
Fly Pollination of Kettle Trap Flowers of Riocreuxia torulosa (Ceropegieae-Anisotominae): A Generalized System of Floral Deception

 , ,
, ,  , ,
, ,  and
and
Abstract
:
1. Introduction
2. Results
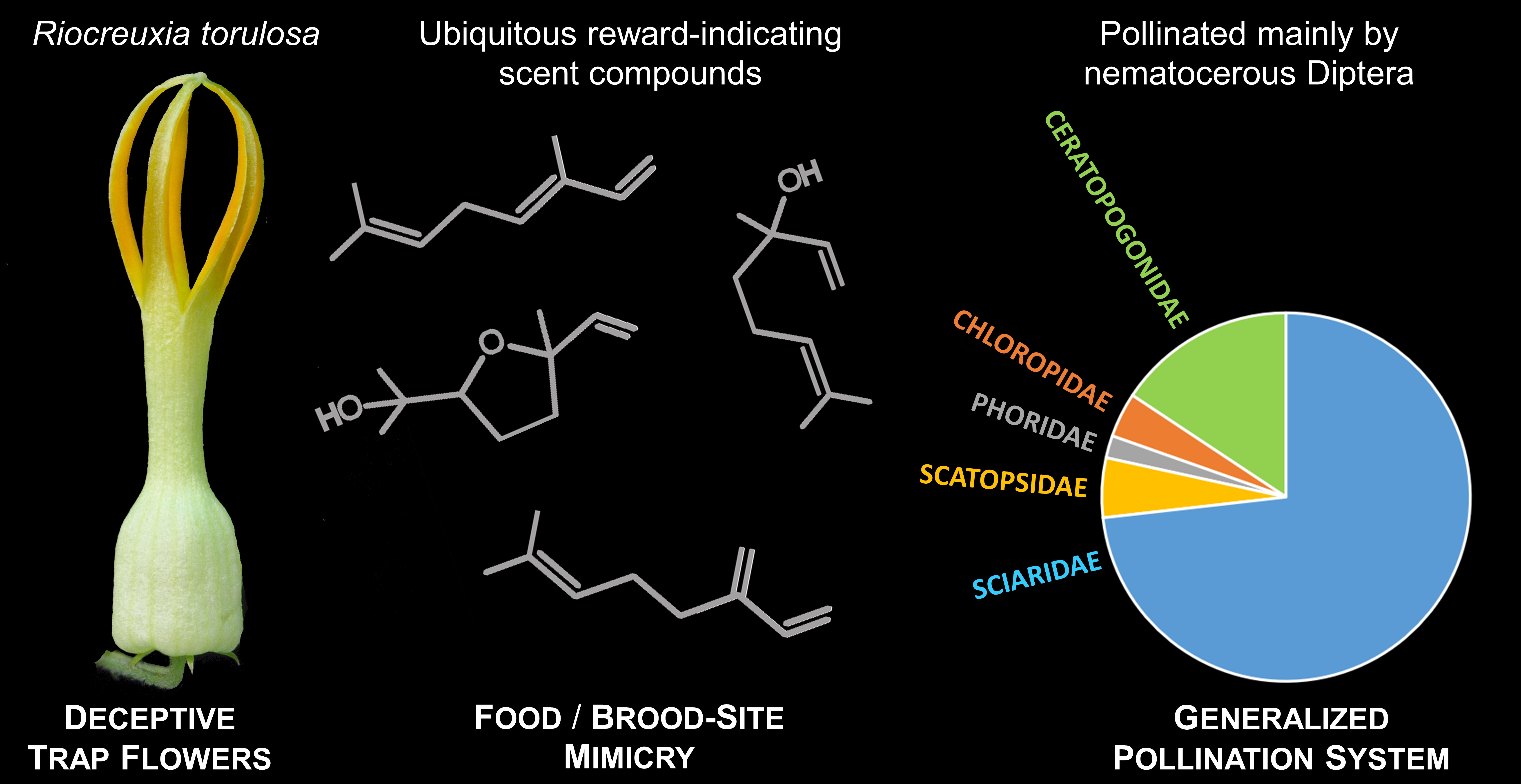
2.1. Flower Visitors and Pollinators
2.2. Pollination Success
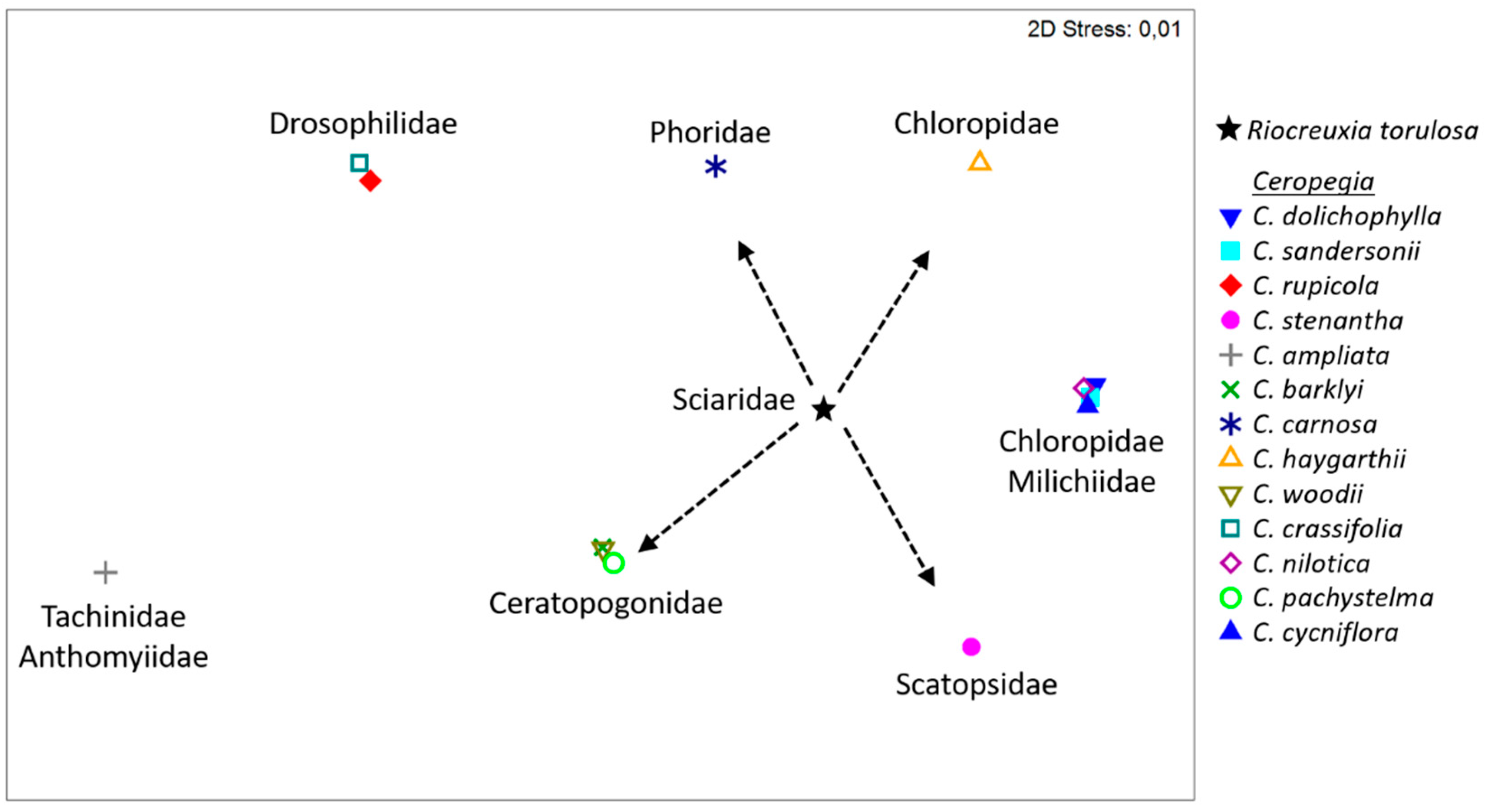
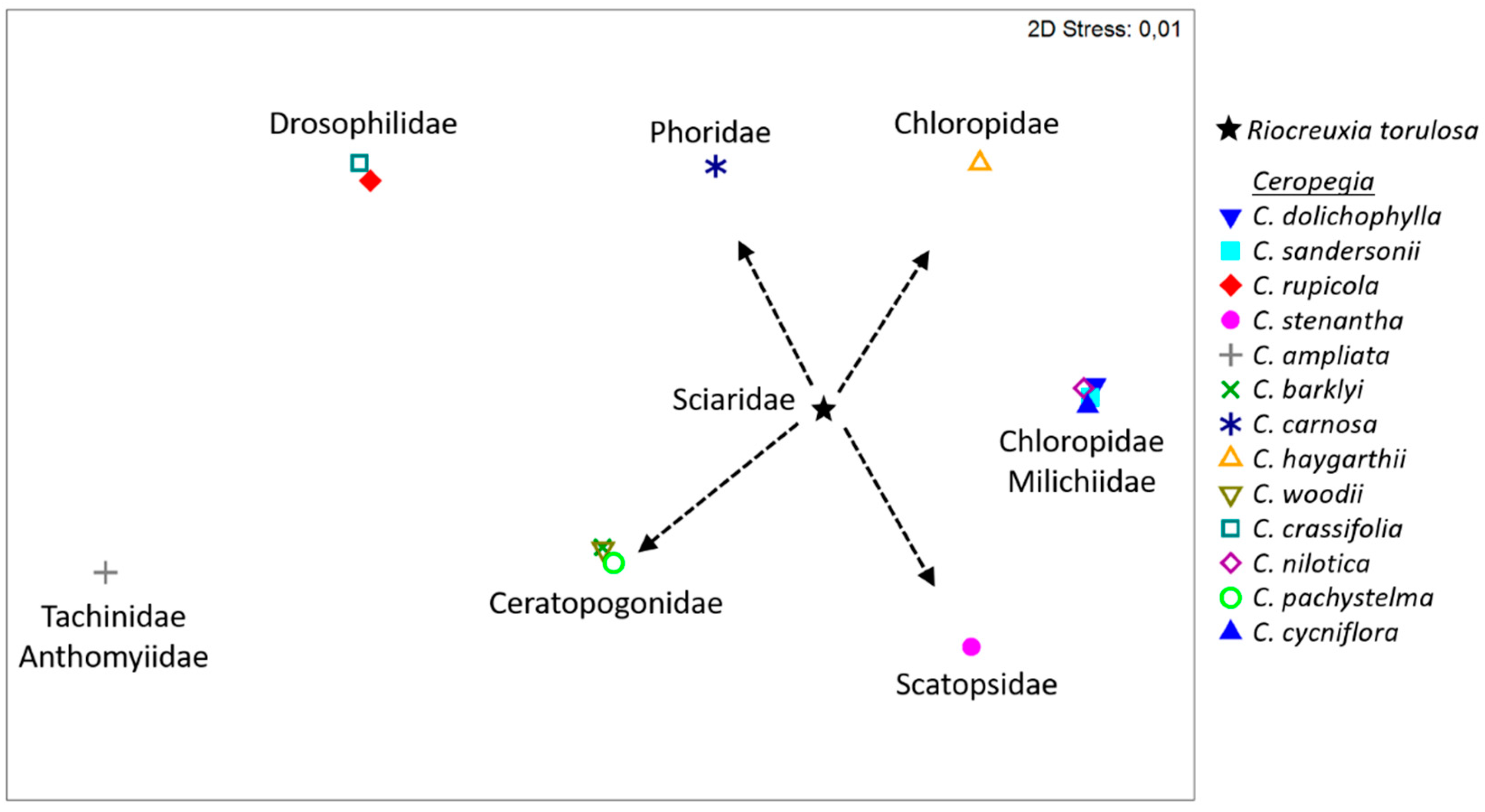
2.3. Floral Scent
3. Discussion
4. Conclusions
5. Materials and Methods
5.1. Plant Material and Study Sites
5.2. Flower Visitors and Pollinators
5.3. Pollination Success and Natural Seed Set
5.4. Collection of Floral Volatiles
5.5. Chemical Analyses
5.6. Statistical Analyses
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Stebbins, G. Adaptive radiation of reproductive characteristics in angiosperms, I: Pollination mechanisms. Annu. Rev. Ecol. Evol. Syst. 1970, 1, 307–326. [Google Scholar] [CrossRef]
- Kay, K.M.; Voelckel, C.; Yang, J.Y.; Hufford, K.M.; Kaska, D.D.; Hodges, S.A. Floral characters and species diversification. In Ecology and Evolution of Flowers; Harder, L.D., Sch, B., Eds.; Oxford University Press: Oxford, UK, 2006; pp. 311–325. [Google Scholar]
- Ollerton, J.; Masinde, S.; Meve, U.; Picker, M.; Whittington, A. Fly pollination in Ceropegia (Apocynaceae: Asclepiadoideae): Biogeographic and phylogenetic perspectives. Ann. Bot. 2009, 103, 1501–1514. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vogel, S. Kesselfallen-Blumen. Umsch. Wiss. Tech. 1965, 65, 12–17. [Google Scholar]
- Johnson, S.D.; Schiestl, F.P. Floral Mimicry; Oxford University Press: Oxford, UK, 2016. [Google Scholar]
- Meve, U.; Heiduk, A.; Liede-Schumann, S. Origin and early evolution of Ceropegieae (Apocynaceae-Asclepiadoideae). Syst. Biodivers. 2017, 15, 143–155. [Google Scholar] [CrossRef]
- Endress, M.E.; Meve, U.; Middleton, D.J.; Liede-Schumann, S. Apocynaceae. In Flowering Plants. The Families and Genera of Vascular Plants, Vol. 15, Eudicots, Apiales, Gentianales (excl. Rubiaceae); Kadereit, J.W., Bittrich, V., Eds.; Springer: Berlin/Heidelberg, Germany; New York, NY, USA, 2018; pp. 207–411. [Google Scholar] [CrossRef]
- Meve, U.; Liede-Schumann, S. Ceropegia (Apocynaceae, Ceropegieae, Stapeliinae): Paraphyletic but still taxonomically sound. Ann. Mo. Bot. Gard. 2007, 94, 392–406. [Google Scholar] [CrossRef]
