology: Gothenburg, Sweden, 2019; pp. 29–37. [Google Scholar]
- Mayhew, P.J. Global Climate and Extinction: Evidence from the fossil records. In Climate Change, Ecology and Systematics; Hodkinson, T.R., Jones, M.B., Waldren, S., Parnell, J.A.N., Eds.; Cambridge University Press: Cambridge, UK, 2011; pp. 99–121. [Google Scholar]
- Mayhew, P.J.; Jenkins, G.B.; Benton, T.G. A Long-Term Association between Global Temprature and Biodiversity. Origination and Extinction in the Fossil Record. Proc. R. Soc. B Biol. Sci. 2008, 275, 47–53. [Google Scholar] [CrossRef]
- Vellend, M. Habitat Loss Inhibits Recovery of Plant Diversity as Forests Regrow. Ecology 2003, 84, 1158–1164. [Google Scholar] [CrossRef]
- Watson, J.E.; Evans, T.D.; Venter, O.; Maxwell, S.L. Manage Forests as Protection against Warming. Nature 2019, 567, 311–312. [Google Scholar] [CrossRef]
- Turney, C.; Ausseil, A.-G.; Broadhurst, L. Urgent Need for an Integrated Policy Framework for Biodiversity Loss and Climate Change. Nat. Ecol. Evol. 2020, 4, 996. [Google Scholar] [CrossRef]
- Thomas, K.; Hardy, R.D.; Lazrus, H.; Mendez, M.; Orlove, B.; Rivera-Collazo, I.; Roberts, J.T.; Rockman, M.; Warner, B.P.; Winthrop, R. Explaining Differential Vulnerability to Climate Change: A Social Science Review. Wiley Interdiscip. Rev. Clim. Chang. 2019, 10, e565. [Google Scholar] [CrossRef]
- Thuiller, W.; Lavorel, S.; Araújo, M.B.; Sykes, M.T.; Prentice, I.C. Climate Change Threats to Plant Diversity in Europe. Proc. Natl. Acad. Sci. USA 2005, 102, 8245–8250. [Google Scholar] [CrossRef] [PubMed]
- Petelka, J.; Plagg, B.; Säumel, I.; Zerbe, S. Traditional Medicinal Plants in South Tyrol (Northern Italy, Southern Alps): Biodiversity and Use. J. Ethnobiol. Ethnomed. 2020, 16, 1–15. [Google Scholar] [CrossRef]
- Di Sacco, A.; Hardwick, K.A.; Blakesley, D.; Brancalion, P.H.; Breman, E.; Cecilio Rebola, L.; Chomba, S.; Dixon, K.; Elliott, S.; Ruyonga, G. Ten Golden Rules for Reforestation to Optimize Carbon Sequestration, Biodiversity Recovery and Livelihood Benefits. Glob. Chang. Biol. 2021, 27, 1328–1348. [Google Scholar] [CrossRef]
- Allan, R.P.; Hawkins, E.; Bellouin, N.; Collins, B. IPC. Summary for policymakers. In Climate Change 2021: The Physical Science Basis; Cambridge University Press: Cambridge, UK, 2021; Available online: https://www.ipcc.ch/report/ar6/wg1/downloads/report/IPCC_AR6_WGI_SPM_final.pdf (accessed on 7 November 2021).
- Oldekop, J.A.; Sims, K.R.E.; Karna, B.K.; Whittingham, M.J.; Agrawal, A. Reductions in Deforestation and Poverty from Decentralized Forest Management in Nepal. Nat. Sustain. 2019, 2, 421–428. [Google Scholar] [CrossRef]
- Strassburg, B.B.N.; Iribarrem, A.; Beyer, H.L.; Cordeiro, C.L.; Crouzeilles, R.; Jakovac, C.C.; Braga Junqueira, A.; Lacerda, E.; Latawiec, A.E.; Balmford, A.; et al. Global Priority Areas for Ecosystem Restoration. Nature 2020, 586, 724–729. [Google Scholar] [CrossRef]
- Verschuuren, B. Conclusions: How the cultural, spiritual and philosophical underpinnings of sacred natural sites can make conservation in Asia more effective and sustainable. In Asian Sacred Natural Sites; Routledge: London, UK, 2016; pp. 319–331. ISBN 1-315-67627-3. [Google Scholar]
- Zecca, G.; Casazza, G.; Piscopo, S.; Minuto, L.; Grassi, F. Are the Responses of Plant Species to Quaternary Climatic Changes Idiosyncratic? A Demographic Perspective from the Western Alps. Plant Ecol. Divers. 2017, 10, 273–281. [Google Scholar] [CrossRef]
- Christmas, M.J.; Breed, M.F.; Lowe, A.J. Constraints to and Conservation Implications for Climate Change Adaptation in Plants. Conserv. Genet. 2016, 17, 305–320. [Google Scholar] [CrossRef]
- Sayre, M.; Stenner, T.; Argumedo, A. You Can’t Grow Potatoes in the Sky: Building Resilience in the Face of Climate Change in the Potato Park of Cuzco, Peru. Cult. Agric. Food Environ. 2017, 39, 100–108. [Google Scholar] [CrossRef]
- Pecl, G.T.; Araújo, M.B.; Bell, J.D.; Blanchard, J.; Bonebrake, T.C.; Chen, I.-C.; Clark, T.; Colwell, R.K.; Danielsen, F.; Evengård, B.; et al. Biodiversity Redistribution under Climate Change: Impacts on Ecosystems and Human Well-Being. Science 2017, 355, eaai9214. [Google Scholar] [CrossRef]
- Dullinger, S.; Gattringer, A.; Thuiller, W.; Moser, D.; Zimmermann, N.E.; Guisan, A.; Willner, W.; Plutzar, C.; Leitner, M.; Mang, T.; et al. Extinction Debt of High-Mountain Plants under Twenty-First-Century Climate Change. Nat. Clim. Chang. 2012, 2, 619–622. [Google Scholar] [CrossRef]
- Salick, J.; Fang, Z.; Byg, A. Eastern Himalayan Alpine Plant Ecology, Tibetan Ethnobotany, and Climate Change. Glob. Environ. Chang. 2009, 19, 147–155. [Google Scholar] [CrossRef]
- Steinbauer, M.J.; Grytnes, J.-A.; Jurasinski, G.; Kulonen, A.; Lenoir, J.; Pauli, H.; Rixen, C.; Winkler, M.; Bardy-Durchhalter, M.; Barni, E.; et al. Accelerated Increase in Plant Species Richness on Mountain Summits Is Linked to Warming. Nature 2018, 556, 231–234. [Google Scholar] [CrossRef] [PubMed]
- Pertoldi, C.; Bach, L.A. Evolutionary Aspects of Climate-Induced Changes and the Need for Multidisciplinarity. J. Therm. Biol. 2007, 32, 118–124. [Google Scholar] [CrossRef]
- Corradi, E.; Abbet, C.; Gafner, F.; Hamburger, M.; Potterat, O. Screening of Alpine Plant Extracts as Protective Agents against UV-Induced Skin Damage. Planta Med. 2013, 79, PC3. [Google Scholar] [CrossRef]
- Descola, P. Beyond Nature and Culture; University of Chicago Press: Chicago, IL, USA, 2013. [Google Scholar]
