
Three Chemically Distinct Floral Ecotypes in Drakaea livida, an Orchid Pollinated by Sexual Deception of Thynnine Wasps


Abstract
:1. Introduction
2. Results
2.1. Determining the Pollinator Species of D. livida Populations
2.2. Pollinator Choice Experiments
2.2.1. Response of Zaspilothynnus dilatatus to Flowers from Populations That Attract Z. nigripes and Catocheilus sp.
2.2.2. Response of Zaspilothynnus nigripes to Flowers from Populations That Attract Zaspilothynnus dilatatus
2.2.3. Response of Zaspilothynnus nigripes to Flowers from Populations That Attract Catocheilus sp.
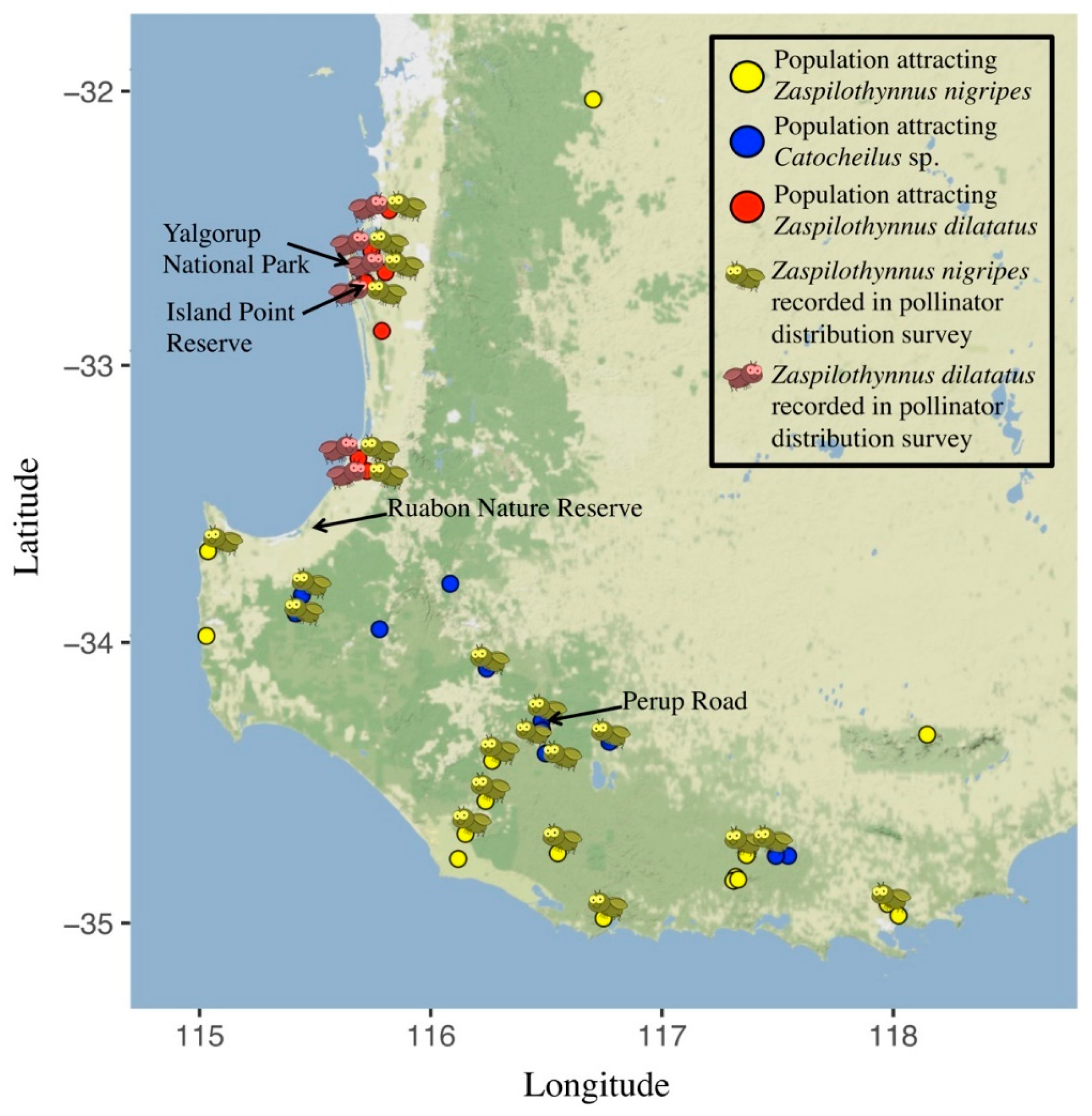
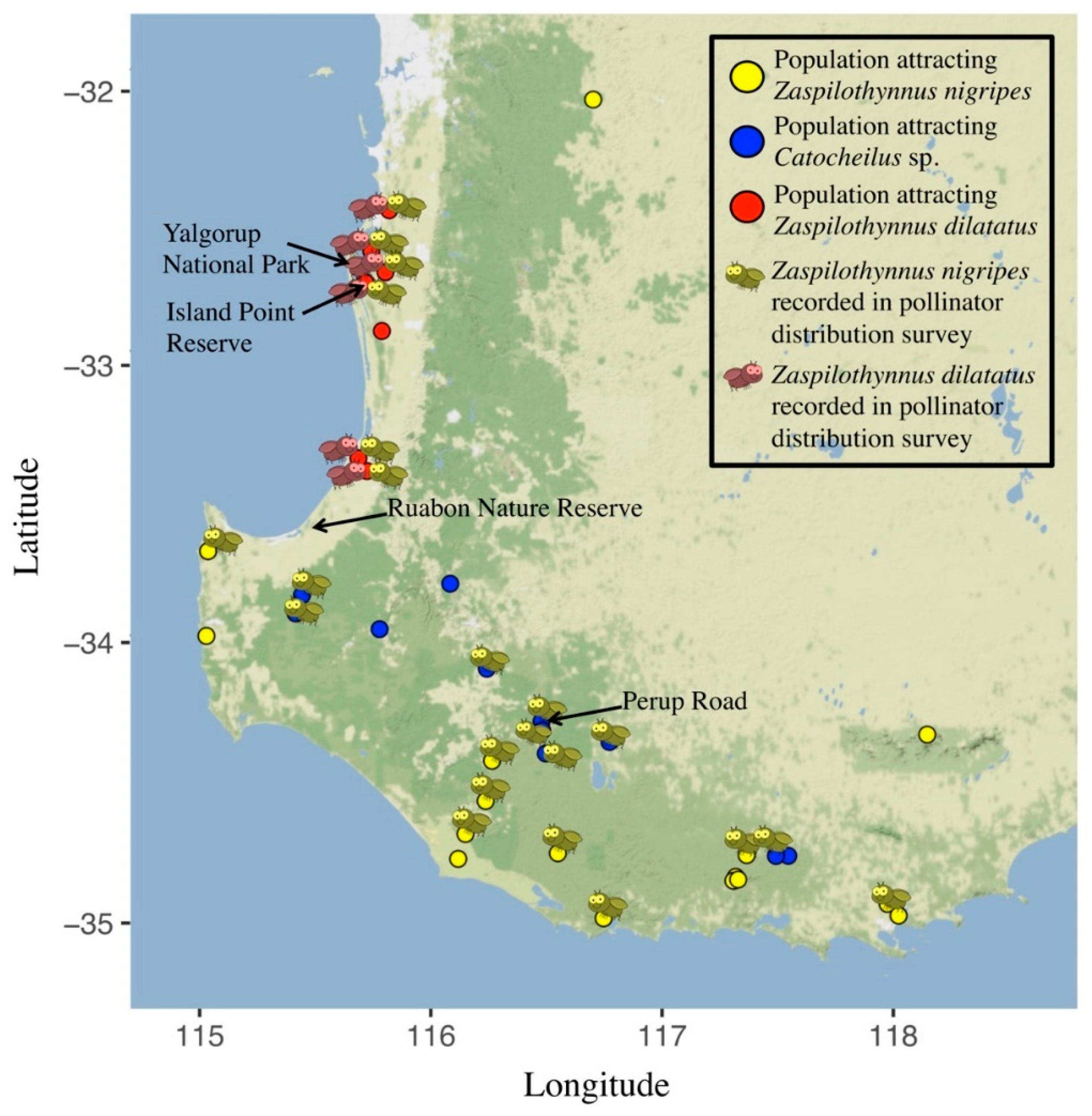
2.3. Correlation of Plant Distribution and Pollinator Availability
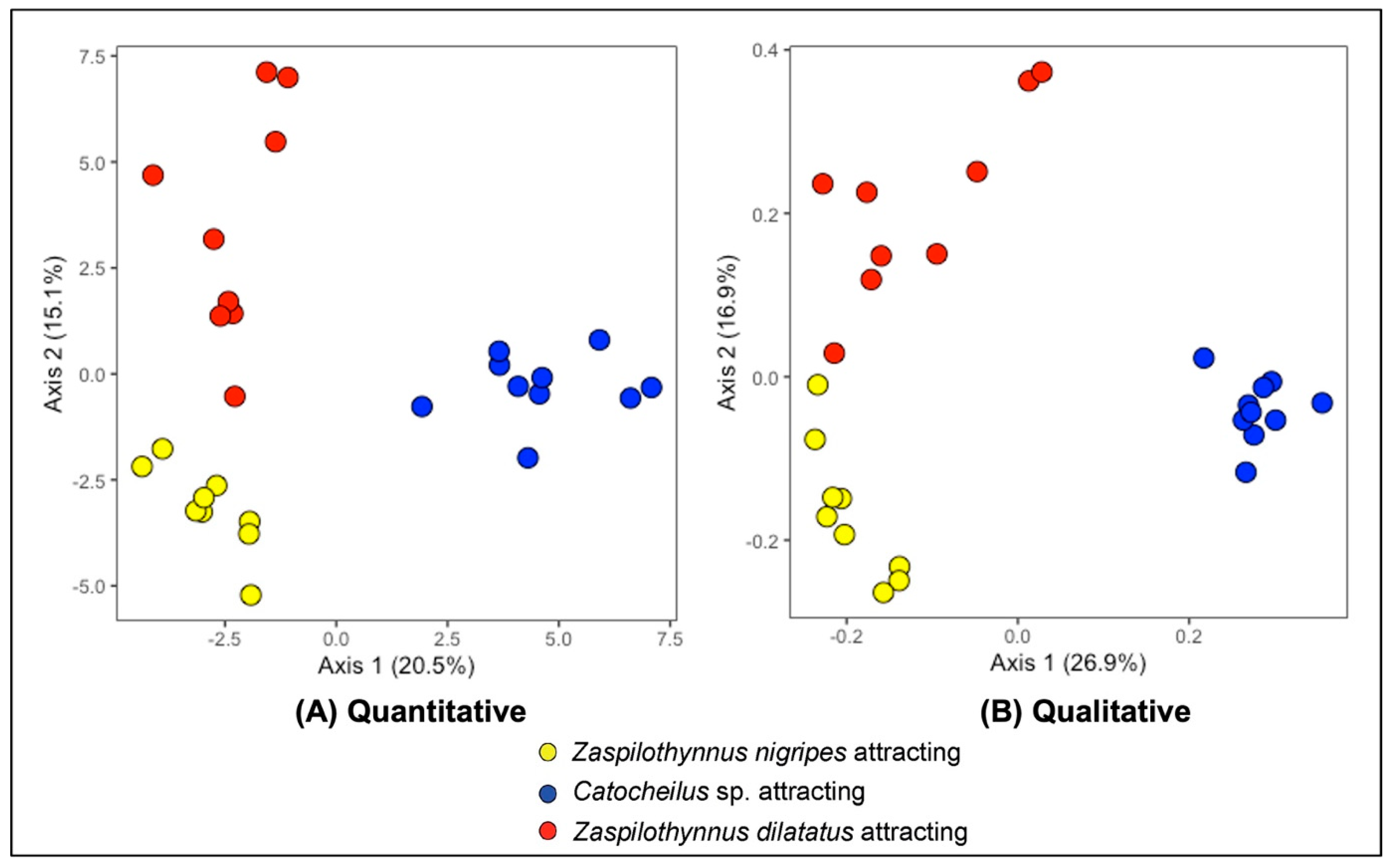
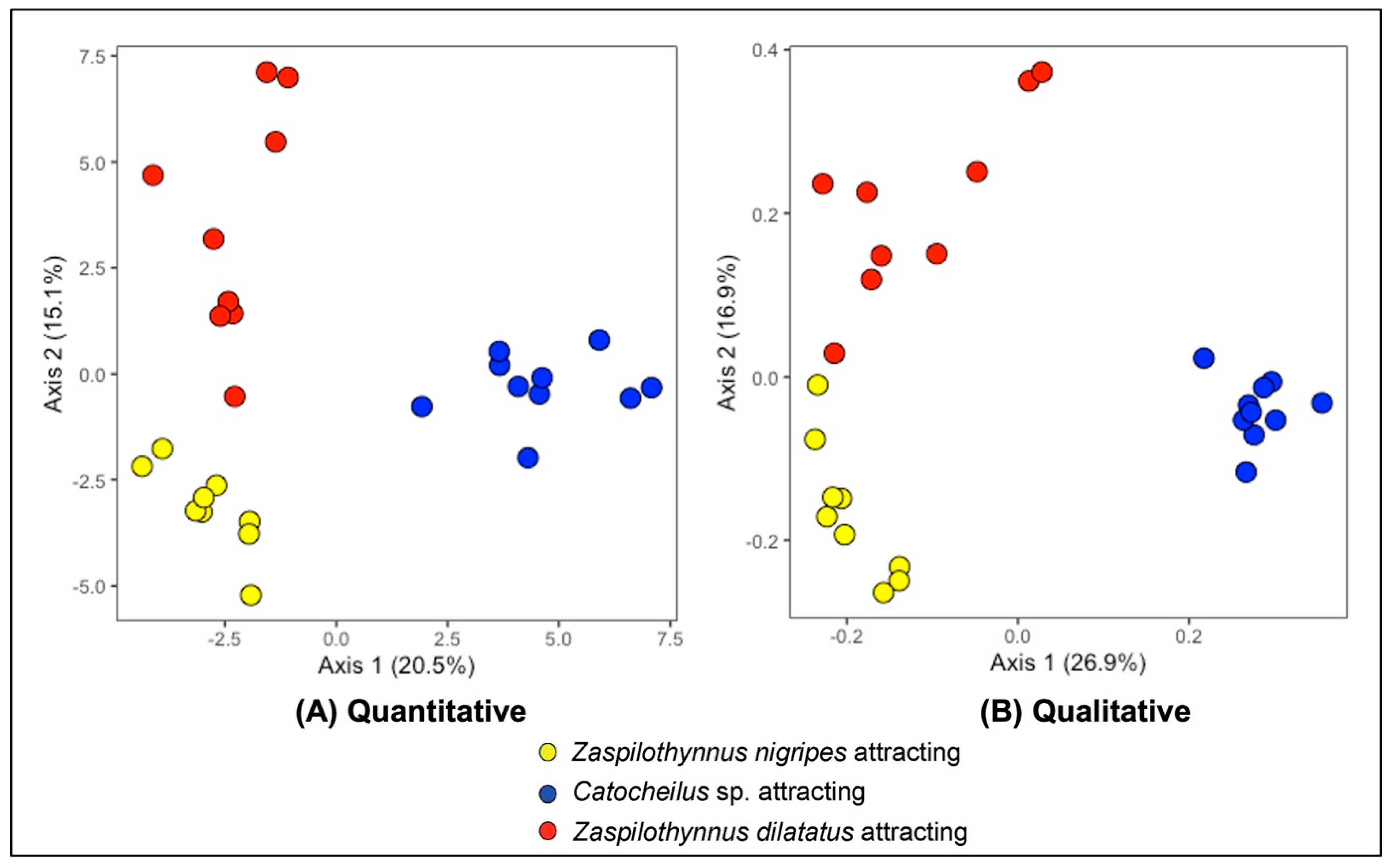
2.4. Floral Volatile Composition of Plants That Attract Different Pollinator Species
2.5. Presence of Electrophysiologically Active Compounds
2.5.1. Gas Chromatography/Mass Spectrometry-Electroantennographic Detection Studies of Z. dilatatus
2.5.2. Screening of Floral Extracts for Electrophysiologically Active Compounds
2.6. Compounds That Differ between Ecotypes
3. Discussion
3.1. Presence of Floral Ecotypes in Drakaea livida
3.2. Correlation of Ecotype Distribution and Pollinator Availability
3.3. Floral Volatile Composition of the Ecotypes
3.4. The Discovery of (Methylthio)Phenols in D. livida
3.5. Conservation Implications of the Presence of Ecotypes
4. Materials and Methods
4.1. Study Species
4.2. Testing for the Presence of Floral Ecotypes in D. livida
4.3. Determining the Pollinator Species of D. livida Populations
4.4. Pollinator Choice Experiments
4.4.1. Response of Zaspilothynnus dilatatus to Flowers from Populations That Attract Other Pollinator Species
4.4.2. Response of Zaspilothynnus nigripes to Flowers from Populations That Attract Zaspilothynnus dilatatus
4.4.3. Response of Zaspilothynnus nigripes to Flowers from Populations That Attract Catocheilus sp.
4.5. Correlation of Plant Geographic Range and Pollinator Availability
4.6. Floral Volatile Composition of Plants That Attract Different Pollinator Species
4.7. Presence of Electrophysiologically Active Compounds
4.7.1. Gas Chromatography/Mass Spectrometry-Electroantennographic Detection Studies of Z. dilatatus
4.7.2. Screening of Floral Extracts for Electrophysiologically Active Compounds
4.8. Compounds That Differ between Ecotypes
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Ayasse, M.; Stökl, J.; Francke, W. Chemical ecology and pollinator-driven speciation in sexually deceptive orchids. Phytochemistry 2011, 72, 1667–1677. [Google Scholar] [CrossRef]
- Xu, S.; Schlüter, P.M.; Schiestl, F.P. Pollinator-driven speciation in sexually deceptive orchids. Int. J. Ecol. 2012, 2012, 285081. [Google Scholar] [CrossRef] [Green Version]
