
The Effects of Temperature and Water on the Seed Germination and Seedling Development of Rapeseed (Brassica napus L.)


 , ,
, , 
Abstract
1. Introduction
2. Results
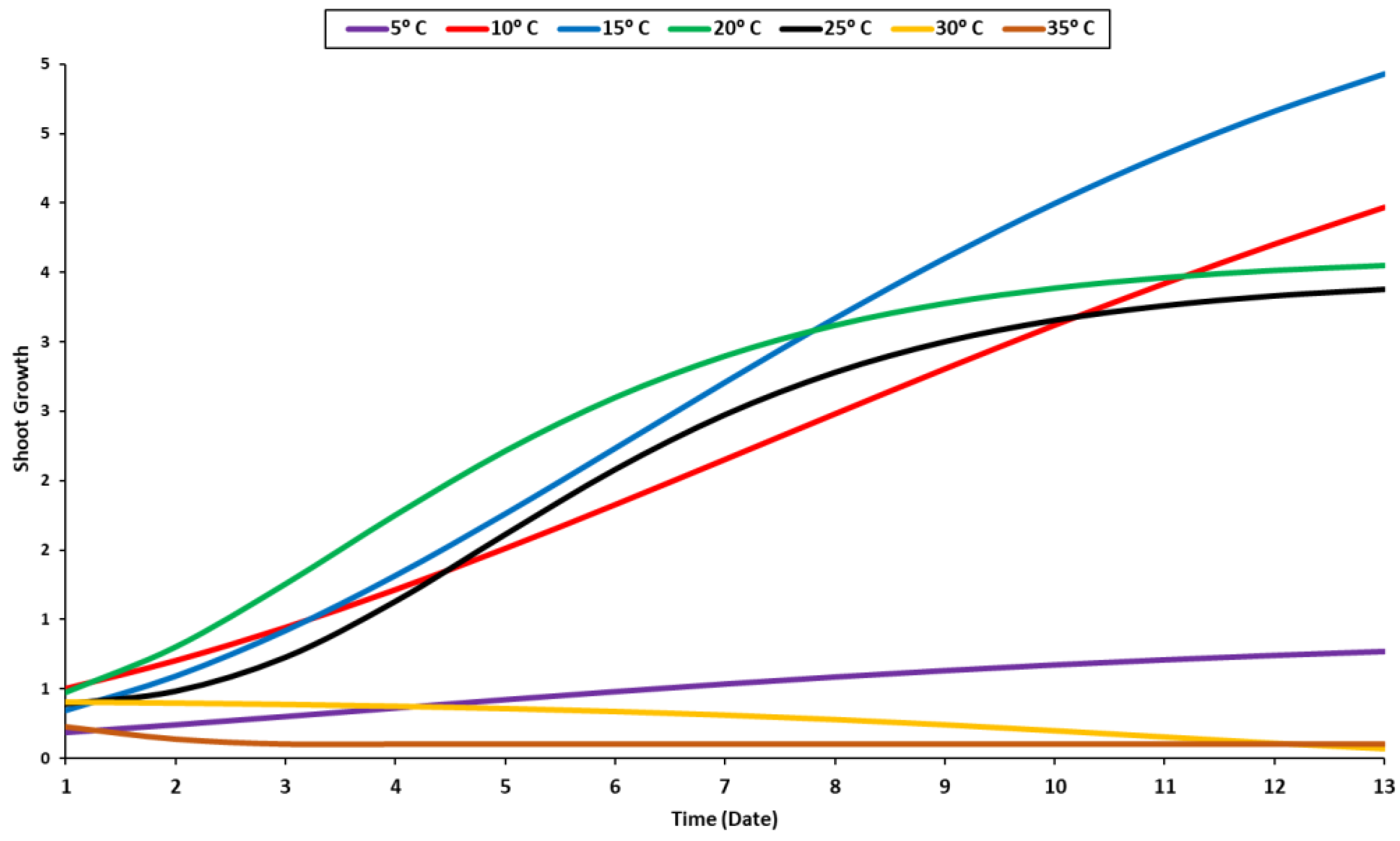
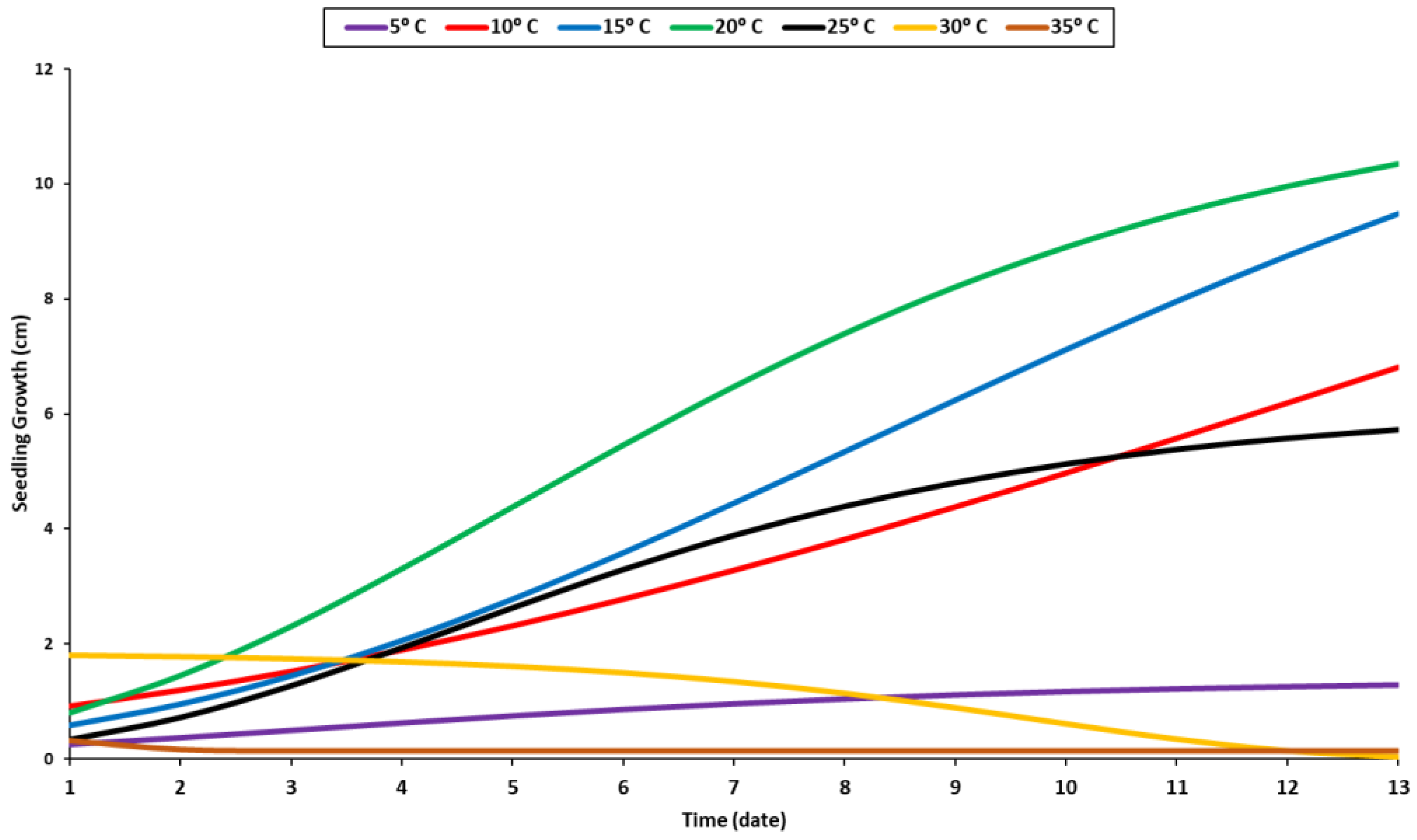
2.1. Temperature Trial
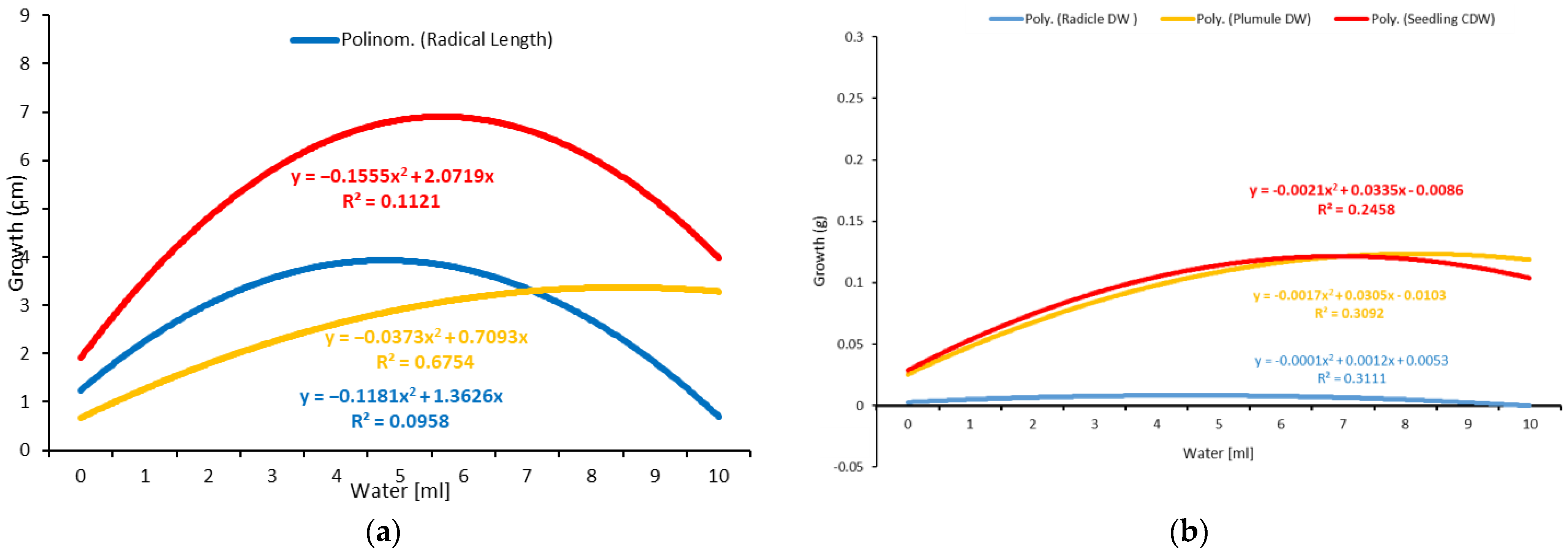
2.2. Water Trial
2.3. Seed Number Trial
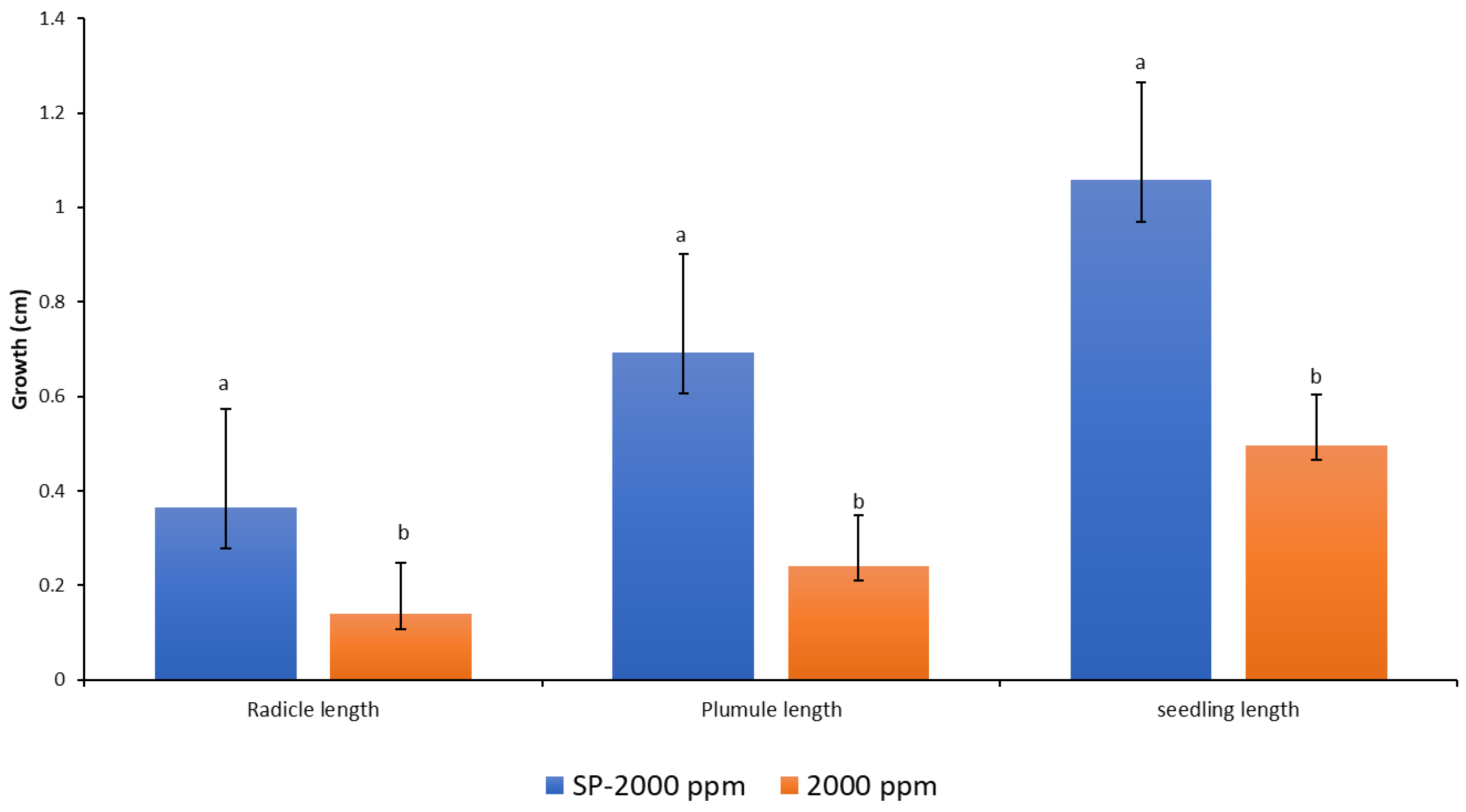
2.4. Antifungal Trial
3. Discussion
4. Materials and Methods
4.1. Temperature Stress Trail
4.2. Water Stress Trail
4.3. Seed Number Trial
4.4. Antifungal Trial
4.5. Statistical Analysis
5. Conclusions
Author Contributions
Funding
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Zhang, H.; Flottmann, S. Seed Yield of Canola (Brassica Napus L.) Is Determined Primarily by Biomass in a High-Yielding Environment. Crop Pasture Sci. 2016, 67, 369–380. [Google Scholar] [CrossRef]
- Food and Agriculture Organization of the United Nations. Available online: https://www.fao.org/faostat/en/#data/QCL (accessed on 20 February 2022).
- Elferjani, R.; Soolanayakanahally, R. Canola Responses to Drought, Heat, and Combined Stress: Shared and Specific Effects on Carbon Assimilation, Seed Yield, and Oil Composition. Front. Plant. Sci. 2018, 9, 1224. [Google Scholar] [CrossRef] [PubMed]
- Friedt, W.; Tu, J.; Fu, T. Academic and Economic Importance of Brassica Napus Rapeseed. In The Brassica napus Genome; Springer: Berlin/Heidelberg, Germany, 2018; pp. 1–20. [Google Scholar] [CrossRef]
- Wittkop, B.; Snowdon, R.J.; Friedt, W. Status and Perspectives of Breeding for Enhanced Yield and Quality of Oilseed Crops for Europe. Euphytica 2009, 170, 131–140. [Google Scholar] [CrossRef]
- McVetty, P.B.E.; Lukow, O.M.; Hall, L.M.; Rajcan, I.; Rahman, H. Grain Production and Consumption: Oilseeds in North America. In Encyclopedia of Food Grains: Second Edition; Elsevier Inc.: Amsterdam, The Netherlands, 2015; Volume 1–4, pp. 401–408. ISBN 9780123947864. [Google Scholar]
- Ali, A.S.; Elozeiri, A.A. Metabolic Processes During Seed Germination. In Advances in Seed Biology; InTech: London, UK, 2017. [Google Scholar]
- Nonogaki, H.; Bassel, G.W.; Bewley, J.D. Germination-Still a Mystery. Plant Sci. 2010, 179, 574–581. [Google Scholar] [CrossRef]
- Szczerba, A.; Płażek, A.; Pastuszak, J.; Kopeć, P.; Hornyák, M.; Dubert, F. Effect of Low Temperature on Germination, Growth, and Seed Yield of Four Soybean (Glycine Max l.) Cultivars. Agronomy 2021, 11, 800. [Google Scholar] [CrossRef]
- Miransari, M.; Smith, D.L. Plant Hormones and Seed Germination. Environ. Exp. Bot. 2014, 99, 110–121. [Google Scholar] [CrossRef]
- Bhardwaj, D.; Wahid Ansari, M.; Kumar Sahoo, R.; Tuteja, N. Biofertilizers Function as Key Player in Sustainable Agriculture by Improving Soil Fertility, Plant Tolerance and Crop Productivity. Microvial Cell Factories 2014, 13, 66. [Google Scholar] [CrossRef]
- Makena, I.M.; Matsiliza-Mlathi, B.; Kleynhans, R. Seed Propagation and Seed Anatomy of Three Eucomis Species. Acta Hortic. 2018, 1204, 263–271. [Google Scholar] [CrossRef]
