
Integrated Physiological and Metabolomic Analyses of the Effect of Potassium Fertilizer on Citrus Fruit Splitting


Abstract
:1. Introduction
2. Results
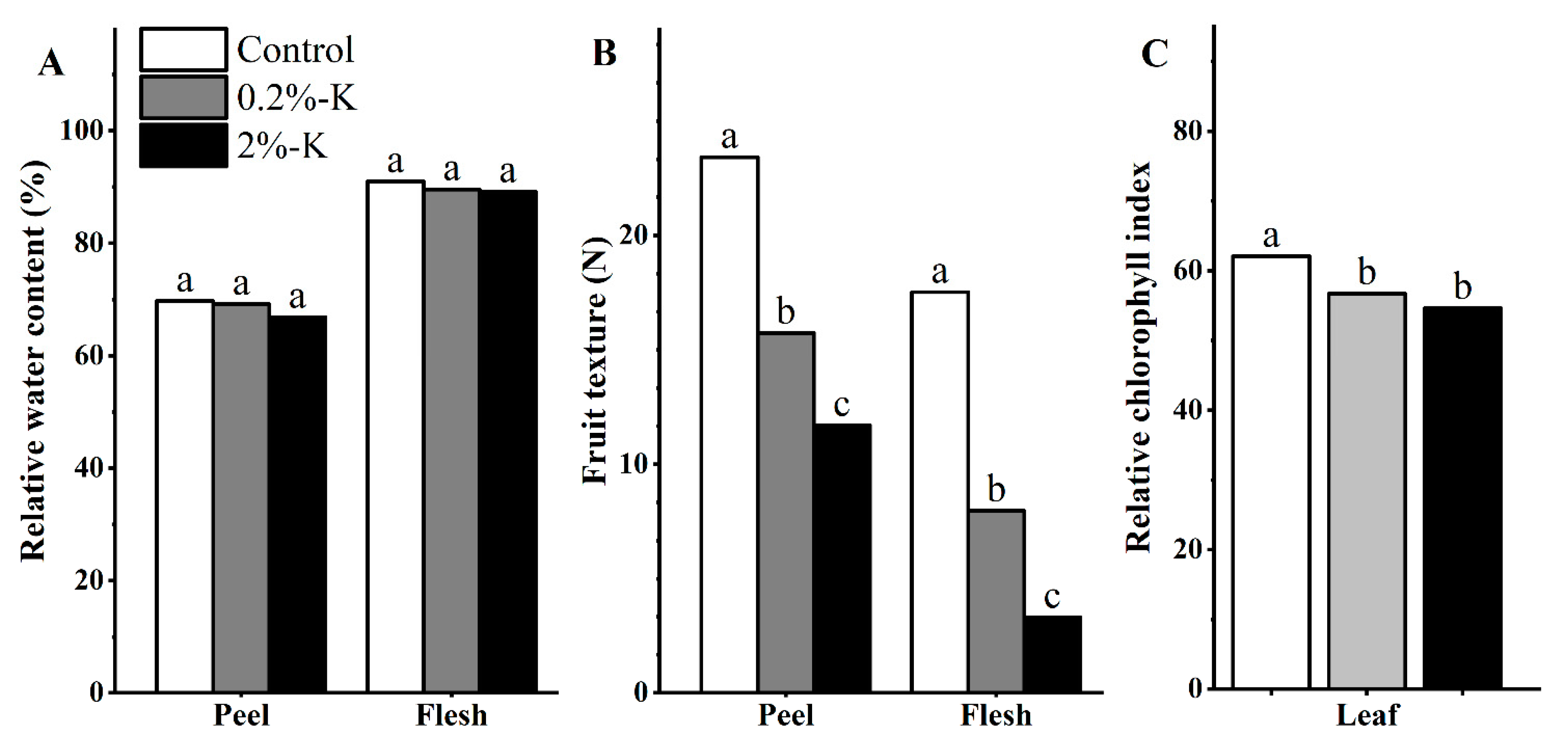
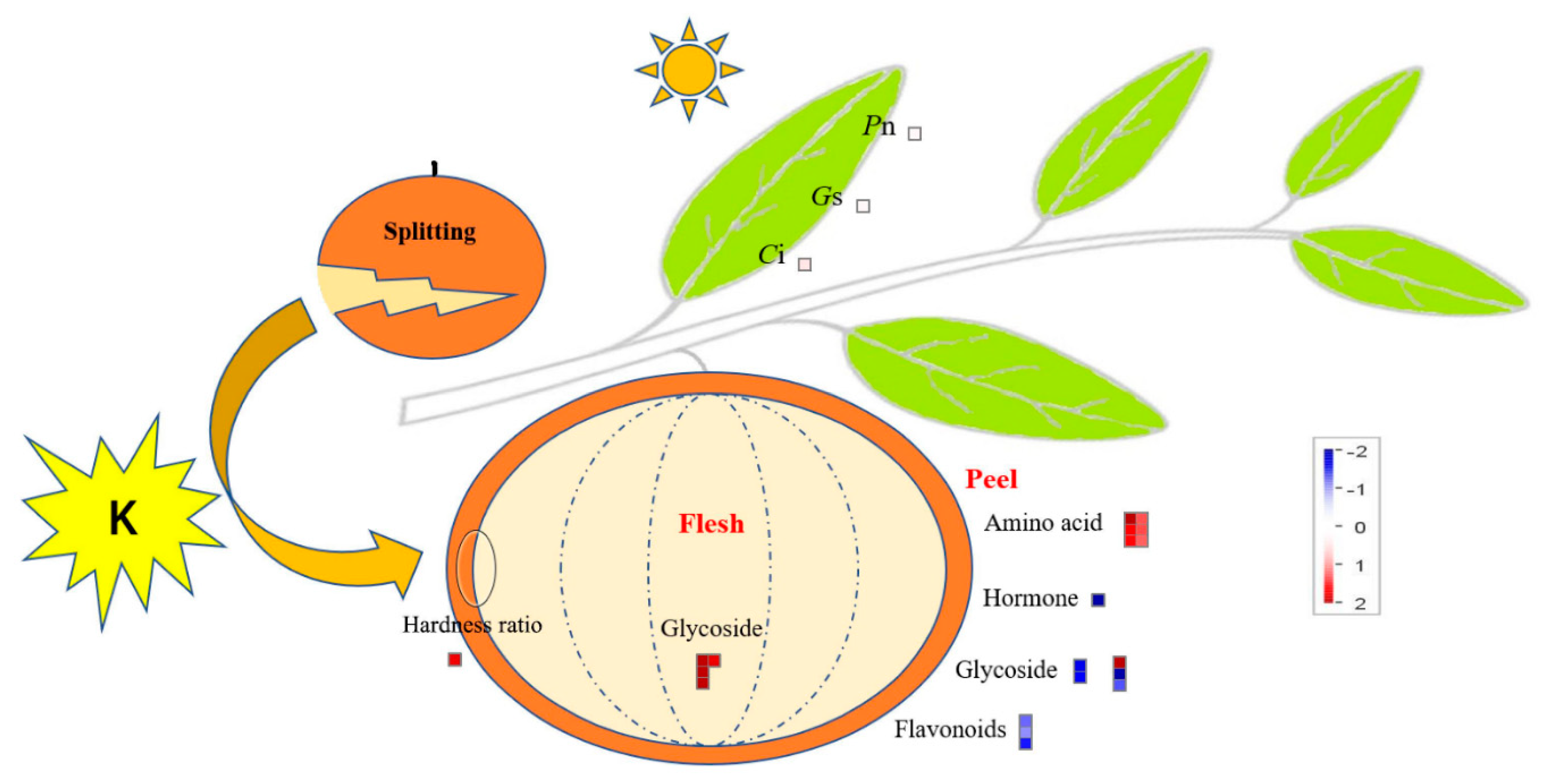
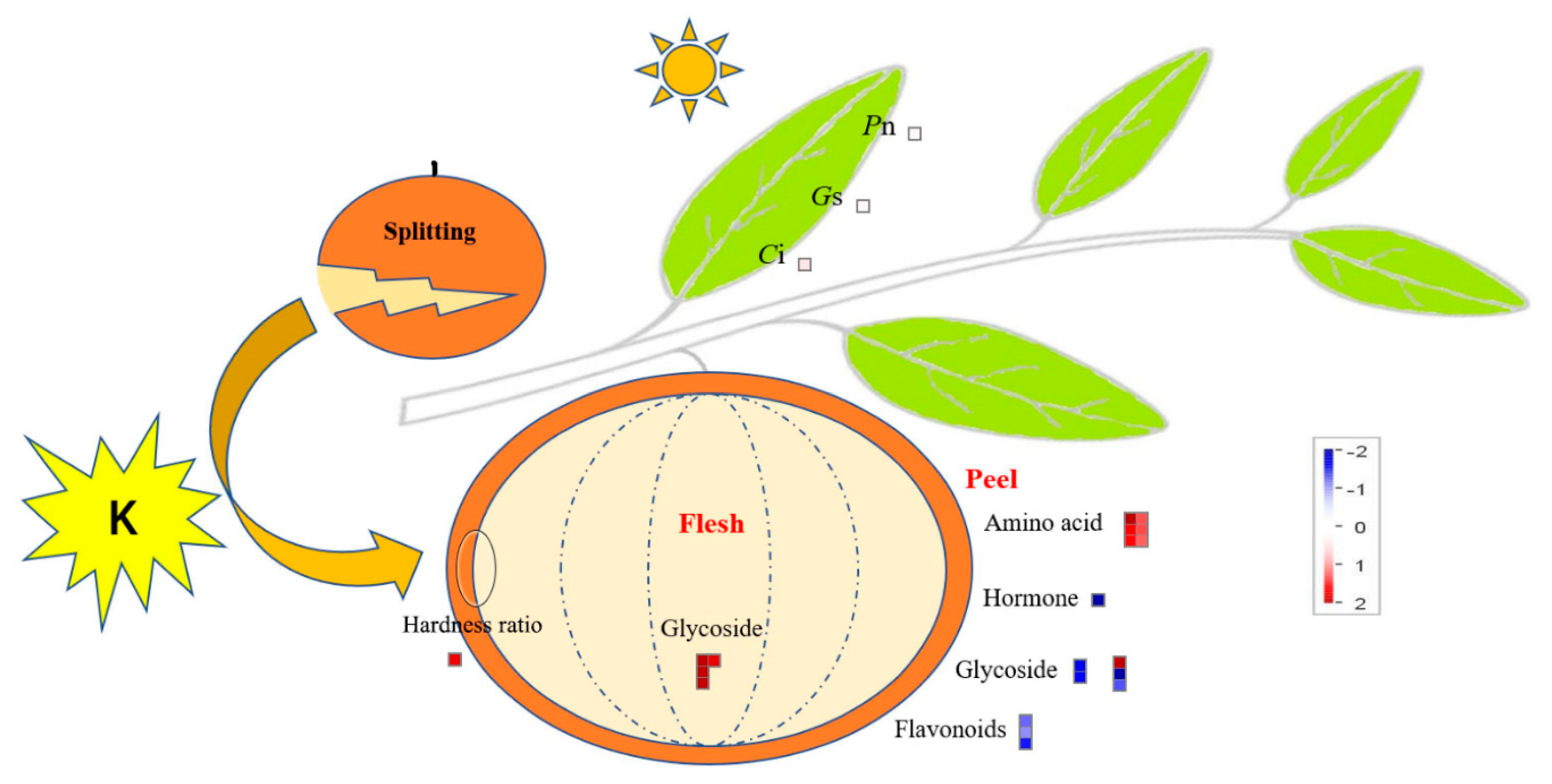
2.1. Effect of Potassium on the Quality of Citrus Fruit
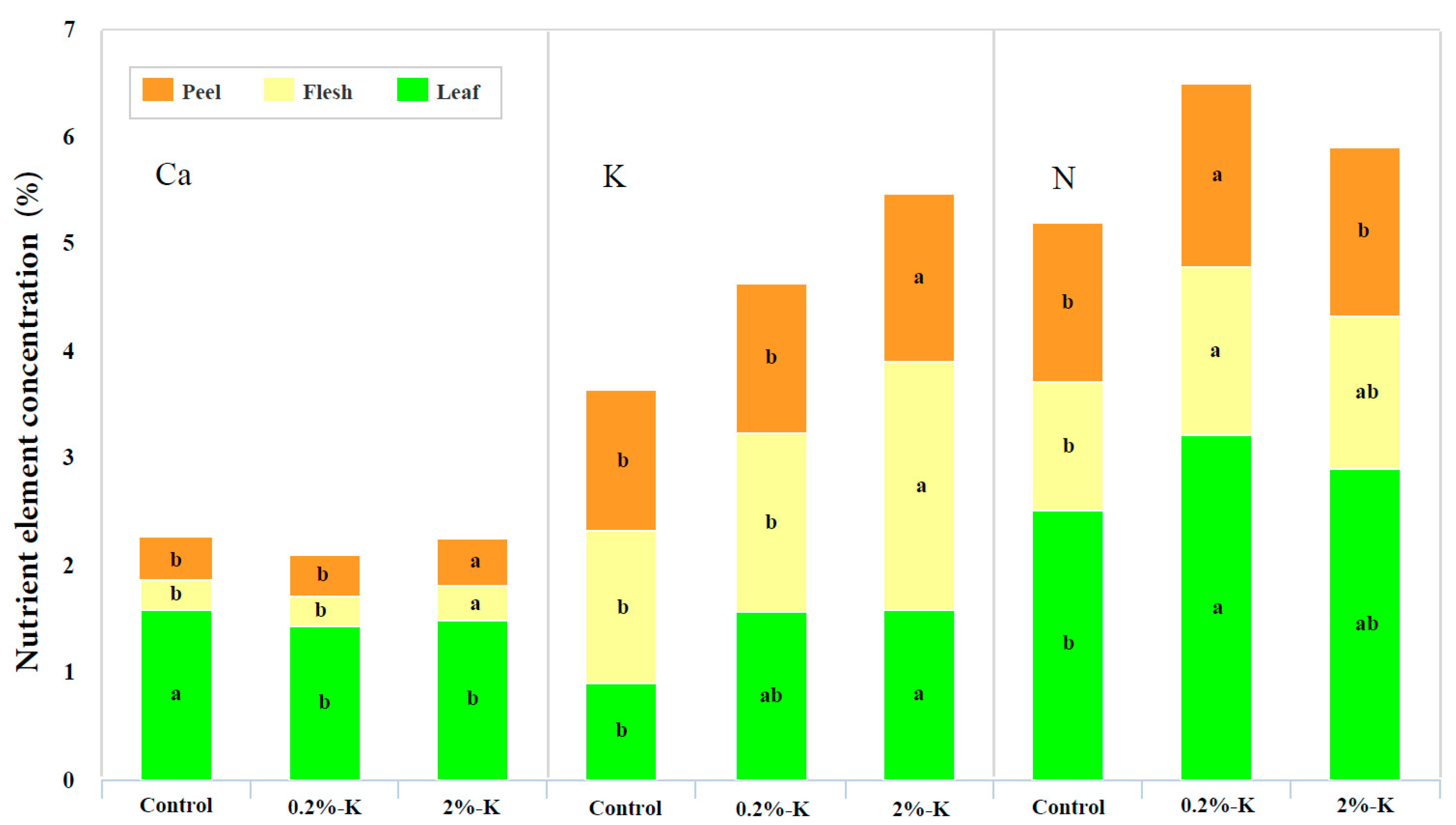
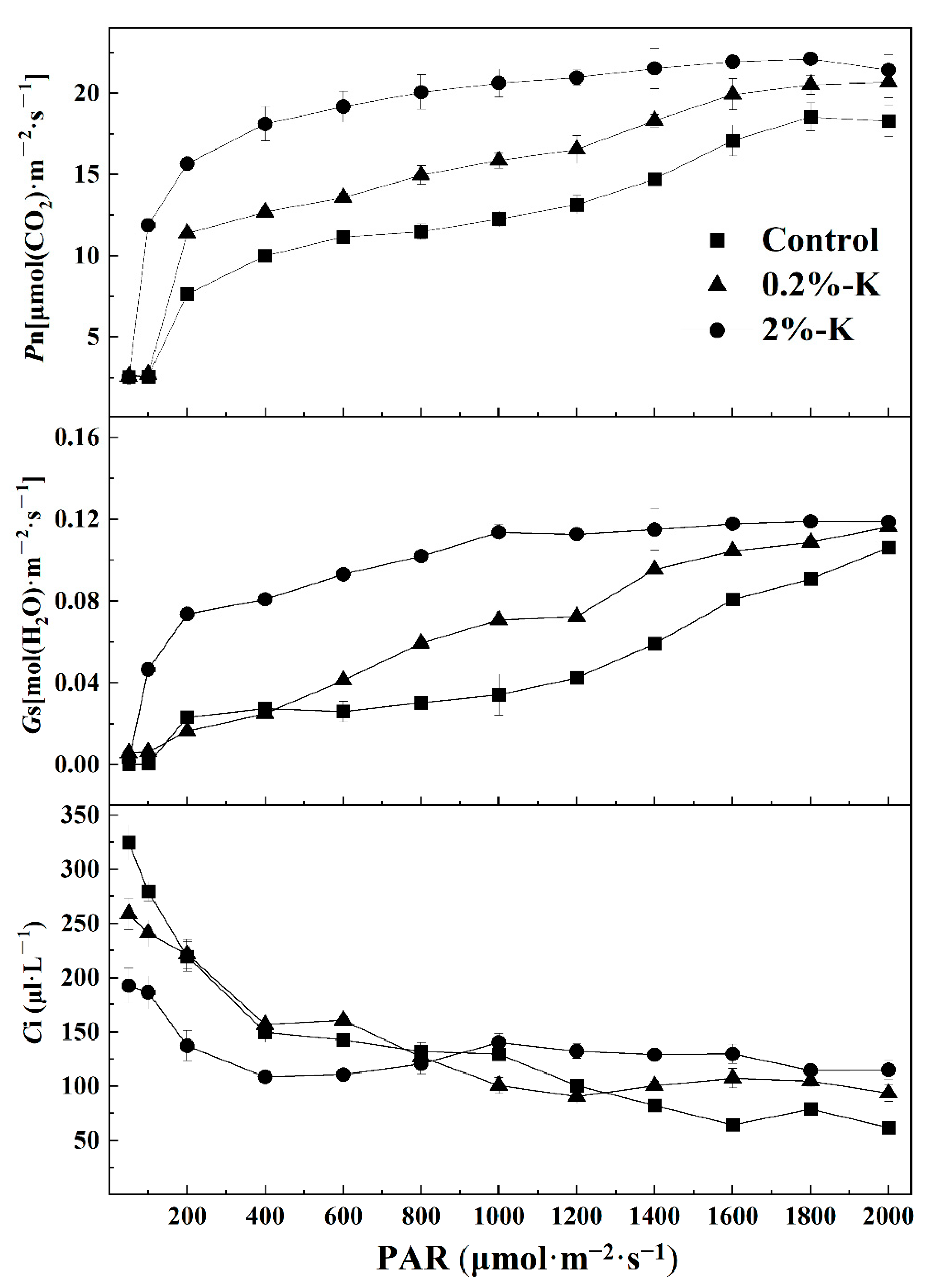
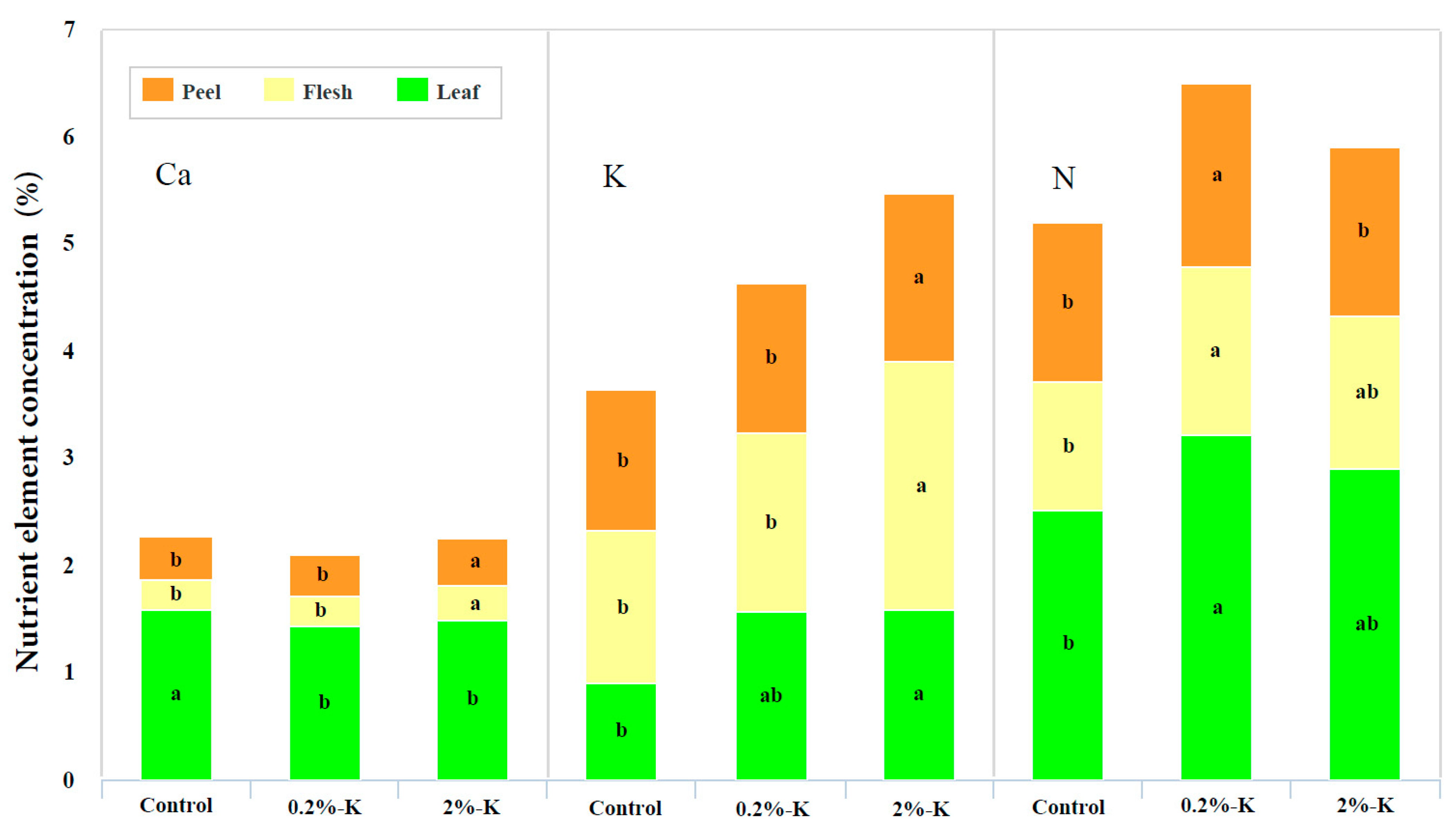
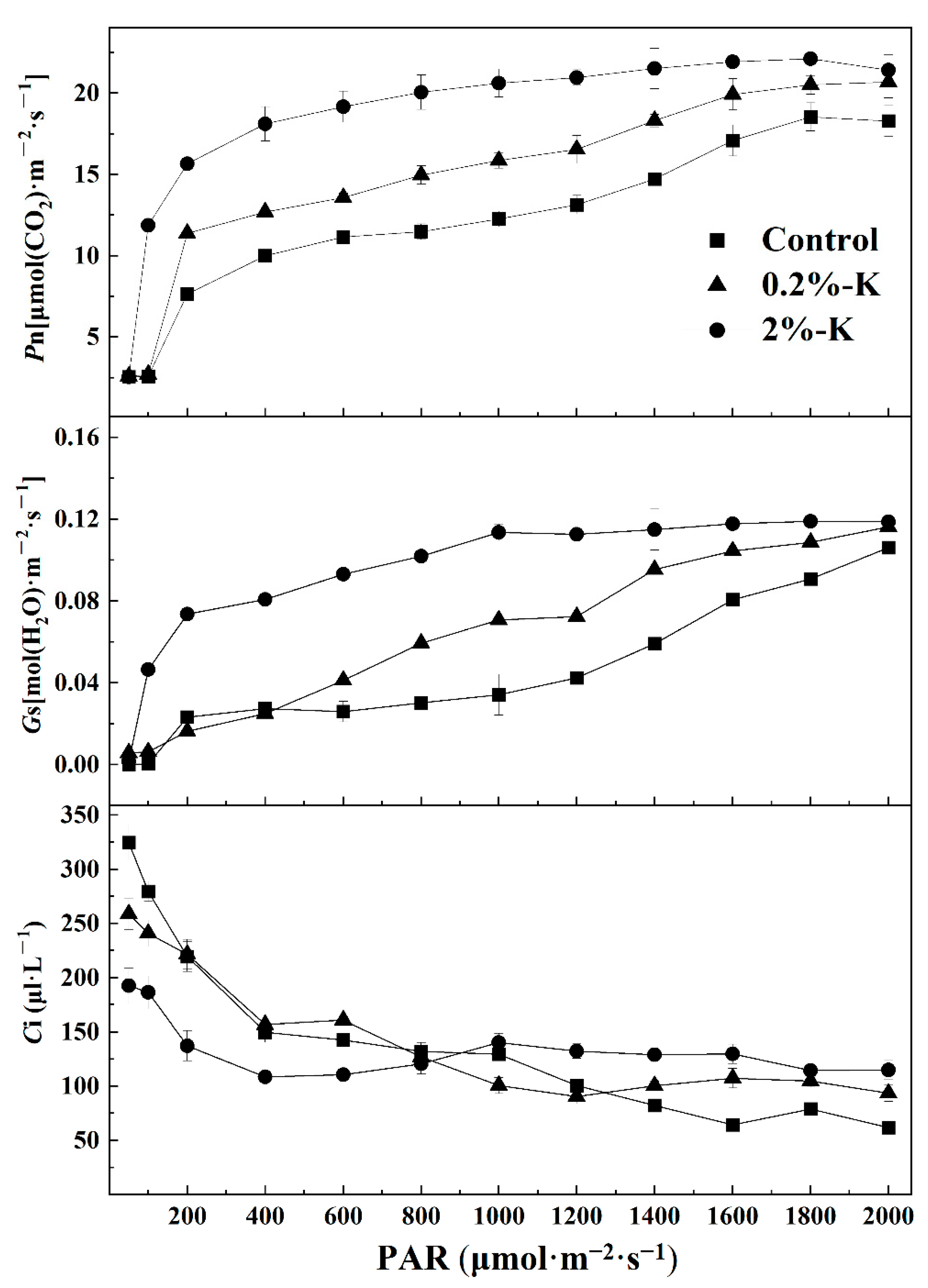
2.2. Effect of Potassium on the Contents of Nutrient Elements and Photosynthesis in Citrus Fruits and Leaves
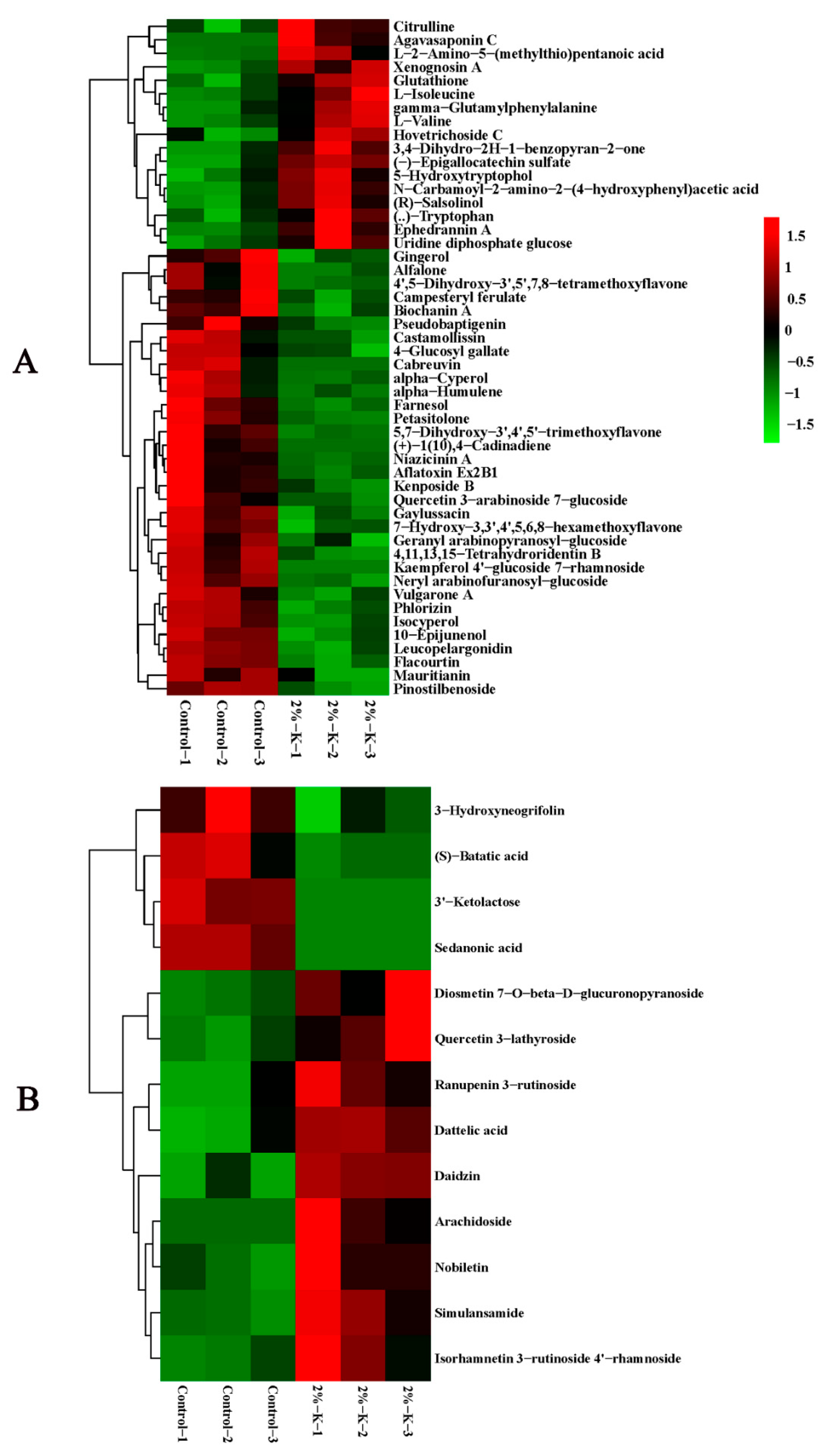
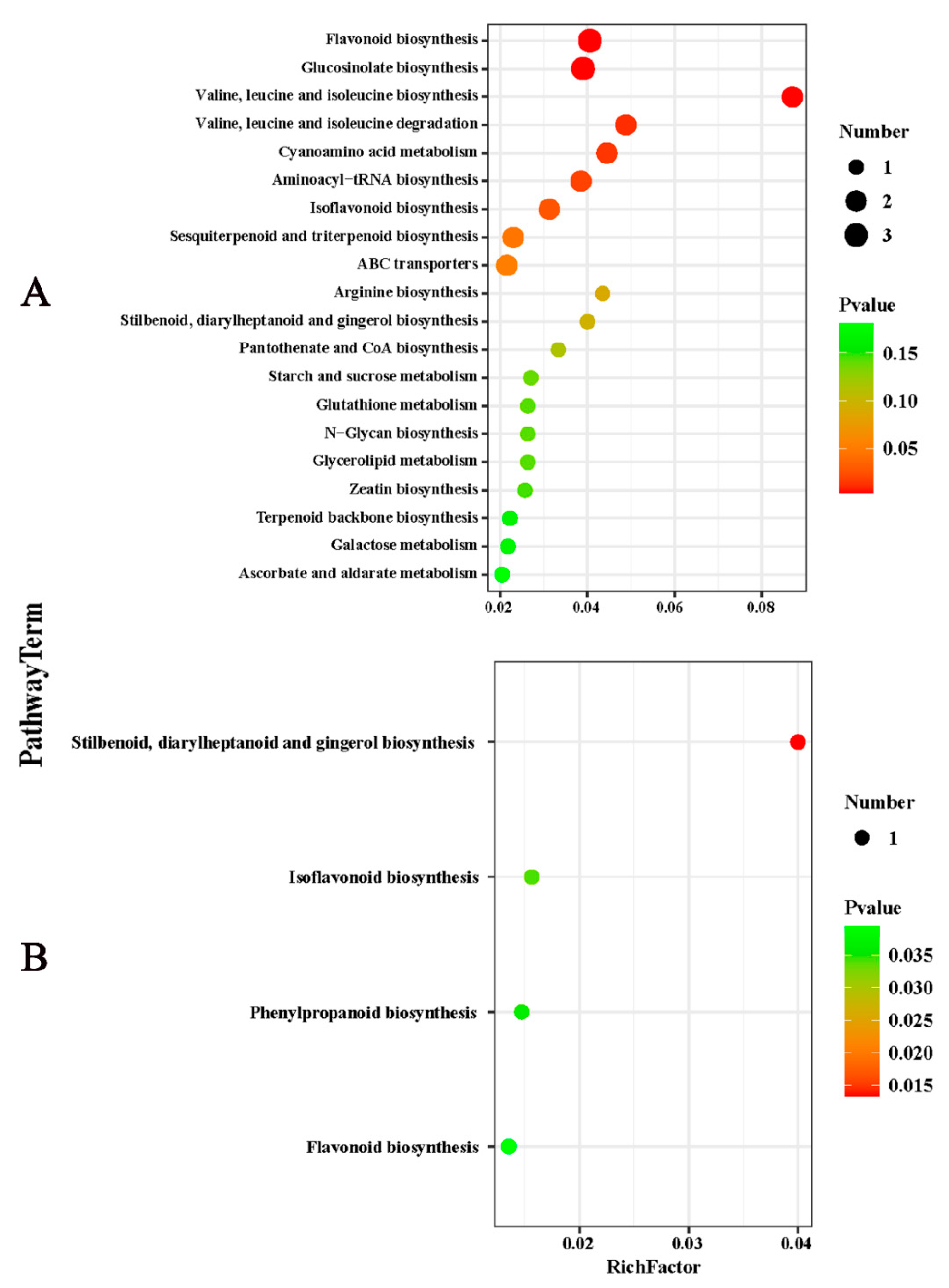
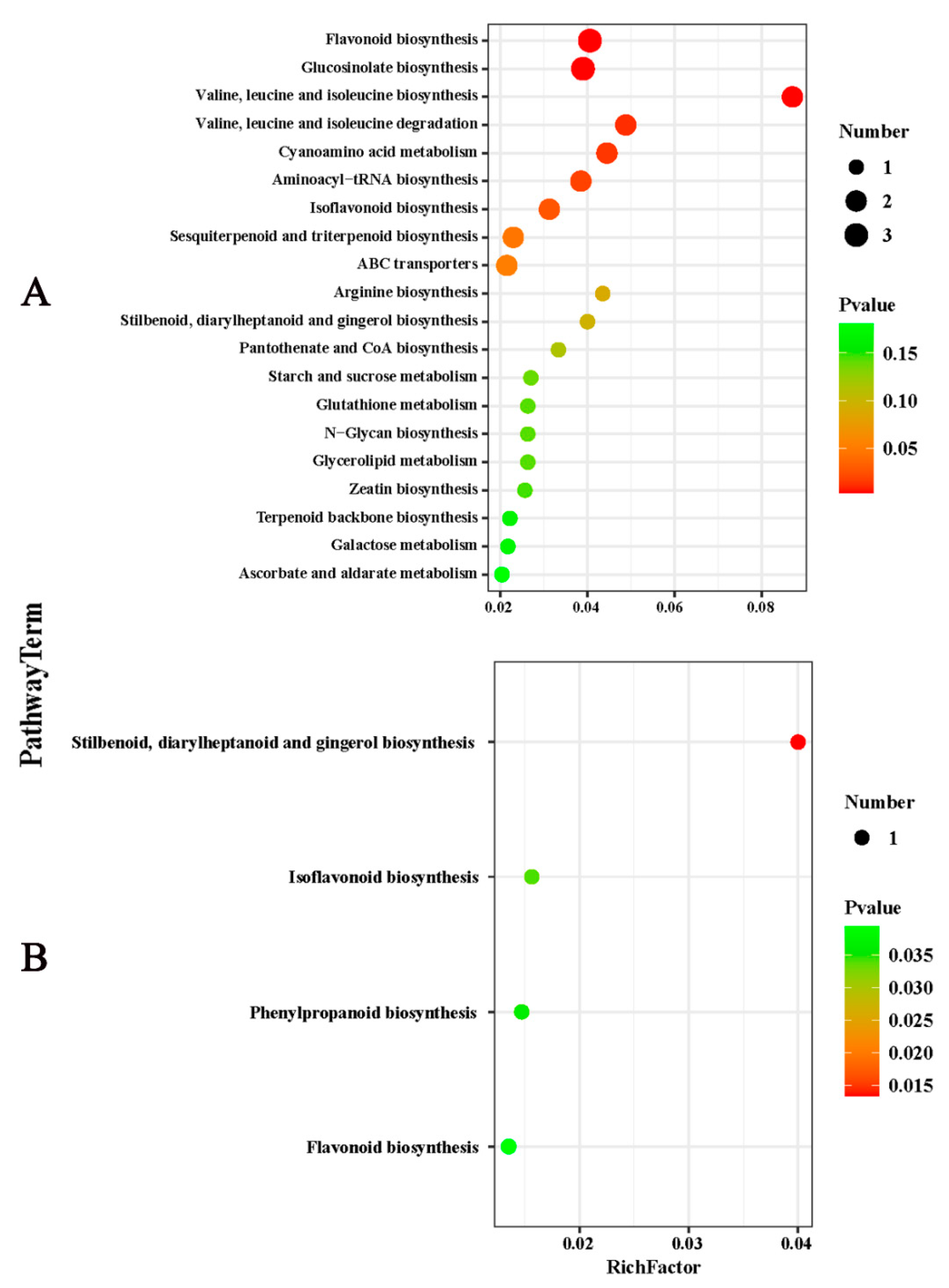
2.3. Effects of Potassium on the Components of Metabolites in Citrus Fruits
2.4. Overview of the Core DEMs in Citrus Fruit following Potassium Treatment
3. Discussion
4. Materials and Methods
4.1. Plant Treatment and Sampling
4.2. Measurements of Physiological Characteristics
4.3. LC/MS Untargeted Metabolomics Analysis
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hyun, J.W.; Hwang, R.Y.; Choi, C.W.; Jung, K.E.; Han, S.G. Symptomatology of Citrus mosaic sadwavirus (CiMV) in Some Citrus Cultivars and Effect of CiMV Infection on Citrus Fruit Quality. Plant Pathol. J. 2020, 3, 106. [Google Scholar] [CrossRef] [PubMed]
- Imai, A.; Kuniga, T. Genome-wide estimation of pedigree haplotypes offers genetic compositions and founder origins in Japanese citrus breeding materials. Sci. Hortic. 2021, 282, 110000. [Google Scholar] [CrossRef]