- Bruyns, P.; Klak, C.; Hanáček, P. A revised, phylogenetically-based concept of Ceropegia (Apocynaceae). S. Afr. J. Bot. 2017, 112, 399–436. [Google Scholar] [CrossRef]
- Meyer, E. Commentatorium de Plantis Africae Australiorum; fasc. 2: 194; Leopold Voss: Leipzig, Germany, 1838; Volume I. [Google Scholar]
- Masinde, P.S. A revision of the African genus Riocreuxia Decne. (Apocynaceae: Asclepiadoideae-Ceropegieae). Kew Bull. 2005, 60, 401–434. [Google Scholar]
- Vogel, S. Die Bestäubung der Kesselfallen-Blüten von Ceropegia. Beitr. Biol. Pfl. 1961, 36, 159–237. [Google Scholar]
- Dyer, R.A. Ceropegia, Brachystelma and Riocreuxia in Southern Africa; AA Balkema: Rotterdam, The Netherlands, 1983. [Google Scholar]
- Vogel, S. Über die “Uvula” von Ceropegia sandersonii Hook. f.-zugleich über einen merkwürdigen Fall postgenitaler Verwachsung. Beitr. Biol. Pfl. 1960, 35, 395–412. [Google Scholar]
- Masinde, P.S. Trap-flower fly pollination in East African Ceropegia L. (Apocynaceae). Int. J. Trop. Insect Sci. 2004, 24, 55–72. [Google Scholar] [CrossRef]
- Ollerton, J.; Dötterl, S.; Ghorpadé, K.; Heiduk, A.; Liede-Schumann, S.; Masinde, S.; Meve, U.; Craig, P.I.; Prieto-Benítez, S.; Punekar, S.; et al. Diversity of Diptera families that pollinate Ceropegia (Apocynaceae) trap flowers: An update in light of new data and phylogenetic analyses. Flora 2017, 234, 233–244. [Google Scholar] [CrossRef] [Green Version]
- Coombs, G.; Dold, A.P.; Peter, C.I. Generalized fly-pollination in Ceropegia ampliata (Apocynaceae-Asclepiadoideae): The role of trapping hairs in pollen export and receipt. Plant Sys. Evol. 2011, 296, 137–148. [Google Scholar] [CrossRef]
- Heiduk, A.; Brake, I.; Tolasch, T.; Frank, J.; Jürgens, A.; Meve, U.; Dötterl, S. Scent chemistry and pollinator attraction in the deceptive trap flowers of Ceropegia dolichophylla. S. Afr. J. Bot. 2010, 76, 762–769. [Google Scholar] [CrossRef] [Green Version]
- Heiduk, A.; Brake, I.; von Tschirnhaus, M.; Haenni, J.-P.; Miller, R.; Hash, J.; Prieto-Benítez, S.; Jürgens, A.; Johnson, S.D.; Schulz, S.; et al. Floral scent and pollinators of Ceropegia trap flowers. Flora 2017, 232, 169–182. [Google Scholar] [CrossRef]
- Heiduk, A.; Brake, I.; von Tschirnhaus, M.; Göhl, M.; Jürgens, A.; Johnson, S.D.; Meve, U.; Dötterl, S. Ceropegia sandersonii mimics attacked honeybees to attract kleptoparasitic flies for pollination. Curr. Biol. 2016, 26, 2787–2793. [Google Scholar] [CrossRef] [PubMed]