- Coleman, E. Pollination of an australian orchid by the male ichneumonid Lissopimpla semipunctata, Kirby. Trans. Entomol. Soc. Lond. 1928, 76, 533–539. [Google Scholar] [CrossRef]
- Kullenberg, B. Studies in Ophrys pollination. Zool. Bidr. Frân Upps. 1961, 34, 1–340. [Google Scholar]
- Stoutamire, W.P. Australian terrestrial orchids, thynnid wasps, and pseudocopulation. Am. Orchid. Soc. Bull. 1974, 43, 13–18. [Google Scholar]
- Schiestl, F.P.; Peakall, R.; Mant, J.G.; Ibarra, F.; Schulz, C.; Franke, S.; Francke, W. The chemistry of sexual deception in an orchid-wasp pollination system. Science 2003, 302, 437–438. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, F.P.; Ayasse, M.; Paulus, H.F.; Lofstedt, C.; Hansson, B.S.; Ibarra, F.; Francke, W. Orchid pollination by sexual swindle. Nature 1999, 399, 421. [Google Scholar] [CrossRef]
- Ellis, A.G.; Johnson, S.D. Floral mimicry enhances pollen export: The evolution of pollination by sexual deceit outside of the Orchidaceae. Am. Nat. 2010, 176, E143–E151. [Google Scholar] [CrossRef] [PubMed]
- Vereecken, N.J.; Wilson, C.A.; Hötling, S.; Schulz, S.; Banketov, S.A.; Mardulyn, P. Pre-adaptations and the evolution of pollination by sexual deception: Cope’s rule of specialization revisited. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2012, 279, 4786–4794. [Google Scholar] [CrossRef] [Green Version]
- Bohman, B.; Flematti, G.R.; Barrow, R.A.; Pichersky, E.; Peakall, R. Pollination by sexual deception—It takes chemistry to work. Curr. Opin. Plant Biol. 2016, 32, 37–46. [Google Scholar] [CrossRef]
- Cohen, C.; Liltved, W.R.; Colville, J.F.; Shuttleworth, A.; Weissflog, J.; Svatoš, A.; Bytebier, B.; Johnson, S.D. Sexual deception of a beetle pollinator through floral mimicry. Curr. Biol. 2021, 31, 1962–1969.e6. [Google Scholar] [CrossRef]
- Paulus, H.F.; Gack, C. Pollination of Ophrys (Orchidaceae) in Cyprus. Plant Syst. Evol. 1990, 169, 177–207. [Google Scholar] [CrossRef]
- Bower, C.C.; Brown, G.R. Pollinator specificity, cryptic species and geographical patterns in pollinator responses to sexually deceptive orchids in the genus Chiloglottis: The Chiloglottis gunnii complex. Aust. J. Bot. 2009, 57, 37–55. [Google Scholar] [CrossRef]
- Peakall, R.; Ebert, D.; Poldy, J.; Barrow, R.A.; Francke, W.; Bower, C.C.; Schiestl, F.P. Pollinator specificity, floral odour chemistry and the phylogeny of Australian sexually deceptive Chiloglottis orchids: Implications for pollinator-driven speciation. New Phytol. 2010, 188, 437–450. [Google Scholar] [CrossRef]
- Phillips, R.; Brown, G.R.; Dixon, K.W.; Hayes, C.; Linde, C.; Peakall, R. Evolutionary relationships among pollinators and repeated pollinator sharing in sexually deceptive orchids. J. Evol. Biol. 2017, 30, 1674–1691. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gaskett, A.C. Orchid pollination by sexual deception: Pollinator perspectives. Biol. Rev. 2011, 86, 33–75. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, F.P.; Ayasse, M.; Paulus, H.F.; Löfstedt, C.; Hansson, B.S.; Ibarra, F.; Francke, W. Sex pheromone mimicry in the early spider orchid (Ophrys sphegodes): Patterns of hydrocarbons as the key mechanism for pollination by sexual deception. J. Comp. Physiol. A 2000, 186, 567–574. [Google Scholar] [CrossRef] [PubMed]