- Ripple, W.J.; Wolf, C.; Newsome, T.M.; Barnard, P.; Moomaw, W.R. World Scientists’ Warning of a Climate Emergency. BioScience 2020, 70, 8–12. [Google Scholar] [CrossRef]
- Maffi, L. Linguistic, Cultural, and Biological Diversity. Annu. Rev. Anthropol. 2005, 34, 599–617. [Google Scholar] [CrossRef]
- Corlett, R.T. The Anthropocene Concept in Ecology and Conservation. Trends Ecol. Evol. 2015, 30, 36–41. [Google Scholar] [CrossRef] [PubMed]
- Simlai, T.; Sandbrook, C. Digital surveillance technologies in conservation and their social implications. In Conservation Technology; Wich, S., Piel, A., Eds.; Oxford University Press: Oxford, UK, 2021; pp. 239–249. [Google Scholar]
- Gavin, M.C.; McCarter, J.; Mead, A.; Berkes, F.; Stepp, J.R.; Peterson, D.; Tang, R. Defining Biocultural Approaches to Conservation. Trends Ecol. Evol. 2015, 30, 140–145. [Google Scholar] [CrossRef] [PubMed]
- Meine, C. Conservation Biology: Past and Present—Conservation Biology for All; Meine, C., Ed.; Oxford University Press: Oxford, UK, 2010; pp. 1–5. [Google Scholar]
- Lorimer, J. Wildlife in the Anthropocene: Conservation after Nature; University of Minnesota Press: Milwaukee, WI, USA, 2015; ISBN 1-4529-4429-6. [Google Scholar]
- Bain, R. Technology and State Government. Am. Sociol. Rev. 1937, 2, 860–874. [Google Scholar] [CrossRef]
- Stiegler, B. Technics and Time: The Fault of Epimetheus; Stanford University Press: Redwood City, CA, USA, 1998. [Google Scholar]
- Arthur, W.B. The Nature of Technology: What It Is and How It Evolves; Simon and Schuster: New York, NY, USA, 2009; ISBN 1-4391-6578-5. [Google Scholar]
- Jørgensen, D. Not by Human Hands: Five Technological Tenets for Environmental History in the Anthropocene. Environ. Hist. 2014, 20, 479–489. [Google Scholar] [CrossRef]
- Verbeek, P.-P. Toward a theory of technological mediation. In Technoscience and Postphenomenology; Friis, J., Crease, R.P., Eds.; The Manhattan Papers; Lexington Books: London, UK, 2015; pp. 189–203. [Google Scholar]
- Latour, B. The more manipulations, the better. In Representation in Scientific Practice Revisited; Coopmans, C., Vertesi, J., Lynch, M., Woolgar, S., Eds.; Massachusetts Institute of Technology: Cambridge, MA, USA, 2014; pp. 10–19. [Google Scholar]
- Sismondo, S. An Introduction to Science and Technology Studies; Wiley-Blackwell Chichester: Chichester, UK, 2010. [Google Scholar]
- Holmes, G.; Cavanagh, C.J. A Review of the Social Impacts of Neoliberal Conservation: Formations, Inequalities, Contestations. Geoforum 2016, 75, 199–209. [Google Scholar] [CrossRef]
- Berger-Tal, O.; Lahoz-Monfort, J.J. Conservation Technology: The next Generation. Conserv. Lett. 2018, 11, e12458. [Google Scholar] [CrossRef]
- Arneth, A.; Shin, Y.-J.; Leadley, P.W.; Rondinini, C.; Bukvareva, E.; Kolb, M.; Midgley, G.F.; Oberdorff, T.; Palomo, I.; Saito, O. Post-2020 Biodiversity Targets Need to Embrace Climate Change. Proc. Natl. Acad. Sci. USA 2020, 49, 30882–30891. [Google Scholar] [CrossRef]
- O’Connor, L.M.J.; Fugère, V.; Gonzalez, A. Evolutionary Rescue Is Mediated by the History of Selection and Dispersal in Diversifying Metacommunities. Front. Ecol. Evol. 2020, 8, 437. [Google Scholar] [CrossRef]
- IPBES. Summary for Policymakers of the Global Assessment Report on Biodiversity and Ecosystem Services of the Intergovernmental Science-Policy Plantform on Biodiversity and Ecosystem Services; IPBES Secretariat: Bonn, Germany, 2019; p. 56. [Google Scholar]
- Brey, P. Is Information Ethics Culture-Relative? Int. J. Technol. Hum. Interact. 2007, 3, 13. [Google Scholar] [CrossRef][Green Version]
- Maffi, L. Biocultural diversity and sustainability. In The Sage Handbook of Environment and Society; Sage Publishing: London, UK, 2007; pp. 267–277. [Google Scholar]
- Maffi, L.; Woodley, E. Biocultural Diversity Conservation: A Global Sourcebook; Routledge: New York, NY, USA, 2012; ISBN 1-136-54426-7. [Google Scholar]
- Upreti, B.; Tewari, L.; Tewari, A. Role of Plants Used in Religious and Cultral System by Local Inhabitnats of Sacred Forests of District Pothoragarth Kumaun Himalaya. Biolife 2017, 5, 7–11. [Google Scholar]
- Ticktin, T.; Mondragón, D.; Lopez-Toledo, L.; Dutra-Elliott, D.; Aguirre-León, E.; Hernández-Apolinar, M. Synthesis of Wild Orchid Trade and Demography Provides New Insight on Conservation Strategies. Conserv. Lett. 2020, 13, e12697. [Google Scholar] [CrossRef]
- Loh, J.; Harmon, D. Biocultural Diversity: Threatened Species, Endangered Languages; WWF Netherlands: Zeist, The Netherlands, 2014; Volume 1, pp. 30–43. [Google Scholar]
- Cámara-Leret, R.; Bascompte, J. Language Extinction Triggers the Loss of Unique Medicinal Knowledge. Proc. Natl. Acad. Sci. USA 2021, 118, e2103683118. [Google Scholar] [CrossRef]
- Mackey, B.; Claudie, D. Points of Contact: Integrating Traditional and Scientific Knowledge for Biocultural Conservation. Environ. Ethics 2015, 37, 341–357. [Google Scholar] [CrossRef]
- Sandbrook, C. The Social Implications of Using Drones for Biodiversity Conservation. Ambio 2015, 44, 636–647. [Google Scholar] [CrossRef]
- Tripathi, M. Critical Review of Some Important Medicinal and Aromatic Plants of Western Himalaya. Res. J. Pharm. Sci. 2020, 9, 7–16. [Google Scholar]
- Manuel-Navarrete, D. Double Coupling: Modeling Subjectivity and Asymmetric Organization in Social-Ecological Systems. Ecol. Soc. 2015, 20, 1–10. [Google Scholar] [CrossRef]
- Bonneau, A. Indigenous Knowledge Systems Often Overlooked in Academia. 2021. Available online: https://thediscourse.ca/okanagan/indigenous-knowledge-systems-overlooked-in-academia (accessed on 5 November 2021).