- Cronje, P.J.; Stander, O.P.; Theron, K.I. Fruit splitting in citrus. Hortic. Rev. 2013, 41, 177–200. [Google Scholar]
- Fan, Z.; Xiong, H.; Luo, Y.; Wang, Y.; Zhao, H.; Li, W.; He, X.; Wang, J.; Shi, X.; Zhang, Y. Fruit Yields Depend on Biomass and Nutrient Accumulations in New Shoots of Citrus Trees. Agronomy 2020, 10, 1988. [Google Scholar] [CrossRef]
- Wu, S.; Zhang, C.; Li, M.; Tan, Q.; Sun, X.; Pan, Z.; Deng, X.; Hu, C. Effects of potassium on fruit soluble sugar and citrate accumulations in Cara Cara navel orange (Citrus sinensis L. Osbeck). Sci. Hortic. 2021, 283, 110057. [Google Scholar] [CrossRef]
- Mangali Mounika, D.T.; Kumar, A.K.; Joshi, V.; Sunil, N. Studies on the effect of foliar application of calcium, potassium and silicon on quality and shelf life of sweet orange (Citrus sinensis L.) cv. Sathgudi. J. Pharmacogn. Phytochem. 2021, 10, 1711–1713. [Google Scholar]
- Kadyampakeni, D.M.; Chinyukwi, T. Are macronutrients and micronutrients therapeutic for restoring performance of trees affected by citrus greening? A discussion of current practices and future research opportunities. J. Plant Nutr. 2021, 44, 2949–2969. [Google Scholar] [CrossRef]
- Abbas, F.; Fares, A. Best management practices in citrus production. Tree For. Sci. Biotechnol. 2009, 3, 1–11. [Google Scholar]
- Savreet, S.; Bal, J. Quality improvement in lemon (Citrus limon (L.) Burm.) through integrated management of fruit cracking. Afr. J. Agric. Res. 2013, 8, 3552–3557. [Google Scholar] [CrossRef] [Green Version]
- Amin, A.; Al-Abbasi, G.; Alkurdi, H. Effect of foliar spray with nano-optimus plus and potassium chelated with amino acids in some growth characters of Citrus aurantifolia L. Saplings. Plant Arch. 2020, 20, 897–900. [Google Scholar]