- Mant, J.; Brändli, C.; Vereecken, N.J.; Schulz, C.M.; Francke, W.; Schiestl, F.P. Cuticular hydrocarbons as sex pheromone of the bee Colletes cunicularius and the key to its mimicry by the sexually deceptive orchid, Ophrys exaltata. J. Chem. Ecol. 2005, 31, 1765–1787. [Google Scholar] [CrossRef] [PubMed]
- Gervasi, D.D.; Selosse, M.A.; Sauve, M.; Francke, W.; Vereecken, N.J.; Cozzolino, S.; Schiestl, F.P. Floral scent and species divergence in a pair of sexually deceptive orchids. Ecol. Evol. 2017, 7, 6023–6034. [Google Scholar] [CrossRef] [Green Version]
- Ayasse, M.; Schiestl, F.P.; Paulus, H.F.; Ibarra, F.; Francke, W. Pollinator attraction in a sexually deceptive orchid by means of unconventional chemicals. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2003, 270, 517–522. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohman, B.; Phillips, R.D.; Flematti, G.R.; Peakall, R. (Methylthio) phenol semiochemicals are exploited by deceptive orchids as sexual attractants for thynnine wasps. Fitoterapia 2017, 126, 78–82. [Google Scholar] [CrossRef] [Green Version]
- Bohman, B.; Phillips, R.D.; Flematti, G.R.; Barrow, R.A.; Peakall, R. The spider orchid Caladenia crebra produces sulfurous pheromone mimics to attract its male wasp pollinator. Angew. Chem. Int. Ed. 2017, 56, 8455–8458. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, H.; Bohman, B.; Wong, D.C.; Rodriguez-Delgado, C.; Scaffidi, A.; Flematti, G.R.; Phillips, R.D.; Pichersky, E.; Peakall, R. Complex sexual deception in an orchid is achieved by co-opting two independent biosynthetic pathways for pollinator attraction. Curr. Biol. 2017, 27, 1867–1877. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Xu, S.; Schlüter, P.M.; Scopece, G.; Breitkopf, H.; Gross, K.; Cozzolino, S.; Schiestl, F.P. Floral isolation is the main reproductive barrier among closely related sexually deceptive orchids. Evolution 2011, 65, 2606–2620. [Google Scholar] [CrossRef]
- Peakall, R.; Whitehead, M. Floral odour chemistry defines species boundaries and underpins strong reproductive isolation in sexually deceptive orchids. Ann. Bot. 2014, 113, 341–355. [Google Scholar] [CrossRef] [Green Version]
- de Jager, M.L.; Peakall, R. Does morphology matter? An explicit assessment of floral morphology in sexual deception. Funct. Ecol. 2016, 30, 537–546. [Google Scholar] [CrossRef]
- Bower, C.C. Specific pollinators reveal a cryptic taxon in the bird orchid, Chiloglottis valida sensu lato (Orchidaceae) in south-eastern Australia. Aust. J. Bot. 2006, 54, 53–64. [Google Scholar] [CrossRef]