- Lewis, D.; Williams, L.; Jones, R.A. A Radical Revision of the Public Health Response to Environmental Crisis in a Warming World: Contributions of Indigenous Knowledges and Indigenous Feminist Perspectives. Can. J. Public Health 2020, 111, 897–900. [Google Scholar] [CrossRef]
- Institutional Labour Organization (ILO) C169—Indigenous and Tribal Peoples Convention; Article 1. 2021. Available online: https://www.ilo.org/dyn/normlex/en/f?p=NORMLEXPUB:12100:0::NO::P12100_ILO_CODE:C169 (accessed on 6 June 2021).
- Ouane, A.; Glanz, C. Optimising Learning, Education and Publishing in Africa: The Language Factor—A Review and Analysis of Theory and Practice in Mother-Tongue and Bilingual Education in Sub-Saharan Africa; UNESCO Institute for Lifelong Learning Press: Hamburg, Germany, 2011. [Google Scholar]
- McGregor, D. Indigenous Knowledge Systems in Environmental Governance in Canada. KULA 2021, 5, 1–10. [Google Scholar] [CrossRef]
- Turner, N.J.; Luczaj, L.; Migliorini, P.; Pieroni, A.; Dreon, A.L.; Sacchetti, L.E.; Paoletti, M.G. Edible and Tended Wild Plants, Traditional Ecological Knowledge and Agroecology. Crit. Rev. Plant Sci. 2011, 30, 198–225. [Google Scholar] [CrossRef]
- Love, T.R. Indigenous Knowledges, Priorities and Processes in Qualitative Organization and Management Research. Qual. Res. Organ. Manag. Int. J. 2020, 15, 6–20. [Google Scholar] [CrossRef]
- Batumike, R.; Imani, G.; Bisimwa, B.; Urom, C.; Mambo, H.; Kalume, J.; Kavuba, F.; Cuni-Sanchez, A. From Tree Species to Forest Services: Ethnic Differences in Lomami, Democratic Republic of the Congo. Econ. Bot. 2021, 1–12. [Google Scholar] [CrossRef]
- Whyte, K.P. Our ancestors’ dystopia now: Indigenous conservation and the anthropocene. In The Routledge Companion to the Environmental Humanities; Heise, U., Christensen, J., Niemann, M., Eds.; Routledge: New York, NY, USA, 2017; pp. 208–215. [Google Scholar]
- Reid, A.J.; Eckert, L.; Lane, J.-F.; Young, N.; Hinch, S.; Darimont, C.; Cooke, S.; Ban, N.C.; Marshall, A. “Two-Eyed Seeing”: An Indigenous Framework to Transform Fisheries Research and Management. Fish Fish. 2021, 22, 243–261. [Google Scholar] [CrossRef]
- Broadhead, L.-A.; Howard, S. Confronting the Contradictions between Western and Indigenous Science: A Critical Perspective on Two-Eyed Seeing. AlterNative Int. J. Indig. Peoples 2021, 17, 111–119. [Google Scholar] [CrossRef]
- Ross, A.; Sherman, K.P.; Snodgrass, J.G.; Delcore, H.D.; Sherman, R. Indigenous Peoples and the Collaborative Stewardship of Nature: Knowledge Binds and Institutional Conflicts; Routledge: Walnut Creek, CA, USA, 2016; ISBN 1-315-42661-7. [Google Scholar]
- Kimmerer, R. Braiding Sweetgrass: Indigenous Wisdom, Scientific Knowledge and the Teachings of Plants, 2nd ed.; Milkweed Editions: Minneapolis, MN, USA, 2020; ISBN 978-1-57131-335-5. [Google Scholar]
- Ludwig, D.; Macnaghten, P. Traditional Ecological Knowledge in Innovation Governance: A Framework for Responsible and Just Innovation. J. Responsible Innov. 2020, 7, 26–44. [Google Scholar] [CrossRef]
- Pence, V.C.; Ballesteros, D.; Walters, C.; Reed, B.M.; Philpott, M.; Dixon, K.W.; Pritchard, H.W.; Culley, T.M.; Vanhove, A.-C. Cryobiotechnologies: Tools for Expanding Long-Term Ex Situ Conservation to All Plant Species. Biol. Conserv. 2020, 250, 108736. [Google Scholar] [CrossRef]
- Barnes, J.C.; Delborne, J.A. The Politics of Genetic Technoscience for Conservation: The Case of Blight-Resistant American Chestnut. Environ. Plan. E Nat. Space 2021, 25148486211024910. [Google Scholar] [CrossRef]
- Rossi, J. Genes Are Not Information: Rendering Plant Genetic Resources Untradeable through Genetic Restoration Practices. Geoforum 2014, 55, 66–75. [Google Scholar] [CrossRef]
- Jacobs, D.F.; Dalgleish, H.J.; Nelson, C.D. A Conceptual Framework for Restoration of Threatened Plants: The Effective Model of American Chestnut (Castanea Dentata) Reintroduction. New Phytol. 2013, 197, 378–393. [Google Scholar] [CrossRef]
- Barnhill-Dilling, S.K.; Rivers, L.; Delborne, J.A. Rooted in Recognition: Indigenous Environmental Justice and the Genetically Engineered American Chestnut Tree. Soc. Nat. Resour. 2020, 33, 83–100. [Google Scholar] [CrossRef]
- Barnes, J.C. Engineering Conservation: The Biogeography, Biopolitics, and Biotechnology of American Chestnut Restoration. Ph.D. Dissertation, North Carolina State University, Raleigh, NC, USA, 2018. [Google Scholar]
- Foster, A.D.; Rosenzweig, M.R. Microeconomics of Technology Adoption. Annu. Rev. Econ. 2010, 2, 395–424. [Google Scholar] [CrossRef] [PubMed]