- Zhang, J.; Ding, J.; Ibrahim, M.; Jiao, X.; Song, X.; Bai, P.; Li, J. Effects of the interaction between vapor-pressure deficit and potassium on the photosynthesis system of tomato seedlings under low temperature. Sci. Hortic. 2021, 283, 110089. [Google Scholar] [CrossRef]
- Gao, J.; Su, Y.; Yu, M.; Huang, Y.; Wang, F.; Shen, A. Potassium alleviates post-anthesis photosynthetic reductions in winter wheat caused by waterlogging at the stem elongation stage. Front. Plant Sci. 2021, 11, 1982. [Google Scholar] [CrossRef] [PubMed]
- Sarkar, S.; Khatun, M.; Era, F.M.; Islam, A.; Anwar, M.; Danish, S.; Datta, R.; Islam, A. Abiotic stresses: Alteration of composition and grain quality in food legumes. Agronomy 2021, 11, 2238. [Google Scholar] [CrossRef]
- Gao, C.; El-Sawah, A.M.; Ali, D.F.I.; Alhaj Hamoud, Y.; Shaghaleh, H.; Sheteiwy, M.S. The integration of bio and organic fertilizers improve plant growth, grain yield, quality and metabolism of hybrid maize (Zea mays L.). Agronomy 2020, 10, 319. [Google Scholar] [CrossRef] [Green Version]
- Xu, H.-j.; Johkan, M.; Maruo, T.; Kagawa, N.; Tsukagoshi, S. New Insight on Low-K Lettuce: From Photosynthesis to Primary and Secondary Metabolites. HortScience 2021, 56, 407–413. [Google Scholar] [CrossRef]
- Ibrahim, M.H.; Jaafar, H.Z.; Karimi, E.; Ghasemzadeh, A. Primary, secondary metabolites, photosynthetic capacity and antioxidant activity of the Malaysian Herb Kacip Fatimah (Labisia pumila Benth) exposed to potassium fertilization under greenhouse conditions. Int. J. Mol. Sci. 2012, 13, 15321–15342. [Google Scholar] [CrossRef] [PubMed]