- Menz, M.H.; Phillips, R.D.; Anthony, J.M.; Bohman, B.; Dixon, K.W.; Peakall, R. Ecological and genetic evidence for cryptic ecotypes in a rare sexually deceptive orchid, Drakaea elastica. Bot. J. Linn. Soc. 2015, 177, 124–140. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.; Bohman, B.; Anthony, J.; Krauss, S.; Dixon, K.; Peakall, R. Mismatch in the distribution of floral ecotypes and pollinators: Insights into the evolution of sexually deceptive orchids. J. Evol. Biol. 2015, 28, 601–612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Breitkopf, H.; Onstein, R.E.; Cafasso, D.; Schlüter, P.M.; Cozzolino, S. Multiple shifts to different pollinators fuelled rapid diversification in sexually deceptive Ophrys orchids. New Phytol. 2015, 207, 377–389. [Google Scholar] [CrossRef] [PubMed]
- Van der Niet, T.; Peakall, R.; Johnson, S.D. Pollinator-driven ecological speciation in plants: New evidence and future perspectives. Ann. Bot. 2014, 113, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Phillips, R.D.; Reiter, N.; Peakall, R. Orchid conservation: From theory to practice. Ann. Bot. 2020, 126, 345–362. [Google Scholar] [CrossRef]
- Velasco, L.; Goffman, F.D. Chemotaxonomic significance of fatty acids and tocopherols in Boraginaceae. Phytochemistry 1999, 52, 423–426. [Google Scholar] [CrossRef]
- Goffman, F.D.; Thies, W.; Velasco, L. Chemotaxonomic value of tocopherols in Brassicaceae. Phytochemistry 1999, 50, 793–798. [Google Scholar] [CrossRef]
- Özcan, T. Fatty Acid Profiles of the Seed Oils in Two Groups of Anchusa officinalis L. IUFS J. Biol. 2008, 67, 65–71. [Google Scholar]
- Coutinho, D.J.G.; Barbosa, M.O.; Silva, R.M.; da Silva, S.I.; de Oliveira, A.F.M. Fatty-Acid Composition of Seeds and Chemotaxonomic Evaluation of Sixteen Sapindaceae Species. Chem. Biodivers. 2015, 12, 1271–1280. [Google Scholar] [CrossRef] [PubMed]
- Velasco, L.; Goffman, F.D.; Pujadas-Salvà, A.J. Fatty acids and tocochromanols in seeds of Orobanche. Phytochemistry 2000, 54, 295–300. [Google Scholar] [CrossRef]
- Li, H.; Madden, J.; Potts, B. Variation in volatile leaf oils of the Tasmanian Eucalyptus species—1. Subgenus Monocalyptus. Biochem. Syst. Ecol. 1995, 23, 299–318. [Google Scholar] [CrossRef]
- Véla, E.; Tirard, A.; Renucci, M.; Suehs, C.M.; Provost, E. Floral chemical signatures in the genus Ophrys L.(Orchidaceae): A preliminary test of a new tool for taxonomy and evolution. Plant Mol. Biol. Rep. 2007, 25, 83–97. [Google Scholar] [CrossRef]
- Joffard, N.; Buatois, B.; Schatz, B. Integrative taxonomy of the fly orchid group: Insights from chemical ecology. Sci. Nat. 2016, 103, 77. [Google Scholar] [CrossRef]
- Mant, J.; Peakall, R.; Schiestl, F.P. Does selection on floral odor promote differentiation among populations and species of the sexually deceptive orchid genus Ophrys? Evolution 2005, 59, 1449–1463. [Google Scholar] [CrossRef]