- Delborne, J.A.; Binder, A.R.; Rivers, L.; Barnes, J.C.; Barnhill-Dilling, S.; George, D.; Sudweeks, J. Biotechnology, the American Chestnut Tree, and Public Engagement (Workshop Report); Genetic Engineering and Society Center, North Carolina State University: Raleigh, NC, USA, 2018. [Google Scholar]
- Juiling, S.; Leon, S.; Jumian, J.; Tsen, S.; Lee, Y.L.; Khoo, E.; Sugau, J.B.; Nilus, R.; Pereira, J.; Damit, A.; et al. Conservation Assessment and Spatial Distribution of Endemic Orchids in Sabah, Borneo. Nat. Conserv. Res. 2020, 5, 136–144. [Google Scholar] [CrossRef]
- Sharrock, S.; Hoft, R.; Dias, B.F.D.S. An Overview of Recent Progress in the Implementation of the Global Strategy for Plant Conservation—A Global Perspective. Rodriguésia 2018, 69, 1489–1511. [Google Scholar] [CrossRef]
- Wandersee, J.H.; Schussler, E.E. Preventing Plant Blindness. Am. Biol. Teach. 1999, 61, 82–86. [Google Scholar] [CrossRef]
- Shannon, L. Plant Speak. PAN 2012, 1, 65–71. [Google Scholar] [CrossRef]
- Pouteau, S. Plants as Open Beings: From Aesthetics to plant–human ethics. In Plant Ethics; Kallhof, A., Di Paola, M., Schorgenhumer, M., Eds.; Routledge: London, UK, 2018; pp. 82–97. ISBN 1-315-11439-9. [Google Scholar]
- Balding, M.; Williams, K. Plant Blindness and the Implications for Plant Conservation. Conserv. Biol. 2016, 30, 1192–1199. [Google Scholar] [CrossRef] [PubMed]
- Margulies, J.; Bullough, L.-A.; Hinsley, A.; Ingram, D.; Cowell, C.; Goettsch, B.; Klitgard, B.; Lavorgna, A.; Sinovas, P.; Phelps, J. Illegal Wildlife Trade and the Persistence of “Plant Blindness”. Plants People Planet 2019, 1, 173–182. [Google Scholar] [CrossRef]
- Walters, R. Eco-crime and green activism. In Forging a Scio-Legal Approach to Environmental Harms; Bergin, T., Orlando, E., Eds.; Routledge: Oxon, UK, 2017; pp. 220–236. [Google Scholar]
- Cousins, S.R.; Williams, V.L.; Witkowski, E.T.F. Uncovering the Cycad Taxa (Encephalartos Species) Traded for Traditional Medicine in Johannesburg and Durban, South Africa. S. Afr. J. Bot. 2012, 78, 129–138. [Google Scholar] [CrossRef]
- Kenny, J.G. Conservation and Murder: The Plight of Indigenous Land Defenders in Mexico, Costa Rica, New Zealand and the Philippines. Ph.D. Thesis, Fordham University, New York, NY, USA, 2021. [Google Scholar]
- Medin, D.L.; García, S.G. Conceptualizing Agency: Folkpsychological and Folkcommunicative Perspectives on Plants. Cognition 2017, 162, 103–123. [Google Scholar] [CrossRef]
- Ojalehto, B.L.; Medin, D.L.; Horton, W.S.; Garcia, S.G.; Kays, E.G. Seeing Cooperation or Competition: Ecological Interactions in Cultural Perspectives. Top. Cogn. Sci. 2015, 7, 624–645. [Google Scholar] [CrossRef]
- Alpert, P.; Bone, E.; Holzapfel, C. Invasiveness, Invasibility and the Role of Environmental Stress in the Spread of Non-Native Plants. Perspect. Plant Ecol. Evol. Syst. 2000, 3, 52–66. [Google Scholar] [CrossRef]
- Pleasant, J.M. Food Yields and Nutrient Analyses of the Three Sisters: A Haudenosaunee Cropping System. Ethnobiol. Lett. 2016, 7, 87–98. [Google Scholar]
- Van Deynze, A.; Zamora, P.; Delaux, P.-M.; Heitmann, C.; Jayaraman, D.; Rajasekar, S.; Graham, D.; Maeda, J.; Gibson, D.; Schwartz, K.D.; et al. Nitrogen Fixation in a Landrace of Maize Is Supported by a Mucilage-Associated Diazotrophic Microbiota. PLoS Biol. 2018, 16, e2006352. [Google Scholar] [CrossRef] [PubMed]
- Balick, M.J.; Cox, P.A. Plants, People, and Culture: The Science of Ethnobotany, 2nd ed.; Garland Science CRC Press: Boca Raton, FL, USA, 2020; ISBN 1-00-009848-6. [Google Scholar]
- Sandbrook, C.; Luque-Lora, R.; Adams, W.M. Human Bycatch: Conservation Surveillance and the Social Implications of Camera Traps. Conserv. Soc. 2018, 16, 493–504. [Google Scholar] [CrossRef]
- Bonga, J. Can Explant Choice Help Resolve Recalcitrance Problems in in Vitro Propagation, a Problem Still Acute Especially for Adult Conifers? Trees 2017, 31, 781–789. [Google Scholar] [CrossRef]
- Carlín, A.P.; Tafoya, F.; Solís, A.G.A.; Pérez-Molphe-Balch, E. Effects of Different Culture Media and Conditions on Biomass Production of Hairy Root Cultures in Six Mexican Cactus Species. In Vitro Cell. Dev. Biol.-Plant 2015, 51, 332–339. [Google Scholar] [CrossRef]
- CITES Turbinicarpus. Available online: https://cites.org/eng/taxonomy/term/9622 (accessed on 17 November 2021).
- Muneta, J.D. Peyote Crisis Confronting Modern Indigenous Peoples: The Declining Peyote Population and the Demand for Conservation. Am. Indian Law J. 2020, 9, 135. [Google Scholar]
- Fitz Maurice, W.A.; Sanchez, E.; Fitz Maurice, B.; Guadalupe Martinez, J. Turbinicarpus Alonsoi; The IUCN Red List of Threatened Species; International Union for the Conservation of Nature and Natural Resources. Available online: https://www.iucn.org/resources/conservation-tools/iucn-red-list-threatened-species (accessed on 9 November 2021).