- Heiduk, A.; Haenni, J.-P.; Meve, U.; Schulz, S.; Dötterl, S. Flower scent of Ceropegia stenantha: Electrophysiological activity and synthesis of novel components. J. Comp. Physiol. A 2019, 205, 301–310. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Heiduk, A.; Kong, H.; Brake, I.; von Tschirnhaus, M.; Tolasch, T.; Tröger, A.; Wittenberg, E.; Francke, W.; Meve, U.; Dötterl, S. Deceptive Ceropegia dolichophylla fools its kleptoparasitic fly pollinators with exceptional floral scent. Front. Ecol. Evol. 2015, 3. [Google Scholar] [CrossRef] [Green Version]
- Ollerton, J.; Liede-Schumann, S.; Endress, M.E.; Meve, U.; Rech, A.R.; Shuttleworth, A.; Keller, H.A.; Fishbein, M.; Alvarado-Cárdenas, L.O.; Amorim, F.W. The diversity and evolution of pollination systems in large plant clades: Apocynaceae as a case study. Ann. Bot. 2019, 123, 311–325. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Koschnitzke, C. Bradysia sp. (Diptera: Sciaridae), a pollinator that can die in flowers of Ditassa banksii Schult. (Apocynaceae, Asclepiadoideae). Hoehnea 2018, 45, 126–128. [Google Scholar] [CrossRef] [Green Version]
- Sakai, S. Aristolochia spp. (Aristolochiaceae) pollinated by flies breeding on decomposing flowers in Panama. Am. J. Bot. 2002, 89, 527–534. [Google Scholar] [CrossRef] [Green Version]
- Borkent, C.; Hauser, M.; Shepard, W. Arthropods visiting flowers of Aristolochia californica Torr. (Aristolochiaceae). Pan Pac. Entomol. 2020, 96, 79–89. [Google Scholar] [CrossRef]
- Rulik, B.; Wanke, S.; Nuss, M.; Neinhuis, C. Pollination of Aristolochia pallida Willd. (Aristolochiaceae) in the Mediterranean. Flora 2008, 203, 175–184. [Google Scholar] [CrossRef]
- Vogel, S.; Martens, J. A survey of the function of the lethal kettle traps of Arisaema (Araceae), with records of pollinating fungus gnats from Nepal. Bot. J. Linn. Soc. 2000, 133, 61–100. [Google Scholar] [CrossRef]
- Matsumoto, T.K.; Hirobe, M.; Sueyoshi, M.; Miyazaki, Y. Selective pollination by fungus gnats potentially functions as an alternative reproductive isolation among five Arisaema species. Ann. Bot. 2021, 127, 633–644. [Google Scholar] [CrossRef]