- Bohman, B.; Jeffares, L.; Flematti, G.R.; Byrne, L.T.; Skelton, B.W.; Phillips, R.D.; Dixon, K.W.; Peakall, R.; Barrow, R.A. Discovery of tetrasubstituted pyrazines as semiochemicals in a sexually deceptive orchid. J. Nat. Prod. 2012, 75, 1589–1594. [Google Scholar] [CrossRef]
- Bohman, B.; Peakall, R. Pyrazines attract Catocheilus thynnine wasps. Insects 2014, 5, 474–487. [Google Scholar] [CrossRef]
- Bohman, B.; Phillips, R.D.; Menz, M.H.M.; Berntsson, B.W.; Flematti, G.R.; Barrow, R.A.; Dixon, K.W.; Peakall, R. Discovery of pyrazines as pollinator sex pheromones and orchid semiochemicals: Implications for the evolution of sexual deception. New Phytol. 2014, 203, 939–952. [Google Scholar] [CrossRef] [PubMed]
- Bohman, B.; Tan, M.; Phillips, R.; Scaffidi, A.; Sobolev, A.; Moggach, S.; Flematti, G.; Peakall, R. A specific blend of drakolide and hydroxymethylpyrazines–an unusual pollinator sexual attractant used by the endangered orchid Drakaea micrantha. Angew. Chem. Int. Ed. 2019, 59, 1124–1128. [Google Scholar] [CrossRef] [PubMed]
- Bohman, B.; Jeffares, L.; Flematti, G.R.; Phillips, R.D.; Dixon, K.W.; Peakall, R.; Barrow, R.A. The discovery of 2-hydroxymethyl-3-(3-methylbutyl)-5-methylpyrazine: A semiochemical in orchid pollination. Org. Lett. 2012, 14, 2576–2578. [Google Scholar] [CrossRef] [PubMed]
- Hopper, S.D.; Brown, A.P. A revision of Australia’s hammer orchids (Drakaea: Orchidaceae), with some field data on species-specific sexually deceived wasp pollinators. Aust. Syst. Bot. 2007, 20, 252–285. [Google Scholar] [CrossRef]
- Phillips, R.D.; Peakall, R.; Hutchinson, M.F.; Linde, C.C.; Xu, T.; Dixon, K.W.; Hopper, S.D. Specialized ecological interactions and plant species rarity: The role of pollinators and mycorrhizal fungi across multiple spatial scales. Biol. Conserv. 2014, 169, 285–295. [Google Scholar] [CrossRef]
- Bohman, B.; Flematti, G. Oxidation of hydroxymethylpyrazines and hydroxylated phenyl compounds in a gas chromatography inlet. Anal. Methods 2015, 7, 2283–2286. [Google Scholar] [CrossRef]
- Phillips, R.D.; Xu, T.; Hutchinson, M.F.; Dixon, K.W.; Peakall, R. Convergent specialization–the sharing of pollinators by sympatric genera of sexually deceptive orchids. J. Ecol. 2013, 101, 826–835. [Google Scholar] [CrossRef]
- Gaskett, A.C.; Winnick, C.G.; Herberstein, M.E. Orchid sexual deceit provokes ejaculation. Am. Nat. 2008, 171, E206–E212. [Google Scholar] [CrossRef] [Green Version]
- Emlen, S.T. Lek organization and mating strategies in the bullfrog. Behav. Ecol. Sociobiol. 1976, 1, 283–313. [Google Scholar] [CrossRef]
- Bretman, A.; Gage, M.J.; Chapman, T. Quick-change artists: Male plastic behavioural responses to rivals. Trends Ecol. Evol. 2011, 26, 467–473. [Google Scholar] [CrossRef]
- Robertson, J.L.; Wyatt, R. Evidence for pollination ecotypes in the yellow-fringed orchid, Platanthera ciliaris. Evolution 1990, 44, 121–133. [Google Scholar] [CrossRef] [PubMed]