- Stephens, L.; Fuller, D.; Boivin, N.; Rick, T.; Gauthier, N.; Kay, A.; Marwick, B.; Armstrong, C.G.; Barton, C.M.; Denham, T. Archaeological Assessment Reveals Earth’s Early Transformation through Land Use. Science 2019, 365, 897–902. [Google Scholar] [CrossRef] [PubMed]
- Larson, G.; Piperno, D.R.; Allaby, R.G.; Purugganan, M.D.; Andersson, L.; Arroyo-Kalin, M.; Barton, L.; Climer Vigueira, C.; Denham, T.; Dobney, K.; et al. Current Perspectives and the Future of Domestication Studies. Proc. Natl. Acad. Sci. USA 2014, 111, 6139. [Google Scholar] [CrossRef]
- Corlett, R.T. A Bigger Toolbox: Biotechnology in Biodiversity Conservation. Trends Biotechnol. 2017, 35, 55–65. [Google Scholar] [CrossRef]
- Lee, K.; Seo, P.J. Dynamic Epigenetic Changes during Plant Regeneration. Trends Plant Sci. 2018, 23, 235–247. [Google Scholar] [CrossRef] [PubMed]
- Engelmann, F. Use of Biotechnologies for the Conservation of Plant Biodiversity. In Vitro Cell. Dev. Biol.-Plant 2011, 47, 5–16. [Google Scholar] [CrossRef]
- Shahzad, A.; Parveen, S.; Sharma, S.; Shaheen, A.; Saeed, T.; Yadav, V.; Akhtar, R.; Ahmad, Z.; Upadhyay, A. Plant tissue culture: Applications in plant improvement and conservation. In Plant Biotechnology: Principles and Applications; Abdin, M., Kiran, U., Kamaluddin, A.A., Eds.; Springer: Singapore, 2017; pp. 37–72. [Google Scholar]
- Saxena, A.; Shukla, M.R.; Saxena, P.K. Synthetic seeds: Relevance to endangered germplasm conservation in vitro. In Synthetic Seeds; Faisal, M., Alatar, A.A., Eds.; Springer: Cham, Switzerland, 2019; pp. 21–60. [Google Scholar]
- Thorpe, T.A. History of Plant Tissue Culture. Mol. Biotechnol. 2007, 37, 169–180. [Google Scholar] [CrossRef] [PubMed]
- Ashmore, S.E.; Hamilton, K.N.; Offord, C.A. Conservation Technologies for Safeguarding and Restoring Threatened Flora: Case Studies from Eastern Australia. In Vitro Cell. Dev. Biol.-Plant 2011, 47, 99–109. [Google Scholar] [CrossRef]
- Coelho, N.; Gonçalves, S.; Romano, A. Endemic Plant Species Conservation: Biotechnological Approaches. Plants 2020, 9, 345. [Google Scholar] [CrossRef]
- Grigoriadou, K.; Krigas, N.; Sarropoulou, V.; Papanastasi, K.; Tsoktouridis, G.; Maloupa, E. In Vitro Propagation of Medicinal and Aromatic Plants: The Case of Selected Greek Species with Conservation Priority. In Vitro Cell. Dev. Biol. 2019, 55, 635–646. [Google Scholar] [CrossRef]
- Cui, Y.; Deng, Y.; Zheng, K.; Hu, X.; Zhu, M.; Deng, X.; Xi, R. An Efficient Micropropagation Protocol for an Endangered Ornamental Tree Species (Magnolia Sirindhorniae Noot. & Chalermglin) and Assessment of Genetic Uniformity through DNA Markers. Sci. Rep. 2019, 9, 9634. [Google Scholar] [CrossRef]
- Komakech, R.; Kim, J.; Kim, W.J.; Omujal, F.; Yang, S.; Moon, B.C.; Okello, D.; Rahmat, E.; Nambatya Kyetune, G.; Matsabisa, M.G. A Micropropagation Protocol for the Endangered Medicinal Tree Prunus Africana (Hook f.) Kalkman: Genetic Fidelity and Physiological Parameter Assessment. Front. Plant Sci. 2020, 11, 1871. [Google Scholar] [CrossRef]
- Singh, R.K.; Anandhan, S.; García-Pérez, L.M.; Ruiz-May, E.; Nava Pérez, E.; Quiroz-Figueroa, F.R. An Efficient Protocol for in Vitro Propagation of the Wild Legume Cicer microphyllum Benth., a Crop Wild Relative of Chickpea (Cicer arietinum L.). In Vitro Cell. Dev. Biol.-Plant 2019, 55, 9–14. [Google Scholar] [CrossRef]
- Benson, E. Plant Conservation Biotechnology; CRC Press: Boca Raton, FL, USA, 1999; ISBN 1-4822-7303-9. [Google Scholar]
- Bi, W.; Saxena, A.; Ayyanath, M.-M.; Harpur, C.; Shukla, M.R.; Saxena, P.K. Conservation, Propagation, and Redistribution (CPR) of Hill’s Thistle: Paradigm for Plant Species at Risk. Plant Cell Tissue Organ Cult. 2021, 145, 75–88. [Google Scholar] [CrossRef]
- Panis, B.; Nagel, M.; Van den Houwe, I. Challenges and Prospects for the Conservation of Crop Genetic Resources in Field Genebanks, in In Vitro Collections and/or in Liquid Nitrogen. Plants 2020, 9, 1634. [Google Scholar] [CrossRef] [PubMed]
- Reed, B.M. Plant Cryopreservation: A Continuing Requirement for Food and Ecosystem Security. In Vitro Cell. Dev. Biol.-Plant 2017, 53, 285–288. [Google Scholar] [CrossRef]
- Popova, E.; Shukla, M.R.; Kim, H.H.; Saxena, P.K. Plant Cryopreservation for biotechnology and breeding. In Advances in Plant Breeding Strategies: Breeding, Biotechnology and Molecular Tools; Al-Khayri, J., Jain, S., Johnson, D., Eds.; Springer: Berlin, Germany, 2015; pp. 63–93. [Google Scholar]