- Troufflard, S.; Mullen, W.; Larson, T.R.; Graham, I.A.; Crozier, A.; Amtmann, A.; Armengaud, P. Potassium deficiency induces the biosynthesis of oxylipins and glucosinolates in Arabidopsis thaliana. BMC Plant Biol. 2010, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- El-Sayed, S.A. Some factors affecting orange fruit splitting of washington navel orange under kafr elsheikh conditions. a-the effect of rootstock. J. Plant Product. 2016, 7, 343–349. [Google Scholar] [CrossRef]
- Li, Y.; Jin, G.; Jiang, X.; Yi, S.; Tian, X. Non-destructive determination of soluble solids content using a multi-region combination model in hybrid citrus. Infrared Phys. Technol. 2020, 104, 103138. [Google Scholar] [CrossRef]
- Alikhani, M.; Babakhani, B.; Golein, B.; Asadi, M.; Rahdari, P. Foliar application of potassium nitrate and 2,4-dichlorophenoxyacetic acid affect some fruit splitting related characteristics and biochemical traits of mandarin cv.‘page’. Eur. Asian J. BioSci. 2020, 14, 4251–4260. [Google Scholar]
- Ali, M.M.; Yousef, A.F.; Li, B.; Chen, F. Effect of Environmental Factors on Growth and Development of Fruits. Trop. Plant Biol. 2021, 14, 226–238. [Google Scholar] [CrossRef]
- Chang, B.-M.; Keller, M. Cuticle and skin cell walls have common and unique roles in grape berry splitting. Hortic. Res. 2021, 8, 168. [Google Scholar] [CrossRef] [PubMed]
- Cui, J.; Lamade, E.; Tcherkez, G. Potassium deficiency reconfigures sugar export and induces catecholamine accumulation in oil palm leaves. Plant Sci. 2020, 300, 110628. [Google Scholar] [CrossRef]