- Savaedi, Z.; Parmoon, G.; Moosavi, S.A.; Bakhshande, A. The Role of Light and Gibberellic Acid on Cardinal Temperatures and Thermal Time Required for Germination of Charnushka (Nigella Sativa) Seed. Ind. Crops Prod. 2019, 132, 140–149. [Google Scholar] [CrossRef]
- Luna, B.; Chamorro, D. Germination Sensitivity to Water Stress of Eight Cistaceae Species from the Western Mediterranean. Seed Sci. Res. 2016, 26, 101–110. [Google Scholar] [CrossRef]
- Ramírez-Tobías, H.M.; Peña-Valdivia, C.B.; Trejo, C.; Aguirre R, J.R.; Vaquera H, H. Seed Germination of Agave Species as Influenced by Substrate Water Potential. Biol. Res. 2014, 47, 11. [Google Scholar] [CrossRef]
- Rao, A.B.G.; Gupta, R.K. Influence of Soil Moisture Suction and Depth of Sowing on Seedling Emergence of Soybean in a Clayey Soil. Seed Res. 1976, 4, 118–123. [Google Scholar]
- Muscolo, A.; Sidari, M.; Anastasi, U.; Santonoceto, C.; Maggio, A. Effect of PEG-Induced Drought Stress on Seed Germination of Four Lentil Genotypes. J. Plant Interact. 2014, 9, 354–363. [Google Scholar] [CrossRef]
- Abdul Jaleel, C.; Gopi, R.; Sankar, B.; Manivannan, P.; Kishorekumar, A.; Sridharan, R.; Panneerselvam, R. Studies on Germination, Seedling Vigour, Lipid Peroxidation and Proline Metabolism in Catharanthus Roseus Seedlings under Salt Stress. S. Afr. J. Bot. 2007, 73, 190–195. [Google Scholar] [CrossRef]
- Kovács, Z.; Bedő, J.; Pápai, B.; Tóth-Lencsés, A.K.; Csilléry, G.; Szőke, A.; Bányai-Stefanovits, É.; Kiss, E.; Veres, A. Ripening-Induced Changes in the Nutraceutical Compounds of Differently Coloured Pepper (Capsicum Annuum L.) Breeding Lines. Antioxidants 2022, 11, 637. [Google Scholar] [CrossRef]
- Kunos, V.; Cséplő, M.; Seress, D.; Eser, A.; Kende, Z.; Uhrin, A.; Bányai, J.; Bakonyi, J.; Pál, M.; Mészáros, K. The Stimulation of Superoxide Dismutase Enzyme Activity and Its Relation with the Pyrenophora Teres f. Teres Infection in Different Barley Genotypes. Sustainability 2022, 14, 2597. [Google Scholar] [CrossRef]
- Mouradi, M.; Bouizgaren, A.; Farissi, M.; Makoudi, B.; Kabbadj, A.; Very, A.A.; Sentenac, H.; Qaddoury, A.; Ghoulam, C. Osmopriming Improves Seeds Germination, Growth, Antioxidant Responses and Membrane Stability during Early Stage of Moroccan Alfalfa Populations under Water Deficit. Chil. J. Agric. Res. 2016, 76, 265–272. [Google Scholar] [CrossRef][Green Version]
- Chen, K.; Arora, R. Priming Memory Invokes Seed Stress-Tolerance. Environ. Exp. Bot. 2013, 94, 33–45. [Google Scholar] [CrossRef]
- Jisha, K.C.; Vijayakumari, K.; Puthur, J.T. Seed Priming for Abiotic Stress Tolerance: An Overview. Acta Physiol. Plant. 2013, 35, 1381–1396. [Google Scholar] [CrossRef]
- Savvides, A.; Ali, S.; Tester, M.; Fotopoulos, V. Chemical Priming of Plants Against Multiple Abiotic Stresses: Mission Possible? Trends Plant Sci. 2016, 21, 329–340. [Google Scholar] [CrossRef] [PubMed]