- Salama, A.; Popova, E.; Jones, M.P.; Shukla, M.R.; Fisk, N.S.; Saxena, P.K. Cryopreservation of the Critically Endangered Golden Paintbrush (Castilleja levisecta Greenm.): From Nature to Cryobank to Nature. In Vitro Cell. Dev. Biol.-Plant 2018, 54, 69–78. [Google Scholar] [CrossRef]
- Rathwell, R.; Popova, E.; Shukla, M.R.; Saxena, P.K. Development of Cryopreservation Methods for Cherry Birch (Betula lenta L.), an Endangered Tree Species in Canada. Can. J. For. Res. 2016, 46, 1284–1292. [Google Scholar] [CrossRef]
- Whiteley, S.E.; Bunn, E.; Menon, A.; Mancera, R.L.; Turner, S.R. Ex Situ Conservation of the Endangered Species Androcalva Perlaria (Malvaceae) by Micropropagation and Cryopreservation. Plant Cell Tissue Organ Cult. 2016, 125, 341–352. [Google Scholar] [CrossRef]
- Popova, E.V.; Shukla, M.R.; McIntosh, T.; Saxena, P.K. In Vitro and Cryobiotechnology Approaches to Safeguard Lupinus rivularis Douglas Ex Lindl., an Endangered Plant in Canada. Agronomy 2021, 11, 37. [Google Scholar] [CrossRef]
- Barnhill-Dilling, S.K.; Delborne, J.A. The Genetically Engineered American Chestnut Tree as Opportunity for Reciprocal Restoration in Haudenosaunee Communities. Biol. Conserv. 2019, 232, 1–7. [Google Scholar] [CrossRef]
- Sharma, N. In Vitro Conservation of Gentiana Kurroo Royle: An Indigenous Threatened Medicinal Plant. Indian J. Plant Genet. Resour. 2001, 14, 99–100. [Google Scholar]
- McComb, J.; Bennett, I.; Tonkin, C. In Vitro Propagation of Eucalyptus Species. In Tissue Culture of Australian Plants; Taki, A., Williams, R., Eds.; University of New England: Armidale, NSW, Australia, 1996; pp. 112–156. [Google Scholar]
- Reshi, N.A.; Sudarshana, M.; Girish, H. In Vitro Micropropagation of Rhinacanthus nasutus (L) Kurz. Int. J. Biodivers. Conserv. 2018, 10, 357–364. [Google Scholar] [CrossRef]
- Daniëls, C.W. A Study of the Propagation and Cultivation of Gethyllis multifolia and G. villosa. Master’s Thesis, Cape Peninsula University of Technology, Cape Town, Western Cape, South Africa, June 2007. [Google Scholar]
- Witbooi, H. In Vitro Propagation of Agathosma Betulina an Indigenous Plant of Economic Importance. Master’s Thesis, Cape Peninsula University of Technology, Cape Town, Western Cape, South Africa, June 2013. [Google Scholar]
- Mridula, M.R.; Nair, A.S. Rapid Micro-Propagation of Wrightia Tinctoria (ROXB.) R Br: A Medicinal Tree. Int. J. Bot. Stud. 2018, 3, 126–131. [Google Scholar]
- Supinrach, S.; Supinrach, I. Study of cuttings and cutting environment of dutchman’s pipe (Aristolochia ringens Vahl.). In Proceedings of the 52nd Kasetsart University Annual Conference, Kasetsart University, Bankok, Thailand, 7 February 2014; pp. 355–362. [Google Scholar]
- Chinnadurai, V.; Viswanathan, P.; Kalimuthu, K.; Vanitha, A.; Ranjitha, V.; Pugazhendhi, A. Comparative Studies of Phytochemical Analysis and Pharmacological Activities of Wild and Micropropagated Plant Ethanol Extracts of Manihot Esculenta. Biocatal. Agric. Biotechnol. 2019, 19, 101166. [Google Scholar] [CrossRef]
- Barron, R. In Vitro Regeneration, Rooting, and Cloning of Artemisia Tridentata. Master’s Thesis, Boise State University, Boise, ID, USA, 6 July 2020. [Google Scholar]
- He, T.; Xu, J.; Yang, L.; Wang, H. An Efficient Method for Plant Regeneration from Calli of Swertia Mussotii, an Endangered Medicinal Herb. Am. J. Plant Sci. 2012, 3, 904–908. [Google Scholar] [CrossRef]
- Bose, B.; Kumaria, S.; Choudhury, H.; Tandon, P. Assessment of Genetic Homogeneity and Analysis of Phytomedicinal Potential in Micropropagated Plants of Nardostachys Jatamansi, a Critically Endangered, Medicinal Plant of Alpine Himalayas. Plant Cell Tissue Organ Cult. 2016, 124, 331–349. [Google Scholar] [CrossRef]
- Monteuuis, O.; Doulbeau, S.; Verdeil, J.-L. DNA Methylation in Different Origin Clonal Offspring from a Mature Sequoiadendron Giganteum Genotype. Trees 2008, 22, 779. [Google Scholar] [CrossRef]
- Murch, S.J.; Ragone, D.; Shi, W.L.; Alan, A.R.; Saxena, P.K. In vitro conservation and sustained production of breadfruit (artocarpus altilis, moraceae): Modern technologies for a traditional tropical crop. In Protocols for Micropropagation of Woody Trees and Fruits; Jain, S., Hagman, H., Eds.; Springer: Dordrecht, The Netherlands, 2007; Volume 95, pp. 279–288. ISBN 978-1-4020-6352-7. [Google Scholar]
- Kim, J.; Soh, S.Y.; Bae, H.; Nam, S.-Y. Antioxidant and Phenolic Contents in Potatoes (Solanum tuberosum L.) and Micropropagated Potatoes. Appl. Biol. Chem. 2019, 62, 1–9. [Google Scholar] [CrossRef]