- Kakishima, S.; Okuyama, Y. Pollinator assemblages of Arisaema heterocephalum subsp. majus (Araceae), a critically endangered species endemic to Tokunoshima island, central Ryukyus. Bull. Natl. Mus. Nat. Sci. Ser. B Bot. 2018, 44, 173–179. [Google Scholar]
- Kakishima, S.; Sueyoshi, M.; Okuyama, Y. Floral visitors of critically endangered Arisaema cucullatum (Araceae) endemic to Kinki region of Japan. Bull. Natl. Mus. Nat. Sci. Ser. B Bot. 2020, 46, 47–53. [Google Scholar]
- Barriault, I.; Barabé, D.; Cloutier, L.; Gibernau, M. Pollination ecology and reproductive success in Jack-in-the-pulpit (Arisaema triphyllum) in Québec (Canada). Plant Biol. 2010, 12, 161–171. [Google Scholar] [CrossRef]
- Borba, E.L.; Semir, J. Pollinator specificity and convergence in fly-pollinated Pleurothallis (Orchidaceae) species: A multiple population approach. Ann. Bot. 2001, 88, 75–88. [Google Scholar] [CrossRef] [Green Version]
- Bogarín, D.; Fernández, M.; Borkent, A.; Heemskerk, A.; Pupulin, F.; Ramírez, S.; Smets, E.; Gravendeel, B. Pollination of Trichosalpinx (Orchidaceae: Pleurothallidinae) by biting midges (Diptera: Ceratopogonidae). Bot. J. Linn. Soc. 2018, 186, 510–543. [Google Scholar] [CrossRef]
- Blanco, M.A.; Barboza, G. Pseudocopulatory pollination in Lepanthes (Orchidaceae: Pleurothallidinae) by fungus gnats. Ann. Bot. 2005, 95, 763–772. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Phillips, R.D.; Scaccabarozzi, D.; Retter, B.A.; Hayes, C.; Brown, G.R.; Dixon, K.W.; Peakall, R. Caught in the act: Pollination of sexually deceptive trap-flowers by fungus gnats in Pterostylis (Orchidaceae). Ann. Bot. 2014, 113, 629–641. [Google Scholar] [CrossRef] [Green Version]
- Hayashi, T.; Bohman, B.; Scaffidi, A.; Peakall, R.; Flematti, G.R. An unusual tricosatriene is crucial for male fungus gnat attraction and exploitation by sexually deceptive Pterostylis orchids. Curr. Biol. 2021, 31, 1954–1961.e7. [Google Scholar] [CrossRef]
- Toledo-Hernández, M.; Wanger, T.C.; Tscharntke, T. Neglected pollinators: Can enhanced pollination services improve cocoa yields? A review. Agric. Ecosyst. Environ. 2017, 247, 137–148. [Google Scholar] [CrossRef]
- Hartop, E.; Häggqvist, S.; Ulefors, S.O.; Ronquist, F. Scuttling towards monophyly: Phylogeny of the mega-diverse genus Megaselia (Diptera: Phoridae). Syst. Entomol. 2021, 46, 71–82. [Google Scholar] [CrossRef]