- Johnson, S. Pollination ecotypes of Satyrium hallackii (Orchidaceae) in South Africa. Bot. J. Linn. Soc. 1997, 123, 225–235. [Google Scholar] [CrossRef]
- Ayasse, M.; Schiestl, F.P.; Paulus, H.F.; Löfstedt, C.; Hansson, B.; Ibarra, F.; Francke, W. Evolution of reproductive strategies in the sexually deceptive orchid Ophrys sphegodes: How does flower-specific variation of odor signals influence reproductive success? Evolution 2000, 54, 1995–2006. [Google Scholar] [CrossRef] [PubMed]
- Schiestl, F.P.; Cozzolino, S. Evolution of sexual mimicry in the orchid subtribe orchidinae: The role of preadaptations in the attraction of male bees as pollinators. BMC Evol. Biol. 2008, 8, 1–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bohman, B.R.; Weinstein, A.M.; Phillips, R.D.; Peakall, R.; Flematti, G.R. 2-(Tetrahydrofuran-2-yl) acetic acid and ester derivatives as long-range pollinator attractants in the sexually deceptive orchid Cryptostylis ovata. J. Nat. Prod. 2019, 82, 1107–1113. [Google Scholar] [CrossRef] [PubMed]
- Bohman, B.; Phillips, R.D.; Flematti, G.; Peakall, R.; Barrow, R.A. Sharing of pyrazine semiochemicals between genera of sexually deceptive orchids. Nat. Prod. Commun. 2013, 8, 701–702. [Google Scholar] [CrossRef] [Green Version]
- Jacob, V.E.J.M. Current source density analysis of electroantennogram recordings: A tool for mapping the olfactory response in an insect antenna. Front. Cell. Neurosci. 2018, 12, 287. [Google Scholar] [CrossRef] [Green Version]
- Mayr, E. Systematics and the Origin of Species, from the Viewpoint of a Zoologist; Harvard University Press: Cambridge, MA, USA, 1942. [Google Scholar]
- Phillips, R.D.; Brown, A.P.; Dixon, K.W.; Hopper, S.D. Orchid biogeography and factors associated with rarity in a biodiversity hotspot, the Southwest Australian Floristic Region. J. Biogeogr. 2011, 38, 487–501. [Google Scholar] [CrossRef]
- Horwitz, P.; Bradshaw, D.; Hopper, S.; Davies, P.; Froend, R.; Bradshaw, F. Hydrological change escalates risk of ecosystem stress in Australia’s threatened biodiversity hotspot. J. R. Soc. West. Aust. 2008, 91, 1–11. [Google Scholar]
- Phillips, R.D.; Peakall, R.; Retter, B.A.; Montgomery, K.; Menz, M.H.; Davis, B.J.; Hayes, C.; Brown, G.R.; Swarts, N.D.; Dixon, K.W. Pollinator rarity as a threat to a plant with a specialized pollination system. Bot. J. Linn. Soc. 2015, 179, 511–525. [Google Scholar] [CrossRef] [Green Version]
- Gaskett, A.C.; Endler, J.A.; Phillips, R.D. Convergent evolution of sexual deception via chromatic and achromatic contrast rather than colour mimicry. Evol. Ecol. 2017, 31, 205–227. [Google Scholar] [CrossRef]