- Muneta-Arrate, I.; Diez-Alarcia, R.; Horrillo, I.; Meana, J.J. Pimavanserin exhibits serotonin 5-HT2A receptor inverse agonism for Gαi1-and neutral antagonism for Gαq/11-proteins in human brain cortex. Eur. Neuropsychopharm. 2020, 36, 83–89. [Google Scholar] [CrossRef] [PubMed]
- Sprangers, K.; Thys, S.; van Dusschoten, D.; Beemster, G.T. Gibberellin enhances the anisotropy of cell expansion in the growth zone of the maize leaf. Front. Plant Sci. 2020, 11, 1163. [Google Scholar] [CrossRef] [PubMed]
- Barker, R.; Fernandez Garcia, M.N.; Powers, S.J.; Vaughan, S.; Bennett, M.J.; Phillips, A.L.; Thomas, S.G.; Hedden, P. Mapping sites of gibberellin biosynthesis in the Arabidopsis root tip. New Phytol. 2021, 229, 1521–1534. [Google Scholar] [CrossRef] [PubMed]
- Wang, Y.; Guo, L.; Zhao, X.; Zhao, Y.; Hao, Z.; Luo, H.; Yuan, Z. Advances in Mechanisms and Omics Pertaining to Fruit Cracking in Horticultural Plants. Agronomy 2021, 11, 1045. [Google Scholar] [CrossRef]
- Faizy, A.H.; Ozturk, B.; Aglar, E.; Yıldız, K. Role of methyl jasmonate application regime on fruit quality and bioactive compounds of sweet cherry at harvest and during cold storage. J. Food Process. Pres. 2021, 45, e15882. [Google Scholar] [CrossRef]
- Tang, H.; Zhang, X.; Gong, B.; Yan, Y.; Shi, Q. Proteomics and metabolomics analysis of tomato fruit at different maturity stages and under salt treatment. Food Chem. 2020, 311, 126009. [Google Scholar] [CrossRef]