- Borkent, A. 34. Ceratopogonideae (Biting Midges). In Manual of Afrotropical Diptera. Volume 2. Nematocerous Diptera and lower Brachycera; Kirk-Spriggs, A., Sinclair, B., Eds.; SANBI South African National Biodiversity Institute: Pretoria, South Africa, 2017; pp. 733–812. [Google Scholar]
- Menzel, F.; Smith, J.E. 21. Sciaridae (Black Fungus Gnats). In Manual of Afrotropical Diptera. Volume 2. Nematocerous Diptera and lower Brachycera; Kirk-Spriggs, A., Sinclair, B., Eds.; SANBI South African National Biodiversity Institute: Pretoria, South Africa, 2017; pp. 557–580. [Google Scholar]
- Auttama, P.; McKey, D.; Kidyoo, A. Flowering phenology and trap pollination of the rare endemic plant Ceropegia thaithongiae in montane forest of northern Thailand. Botany 2018, 96, 601–620. [Google Scholar] [CrossRef]
- Bauer, A.A.; Clayton, M.K.; Brunet, J. Floral traits influencing plant attractiveness to three bee species: Consequences for plant reproductive success. Am. J. Bot. 2017, 104, 772–781. [Google Scholar] [CrossRef] [Green Version]
- Sun, H.Q.; Huang, B.Q.; Yu, X.H.; Tian, C.B.; Peng, Q.X.; An, D.J. Pollen limitation, reproductive success and flowering frequency in single-flowered plants. J. Ecol. 2018, 106, 19–30. [Google Scholar] [CrossRef] [Green Version]
- Rupp, T.; Oelschlägel, B.; Rabitsch, K.; Mahfoud, H.; Wenke, T.; Disney, R.H.L.; Neinhuis, C.; Wanke, S.; Dötterl, S. Flowers of deceptive Aristolochia microstoma are pollinated by phorid flies and emit volatiles known from invertebrate carrion. Front. Ecol. Evol. 2021, 9. [Google Scholar] [CrossRef]
- Jürgens, A.; Shuttleworth, A. Carrion and dung mimicry in plants. In Carrion Ecology, Evolution, and Their Applications; Benbow, E.M., Tomberlin, J.K., Tarone, A.M., Eds.; CRC Press: Boca Raton, FL, USA, 2015; pp. 361–386. [Google Scholar]
- Jürgens, A.; Wee, S.-L.; Shuttleworth, A.; Johnson, S.D. Chemical mimicry of insect oviposition sites: A global analysis of convergence in angiosperms. Ecol. Lett. 2013, 16, 1157–1167. [Google Scholar] [CrossRef] [PubMed]
- Fontes, A.L.; Valentin-Silva, A.; Demuner, A.J.; Vieira, M.F. Functional morphology and floral odor of Ditassa burchellii (Apocynaceae, Asclepiadoideae), a micromyiophilous species from southeastern Brazil. Int. J. Plant Sci. 2018, 179, 450–460. [Google Scholar] [CrossRef]