- Vollmer, R.; Espirilla, J.; Villagaray, R.; Cardenas, J.; Castro, M.; Sanchez, J.C.; Anglin, N.L. Cryopreservation of Potato Shoot Tips for Long-Term Storage. In Solanum Tuberosum, Methods and Protocols; Dobnik, D., Gruden, K., Ramsak, Z., Coll, A., Eds.; Humana Press: Totowa, NJ, USA, 2021; pp. 21–54. [Google Scholar]
- Uchendu, E.E.; Shukla, M.; Saxena, P.K.; Keller, J.E.R. Cryopreservation of Potato Microtubers: The Critical Roles of Sucrose and Desiccation. Plant Cell Tissue Organ Cult. 2016, 124, 649–656. [Google Scholar] [CrossRef]
- Parra-Rondinel, F.; Casas, A.; Begazo, D.; Paco, A.; Márquez, E.; Cruz, A.; Segovia, J.; Torres-García, I.; Zarazúa, M.; Lizárraga, L. Natural and Cultural Processes Influencing Gene Flow Among Wild (Atoq Papa), Weedy (Araq Papa and k’ipa Papa), and Crop Potatoes in the Andean Region of Southern Peru. Front. Ecol. Evol. 2021, 9, 327. [Google Scholar] [CrossRef]
- Fretz, A.; Jahne, A.; Lorz, H. Cryopreservation of Embryogenic Suspension Cultures of Barley (Hordeum vulgare L.). Botanica Acta 1992, 105, 140–145. [Google Scholar] [CrossRef]
- da Silva, P.; Contim, L.; de Freitas, D.; Aride, P.; dos Santos, A. In Vitro Establishment of Kapok Tree (Ceiba pentandra L. Gaertn) Apical Shoots. Sci. Agrar. 2010, 11, 437–443. [Google Scholar]
- Usman, I.S.; Abdulmalik, M. Cryopreservation of Embryonic Axes of Maize (Zea mays L.) by Vitrification Protocol. Afr. J. Biotechnol. 2010, 9, 8955–8957. [Google Scholar]
- Perez, J.; Araya-Valverde, E.; Garro, G.; Abdelnour-Esquivel, A. Analysis of Stress Indicators during Cryopreservation of Seeds of Landrace Maize (Zea Mays). CryoLetters 2017, 38, 445–454. [Google Scholar]
- Rossetto, M.; Yap, J.-Y.S.; Lemmon, J.; Bain, D.; Bragg, J.; Hogbin, P.; Gallagher, R.; Rutherford, S.; Summerell, B.; Wilson, T.C. A Conservation Genomics Workflow to Guide Practical Management Actions. Glob. Ecol. Conserv. 2021, 26, e01492. [Google Scholar] [CrossRef]
- Sheikholeslami, B.; Shukla, M.R.; Turi, C.E.; Harpur, C.; Saxena, P.K. Saving Threatened Plant Species: Reintroduction of Hill’s Thistle (Cirsium Hillii (Canby) Fernald) to Its Natural Habitat. PLoS ONE 2020, 15, e0231741. [Google Scholar] [CrossRef] [PubMed]
- Cruz-Cruz, C.A.; González-Arnao, M.T.; Engelmann, F. Biotechnology and Conservation of Plant Biodiversity. Resources 2013, 2, 73–95. [Google Scholar] [CrossRef]
- Wang, M.-R.; Chen, L.; Teixeira da Silva, J.A.; Volk, G.M.; Wang, Q.-C. Cryobiotechnology of Apple (Malus spp.): Development, Progress and Future Prospects. Plant Cell Rep. 2018, 37, 689–709. [Google Scholar] [CrossRef] [PubMed]
- Reiter, N.; Whitfield, J.; Pollard, G.; Bedggood, W.; Argall, M.; Dixon, K.; Davis, B.; Swarts, N. Orchid Re-Introductions: An Evaluation of Success and Ecological Considerations Using Key Comparative Studies from Australia. Plant Ecol. 2016, 217, 81–95. [Google Scholar] [CrossRef]
- Bunn, E.; Turner, S.; Panaia, M.; Dixon, K.W. The Contribution of in Vitro Technology and Cryogenic Storage to Conservation of Indigenous Plants. Aust. J. Bot. 2007, 55, 345–355. [Google Scholar] [CrossRef]
- Espinosa-Leal, C.A.; Puente-Garza, C.A.; García-Lara, S. In Vitro Plant Tissue Culture: Means for Production of Biological Active Compounds. Planta 2018, 248, 1–18. [Google Scholar] [CrossRef]
- Shaw, R.; Greggor, A.; Plotnik, J. The Challenges of Replicating Research on Endangered Species. Anim. Behav. Cogn. 2021, 8, 240–246. [Google Scholar] [CrossRef]
- Sandbrook, C.; Fisher, J.A.; Holmes, G.; Luque-Lora, R.; Keane, A. The Global Conservation Movement Is Diverse but Not Divided. Nat. Sustain. 2019, 2, 316–323. [Google Scholar] [CrossRef]
- Ryder, O.A. Cloning Advances and Challenges for Conservation. Trends Biotechnol. 2002, 20, 231–232. [Google Scholar] [CrossRef]
- Snaddon, J.; Petrokofsky, G.; Jepson, P.; Willis, K.J. Biodiversity Technologies: Tools as Change Agents. Biol. Lett. 2013, 9, 20121029. [Google Scholar] [CrossRef] [PubMed]
- Zimov, S. Pleistocene Park: Return of the Mammoth’s Ecosystem. Science 2005, 308, 796–798. [Google Scholar] [CrossRef] [PubMed]
- Kordyum, E.; Hasenstein, K.H. Plant Biology for Space Exploration—Building on the Past, Preparing for the Future. Life Sci. Space Res. 2021, 29, 1–7. [Google Scholar] [CrossRef]
- Preil, W. In vitro propagation and breeding of ornamental plants: Advantages and disadvantages of variability. In Genetic Manipulation in Plant Breeding; Preil, W., Ed.; De Gruyter Press: Ahrensburg, Germany, 2019; pp. 377–404. ISBN 3-11-087194-7. [Google Scholar]
- United Nations. Convention on Biological Diversity. 1992. Available online: https://www.cbd.int/doc/legal/cbd-en.pdf (accessed on 7 October 2021).