- Liu, A.; Xiao, Z.; Li, M.W.; Wong, F.L.; Yung, W.S.; Ku, Y.S.; Wang, Q.; Wang, X.; Xie, M.; Yim, A.K.Y. Transcriptomic reprogramming in soybean seedlings under salt stress. Plant Cell Environ. 2019, 42, 98–114. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Liu, S.; Wang, H.; Chen, K.; Zhang, P. The flavonoid 3′-hydroxylase gene from the Antarctic moss Pohlia nutans is involved in regulating oxidative and salt stress tolerance. Biotechnol. Appl. Biochem. 2021, bab2143, 1–11. [Google Scholar] [CrossRef]
- Mokhtarzadeh, Z.; Shahsavar, A.R. Effects of gibberellic acid, potassium nitrate and calcium sulfate on pomegranate fruit splitting and fruit characteristics. Agric. Conspec. Sci. 2020, 85, 237–245. [Google Scholar]
- Ghanbarpour, E.; Rezaei, M.; Lawson, S. Reduction of cracking in pomegranate fruit after foliar application of humic acid, calcium-boron and kaolin during water stress. Erwerbs-Obstbau 2019, 61, 29–37. [Google Scholar] [CrossRef]
- Singh, A.; Shukla, A.; Meghwal, P. Fruit cracking in pomegranate: Extent, cause, and management. Int. J. Fruit Sci. 2020, 20, S1234–S1253. [Google Scholar] [CrossRef]
- Xie, R.-j.; Zheng, L.; Jiao, Y.; Huang, X. Understanding physiological and molecular mechanisms of citrus rootstock seedlings in response to root zone hypoxia by RNA-Seq. Environ. Exp. Bot. 2021, 192, 104647. [Google Scholar] [CrossRef]
- Dong, T.; Shang, J.; Chen, J.M.; Liu, J.; Qian, B.; Ma, B.; Morrison, M.-J.; Zhang, C.; Liu, Y.-P.; Shi, Y.-C.; et al. Assessment of portable chlorophyll meters for measuring crop leaf chlorophyll concentration. Remote Sens. 2019, 11, 2706. [Google Scholar] [CrossRef] [Green Version]