- Johnson, S.; Jürgens, A. Convergent evolution of carrion and faecal scent mimicry in fly-pollinated angiosperm flowers and a stinkhorn fungus. S. Afr. J. Bot. 2010, 76, 796–807. [Google Scholar] [CrossRef] [Green Version]
- Knudsen, J.T.; Eriksson, R.; Gershenzon, J.; Ståhl, B. Diversity and distribution of floral scent. Bot. Rev. 2006, 72, 1–120. [Google Scholar] [CrossRef]
- Andersson, S. Antennal responses to floral scents in the butterflies Inachis io, Aglais urticae (Nymphalidae), and Gonepteryx rhamni (Pieridae). Chemoecology 2003, 13, 13–20. [Google Scholar] [CrossRef]
- Andersson, S.; Nilsson, L.A.; Groth, I.; Bergström, G. Floral scents in butterfly-pollinated plants: Possible convergence in chemical composition. Bot. J. Linn. Soc. 2002, 140, 129–153. [Google Scholar] [CrossRef] [Green Version]
- Steenhuisen, S.-L.; Jürgens, A.; Johnson, S.D. Effects of volatile compounds emitted by Protea species (Proteaceae) on antennal electrophysiological responses and attraction of Cetoniine beetles. J. Chem. Ecol. 2013, 39, 438–446. [Google Scholar] [CrossRef] [PubMed]
- Dötterl, S.; Vereecken, N. The chemical ecology and evolution of bee-flower interactions: A review and perspectives. Can. J. Zool. 2010, 88, 668–697. [Google Scholar] [CrossRef]
- Kotze, M. Olfactory Responses of Dasineura dielsi Rübsaamen (Diptera: Cecidomyiidae) Females to Host Plant Volatiles; University of Cape Town: Cape Town, South Africa, 2012. [Google Scholar]
- Jhumur, U.S.; Dötterl, S.; Jürgens, A. Floral odors of Silene otites: Their variability and attractiveness to mosquitoes. J. Chem. Ecol. 2008, 34, 14–25. [Google Scholar] [CrossRef]
- Martos, F.; Cariou, M.L.; Pailler, T.; Fournel, J.; Bytebier, B.; Johnson, S.D. Chemical and morphological filters in a specialized floral mimicry system. New Phytol. 2015, 207, 225–234. [Google Scholar] [CrossRef]
- Ollerton, J. Fly trapping in Ceropegia flowers—Evidence of ant predation of pollinators. Asklepios 1999, 77, 31–32. [Google Scholar]
- Harder, L.D.; Johnson, S.D. Function and evolution of aggregated pollen in angiosperms. Int. J. Plant Sci. 2008, 169, 59–78. [Google Scholar] [CrossRef]
- Dötterl, S.; Wolfe, L.M.; Jürgens, A. Qualitative and quantitative analyses of flower scent in Silene latifolia. Phytochemistry 2005, 66, 203–213. [Google Scholar] [CrossRef] [PubMed]