- Antonelli, A.; Smith, R.J.; Simmonds, M.S.J. Unlocking the Properties of Plants and Fungi for Sustainable Development. Nat. Plants 2019, 5, 1100–1102. [Google Scholar] [CrossRef]
- Sas-Rolfes, M.; Challender, D.W.S.; Hinsley, A.; Veríssimo, D.; Milner-Gulland, E.J. Illegal Wildlife Trade: Scale, Processes, and Governance. Annu. Rev. Environ. Resour. 2019, 44, 201–228. [Google Scholar] [CrossRef]
- De, L.; Pathak, P. Conservation, Management and Utilization of Orchid Genetic Resources. Orchid Soc. India 2018, 32, 81–91. [Google Scholar]
- Hinsley, A.; Nuno, A.; Ridout, M.; St John, F.; Roberts, D. Estimating the Extent of CITES Noncompliance among Traders and End-consumers; Lessons from the Global Orchid Trade. Conserv. Lett. 2017, 10, 602–609. [Google Scholar] [CrossRef]
- Hinsley, A.; De Boer, H.J.; Fay, M.F.; Gale, S.W.; Gardiner, L.M.; Gunasekara, R.S.; Kumar, P.; Masters, S.; Metusala, D.; Roberts, D.L. A Review of the Trade in Orchids and Its Implications for Conservation. Bot. J. Linn. Soc. 2018, 186, 435–455. [Google Scholar] [CrossRef]
- Popova, E.; Kim, H.H.; Saxena, P.K.; Engelmann, F.; Pritchard, H.W. Frozen Beauty: The Cryobiotechnology of Orchid Diversity. Biotechnol. Adv. 2016, 34, 380–403. [Google Scholar] [CrossRef]
- Brand, R.; Fischer, J. Overcoming the Technophilia/Technophobia Split in Environmental Discourse. Environ. Politics 2013, 22, 235–254. [Google Scholar] [CrossRef]
- Solis-Castañeda, G.J.; Zamilpa, A.; Cabañas-García, E.; Bahena, S.M.; Pérez-Molphe-Balch, E.; Gómez-Aguirre, Y.A. Identification and Quantitative Determination of Feruloyl-Glucoside from Hairy Root Cultures of Turbinicarpus Lophophoroides (Werderm.) Buxb. & Backeb. (Cactaceae). In Vitro Cell. Dev. Biol.-Plant 2020, 56, 8–17. [Google Scholar] [CrossRef]
- Gallia, M.C.; Echeverri Del Sarto, J.; Bongiovanni, G.A. Sustainable and Efficient Protocols for in Vitro Germination and Antioxidants Production from Seeds of the Endangered Species Araucaria Araucana. J. Genet. Eng. Biotechnol. 2021, 19, 181. [Google Scholar] [CrossRef] [PubMed]
- Gehlot, H.S.; Tak, N.; Dagla, H.R.; Davis, T.D. Indigenous and Modern Scientific Strategies for Characterization, Conservation and Sustainable Utilization of Bio-Resources of the Indian Thar Desert. J. Arid. Land Stud. 2014, 24, 5–8. [Google Scholar]
- Goswami, D.; Dagla, H.R. In vitro culture of haloxylon recurvum and haloxylon salicornicum: Valuable source of food additives and pharmaceutical and nutritional components from extreme arid zone. In Exploring Plant Cells for the Production of Compuounds of Interest; Malik, S., Ed.; Springer: Cham, Switzerland, 2021; pp. 335–339. [Google Scholar]
- Moon, K.; Blackman, D. A Guide to Understanding Social Science Research for Natural Scientists. Conserv. Biol. 2014, 28, 1167–1177. [Google Scholar] [CrossRef]
- Kelly, R.; Mackay, M.; Nash, K.L.; Cvitanovic, C.; Allison, E.H.; Armitage, D.; Bonn, A.; Cooke, S.J.; Frusher, S.; Fulton, E.A. Ten Tips for Developing Interdisciplinary Socio-Ecological Researchers. Socio-Ecol. Pract. Res. 2019, 1, 149–161. [Google Scholar] [CrossRef]
- Romeis, J.; Meissle, M.; Brunner, S.; Tschamper, D.; Winzeler, M. Plant Biotechnology: Research behind Fences. Trends Biotechnol. 2013, 31, 222–224. [Google Scholar] [CrossRef] [PubMed]
- Moyo, M.; Bairu, M.; Amoo, S.; Van Staden, J. Plant Biotechnology in South Africa: Micropropagation Research Endeavours, Prospects and Challenges. S. Afr. J. Bot. 2011, 77, 996–1011. [Google Scholar] [CrossRef][Green Version]



{kind=link}
{kind=link}
| Scientific Name | Traditional Uses | In Vitro Method Used | Geographical Location | Reference |
|---|---|---|---|---|
| Castanea americana | food, wood | transgenic * modification * | Northeastern USA | [145] |
| Turbinicarpus sp. | medicinal, ceremonial | tissue culture *^ | Mexico | [118] |
| Gentiana kurroo | medicinal | shoot culture * | India | [146] |
| Eucalyptus spp. | medicinal | tissue culture * | Australia, Tasmania | [147] |
| Rhinacanthus nasutus | medicinal, dye | tissue culture * | Southwest Bengal | [148] |
| Gethyllis multifolia | medicinal | hydro culture * | Worcester, South Africa | [149] |
| Agathosma betulina | medicinal, food | micropropagation * | Western Cape, South Africa | [150] |
| Wrightia tinctoria | medicinal | stem cuttings * | India | [151] |
| Aristolochia ringens | medicinal | root, stem cuttings * | Nigeria | [152] |
| Manihot esculenta | medicinal, food | micropropagation * | Global distribution | [153] |
| Artemisia tridentata | medicinal, ceremonial | micropropagation * | Western North America | [154] |
| Swertia mussotii | medicinal, ceremonial | micropropagation * | Qinghai-Tibet Plateau, China | [155] |
| Nardostachys jatamansi | medicinal | micropropagation * | Himalayan region | [156] |
| Sequoiadendron giganteum | ornamental | meristem culture | Sierra Nevada, USA | [157] |
| Artocarpus altilis | medicinal, food | meristem culture *^ | Pacific Islands | [158] |
| Solanum tuberosum | food, ceremonial | nodal explant tissue culture *^ shoot tip, micro tuber cryopreservation | Global distribution | [159,160,161,162] |
| Hordeum vulgare | food | embryo cryopreservation * | SW Asia, Himalayas | [163] |
| Ceiba pentandra | medicinal, wood | apical shoot culture | Tropical forests, global distribution | [164] |
| Zea mays | food, ceremonial | embryo, seed cryopreservation | Global distribution | [165,166] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Kulak, V.; Longboat, S.; Brunet, N.D.; Shukla, M.; Saxena, P. In Vitro Technology in Plant Conservation: Relevance to Biocultural Diversity. Plants 2022, 11, 503. https://doi.org/10.3390/plants11040503
Kulak V, Longboat S, Brunet ND, Shukla M, Saxena P. In Vitro Technology in Plant Conservation: Relevance to Biocultural Diversity. Plants. 2022; 11(4):503. https://doi.org/10.3390/plants11040503
Chicago/Turabian StyleKulak, Verena, Sheri Longboat, Nicolas D. Brunet, Mukund Shukla, and Praveen Saxena. 2022. "In Vitro Technology in Plant Conservation: Relevance to Biocultural Diversity" Plants 11, no. 4: 503. https://doi.org/10.3390/plants11040503
APA StyleKulak, V., Longboat, S., Brunet, N. D., Shukla, M., & Saxena, P. (2022). In Vitro Technology in Plant Conservation: Relevance to Biocultural Diversity. Plants, 11(4), 503. https://doi.org/10.3390/plants11040503