- Linde, C.C.; May, T.W.; Phillips, R.D.; Ruibal, M.; Smith, L.M.; Peakall, R. New species of Tulasnella associated with terrestrial orchids in Australia. IMA Fungus 2017, 8, 28–48. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R. Responses of male Zaspilothynnus trilobatus Turner wasps to females and the sexually deceptive orchid it pollinates. Funct. Ecol. 1990, 4, 159–167. [Google Scholar] [CrossRef]
- Bower, C.C. Demonstration of pollinator-mediated reproductive isolation in sexually deceptive species of Chiloglottis (Orchidaceae: Caladeniinae). Aust. J. Bot. 1996, 44, 15–33. [Google Scholar] [CrossRef]
- Menz, M.H.M.; Phillips, R.D.; Dixon, K.W.; Peakall, R.; Didham, R.K. Mate-Searching Behaviour of Common and Rare Wasps and the Implications for Pollen Movement of the Sexually Deceptive Orchids They Pollinate. PLoS ONE 2013, 8, e59111. [Google Scholar] [CrossRef] [Green Version]
- Peakall, R.; Smouse, P.E. GENALEX 6: Genetic analysis in Excel. Population genetic software for teaching and research. Mol. Ecol. Notes 2006, 6, 288–295. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2018. [Google Scholar]
- Falara, V.; Amarasinghe, R.; Poldy, J.; Pichersky, E.; Barrow, R.A.; Peakall, R. The production of a key floral volatile is dependent on UV light in a sexually deceptive orchid. Ann. Bot. 2013, 111, 21–30. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- O’Callaghan, S.; De Souza, D.P.; Isaac, A.; Wang, Q.; Hodkinson, L.; Olshansky, M.; Erwin, T.; Appelbe, B.; Tull, D.L.; Roessner, U. PyMS: A Python toolkit for processing of gas chromatography-mass spectrometry (GC-MS) data. Application and comparative study of selected tools. BMC Bioinform. 2012, 13, 115. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hervé, M.R.; Nicolè, F.; Lê Cao, K.-A. Multivariate analysis of multiple datasets: A practical guide for chemical ecology. J. Chem. Ecol. 2018, 44, 215–234. [Google Scholar] [CrossRef] [PubMed]
- Wold, S.; Sjöström, M.; Eriksson, L. PLS-regression: A basic tool of chemometrics. Chemom. Intell. Lab. Syst. 2001, 58, 109–130. [Google Scholar] [CrossRef]
- Finch, H. Comparison of distance measures in cluster analysis with dichotomous data. J. Data Sci. 2005, 3, 85–100. [Google Scholar] [CrossRef]
- Oksanen, J.; Guillaume Blanchet, F.; Friendly, M.; Kindt, R.; Legendre, P.; McGlinn, D.; Minchin, P.R.; O’Hara, R.B.; Simpson, G.L.; Solymos, P.; et al. Vegan: Community Ecology Package. R Package Version 2.5-3. 2018. Available online: https://CRAN.R-project.org/web/packages/vegan/index.html (accessed on 13 January 2022).
- Paradis, E.; Claude, J.; Strimmer, K. APE: Analyses of phylogenetics and evolution in R language. Bioinformatics 2004, 20, 289–290. [Google Scholar] [CrossRef] [Green Version]
- Davies, T. The new automated mass spectrometry deconvolution and identification system (AMDIS). Spectrosc. Eur. 1998, 10, 24–27. [Google Scholar]
- Dowle, M.; Srinivasan, A. Data.Table: Extension of ‘Data.Frame’. R Package Version 1.11.8. 2018. Available online: https://CRAN.R-project.org/package=data.table (accessed on 13 January 2022).