- León García, I.V. Warming Effect on Thermal Tolerance and Photosynthetic Performance of Paramo Plants; Uniandes-Universidad de los Andes: Bogota, Colombia, 2017. [Google Scholar]
- Hervalejo, A.; Cardenosa, V.; Forner-Giner, M.A.; Salguero, A.; Pradas, I.C.; Moreno, J.M.; Merino, C.; Arenas-Arenas, F.J. Preliminary Data on Influence of Six Citrus Rootstocks on Fruit Quality of ‘Lane Late’Navel Orange. Acta Hort. 2015, 1065, 363–366. [Google Scholar] [CrossRef]
- Zhu, S.; Nong, J.; Luo, G.; Li, Q.; Wang, F.; Jiang, D.; Zhao, X. Varied tolerance and different responses of five citrus rootstocks to acid stress by principle component analysis and orthogonal analysis. Sci. Hortic. 2021, 278, 109853. [Google Scholar] [CrossRef]
- Jones, J. Kjeldahl Method for Nitrogen (N) Determination; Micromacro Publishing: Athens, GA, USA, 1991; p. 67. [Google Scholar]
- Qi, N.L.; Li, P.W.; Zeng, X.H.; Huang, H.H.; Yang, Z.M.; Gong, X. Comparison of Kjeldahl and the elemental analysis methods for determination of nitrogen content in raw natural rubber. In Advanced Materials Research; Trans Tech Publication: Zurich, Switzerland, 2013. [Google Scholar]
- Ai, J.; Wu, Q.; Battino, M.; Bai, W.; Tian, L. Using untargeted metabolomics to profile the changes in roselle (Hibiscus sabdariffa L.) anthocyanins during wine fermentation. Food Chem. 2021, 364, 130425. [Google Scholar] [CrossRef]
- Cao, Y.-W.; Qu, R.-J.; Tang, X.-Q.; Sun, L.-Q.; Chen, Q.-Q.; Miao, Y.-J. UPLC-Triple TOF-MS/MS based metabolomics approach to reveal the influence of nitrogen levels on Isatis indigotica seedling leaf. Sci. Hortic. 2020, 266, 109280. [Google Scholar] [CrossRef]