- Vicente, M.J.; Martínez-Díaz, E.; Martínez-Sánchez, J.J.; Franco, J.A.; Bañón, S.; Conesa, E. Effect of Light, Temperature, and Salinity and Drought Stresses on Seed Germination of Hypericum Ericoides, a Wild Plant with Ornamental Potential. Sci. Hortic. 2020, 270, 109433. [Google Scholar] [CrossRef]
- Koger, C.H.; Reddy, K.N.; Poston, D.H. Factors Affecting Seed Germination, Seedling Emergence, and Survival of Texasweed (Caperonia Palustris). Weed Sci. 2004, 52, 989–995. [Google Scholar] [CrossRef]
- Bradford, K.J. Applications of Hydrothermal Time to Quantifying and Modeling Seed Germination and Dormancy. Weed Sci. 2002, 50, 248–260. [Google Scholar] [CrossRef]
- Steinmaus, S.J.; Prather, T.S.; Holt, J.S. Estimation of Base Temperatures for Nine Weed Species. J. Exp. Bot. 2000, 51, 275–286. [Google Scholar] [CrossRef] [PubMed]
- Wang, R.; Cheng, T.; Hu, L. Effect of Wide-Narrow Row Arrangement and Plant Density on Yield and Radiation Use Efficiency of Mechanized Direct-Seeded Canola in Central China. Field Crops Res. 2015, 172, 42–52. [Google Scholar] [CrossRef]
- Zhao, T.J.; Liu, Y.; Yan, Y.B.; Feng, F.; Liu, W.Q.; Zhou, H.M. Identification of the Amino Acids Crucial for the Activities of Drought Responsive Element Binding Factors (DREBs) of Brassica Napus. FEBS Lett. 2007, 581, 3044–3050. [Google Scholar] [CrossRef] [PubMed]
- Okcu, G.; Kaya, M.D.; Atak, M. Effects of Salt and Drought Stresses on Germination and Seedling Growth Pea (Pisum Sativum L.). Turk. J. Agric. For. 2005, 29, 237–242. [Google Scholar]
- Marjanović-Jeromela, A.; Nagl, N.; Gvozdanović-Varga, J.; Hristov, N.; Kondić-Špika, A.; Vasić, M.; Marinković, R. Genotype by Environment Interaction for Seed Yield per Plant in Rapeseed Using AMMI Model. Pesq. Agropec. Bras. Brasilia 2011, 46, 174–181. [Google Scholar] [CrossRef]
- de Figueiredo E Albuquerque, M.C.; de Carvalho, N.M. Effect of the Type of Environmental Stress on the Emergence of Sunflower (Heliantus Annus L.), Soybean (Glycine Max (L.) Merril) and Maize (Zea Mays L.) Seeds with Different Levels of Vigor. Seed Sci. Technol. 2003, 31, 465–479. [Google Scholar] [CrossRef]
- Diepenbrock, W. Yield Analysis of Winter Oilseed Rape (Brassica Napus L.): A Review. Field Crops Res. 2000, 67, 35–49. [Google Scholar] [CrossRef]
- Aksouh-Harradj, N.M.; Campbell, L.C.; Mailer, R.J. Canola Response to High and Moderately High Temperature Stresses during Seed Maturation. Can. J. Plant Sci. 2006, 86, 967–980. [Google Scholar] [CrossRef]
- Drebee, H.A.; Razak, N.A.A. Measuring the Efficiency of Colleges at the University of Al-Qadisiyah-Iraq: A Data Envelopment Analysis Approach. J. Ekon. Malays. 2018, 52, 163–179. [Google Scholar] [CrossRef]
- Balodis, O.; Gaile, Z. Sowing Date and Rate Effect on Winter Oilseed Rape (Brassica Napus L.) Yield Components’ Formation. Proc. Latv. Acad. Sci. Sect. B Nat. Exact Appl. Sci. 2016, 70, 384–392. [Google Scholar] [CrossRef]