- Wickham, H. Reshaping data with the reshape package. J. Stat. Softw. 2007, 21, 1–20. [Google Scholar] [CrossRef]


{kind=link}
{kind=link}
{kind=link}
| Zaspilothynnus nigripes Responses | N Sites Surveyed | % Sites Present | Average Number of Wasps per Survey ± SE | Total Wasps Observed |
|---|---|---|---|---|
| Zaspilothynnus nigripes pollinated populations | 9 | 100 | 12.00 ± 2.49 * | 108 |
| Catocheilus sp. pollinated populations | 12 | 66.7 | 4.42 ± 1.58 | 53 |
| Zaspilothynnus dilatatus pollinated populations | 7 | 71.4 | 3.00 ± 1.11 | 21 |
| Zaspilothynnus dilatatus responses | ||||
| Zaspilothynnus nigripes pollinated populations | 9 | 0 | 0 | 0 |
| Catocheilus sp. pollinated populations | 12 | 0 | 0 | 0 |
| Zaspilothynnus dilatatus pollinated populations | 7 | 85.7 | 2.14 ± 0.46 * | 15 |
| Catocheilus sp. responses | ||||
| Zaspilothynnus nigripes pollinated populations | 9 | 0 | 0 | 0 |
| Catocheilus sp. pollinated populations | 12 | 0 | 0 | 0 |
| Zaspilothynnus dilatatus pollinated populations | not surveyed | not surveyed | not surveyed | not surveyed |
| No. | Pollinator Association | Name | Characteristic Mass Fragments (EI) | RI | Detection Method |
|---|---|---|---|---|---|
| 1 | Zaspilothynnus nigripes | 2-hydroxymethyl- 3-(3-methylbutyl)- 5-methylpyrazine | 194, 163, 138, 109 | 1532 | EAD [46]/Extract analyses |
| 2 | Zaspilothynnus nigripes | Unknown 1 | 168, 150, 139, 122 | 1557 | Extract analyses |
| 3 | Zaspilothynnus nigripes | Unknown 2 | 196, 154, 136, 108 | 1804 | Extract analyses |
| 4 | Zaspilothynnus nigripes/Catocheilus sp. | 4-(2-hydroxyethyl)-2-methoxyphenol (homovanillyl alcohol) | 168, 150, 137, 122 | 1547 | Extract analyses |
| 5 | Zaspilothynnis nigripes/Catocheilus sp. | Unknown 8 | 208, 124, 107, 77 | 1722 | Extract analyses |
| 6 | Zaspilothynnis nigripes/Z. dilatatus | Heneicosene (unknown isomer) | 294, 11, 97, 83, 55 | 2086 | Extract analyses |
| 7 | Catocheilus sp. | 3,5,6-trimethylpyrazine-2-carbaldehyde | 150, 122, 121, 107 | 1207 | Extract analyses |
| 8 | Catocheilus sp. | 2-hydroxymethyl-3,5,6-trimethylpyrazine | 152, 151, 134, 123 | 1299 | EAD [42]/Extract analyses |
| 9 | Catocheilus sp. | 2-(3-methylbutyl)-3,5,6-trimethylpyrazine | 191, 177, 149, 136 | 1389 | EAD [42]/Extract analyses |
| 10 | Catocheilus sp. | Unknown 3 | 168, 151, 139, 121 | 1538 | Extract analyses |
| 11 | Catocheilus sp. | Unknown 4 | 208, 193, 175, 149 | 1568 | Extract analyses |
| 12 | Catocheilus sp. | (3,6-dimethylpyrazin-2-yl)methyl 3-methylbutanoate | 222, 180, 138, 121 | 1580 | EAD [42]/Extract analyses |
| 13 | Catocheilus sp. | (3,5,6-trimethylpyrazin-2-yl)methyl-3-methylbutanoate | 236, 208, 152, 151 | 1660 | EAD [42]/Extract analyses |
| 14 | Catocheilus sp. | (3,5,6-trimethylpyrazin-2-yl)methyl(2S)-methylbutanoate | 236, 194, 152, 151 | 1667 | EAD [42]/Extract analyses |
| 15 | Catocheilus sp. | Unknown 5 | 252, 168, 151, 138 | 1899 | Extract analyses |
| 16 | Catocheilus sp. | Unknown 7 | 253, 168, 151, 121 | 2001 | Extract analyses |
| 17 | Catocheilus sp. | Unknown 6 | 210, 168, 151, 122 | 2022 | Extract analyses |
| 18 | Zaspilothynnus dilatatus | 2-(methylthio)benzene-1,4-diol | 156, 141, 113, 97 | 1507 | EAD |
| 19 | Zaspilothynnus dilatatus | 4-hydroxy-3-(methylthio)benzaldehyde | 168, 167, 139, 97 | 1507 | EAD/Extract analyses |
| 20 | Zaspilothynnus dilatatus | 4-(hydroxymethyl)-2-(methylthio)phenol | 170, 153, 141, 123 | 1560 | Extract analyses |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Weinstein, A.M.; Bohman, B.; Flematti, G.R.; Phillips, R.D. Three Chemically Distinct Floral Ecotypes in Drakaea livida, an Orchid Pollinated by Sexual Deception of Thynnine Wasps. Plants 2022, 11, 260. https://doi.org/10.3390/plants11030260
Weinstein AM, Bohman B, Flematti GR, Phillips RD. Three Chemically Distinct Floral Ecotypes in Drakaea livida, an Orchid Pollinated by Sexual Deception of Thynnine Wasps. Plants. 2022; 11(3):260. https://doi.org/10.3390/plants11030260
Chicago/Turabian StyleWeinstein, Alyssa M., Björn Bohman, Gavin R. Flematti, and Ryan D. Phillips. 2022. "Three Chemically Distinct Floral Ecotypes in Drakaea livida, an Orchid Pollinated by Sexual Deception of Thynnine Wasps" Plants 11, no. 3: 260. https://doi.org/10.3390/plants11030260
APA StyleWeinstein, A. M., Bohman, B., Flematti, G. R., & Phillips, R. D. (2022). Three Chemically Distinct Floral Ecotypes in Drakaea livida, an Orchid Pollinated by Sexual Deception of Thynnine Wasps. Plants, 11(3), 260. https://doi.org/10.3390/plants11030260