- Amirav, A.; Dagan, S. A direct sample introduction device for mass spectrometry studies and gas chromatography mass spectrometry analyses. Eur. Mass Spectrom. 1997, 3, 105–111. [Google Scholar] [CrossRef]
- Adams, R.P. Identification of Essential Oil Components by Gas Chromatography/Mass Spectrometry, 4th ed.; Allured Publishing Corporation: Carol Stream, IL, USA, 2007. [Google Scholar]
- Clarke, K.R.; Gorley, R.N. Primer v6: User Manual/Tutorial; Primer-E Ltd.: Plymouth, UK, 2006. [Google Scholar]



{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Total Number of Diptera: | Male | Female |
|---|---|---|
| 24 (12) | 130 (52) | |
| CERATOPOGONIDAE | 2 | 22 (6) |
| Forcipomyia Meigen, 1818 sp. 1 | 15 (6) | |
| Forcipomyia sp. 2 | 1 | |
| Forcipomyia sp. 3 | 2 | |
| Undetermined sp. 1 | 1 | |
| Undetermined sp. 2 | 1 | |
| Undetermined sp. 3 | 1 | 3 |
| CHLOROPIDAE | 1 | 5 (2) |
| Oscinimorpha cf. minutissima (Strobl, 1900) | 1 | |
| Undetermined sp. 1 | 3 | |
| Undetermined sp. 2 | 1 (1) | |
| Undetermined sp. 3 | 1 (1) | |
| LAUXANIIDAE | 1 | |
| Sapromyza Fallén, 1810 sp. | 1 | |
| PHORIDAE | 3 (3) | |
| Megaselia Rondani, 1856 sp. | 3(3) | |
| SCATOPSIDAE | 3 (2) | 5 (5) |
| Octaseps Haenni and Amorim, 2016 sp. nov. aff. labellata Cook, 1965 | 3 (2) | 4 (4) |
| Thripomorpha Enderlein, 1905 sp. | 1 (1) | |
| SCIARIDAE | 18 (10) | 94 (37) |
| Bradysia Winnertz, 1867 sp. 1 (Bradysia fallaciosa group) | 3 | 1 |
| Bradysia sp. 2 (Bradysia fallaciosa group) | 1 | |
| Bradysiasp. 3 (Bradysia hilaris group) | 1 (1) | 6 (3) |
| Bradysia sp. 4 | 11 | |
| Bradysiasp. 5 | 3 (2) | |
| Bradysiasp. 6 | 3 (2) | |
| Bradysiasp. 7 | 1 (1) | |
| Bradysia sp. 8 | 1 | |
| Bradysia sp. 9 | 1 | |
| Bradysiasp. 10 | 1 (1) | |
| Bradysiasp. 11 | 1 (1) | |
| Corynoptera Winnertz, 1867 sp. | 10 | |
| Pseudolycoriella Menzel & Mohrig, 1998 sp. | 14 (9) | 54 (27) |
| Chemical Compound | A (7 flowers) | B (6 flowers) | C (5 flowers) | D (20 flowers) | Ceropegia Species with Similar Floral Compounds |
|---|---|---|---|---|---|
| Total sampling time [min]: | 35 | 40 | 35 | 35 | |
| Total scent emitted per flower [ng/h]: | 58.9 | 143.7 | 140.9 | 161.9 | |
| Aliphatics | |||||
| (Z)-3-Hexen-1-yl acetate S,EAD | 0.2 | 0.3 | 20.5 | 0.4 | san, rup, amp, den, car, hay, cyc |
| Aromatics | |||||
| Phenylacetonitrile | 1.4 | 1.4 | 1.1 | 1.8 | den |
| Terpenoids | |||||