- Lamichhane, J.R.; You, M.P.; Laudinot, V.; Barbetti, M.J.; Aubertot, J.-N. Revisiting Sustainability of Fungicide Seed Treatments for Field Crops. Plant Dis. 2020, 104, 610–623. [Google Scholar] [CrossRef]
- Khaeim, H.; Kende, Z.; Balla, I.; Gyuricza, C.; Eser, A.; Tarnawa, Á. The Effect of Temperature and Water Stresses on Seed Germination and Seedling Growth of Wheat (Triticum Aestivum L.). Sustainability 2022, 14, 3887. [Google Scholar] [CrossRef]
- Khaeim, H.; Kende, Z.; Jolánkai, M.; Kovács, G.P.; Gyuricza, C.; Tarnawa, Á. Impact of Temperature and Water on Seed Germination and Seedling Growth of Maize (Zea Mays L.). Agronomy 2022, 12, 397. [Google Scholar] [CrossRef]
- Wulff, R. Intrapopulational Variation in the Germination of Seeds in Hyptis Suaveolens. Ecol. Soc. Am. 1973, 54, 646–649. [Google Scholar] [CrossRef]
- Padilla, F.M.; Miranda, J.D.D.; Pugnaire, F.I. Early Root Growth Plasticity in Seedlings of Three Mediterranean Woody Species. Plant Soil 2007, 296, 103–113. [Google Scholar] [CrossRef]
- Larios, E.; Venable, D.L. Selection for Seed Size: The Unexpected Effects of Water Availability and Density. Funct. Ecol. 2018, 32, 2216–2224. [Google Scholar] [CrossRef]
- Larios, E.; Búrquez, A.; Becerra, J.X.; Venable, D.L. Natural Selection on Seed Size through the Life Cycle of a Desert Annual Plant. Ecology 2014, 95, 3213–3220. [Google Scholar] [CrossRef]
- Alvarado, V.; Bradford, K.J. A Hydrothermal Time Model Explains the Cardinal Temperatures for Seed Germination. Plant Cell Environ. 2002, 25, 1061–1069. [Google Scholar] [CrossRef]
- Puppala, N.; Bhardwaj, H. Evaluation of Salinity Tolerance of Canola Germination; Janick, J., Ed.; ASHS Press: Alexandria, VA, USA, 1999. [Google Scholar]
- Luo, T.; Xian, M.; Khan, M.N.; Hu, L.; Xu, Z. Estimation of Base Temperature for Germination of Rapeseed (Brassica Napus) Using Different Models. Int. J. Agric. Biol. 2018, 20, 524–530. [Google Scholar] [CrossRef]
- Liu, F.; Jensen, C.R.; Shahanzari, A.; Andersen, M.N.; Jacobsen, S.E. ABA Regulated Stomatal Control and Photosynthetic Water Use Efficiency of Potato (Solanum Tuberosum L.) during Progressive Soil Drying. Plant Sci. 2005, 168, 831–836. [Google Scholar] [CrossRef]
- Cassaro-Silva, M. Efeito Da Temperatura Na Germinação de Sementes de Manduirana (Senna Macranthera (Collad.) Irwin et Barn. —Caesalpiniaceae). Rev. Bras. Sementes 2001, 23, 92–99. [Google Scholar] [CrossRef][Green Version]
- Windauer, L.B.; Martinez, J.; Rapoport, D.; Wassner, D.; Benech-Arnold, R. Germination Responses to Temperature and Water Potential in Jatropha Curcas Seeds: A Hydrotime Model Explains the Difference between Dormancy Expression and Dormancy Induction at Different Incubation Temperatures. Ann. Bot. 2012, 109, 265–273. [Google Scholar] [CrossRef]
- de Oliveira, A.K.M.; Ribeiro, J.W.F.; Pereira, K.C.L.; Silva, C.A.A. Efeito Da Temperatura Sobre a Germinação de Sementes de Diptychandra Aurantiaca (Fabaceae). Acta Sci. Agron. 2013, 35, 203–208. [Google Scholar] [CrossRef]