- Santos, A.S.; de Andrade, E.M.; de Oliveira Sousa, A.R.; do Carmo Costa, M.M.; Togawa, R.; Grynberg, P.; Pirovani, C.P.; Oliveira, S.A.S.; dos Santos Soares Filho, W.; Coelho Filho, M.A. Transcriptome profile of drought responsive candidate genes in varieties of citrus rootstocks with different tolerance strategies. Sci. Hortic. 2021, 277, 109838. [Google Scholar] [CrossRef]

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Treatment | Total Fruit Split (%) | Vertical (%) | Horizontal (%) | Hardness Ratio |
|---|---|---|---|---|
| Control | 31.0 a | 12.0 a | 19.0 a | 1.34 c |
| 0.2%-K | 15.0 b | 5.00 b | 10.0 b | 1.97 b |
| 2%-K | 5.00 c | 2.00 c | 3.00 c | 3.53 a |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jiao, Y.; Sha, C.; Shu, Q. Integrated Physiological and Metabolomic Analyses of the Effect of Potassium Fertilizer on Citrus Fruit Splitting. Plants 2022, 11, 499. https://doi.org/10.3390/plants11040499
Jiao Y, Sha C, Shu Q. Integrated Physiological and Metabolomic Analyses of the Effect of Potassium Fertilizer on Citrus Fruit Splitting. Plants. 2022; 11(4):499. https://doi.org/10.3390/plants11040499
Chicago/Turabian StyleJiao, Yun, Cunlong Sha, and Qiaoyun Shu. 2022. "Integrated Physiological and Metabolomic Analyses of the Effect of Potassium Fertilizer on Citrus Fruit Splitting" Plants 11, no. 4: 499. https://doi.org/10.3390/plants11040499
APA StyleJiao, Y., Sha, C., & Shu, Q. (2022). Integrated Physiological and Metabolomic Analyses of the Effect of Potassium Fertilizer on Citrus Fruit Splitting. Plants, 11(4), 499. https://doi.org/10.3390/plants11040499