| Monoterpenes | |||||
| Limonene S,EAD | 1 | 0.9 | 0.6 | 0.5 | nil |
| Linalool S,EAD | 9.6 | 5.1 | 2.9 | 7.5 | san, rup, amp, hay |
| Furanoid (E)-linalool oxide S,EAD | 2.5 | 3.6 | 0.8 | 2.8 | |
| Furanoid (Z)-linalool oxide S,EAD | 35.9 | 5.6 | 19.5 | 52.4 | |
| Pyranoid (E) + (Z)-linalool oxide S,EAD | 2.8 | 3.6 | 1.6 | 3.5 | |
| β-Myrcene S,EAD | 0.5 | 7.9 | 2.9 | 5.2 | den |
| Myrcenol | 0 | 0 | 0 | tr | |
| (E)-β-Ocimene S,EAD | 34.6 | 38.6 | 39.2 | 13.3 | san, amp, den, car, hay |
| (Z)-β-Ocimene | 0.8 | 0.7 | 0.3 | 0.5 | san |
| Sesquiterpenes | |||||
| α-Farnesene (isomer not assigned) | 0.4 | 1.7 | 0.1 | 0.1 | |
| (E,E)-α-Farnesene | 1.4 | 10.9 | 0.7 | 1 | san |
| (E)-β-Farnesene S,EAD | 0.1 | 0.2 | tr | tr | |
| Irregular terpenes | |||||
| 4-Oxoisophorone S,EAD | 0.5 | 0.3 | 0.6 | 0.5 | den |
| Homoterpenes | |||||
| (E)-4,8-Dimethyl-1,3,7-nonatriene S | 0.6 | 0.3 | 1.5 | 0.1 | san, amp, car, hay |
| Unknowns(7 in total) | |||||
| m/z: 43, 71, 41, 55/59 | 8 | 17.5 | 7.4 | 10.3 | |
| Minor unknowns a | 0.12 | 1.36 | 0.24 | 0.36 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Heiduk, A.; Meve, U.; Menzel, F.; Haenni, J.-P.; Tschirnhaus, M.v.; Dötterl, S.; Johnson, S.D. Fly Pollination of Kettle Trap Flowers of Riocreuxia torulosa (Ceropegieae-Anisotominae): A Generalized System of Floral Deception. Plants 2021, 10, 1564. https://doi.org/10.3390/plants10081564
Heiduk A, Meve U, Menzel F, Haenni J-P, Tschirnhaus Mv, Dötterl S, Johnson SD. Fly Pollination of Kettle Trap Flowers of Riocreuxia torulosa (Ceropegieae-Anisotominae): A Generalized System of Floral Deception. Plants. 2021; 10(8):1564. https://doi.org/10.3390/plants10081564
Chicago/Turabian StyleHeiduk, Annemarie, Ulrich Meve, Frank Menzel, Jean-Paul Haenni, Michael von Tschirnhaus, Stefan Dötterl, and Steven D. Johnson. 2021. "Fly Pollination of Kettle Trap Flowers of Riocreuxia torulosa (Ceropegieae-Anisotominae): A Generalized System of Floral Deception" Plants 10, no. 8: 1564. https://doi.org/10.3390/plants10081564
APA StyleHeiduk, A., Meve, U., Menzel, F., Haenni, J.-P., Tschirnhaus, M. v., Dötterl, S., & Johnson, S. D. (2021). Fly Pollination of Kettle Trap Flowers of Riocreuxia torulosa (Ceropegieae-Anisotominae): A Generalized System of Floral Deception. Plants, 10(8), 1564. https://doi.org/10.3390/plants10081564