- Vassilevska-Ivanova, R.; Tcekova, Z. Effect of Temperature on Seed Germination and Seedling Growth of Sunflower (Helianthus Annuus L.). Compte Renuds l’Académie Bulg. Sci. 2002, 55, 10–67. [Google Scholar]
- Shaban, M. Effect of Water and Temperature on Seed Germination and Emergence as a Seed Hydrothermal Time Model. Int. J. Adv. Biol. Biomed. Res. 2013, 1, 1686–1691. [Google Scholar]
- Li, P.H.; Davis, D.W.; Shen, Z.-Y. High-Temperature-Acclimation Potential of the Common Bean: Can It Be Used as a Selection Criterion for Improving Crop Performance in High-Temperature Environments? Field Crops Res. 1991, 27, 241–256. [Google Scholar] [CrossRef]
- Aksouh, N.M.; Jacobs, B.C.; Stoddard, F.L.; Mailer, R.J. Response of Canola to Different Heat Stresses. Aust. J. Agric. Res. 2001, 52, 817–824. [Google Scholar] [CrossRef]
- Hawker, J.; Jenner, C. High Temperature Affects the Activity of Enzymes in the Committed Pathway of Starch Synthesis in Developing Wheat Endosperm. Funct. Plant Biol. 1993, 20, 197. [Google Scholar] [CrossRef]
- Campos, H.; Trejo, C.; Peña-Valdivia, C.B.; García-Nava, R.; Víctor Conde-Martínez, F.; Cruz-Ortega, R. Water Availability Effects on Germination, Membrane Stability and Initial Root Growth of Agave Lechuguilla and A. Salmiana. Flora Morphol. Distrib. Funct. Ecol. Plants 2020, 268, 151606. [Google Scholar] [CrossRef]
- Bradford, K.J. A Water Relations Analysis of Seed Germination Rates. Plant Physiol. 1990, 94, 840–849. [Google Scholar] [CrossRef] [PubMed]
- Lopes, M.J.d.S.; Dias-Filho, M.B.; Gurgel, E.S.C. Successful Plant Growth-Promoting Microbes: Inoculation Methods and Abiotic Factors. Front. Sustain. Food Syst. 2021, 5, 606454. [Google Scholar] [CrossRef]
- Pan, J.; Sharif, R.; Xu, X.; Chen, X. Mechanisms of Waterlogging Tolerance in Plants: Research Progress and Prospects. Front. Plant. Sci. 2021, 11, 627331. [Google Scholar] [CrossRef]
- Schütz, W.; Lamont, B.B.; Milberg, P. Germination Requirements and Seedling Responses to Water Availability and Soil Type in Four Eucalypt Species. Acta Oecol. 2002, 23, 23–30. [Google Scholar] [CrossRef]
- Brouwer, R. Some Aspect of the Equilibrium between Overground and Underground Plants Parts. Jaarboek Instituut voor Biologisch en Scheikundig Onderzoek (IBS, Wageningen) 1963, 1963, 31–39. [Google Scholar]
- White, T.C.R. Opposing Paradigms: Regulation or Limitation of Populations? Oikos 2001, 93, 148–152. [Google Scholar] [CrossRef]
- Lortie, C.J.; Turkington, R. The Effect of Initial Seed Density on the Structure of a Desert Annual Plant Community. J. Ecol. 2002, 90, 435–445. [Google Scholar] [CrossRef]
- Vasanthakumari, M.M.; Shridhar, J.; Madhura, R.J.; Nandhitha, M.; Kasthuri, C.; Janardhana, B.; Nataraja, K.N.; Ravikanth, G.; Uma Shaanker, R. Role of Endophytes in Early Seedling Growth of Plants: A Test Using Systemic Fungicide Seed Treatment. Plant Physiol. Rep. 2019, 24, 86–95. [Google Scholar] [CrossRef]
- You, M.P.; Lamichhane, J.R.; Aubertot, J.-N.; Barbetti, M.J. Understanding Why Effective Fungicides Against Individual Soilborne Pathogens Are Ineffective with Soilborne Pathogen Complexes. Plant Dis. 2020, 104, 904–920. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Kingsley, K.L.; Bergen, M.S.; Kowalski, K.P.; White, J.F. Fungal Disease Prevention in Seedlings of Rice (Oryza Sativa) and Other Grasses by Growth-Promoting Seed-Associated Endophytic Bacteria from Invasive Phragmites Australis. Microorganisms 2018, 6, 21. [Google Scholar] [CrossRef] [PubMed]
- Verma, S.K.; Kingsley, K.; Irizarry, I.; Bergen, M.; Kharwar, R.N.; White, J.F. Seed-Vectored Endophytic Bacteria Modulate Development of Rice Seedlings. J. Appl. Microbiol. 2017, 122, 1680–1691. [Google Scholar] [CrossRef] [PubMed]
- Bertelsen, J.R.; de Neergaard, E.; Smedegaard-Petersen, V. Fungicidal Effects of Azoxystrobin and Epoxiconazole on Phyllosphere Fungi, Senescence and Yield of Winter Wheat. Plant Pathol. 2001, 50, 190–205. [Google Scholar] [CrossRef]











{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Water (mL) | Germinated (n) | Radicle (cm) | Plumule (cm) | Seedling (cm) | Plumule DW (g) | Radicle DW (g) | Seedling DW (g) | Corrected DW (g) |
|---|---|---|---|---|---|---|---|---|
| 0 | 0.000 ± 0.00 c | 0.000 ± 0.0 d | 0.000 ± 0.00 f | 0.000 ± 0.00 f | 0.000 ± 0.000 f | 0.000 ± 0.00 d | 0.000 ± 0.000 e | 0.000 ± 0.000 d |
| 1 | 19.800 ± 0.45 a | 5.420 ± 1.41 a | 1.955 ± 0.29 e | 7.375 ± 1.27 ab | 0.080 ± 0.005 de | 0.012 ± 0.01 a | 0.093 ± 0.004 bc | 0.092 ± 0.005 b |
| 2 | 19.400 ± 0.89 a | 5.430 ± 0.42 a | 2.354 ± 0.14 de | 7.784 ± 0.50 a | 0.079 ± 0.007 e | 0.011 ± 0.01 a | 0.089 ± 0.009 bc | 0.087 ± 0.012 b |
| 3 | 19.600 ± 0.55 a | 5.888 ± 1.44 a | 2.885 ± 0.19 bc | 8.773 ± 1.269 a | 0.084 ± 0.003 cde | 0.011 ± 0.00 a | 0.095 ± 0.005 b | 0.093 ± 0.006 ab |
| 4 | 20.000 ± 0.00 a | 3.952 ± 1.25 b | 3.593 ± 0.38 a | 7.545 ± 1.56 ab | 0.088 ± 0.011 bcd | 0.008 ± 0.00 b | 0.096 ± 0.10 b | 0.096 ± 0.010 ab |
| 5 | 19.600 ± 0.55 a | 2.780 ± 1.2 bc | 2.369 ± 0.49 cd | 5.149 ± 1.59 de | 0.098 ± 0.002 a | 0.008 ± 0.00 b | 0.105 ± 0.004 a | 0.104 ± 0.003 a |
| 6 | 16.200 ± 2.28 b | 1.648 ± 0.60 c | 2.367 ± 0.58 cd | 4.015 ± 1.17 e | 0.076 ± 0.012 e | 0.003 ± 0.00 c | 0.079 ± 0.013 d | 0.065 ± 0.020 c |
| 7 | 19.600 ± 0.55 a | 1.796 ± 0.45 c | 2.925 ± 0.33 bc | 4.721 ± 0.73 de | 0.092 ± 0.005 abc | 0.003 ± 0.00 c | 0.095 ± 0.005 b | 0.094 ± 0.006 ab |
| 8 | 19.800 ± 0.45 a | 1.718 ± 0.88 c | 2.971 ± 0.81 b | 4.689 ± 1.65 de | 0.094 ± 0.005 ab | 0.003 ± 0.00 c | 0.096 ± 0.005 b | 0.096 ± 0.005 ab |
| 9 | 19.000 ± 0.71 a | 2.306 ± 0.67 c | 3.692 ± 0.59 a | 5.998 ± 1.18 cd | 0.091 ± 0.002 abc | 0.004 ± 0.00 c | 0.094 ± 0.003 b | 0.090 ± 0.006 b |
| 10 | 17.400 ± 1.67 b | 2.482 ± 0.91 c | 3.661 ± 0.43 a | 6.143 ± 1.29 bcd | 0.081 ± 0.008 de | 0.003 ± 0.00 c | 0.085 ± 0.007 cd | 0.074 ± 0.012 c |
| LSD | 1.251 | 1.202 | 0.564 | 1.545 | 0.008 | 0.0019 | 0.0086 | 0.0119 |
| Water (mL) | Germinated (n) | Radicle (cm) | Plumule (cm) | Seedling (cm) | Plumule 1DW (g) | Radicle 1DW (g) | Seedling 1DW (g) | Corrected 1DW (g) |
|---|---|---|---|---|---|---|---|---|
| 0 | 0.0 ± 0.00 e | 0.000 ± 0.00 j | 0.000 ± 0.00 i | 0.000 ± 0.00 g | 0.000 ± 0.00 i | 0.000 ± 0.00 h | 0.000 ± 0.00 f | 0.000 ± 0.00 g |
| 0.65 | 19.8 ± 0.45 ab | 4.251 ± 1.08 de | 1.453 ± 0.22 h | 5.704 ± 1.26 de | 0.069 ± 0.03 h | 0.010 ± 0.00 cd | 0.079 ± 0.03 e | 0.078 ± 0.03 f |
| 1.05 | 19.2 ± 0.45 abc | 5.960 ± 1.35 bc | 2.182 ± 0.44 fg | 8.142 ± 1.39 c | 0.080 ± 0.00 efgh | 0.013 ± 0.00 ab | 0.092 ± 0.00 bcd | 0.089 ± 0.00 cdef |
| 1.45 | 20.0 ± 0.00 a | 7.202 ± 1.40 ab | 2.713 ± 0.13 cdef | 9.915 ± 1.46 ab | 0.074 ± 0.01 gh | 0.012 ± 0.00 bc | 0.086 ± 0.01 de | 0.086 ± 0.01 cdef |
| 1.85 | 19.8 ± 0.45 ab | 7.754 ± 0.88 a | 2.586 ± 0.17 cdefg | 10.34 ± 0.94 a | 0.078 ± 0.01 fgh | 0.014 ± 0.00 a | 0.092 ± 0.01 bcd | 0.091 ± 0.01 cdef |
| 2.25 | 20.0 ± 0.00 a | 5.310 ± 1.60 cd | 2.363 ± 0.17 efg | 7.673 ± 1.66 c | 0.082 ± 0.01 defgh | 0.010 ± 0.00 c | 0.092 ± 0.01 bcd | 0.092 ± 0.01 cdef |
| 2.65 | 20.0 ± 0.00 a | 5.939 ± 0.84 bc | 2.429 ± 0.27 defg | 8.368 ± 1.06 bc | 0.086 ± 0.00 cdefg | 0.012 ± 0.00 bc | 0.097 ± 0.00 bcd | 0.097 ± 0.00 abc |
| 3.05 | 20.0 ± 0.00 a | 5.900 ± 2.30 c | 2.605 ± 0.25 cdefg | 8.505 ± 2.49 bc | 0.081 ± 0.01 defgh | 0.010 ± 0.00 cd | 0.091 ± 0.01 bcd | 0.091 ± 0.01 cdef |
| 3.45 | 20.0 ± 0.00 a | 3.937 ± 1.17 ef | 3.299 ± 0.39 ab | 7.236 ± 1.49 cd | 0.084 ± 0.01 cdefg | 0.007 ± 0.00 f | 0.091 ± 0.01 bcd | 0.091 ± 0.01 cdef |
| 3.85 | 19.8 ± 0.45 ab | 2.601 ± 0.65 ghi | 2.567 ± 0.37 cdefg | 5.168 ± 1.00 ef | 0.103 ± 0.01 a | 0.007 ± 0.00 f | 0.109 ± 0.01 a | 0.108 ± 0.01 a |
| 4.25 | 18.6 ± 1.14 bcd | 2.310 ± 0.85 ghi | 3.106 ± 0.55 abc | 5.416 ± 1.37 ef | 0.083 ± 0.01 cdefg | 0.004 ± 0.00 g | 0.087 ± 0.01 cde | 0.081 ± 0.01 def |
| 4.7 | 19.6 ± 0.55 abc | 2.909 ± 0.50 fgh | 2.882 ± 0.26 bcde | 5.791 ± 0.62 def | 0.096 ± 0.01 abc | 0.008 ± 0.00 ef | 0.104 ± 0.00 ab | 0.102 ± 0.01 ab |
| 5.1 | 19.2 ± 0.84 abc | 2.219 ± 0.92 ghi | 3.336 ± 0.56 ab | 5.555 ± 1.43 ef | 0.087 ± 0.01 bcdef | 0.004 ± 0.00 g | 0.091 ± 0.01 bcd | 0.088 ± 0.02 cdef |
| 5.5 | 18.4 ± 3.85 cd | 2.053 ± 1.17 hi | 2.122 ± 1.13 g | 4.175 ± 2.28 f | 0.100 ± 0.00 a | 0.008 ± 0.00 ef | 0.109 ± 0.00 a | 0.100 ± 0.02 abc |
| 5.9 | 19.8 ± 0.45 ab | 3.479 ± 0.31 efg | 2.444 ± 0.41 defg | 5.923 ± 0.63 de | 0.099 ± 0.00 ab | 0.010 ± 0.00 cd | 0.109 ± 0.00 a | 0.108 ± 0.00 a |
| 6.3 | 19.0 ± 1.23 abc | 2.836 ± 0.75 fgh | 2.976 ± 0.58 bcd | 5.812 ± 1.30 def | 0.093 ± 0.01 abcd | 0.007 ± 0.00 f | 0.099 ± 0.01 abc | 0.095 ± 0.02 bcde |
| 6.7 | 18.6 ± 1.14 bcd | 1.710 ± 0.58 hi | 2.966 ± 0.59 bcd | 4.676 ± 1.00 ef | 0.090 ± 0.00 abcde | 0.004 ± 0.00 g | 0.094 ± 0.00 bcd | 0.087 ± 0.01 cdef |
| 7 | 19.8 ± 0.45 ab | 1.796 ± 0.45 hi | 2.925 ± 0.33 bcde | 4.721 ± 0.73 ef | 0.092 ± 0.00 abcd | 0.003 ± 0.00 g | 0.095 ± 0.00 bcd | 0.095 ± 0.01 abcd |
| 7.5 | 19.8 ± 0.45 ab | 2.149 ± 0.27 hi | 3.617 ± 0.28 a | 5.767 ± 0.25 def | 0.094 ± 0.01 abcd | 0.004 ± 0.00 g | 0.098 ± 0.01 bcd | 0.097 ± 0.01 abc |
| 7.9 | 19.4 ± 0.55 abc | 1.466 ± 0.76 i | 2.890 ± 0.73 bcde | 4.356 ± 1.48 ef | 0.096 ± 0.01 abc | 0.003 ± 0.00 g | 0.099 ± 0.01 bcd | 0.096 ± 0.01 abcd |
| 8.3 | 17.8 ± 1.92 d | 1.980 ± 0.54 hi | 3.430 ± 0.31 ab | 5.410 ± 0.83 ef | 0.086 ± 0.01 cdefg | 0.004 ± 0.00 g | 0.090 ± 0.01 cde | 0.080 ± 0.02 ef |
| LSD | 1.325 | 1.271 | 0.575 | 1.645 | 0.013 | 0.002 | 0.0129 | 0.0154 |
| 1SN | 2No g % | 3S % | 4R % | 5SP-Germ % | 6N-Germ % | 7Agg-Value |
|---|---|---|---|---|---|---|
| 15 | 0.140 ± 0.12 | 0.153 ± 0.02 | 0.007 ± 0.006 | 0.013 ± 0.009 | 0.667 ± 0.03 b | 0.692 ± 0.034 b |
| 20 | 0.085 ± 0.07 | 0.165 ± 0.02 | 0.005 ± 0.005 | 0.025 ± 0.013 | 0.735 ± 0.03 ab | 0.769 ± 0.024 ab |
| 25 | 0.072 ± 0.07 | 0.104 ± 0.006 | 0.020 ± 0.009 | 0.008 ± 0.005 | 0.792 ± 0.03 a | 0.813 ± 0.029 a |
| LSD | NS | NS | NS | NS | 0.091 | 0.082 |
| Amount of Water Based on 1 mL of Intervals | Amount of Water Based on the TKW | ||||
|---|---|---|---|---|---|
| 1TN | 2WA (mL) | 3TN | 4PW (%) | 5WA (mL) | 6RAW (mL) |
| 1 | 0 | 10 | 500 | 0.668 | 0.65 |
| 2 | 1 | 11 | 800 | 1.0688 | 1.05 |
| 3 | 2 | 12 | 1100 | 1.4696 | 1.45 |
| 4 | 3 | 13 | 1400 | 1.8704 | 1.85 |
| 5 | 4 | 14 | 1700 | 2.2712 | 2.25 |
| 6 | 5 | 15 | 2000 | 2.672 | 2.65 |
| 7 | 6 | 16 | 2300 | 3.0728 | 3.05 |
| 8 | 7 | 17 | 2600 | 3.4736 | 3.45 |
| 9 | 8 | 18 | 2900 | 3.8744 | 3.85 |
| 10 | 9 | 19 | 3200 | 4.2752 | 4.25 |
| 10 | 20 | 3500 | 4.676 | 4.7 | |
| 21 | 3800 | 5.0768 | 5.1 | ||
| 22 | 4100 | 5.4776 | 5.5 | ||
| 23 | 4400 | 5.8784 | 5.9 | ||
| 24 | 4700 | 6.2792 | 6.3 | ||
| 25 | 5000 | 6.68 | 6.7 | ||
| 26 | 5300 | 7.0808 | 7 | ||
| 27 | 5600 | 7.4816 | 7.5 | ||
| 28 | 5900 | 7.8824 | 7.9 | ||
| 29 | 6200 | 8.2832 | 8.3 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Haj Sghaier, A.; Tarnawa, Á.; Khaeim, H.; Kovács, G.P.; Gyuricza, C.; Kende, Z. The Effects of Temperature and Water on the Seed Germination and Seedling Development of Rapeseed (Brassica napus L.). Plants 2022, 11, 2819. https://doi.org/10.3390/plants11212819
Haj Sghaier A, Tarnawa Á, Khaeim H, Kovács GP, Gyuricza C, Kende Z. The Effects of Temperature and Water on the Seed Germination and Seedling Development of Rapeseed (Brassica napus L.). Plants. 2022; 11(21):2819. https://doi.org/10.3390/plants11212819
Chicago/Turabian StyleHaj Sghaier, Asma, Ákos Tarnawa, Hussein Khaeim, Gergő Péter Kovács, Csaba Gyuricza, and Zoltán Kende. 2022. "The Effects of Temperature and Water on the Seed Germination and Seedling Development of Rapeseed (Brassica napus L.)" Plants 11, no. 21: 2819. https://doi.org/10.3390/plants11212819
APA StyleHaj Sghaier, A., Tarnawa, Á., Khaeim, H., Kovács, G. P., Gyuricza, C., & Kende, Z. (2022). The Effects of Temperature and Water on the Seed Germination and Seedling Development of Rapeseed (Brassica napus L.). Plants, 11(21), 2819. https://doi.org/10.3390/plants11212819